Plant root structure
Main functions of the root: ensures anchoring of the plant in the soil, absorption of soil aqueous solution salts and its transport to the above-ground parts of the plant.
Additional functions: storage of nutrients, photosynthesis, respiration, vegetative propagation, excretion, symbiosis with microorganisms, fungi. The first true roots appeared in ferns.
The root embryo is called the embryonic root and is formed simultaneously with the bud in the seed embryo.
In plants there are:
Main root. It is formed from the embryo and persists throughout life. Always alone.
Lateral roots. They branch from the roots (main, additional, lateral). When branching, they form roots of the 2nd, 3rd, etc. order.
Adventitious roots. They are formed in any part of the plant (stem, leaves).
The totality of all the roots of a plant forms the root system. The root system is formed throughout the life of the plant. Its formation is ensured mainly by lateral roots. There are two types of root systems: taproot and fibrous.
The growth of the root and its branching continues throughout the life of the plant organism, that is, it is practically unlimited. Meristems - educational tissues - located at the apex of each root. The proportion of meristematic cells is relatively large (10% by mass versus 1% in the stem).
Determining the size of root systems requires special methods. It turned out that total surface roots usually exceeds the surface of above-ground organs by 104-150 times. When growing a single rye plant, it was found that the total length of its roots reaches 600 km, and 15 billion root hairs are formed on them. These data indicate the enormous potential for growth of root systems. However, this ability is not always manifested. When plants grow in phytocenoses with a sufficiently dense structure, the size of the root systems noticeably decreases.
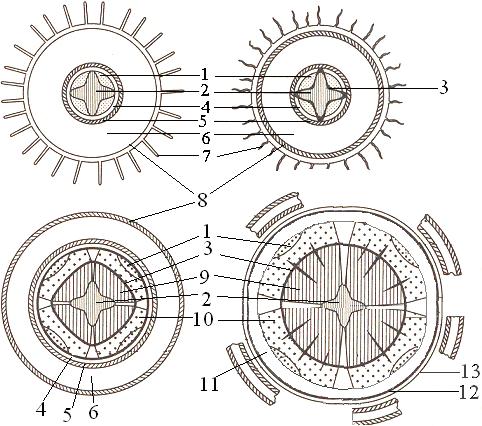
From a physiological point of view, the root system is not homogeneous. Not the entire root surface is involved in the absorption of water. Each root has several zones (Fig. 1). True, not all zones are always expressed equally clearly.
The end of the root is protected from the outside by a root cap, resembling a rounded cap, melting from living thin-walled oblong cells. The root cap serves as protection for the growing point. The cells of the root cap peel off, which reduces friction and facilitates the penetration of the root deep into the soil. The meristematic zone is located under the root cap. The meristem consists of numerous small, rapidly dividing, densely packed cells, almost entirely filled with protoplasm. The next zone is the stretch zone. Here the cells increase in volume (stretch). At the same time, differentiated sieve tubes appear in this zone, followed by a zone of root hairs. With a further increase in cell age, as well as the distance from the root tip, root hairs disappear, cutinization and suberization of cell membranes begins. Water absorption occurs mainly by the cells of the elongation zone and the root hair zone.
Rice. 1. Root structure diagram:
A - longitudinal section: 1-root cap; 2- meristem; 3-stretch zone; 4- zone of root hairs; 5- branching zone;
B - cross section (according to M.F. Danilova): 1 - rhizoderm; 2 - root hair; 3 - parenchyma; 4 - endoderm; 5- Casparian belts; 6 - pericycle; 7 - phloem; 8 - xylem. Dotted arrows represent the paths of movement of substances absorbed from the external solution. Solid arrows are the path of solutions along the simplast; intermittent - path along the apoplast.
