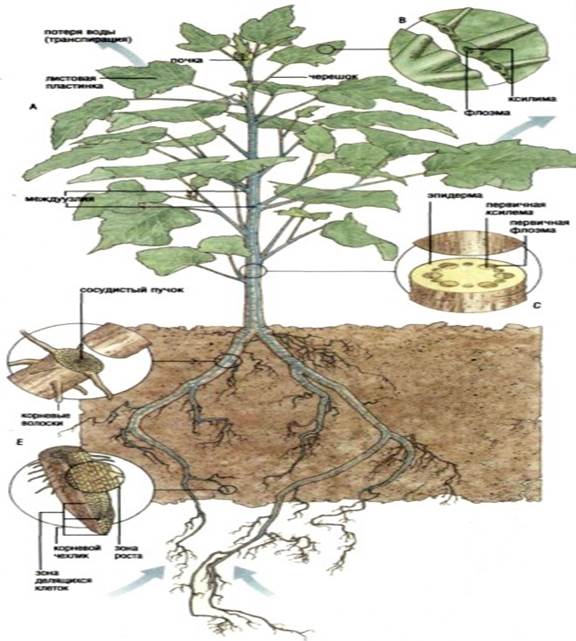
Water moves in the plant along the water potential gradient. Water absorbed by root hairs and other cells of the epidermis moves from the cells of the outer part of the root to the xylem, which occupies the central part of the root (Fig. 6.8). The main pathway for water diffusion in the outer part of the root is the apoplast, a continuous collection of cell walls. However, in the endoderm (a cylindrical layer of cells surrounding the conductive tissue), free diffusion along the cell walls encounters an obstacle - the waterproof cork layer of the Casparian girdle. Water must change its path here and pass through the membrane and protoplast of endoderm cells, which thus plays the role of an osmotic barrier between the root cortex and its central cylinder. In monocots, the inner tangential walls of the cells also become corky, but these walls are permeated with pores, through which, like channels, water can pass.
Rice. 6.7. Mustard sprout root. Numerous thin outgrowths - root hairs
Water rises through the xylem to the aerial parts of the plant. Xylem is made up of several types of cells. Water moves in it mainly through vessels and tracheids (Fig. 2.6 and 6.9). Both those and other cells are perfectly adapted for this purpose: they are elongated, devoid of living content and hollow inside, that is, they are like tubes for water. The lignified secondary cell walls are strong enough to withstand the enormous pressure difference that occurs when water rises to the tops of tall trees. The end, and sometimes the side walls of the segments of the vessels are perforated; vessels, consisting of segments connected end to end, form long tubes through which water with minerals dissolved in it easily passes. There are no perforations in the tracheids, and water, in order to get from one tracheid to another, must pass through their end walls; however, tracheids are very long cells, and therefore this design is well adapted for conducting water. Flowering plants have both vessels and tracheids; more primitive forms of vessels, as a rule, do not.
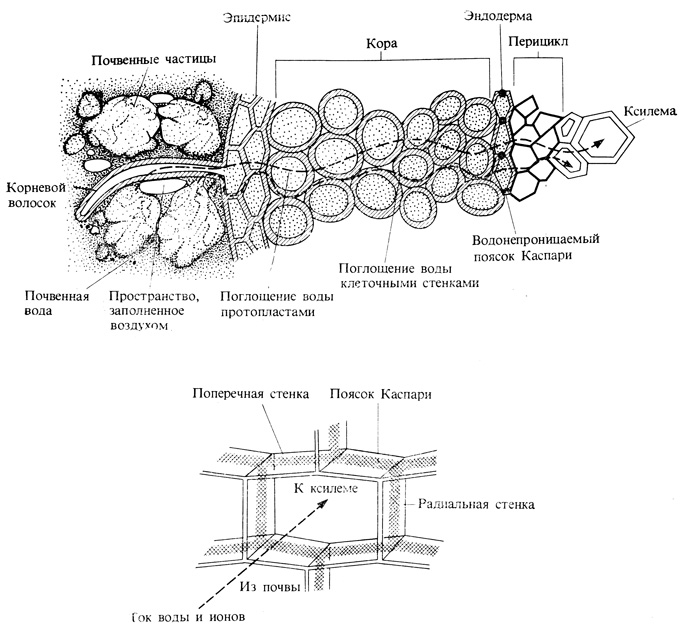
Rice. 6.8. The flow of water from the soil to the roots. Water can move through both the apoplast and the symplast until it reaches the endodermis. Further movement along the apoplast is impossible; here this path is blocked by a barrier - the Caspari belt. The lower part of the figure shows at high magnification the Casparian belt - a waterproof barrier that forces water to leave the apoplast and rush through the membranes of the endoderm cells into the symplast.