The root surface in the area of root hairs is covered with rhizoderm. This is a single-layer tissue with two types of cells that form and do not form root hairs. It has now been shown that the cells that form root hairs have a special type of metabolism. In most plants, rhizoderm cells have thin walls. Following the rhizoderm to the pericycle are the cells of the cortex; the cortex consists of several layers of parenchyma cells. An important feature of the cortex is the development of systemic large intercellular spaces. At the border of the cortex and central cylinder One layer of cells tightly adjacent to each other develops - endoderm, which is characterized by the presence of Casparian belts. The cytoplasm in endodermal cells is tightly adjacent to the cell membranes. As we age, the entire inner surface of endodermal cells, with the exception of passage cells, becomes covered with suberin. With further aging, more layers may be added on top. Apparently, it is the endodermal cells that serve as the main physiological barrier to the movement of both water and nutrients. The central cylinder contains the conducting tissues of the root. When considering the structure of the root in the longitudinal direction, it is important to note that the beginning of the growth of root hairs, the appearance of Casparian hairs in the walls of the endoderm and the differentiation of xylem vessels occur at the same distance from the apical meristem. It is this zone that is the main zone for supplying plants with nutrients. Typically the absorption zone is 5-10 cm in length. Its magnitude depends on the growth rate of the root as a whole. The slower the root grows, the shorter the absorption zone.
The length of the root can be divided into several sections that have different structures and perform various functions. These areas are called root zones. The root cap and the following zones are distinguished: division, extension, absorption and conduction.
Differentiation of root tissue occurs in the absorption zone. These are primary tissues in origin, since they are formed from primary meristem growth cone. Therefore, the microscopic structure of the root in the absorption zone is called primary. U monocots primary structure persists in the venue area. Here, only the most superficial layer with root hairs is missing - the rhizoderm (epiblema). Protective function performed by the underlying tissue - exodermis.
The primary structure of the root is divided into three parts: the rhizoderm, the primary cortex and the axial (central) cylinder.
The primary cortex accounts for the bulk of the primary root tissues. Its cells accumulate starch and other substances. This tissue contains numerous intercellular spaces, which are important for aeration of root cells. The outer cells of the primary cortex, lying immediately below the rhizoderm, are called exoderm. The bulk of the cortex (mesoderm) is formed by parenchyma cells. The innermost layer is called endoderm. This is a series of tightly closed cells (without intercellular spaces).
The central or axial cylinder (stele) consists of conducting tissues surrounded by one or several layers of cells - the pericycle.
The inner part of the central cylinder in most plants is occupied by a continuous strand of primary xylem, which gives projections in the form of ribs to the pericycle. Between them are strands of primary phloem.
In dicotyledonous and gymnosperm plants, already at an early age, a cambium appears in the central cylinder of the root between the xylem and phloem, the activity of which leads to secondary changes and ultimately the formation secondary structure root The cambium deposits secondary xylem cells towards the center, and secondary phloem cells towards the periphery. As a result of the activity of the cambium, the primary phloem is pushed outward, and the primary xylem remains in the center of the root.
Following changes in the central cylinder of the root, changes occur in the cortex. The cells of the pericycle begin to divide along the entire circumference, resulting in the formation of a layer of cells of the secondary meristem - phellogen (cork cambium). The phellogen, in turn, dividing, deposits the phellem outward and the phelloderm inward. The periderm is formed, the cork layer of which isolates the primary cortex from the central cylinder. As a result, the entire primary crust dies and is gradually shed; The periderm becomes the outer layer of the root. The phelloderm cells and remnants of the pericycle subsequently grow and form a parenchymal zone, which is called the secondary root cortex (Fig. 2).
With the development of the storage parenchyma of the main root, the formation of storage roots or root crops occurs. Root vegetables are distinguished:
1. Monocambial (radish, carrot) - only one layer of cambium is laid, and reserve substances can accumulate either in the xylem parenchyma (xylem type - radish) or in the phloem parenchyma (phloem type - carrot);
2. Polycambial - at certain intervals, a new layer of cambium (beets) is formed.
Rice. 2. Transition from the primary structure of the root to the secondary:
1 - primary phloem, 2 - primary xylem, 3 - cambium, 4 - pericycle, 5 - endoderm, 6 - mesoderm, 7 - rhizoderm, 8 - exoderm, 9 - secondary xylem, 10 - secondary phloem, 11 - secondary cortex, 12 - phellogen, 13 - phellem.
It should be noted that in general, root systems are much less diverse compared to above-ground organisms, due to the fact that their habitat is more homogeneous. This does not exclude the possibility that root systems change under the influence of certain conditions. The influence of temperature on the formation of root systems is well demonstrated. As a rule, the optimal temperature for the growth of root systems is slightly lower compared to the growth of above-ground organs of the same plant. Nevertheless, a strong decrease in temperature noticeably inhibits root growth and promotes the formation of thick, fleshy, poorly branched root systems.
Great importance Soil moisture plays a role in the formation of root systems. The distribution of roots across soil horizons is often determined by the distribution of water in the soil. Usually, in the first period of a plant organism’s life, the root system grows extremely intensively and, as a result, reaches more moist layers of the soil more quickly. Some plants develop shallow root systems. Situated close to the surface, strongly branching roots intercept atmospheric precipitation. In dry areas, deep- and shallow-rooted plant species often grow side by side. The former provide themselves with moisture from the deep layers of the soil, the latter from the absorption of precipitation.
Important for the development of root systems is aeration. It is the lack of oxygen that causes poor development of root systems in marshy soils. Plants adapted to grow on poorly aerated soils have a system of intercellular spaces in their roots, which, together with the intercellular spaces in the stems and leaves, form a single ventilation system.
Are of great importance nutritional conditions. It has been shown that the application of phosphorus fertilizers promotes the deepening of root systems, and the application of nitrogen fertilizers promotes their increased branching.
Modification of roots
The root of a plant has many functions. The most important of them for the life of the plant are the retention of the plant in the soil and the absorption of water with dissolved minerals. In addition to the above-mentioned roots, they also tend to perform a number of other functions, which is why their structure changes. Due to metamorphosis, the modified root loses its similarity with a regular root.
Root vegetable
In some biennial plants (turnips, parsley, carrots), the root is transformed into a root vegetable, which is a thickened adventitious root. The main root and the lower part of the stem take part in the formation of the root crop. In the structure of the root crop, the main place belongs to the storage tissue.
Depending on the structure of the root, there are three types of root vegetables: carrot, beet and radish.
1. Carrot-type root vegetables- vegetables with an elongated root shape, which can be cylindrical, conical, elongated conical, spindle-shaped and blunt or sharp at the end. Root crops of this type have a clearly demarcated bark (phloem) and a core (xylem). Between them is the cork cambium. The top of the root crop is covered with natural periderm. In terms of composition and amount of nutrients, the bark is more valuable than the pith. Root vegetables of this type include carrots, parsley, celery, and parsnips.
2. Beet-type root vegetables- vegetables with round, round-flat, oval or elongated root vegetables. Represented by table beet and sugar beet. Only table beets are used as a vegetable crop. The root vegetable has dark red flesh with rings of lighter toga, which is due to the alternation of xylem (light rings) and phloem (dark rings) tissues. The less specific gravity the xylem occupies, the higher the nutritional value of beets.
3.
Rare root vegetables- vegetables with rounded, turnip-shaped, elongated-conical root crops. Their peculiarity internal structure is the radial arrangement of secondary xylem, phloem and parenchyma tissue. The cambium layer is located directly below the periderm. Root vegetables of this type include radishes, radishes, rutabaga and turnips.
Root crops of all types are characterized by common morphological characteristics: a head in the upper part with leaf petioles and buds at the base, a root body (the main edible part) and a root tip (the main one), and beet-type root vegetables have lateral roots. In other root vegetables, thin lateral roots are easily torn off during harvesting and, as a rule, are absent.
The peculiarity of all root vegetables is their ability to heal mechanical damage by suberinization of cells, as well as their easy digestibility. The most easily withered root vegetables are carrots and radishes; the least susceptible are beets, radishes, turnips and rutabaga.
Root tubers (root cones)
Many angiosperms have root tubers (or root cones) on their roots. Their origin is associated with thickening of adventitious and lateral roots. Root tubers are a reservoir of nutrients. They are also used for vegetative propagation of plants.