Leaf veins, consisting of strands of xylem and phloem, form such a dense network in the leaf that any of its cells is close enough to the water source (Fig. 3.9). From the xylem, water diffuses into the cell walls of the mesophyll. Thus, water in the liquid phase fills all the way from the soil - through the root and stem - to the mesophyll cells in the leaf. The total water flow is always directed towards the lower water potential, i.e., ψ is maximum in the soil, somewhat lower in the root cells, and lowest in the cells adjacent to the leaf epidermis. The small value of ψ in these last cells is explained mainly by the evaporation of water from the surface of the leaf, i.e., by transpiration (to which the next section is devoted). Tab. 6.2 gives an idea of the gradients that determine the movement of water in a plant from its entry from the soil to evaporation into the atmosphere.
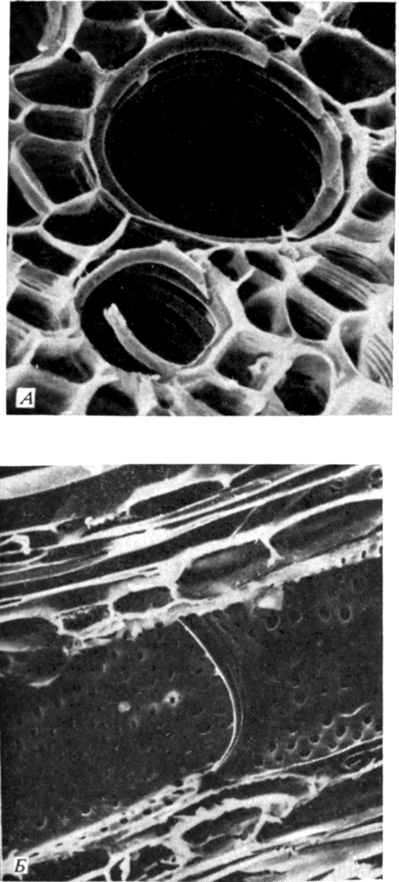
Rice. 6.9. A. Cross section of a castor bean (Ricinus communis) leaf petiole. Conductive elements of xylem with spiral thickenings are visible. Due to this structure, the tube can stretch as the petiole grows. (The larger tube is approximately 15 µm in diameter.) B. Longitudinal section of a Ricinus stem showing the xylem vessel and the perforated end walls of the two vessel segments at their junction. Elliptical and rounded bordered pores with a trailing film are visible, but without a torus, such pores are typical of dicots. (Vessel diameter is approximately 21 µm.) (Courtesy of Milburn J. A., Glasgow University; photographs taken with a scanning electron microscope.)

Table 6.2 Approximate values of water potential (ψ) and water potential difference (Δψ) for a hypothetical soil-plant-air system. [Suggested conditions - relatively small tree, well-drained soil, relative humidity of about 50% at 22 °C (ψ=-1000 bar)1)]
Water passes from the plant into the surrounding air mainly in the vapor state. In the mesophyll of the leaf there are extensive intercellular spaces, and each cell of the mesophyll borders on such an intercellular space at least on one of its sides. Due to the evaporation of water from moist cell walls, the air in the intercellular spaces is saturated with water vapor, and some of these vapors are lost - they go outside. Since the cells of the epidermis in most plants are covered with a waxy waterproof cuticle, water vapor exits the leaf into the atmosphere mainly through stomata (Fig. 6.10).