The most famous of the plants with a typical root tuber is the dahlia. Swollen root cones extend from one point - the base of the old stem. These modified roots provide the plant with nutrients. During the growth period, they grow their own thin roots, which extract water and nutrients. Plants with root cones are propagated by separate cones with a bud (eye) at the end (dahlia, eremurus, clivia, buttercup).
Cling roots, stilt roots, aerial roots, board-shaped roots, sucker roots
Roots-holds- peculiar adventitious roots that allow the plant to easily attach to any support. Such roots are found in ivy and other climbing plants.
An amazing phenomenon in nature is stilted roots, which serve as a support for the plant. Such roots have the ability to withstand heavy loads due to the presence of mechanical tissues in all sections. They are found in plants of the genus Pandaus, growing on oceanic islands in the tropics, where strong hurricane winds prevail.
Aerial (breathing) roots found in tropical trees that grow in oxygen-depleted soils and freshwater tropical swamps. These are lateral roots located above the ground. Thanks to aerial roots, oxygen and water are absorbed by the plant from the air in conditions of high humidity. Thus, the respiratory roots provide the function of additional breathing.
Board-shaped roots- These are vertical above-ground roots characteristic of large trees growing in tropical rain forests. They develop at the base of the trunk, look like boards adjacent to the trunk, providing the plant with additional support.
Mycorrhiza
Mycorrhiza is a symbiosis of roots higher plants with fungal hyphae. This cohabitation is beneficial for both organisms, since the fungus receives ready-made organic matter, and the plant absorbs water and minerals from the fungus. Mycorrhiza is often found on the roots of higher plants, especially woody ones. You can think of fungal hyphae intertwined with thick tree roots as root hairs, since their functions are similar.
Most perennial plants have mycorrhizae. It is assumed that mycorrhiza is one of the factors that contributed to the progress of flowering plants. Plants that feed on mycorrhizal fungi are called mycotrophic.
Distinguish two main types of mycorrhiza: ectotrophic and endotrophic. The hyphae of ectotrophic mycorrhiza cover the root only from the outside, sometimes penetrating into the intercellular spaces of the bark parenchyma. Ectomycorrhiza is common in many woody plants (pine, birch, oak, beech, etc.), as well as a number of herbaceous agricultural plants, especially cereals. The fungus settles on the sprouted root of the seed and in its further development, especially in the tillering phase, provides abundant soil nutrition for the plant.
Endotrophic mycorrhiza is more common. It is characteristic of most flowering plants. Endomycorrhiza does not form a fungal sheath around the root, root hairs do not die, but hyphae penetrate the root and penetrate into the cells of the bark parenchyma.
Epiphytic tropical orchids and some other plants have so-called aerial roots. They are covered with a multi-layered epiblema called velamen. Velamen sometimes performs a photosynthetic function, and later can participate in the absorption of atmospheric moisture, forming a dead spongy hygroscopic root cover.
Without mycorrhiza, the mycelium of symbiotic fungi can exist in the soil for a certain time, but will never form fruiting bodies. Therefore, under artificial conditions it is impossible to obtain fruiting bodies of porcini mushrooms, russula mushrooms, fly agaric mushrooms - all of them are mycorrhiza-formers, and without a certain tree species they will not bear fruit. In turn, a plant without its fungal symbionts develops poorly, slowly, is easily susceptible to diseases and may even die.
Mycorrhiza is of great importance in tropical rain forests. Due to the strong leaching regime (daily precipitation), these forests are practically devoid of soil (all nutrients are washed out of the soil). Plants are faced with an acute problem of nutrition. At the same time, there is a lot of fresh organic matter: fallen branches, leaves, fruits, seeds. But this organic matter is inaccessible to higher plants, and they come into close contact with saprotrophic fungi. Thus, the main source of minerals under these conditions is not the soil, but soil fungi. Minerals enter the root directly from the hyphae of mycorrhizal fungi, which is why rain forest plants have a shallow root system. How effectively mycorrhiza works can be judged by the fact that tropical rain forests are the most productive communities on Earth; the maximum possible biomass develops here.
Bacterial nodules
Plants can also coexist mutually beneficially with nitrogen-fixing bacteria. This is how bacterial nodules appear on the roots of higher plants - modified lateral roots that have adaptations for symbiosis with bacteria. Through root hairs, bacteria enter young roots and provoke the formation of nodules. The role of these bacteria is that they convert nitrogen from the air into mineral form so that it becomes available for absorption by plants. Plants provide protection for bacteria from competing species of soil bacteria. Bacteria also feed on substances from the roots of higher plants. The appearance of nodule bacteria has been recorded, mainly on the roots of plants from the legume family. Therefore, legume seeds are enriched with protein, and agriculture Representatives of this family are used in crop rotation to enrich the soil with nitrogen.
Tropical rainforests contain plants that grow in the canopy of trees. They are called epiphytes(from the Greek “epi” - on and “phyton” - plant). The roots of such plants are called aerial, since they usually hang from the branches and are able, like blotting paper, to absorb moisture and small particles in the air. For example, many orchids are considered epiphytes.
In a longitudinal section through the tip of the root, the following areas can be found: root cap, growth zone, root hair zone, zone with the structure of an adult root(Fig. 59).
Growth zone located slightly away from the root tip. It is composed of several rows of narrow tabular cells with thin membranes and protoplasm filling the cell cavity. This educational fabric — meristem. In fact, there are two meristems: one of the root tip, the other of the root cap.
The cells of the latter detach new cells outward and form root cap . The outermost cells are rounded, separated and exfoliated under the mechanical influence of solid soil particles. Dead cells are replaced by cells located below them and arising from the meristem of the root cap. Inwardly from the meristem of the root tip, cells of the root proper are deposited by division, from which, as further development and growth, various root tissues are formed. Somewhat above the meristem of the root tip, in a longitudinal section, differentiation of root cells into 3 layers is already noticeable:
- the outer layer, which later develops into the skin - epidermis root;
- middle layer giving further beginning conductive part root;
- an intermediate layer that further develops the so-called root bark(Fig. 60 c, e, d).
With further development, these three zones of the internal structure of the root are well differentiated. On a cross section taken in the area of development of root hairs, the root has the structure shown in Figure 61.
The outside of the root is covered with a single layer epidermis. Epidermal cells are living and have thin walls. At the periphery they are elongated into long outgrowths - root hairs (Fig. 61, A). The cell nucleus moves into the root hair, located at its anterior, growing end, on the lower side (Fig. 62).
Root hairs have a length of 0.15–2.5 mm(in humid air and in an artificial environment can reach 8 mm). Their number on the root is very large - on average there are 8 to 425 pieces per 1 lsh. The root hair zone occupies a section of the root 1-2 long cm. Already at a distance of 3-8 mm from the beginning of the zone, root hairs die off, fall off, and as the root grows closer to the root apex, more and more new ones appear. Root hairs protrude outward various substances(acids) that dissolve water-insoluble compounds present in the soil and make them available to plants. Penetrating between soil particles, root hairs grow tightly together with them. When pulled out of the ground and during further replanting, the plant often suffers or even does not take root at all due to the tearing off of young, growing parts of the root with root hairs.
Behind the root hair zone is located root bark (Fig. 61, To). It is represented by several rows of multifaceted parenchyma cells. The outermost layer of the root cortex is composed of larger, somewhat radially elongated cells, tightly adjacent to the epidermis. As the root hairs fall off, the cell membranes of this layer become suberized; in addition, a layer appears underneath them cork cells, which thus protect the outer layer of the root from evaporation. The deeper layers of the root consist of smaller cells.
The innermost layer of the bark forms the so-called endoderm, separating the bark from the middle part of the root, the so-called central cylinder, or lay down(Fig. 61, en). The cells of the endodermis are located in one layer and closely moved towards each other. Their outer walls are thin, while the inner and radial (side) walls become strongly thickened and, saturated with suberin, become suberized, becoming impenetrable to water and gases. Among the endodermal cells there are individual cells with non-thickened walls, rich in content ( access cells); Through these cells, water penetrates from the cortex of the root into the central cylinder.