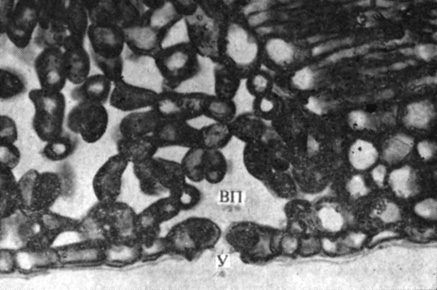
Rice. 6.10. Cross section of a leaf showing an open stoma (U) with a substomatal air cavity (VP). Pay attention to the fact that large, air-filled intercellular spaces penetrate the entire thickness of the leaf.
Movement of water in plants
The main source of moisture is the water in the soil, and the main organ of water absorption into the plant is the root system. The role of this organ, first of all, is that due to the huge surface, water is supplied to the plant from a larger volume of soil.
The formed root system is a complex organ with a well-differentiated external and internal structure. The root system has an absorbing or suction zone - this is the zone of root hairs. Having entered the cell of the root hair, water becomes part of the living system - the plant cell - and obeys the laws that operate in the living cell.
Movement through the plant is determined by two main drivers of water flow in the plant: the lower water flow driver or root pressure, the upper water flow driver or the suction action of the atmosphere.

The main force causing the entry and movement of water in the plant is the process of transpiration, which results in a water potential gradient. The water potential gradient between the cell and the surrounding space creates the driving force for the flow of water across the membrane. If the environment surrounding the cell is hypertonic, more concentrated than cell sap, solutions, then water will come out of the cell to the outside. This will lead to the loss of turgor by the cell, the separation of the plasma membrane from the cell wall and the isolation of the protoplast - the phenomenon of plasmolysis.
The mechanism that ensures the raising of water along the plant due to root pressure is called the lower end motor of the water current.
Root pressure is created when water passes from the root bark into the vascular system of the root when water passes through the passage cells of the pericycle, from which water, under pressure, is, as it were, injected into the xylem vessels. This is evidenced by the phenomena of guttation and "weeping of plants".
Water absorbed by root hairs and other cells of the epidermis moves from the cells of the outer part of the root to the xylem, which occupies the central part of the root. The main pathway for water diffusion in the outer part of the root is the apolast, a continuous collection of cell walls. However, in the endoderm (a cylindrical layer of cells surrounding the conductive tissue), free diffusion along the cell walls encounters an obstacle - a waterproof cork layer of the Casparian girdle. Water must change its path here and pass through the membrane and protoplast of endoderm cells, which thus plays the role of an osmotic barrier between the root cortex and its central cylinder. In monocots, the inner tangential walls of the cells also become corky, but these walls are permeated with pores, through which, like channels, water can pass.
Water rises through the xylem to the aerial parts of the plant. Xylem is made up of several types of cells. Water moves in it mainly through vessels and tracheids. Both those and other cells are perfectly adapted for this purpose: they are elongated, devoid of living content and hollow inside, that is, they are like tubes for water. The lignified secondary cell walls are strong enough to withstand the enormous pressure difference that occurs when water rises to the tops of tall trees. The end, and sometimes the side walls of the segments of the vessels are perforated; vessels, consisting of segments connected end to end, form long tubes through which water with minerals dissolved in it easily passes. There are no perforations in the tracheids, and water, in order to get from one tracheid to another, must pass through their end walls; however, tracheids are very long cells, and therefore this design is quite well adapted for conducting water.
Flowering plants have both vessels and tracheids; in more primitive forms of vessels, as a rule, they do not.
Leaf veins, consisting of strands of xylem and phloem, form such a dense network in the leaf that any of its cells is close enough to the water source. From the xylem, water diffuses into the cell walls of the mesophyll. Thus, water in the liquid phase fills all the way from the soil - through the root and stem - to the mesophyll cells in the leaf.
New in blogs
The total water flow is always directed towards a lower water potential, i.e., it is maximum in the soil, somewhat lower in the root cells, and lowest in the cells adjacent to the leaf epidermis. The low value of the indicator in these last cells is mainly due to the evaporation of water from the leaf surface.
Water passes from the plant into the surrounding air mainly in the vapor state. In the mesophyll of the leaf there are extensive intercellular spaces, and each cell of the mesophyll borders on such an intercellular space at least on one of its sides. Due to the evaporation of water from moist cell walls, the air in the intercellular spaces is saturated with water vapor, and some of these vapors are lost - they go outside. Since the cells of the epidermis of most plants are covered with a waxy waterproof cuticle, water vapor exits the leaf into the atmosphere mainly through stomata.
Guttation is the release of drop-liquid moisture by leaves through hydathodes under conditions of hindered evaporation. Weeping of a plant is the outflow of apiary (water with minerals dissolved in it, located in the xylem) from the stems of plants with cut shoots. Guttation usually takes place under conditions of high air humidity, when the upper terminal motor of the water current is "turned off" - transpiration is the physiological process of water evaporation by the above-ground plant organs. It is carried mainly from the leaves through the stomata and cuticle.
Water makes up to 95% of the mass of plants; all life processes take place in it or with its use. Therefore, water is a necessary condition for the life of the body. With a lack of water, the plant's metabolism is disturbed.
- Water provides the flow of nutrients and minerals through the conducting system of the plant.
- Seed germination depends on the availability of water.
- Water is involved in the process of photosynthesis.
- Aqueous solutions that fill the cells and intercellular spaces provide the plant with elasticity, so the plant retains its shape.
The plant must absorb water. Otherwise, sooner or later, his life will end. Typically, a plant absorbs water exclusively through its root system from the soil. Root hairs are involved in this. Leaves evaporate water through stomata. The point of absorbing excess water in order to evaporate it later, for the most part, is that the flow of water ensures the transport of substances.
If the evaporation of water by the plant exceeds the intake of water, then the plant wilts. This often happens during the day when it is hot. At night, the plant makes up for the deficiency, as evaporation is reduced at this time of day.
Water is absorbed into the plant by osmosis. During osmosis, water, in which there are fewer dissolved substances, is, as it were, sucked into solutions that are more saturated with substances.
How does water exchange occur in plants: processes and movement of water through plants
Cellular solutions of plants are more saturated, so the cells absorb water.
As a result of the constant absorption and evaporation of water in the plant, there is a constant water exchange, which includes three stages: the absorption of water by the roots, its movement through the vessels of the conductive tissue, and the evaporation of water by the leaves. The flow of water goes through all the organs of the plant. How much water a plant absorbs, approximately how much it evaporates. Only a fraction of a percent of the incoming water goes to the synthesis of substances. These are quite large volumes of water. So, for example, only one wheat plant in the field evaporates about 50 g of water per day.
When the roots absorb water, they absorb dissolved mineral salts along with it. When water evaporates, salts are no longer present in it, they remain in the plant and are used in metabolism.
Water flows from bottom to top. Its strength depends on the intensity of absorption by the roots and evaporation by the leaves. The water current unites all the organs of the plant, carries various compounds, and nourishes the cells with water.
New in blogs
Lower and upper end motors. Root pressure, its mechanism and significance in plant life
As a result of the active work of ion pumps in the root and the osmotic (passive) entry of water into the xylem vessels, hydrostatic pressure develops in the vessels, which is called root pressure. It provides the rise of the xylem solution through the vessels of the xylem from the root to the aerial parts. It has been shown that in plants living in cold and poorly aerated soils, as well as as a result of the action of poisons and inhibitors, the root pressure is reduced. The mechanism for raising water over a plant due to developing root pressure is called bottom end motor.
top end motor, ensuring the movement of water up the plant, is created and maintained by the high suction force of the transpiring cells of the leaf parenchyma.
Transpiration - this is the physiological process of evaporation of water by a plant. The main organ of transpiration is the leaf.
The plant has a very large leaf area, which creates a huge evaporation surface. As a result of the loss of water by the cells of the leaves, the water potential in them decreases, i.e., the sucking power increases. This leads to an increase in the absorption of water from the xylem of the veins by the leaf cells and the movement of water along the xylem from the roots to the leaves.