The central cylinder, occupying the middle part of the root, consists of various tissues. The outermost layer (rarely two layers) of cells adjacent to the endoderm from the inside is called pericycle(Fig. 61). Its cells are larger, slightly elongated in radius and have thin cell membranes. In this layer, the formation of lateral roots occurs, which is why the pericycle is often called root layer. They are formed in the following way: above a group of vessels in the pericycle, increased division of its cells is observed, due to which a small tubercle is formed, protruding into the peripheral side of the root. On the outside it is surrounded by endodermal cells.
Gradually, layers characteristic of the meristem of the main root are formed in the tubercle, from which the corresponding tissues are subsequently formed. The tubercle continues to grow, surrounded by a layer of endoderm (the so-called “pocket”), and, breaking through the cortex, comes out. Material from the site
Thus, lateral roots, in contrast to the main root and stem, arise endogenous(intragenous). All adventitious roots arise endogenously, no matter from what part of the plant they are formed.
Behind the pericycle there is parenchymal tissue in which conductive elements (vascular bundles). The vascular bundle of the root is built according to radial type(radial beam). Its xylem is located in groups, rays running from the periphery to the center. The number of rays is usually small - 3, 4, 5. Sometimes the xylem groups do not reach the center, which is then filled with parenchyma cells. The vessels of xylem rays have narrower lumens closer to the periphery and larger ones towards the center. The external vessels are formed earlier than the central ones. The development of xylem in the root proceeds from the periphery to the center. External vessels are usually annular or spiral; larger internal ones are porous. Between the xylem rays in the upper part there are sections of phloem, composed of sieve tubes with accompanying cells and phloem parenchyma.
On this page there is material on the following topics:
Root hair structure
Root hairs are highly elongated outgrowths of the outer cells covering the root. The number of root hairs is very large (per 1 mm2 (square) from 200 to 300 hairs). Their length reaches 10 mm. Hairs form very quickly (in young apple tree seedlings in 30-40 hours). Root hairs are short-lived. They die off after 10-20 days, and new ones grow on the young part of the root. This ensures the development of new soil horizons by the roots. The root continuously grows, forming more and more new areas of root hairs. Hairs can not only absorb ready-made solutions of substances, but also contribute to the dissolution of certain soil substances and then absorb them. The area of the root where the root hairs have died is able to absorb water for a while, but then becomes covered with a plug and loses this ability.
The hair shell is very thin, which facilitates the absorption of nutrients. Almost the entire hair cell is occupied by a vacuole, surrounded by a thin layer of cytoplasm. The nucleus is at the top of the cell. A mucous sheath is formed around the cell, which promotes the gluing of root hairs to soil particles, which improves their contact and increases the hydrophilicity of the system. Absorption is facilitated by the secretion of acids (carbonic, malic, citric) by root hairs, which dissolve mineral salts. Root hairs also play a mechanical role - they serve as support for the root tip, which passes between the soil particles.
Root structure.
Root zones. On a cross section of the main root, areas of different structure are noticeable - these are root zones.
Root cap. The root grows in length from its apex, which is covered with a root cap formed by several layers of living cells that protect the roots. The cap is a sensitive zone of the root; in particular, it perceives the force of gravity and determines the direction of root growth.
The division zone is a group of delicate cells of the forming tissue that constantly divide and give rise to all other cells of the root.
The elongation zone is located above the separation zone; here the cells grow, take on their shape and, together with the root cap, move the root deeper into the soil. The division and elongation zone together form the root growth zone. Then the cells begin to change and acquire the appearance and properties inherent in the tissues they will come to - the differentiation zone. Behind it, higher, is the zone of root hairs, or the suction zone. Root hairs are outgrowths of skin cells that are much larger than the size of the cell itself (up to 1 cm). Root hairs absorb a solution of mineral substances and do not live long (up to 20 days). New root hairs are constantly being formed. As the root grows, the suction zone moves deeper into the soil.
The leading zone (lateral root zone) is located above the suction zone. In this zone, the roots are not able to absorb soil solution, therefore there are no root hairs on it, but it conducts various substances further into the above-ground organs. In it, thickening of the root and branching (formation of lateral roots) occurs.