The strength of the upper end motor will be the greater, the more active transpiration. The upper end motor can operate when the lower end motor is completely turned off, and not metabolic energy is used for its operation, but the energy of the external environment - temperature and air movement.
Previous12345678910111213Next
VIEW MORE:
Plant cells use osmosis to increase the volume of the vacuole so that it expands the cell walls (tug pressure). Plant cells do this by storing sucrose. By increasing or decreasing the concentration of sucrose in the cytoplasm, cells can regulate osmosis. Due to this, the elasticity of the plant as a whole increases. Many movements of plants are associated with changes in turgor pressure (for example, movements of the whiskers of peas and other climbing plants). Freshwater protozoa also have a vacuole, but the task of protozoan vacuoles is only to pump out excess water from the cytoplasm to maintain a constant concentration of substances dissolved in it.
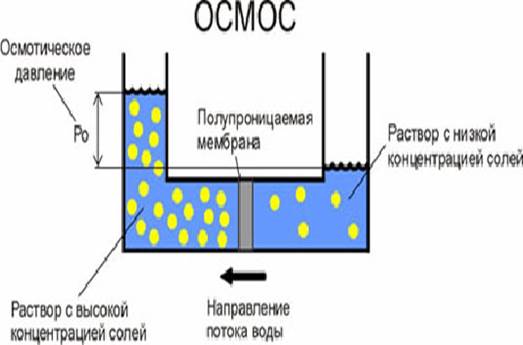
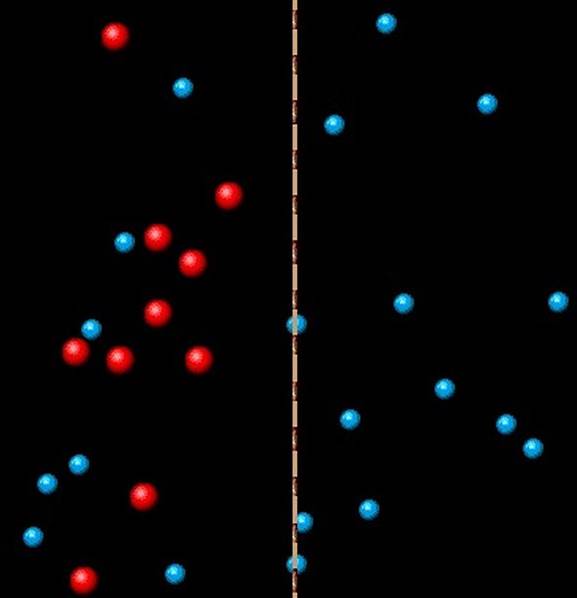
Osmosis through a semi-permeable membrane. Solvent particles (blue) are able to cross the membrane, solute particles (red) are not.
In this chapter, we dealt with osmosis, one of the most important devices for transporting water.
2.3. Transpiration as a mechanism for transporting water
The beginning of the study of transpiration began in the 18th century, but the scientific approach to understanding this phenomenon began in the middle of the 19th century. One of the first researchers of transpiration was G. Mol (1856), who proved that the size of the stomatal openings is determined by the turgor of guard cells and depends on light, heat and humidity. He also conducted experiments, as a result of which he proved that the presence in the guard cells of chloroplasts that synthesize osmotic substances, and thus affect the functioning of stomata and transpiration.
Research work: the movement of water in plants
Other scientists studying transpiration were. Unger (1857), who published a major work on transpiration in 1862. S. Schwendener (1883) suggested that stomata provide not only evaporation, but also the assimilation of CO2. The idea of the active role of trailing, rather than adjacent epidermal cells, as Deitgeb (1886) believed, was finally approved by the son of Charles Darwin F. Darwin (1898). The effect of various rays of the spectrum on the work of stomata was first studied by Kohl (1895). He found that red and blue rays, that is, rays absorbed by chlorophyll, cause the stomata to open. In addition to stomatal transpiration in 1878, cuticular transpiration was also discovered (Khenel). Determinations of the amounts of evaporated water (Gaberlandt, 1877; Henel, 1879, 1880) showed that this value is different depending on the nature of the plant itself and the conditions of its growth. But first things first.
Transpiration is the process by which plants lose water in the form of vapour. The main organ for transpiration is the leaf. Transpiration is analogous to the physical process of evaporation. Transpiration is one of the most important factors in the water regime of plants, since evaporation creates a certain energy impulse, which causes the movement of water through the plant. In this regard, transpiration determines the rate of water uptake by the plant and causes water deficiency in the leaves. Also, due to the loss of water during the process of transpiration, the sucking force increases. This leads to an increase in the absorption of water from the vessels by the leaf cells and the movement of water along the xylem from the roots to the leaves. The process of transpiration can be carried out only when the lower terminal motor is completely turned off for the operation of the upper terminal motor. Two types of transpiration are known to science: cuticular and stomatal. Cuticular transpiration is a mechanism in which the leaves must have a single layered epidermis, the outer cell walls of which are covered with cuticle and wax, forming a barrier to water movement. Hairs are often developed on the surface of the leaves, which also affect the water regime of the leaf, as they reduce the speed of air movement above its surface and scatter light, thereby reducing water loss due to transpiration. Stomatal transpiration. Transpiration through the stomata goes almost at the same rate as normal. Transpiration saves plants from overheating.
So let's sum it up. We have learned that transpiration is basically a physical process of evaporation that is controlled by physical factors. However, transpiration is also a physiological process, which is influenced by such internal factors as the structure and arrangement of leaves, and the behavior of stomata. Transpiration usually occurs in two stages: the evaporation of water from the cell walls into the intercellular spaces and the diffusion of water vapor into the outer atmosphere.
2.4. Root pressure as a mechanism for water transport
Root pressure is one of the most studied physiological processes. It was first described by the English scientist S. Gels in 1727. Interest in studying the mechanisms of this phenomenon does not weaken. In Russia, the famous physiologist D. A. Sabinin was one of the first to study exudation in detail. At first, he considered the root system as an osmotic apparatus, but later came to the conclusion that exudation is a complex physiological process, closely related to cell metabolism and their polarity. At present, a growing number of plant physiologists recognize the osmotic concept of exudation as insufficient to explain the work of the root. Numerous experiments have shown that the activity of the root system consists of two components: osmotic and metabolic. Root pressure is the pressure in the conductive vessels of the roots, which, along with transpiration, ensures the supply of water to aboveground organs. It arises mainly as a result of an increase in the osmotic pressure in the root vessels (usually 1-3 atmospheres) over the osmotic pressure of the soil solution as a result of the active release of mineral and organic substances into the vessels by the root cells. The reverse flow of fluid from the vessels is prevented by a layer of endoderm cells with corked (soaked in suberin) membranes. The result of high root pressure is the “weeping” of plants, as well as the well-known to the inhabitants of Belarus, the USA, Canada and other countries, the spring outflow of juice when the trunks are cut near birch and maple. Root pressure is also of great importance in the absorption of water by the plant in the spring before the leaves open and in seedlings during underground germination. Its role is great in the restoration of torn strands in the vessels of the xylem, through which there is an upward flow of water (from the roots to the leaves). At night, root pressure eliminates the water deficit that has arisen during the day. The suction action of transpiration is transferred to the roots in the form of hydrodynamic tension, which links the operation of these two motors.
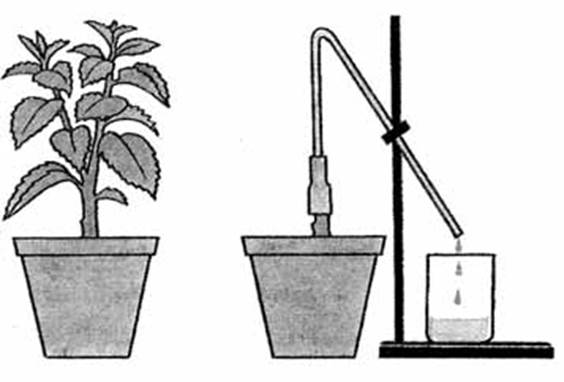
An experiment demonstrating root pressure
3. Conclusion
ADD A COMMENT[possible without registration]
before publication, all comments are considered by the site moderator - spam will not be published