In the zone of root division in the apical meristem, internal tissues arise in a certain sequence and strictly regularly. Moreover, there is a clear division into two departments. From the middle layer of initial cells comes the outer part, which is called periblem . From the upper layer of initial cells comes the internal section, it is called pleroma .
The pleroma subsequently forms the stele ( central cylinder), some of its cells turn into vessels and tracheids, from others sieve tubes arise, from others - core cells, etc.
From the cells of the periblema it is formed primary root cortex , which consists of parenchyma cells of the main tissue.
From dermatogens (outer layer of cells), located on the surface of the root, separates the primary integumentary tissue, which is called epiblema or rhizoderm . Rhizoderm is a single-layer tissue that reaches its full development in the absorption zone.
is the result of differentiation of the apex meristem. In the primary structure of the root in the area of its tip, 3 layers can be distinguished: outer - epibleme , average - primary cortex and the central axial cylinder - stele . See picture below.

In the formed rhizoderm, many thin outgrowths are formed - root hairs (see pictures below).


Root hairs are short-lived. They can actively absorb water and substances dissolved in water only in a growing state. Due to the formation of hairs, the total surface of the suction zone increases by more than 10 times. As a rule, the length of the hairs is no more than 1 mm. They are covered very thin shell, consisting of cellulose and pectin substances.
Water penetrates into the root hair cells passively, namely, due to the difference in osmotic pressure of the soil solution and cell sap. But minerals enter the root hairs as a result active absorption. This process requires energy to overcome the concentration gradient. Once in the cytoplasm, minerals are transferred from the root hair to the xylem from cell to cell. Thanks to root pressure, which is created by the suction force of all root hairs, as well as the evaporation of water from the surface of the plant leaves (transpiration), the movement of the soil solution upwards through the vessels of the root and stem is ensured.
The plant can provide all these energy-intensive processes due to breathing!
As a result of the diffusion of oxygen from the soil into the tissues, respiration occurs. Plants need organic matter to breathe. These organic substances enter the root from the leaves. The energy generated during the breathing process is stored in ATP molecules. This energy will be spent on cell division, growth, synthesis processes, transport of substances, etc. It is for this reason that it is necessary for air to penetrate into the soil, and for this the soil must be loosened. In addition, by loosening the soil, moisture is retained in it, which is why loosening is often called “dry watering.”
The primary cortex, which, as mentioned above, is formed from the periblema, consists of living thin-walled parenchyma cells. In the primary cortex, three distinct layers can be distinguished: endoderm, mesoderm And exodermis.
Endoderm - This is the inner layer of the primary cortex, which is adjacent directly to the central cylinder or stele. The endoderm consists of a single row of cells that have thickenings on the radial walls (also called Casparian belts), alternating with thin-walled passage cells. The endoderm controls the passage of substances from the cortex to the central cylinder and back, the so-called horizontal currents.
The next layer after the endoderm is mesoderm or the middle layer of the primary cortex. The mesoderm consists of cells with a system of intercellular spaces, located loosely. Intense gas exchange occurs in these cells. In the mesoderm, the synthesis of plastic substances occurs and their further movement to other tissues, the accumulation of reserve substances, and mycorrhiza is also located.
The last, outer layer of the primary cortex is called exodermis . The exoderm is located directly under the rhizoderm, and as the root hairs die off, it appears on the surface of the root. In this case, the exodermis can perform the functions cover tissue: thickening and suberization of cell membranes occurs, and the cell contents die. Among these suberized cells, non-suberized passage cells remain. Substances pass through these passage cells.
The outer layer of the stele, which is adjacent to the endodermis, is called pericycle . Its cells retain the ability to divide for a long time. In this layer, the germination of lateral roots occurs, which is why the pericycle is also called the root layer. Characteristic feature roots is the alternation of xylem and phloem sections in the stele. The xylem forms a star. U various groups plants, the number of rays of this star may be different. Between the rays of this star there is phloem. In the very center of the root there may be elements of primary xylem, sclerenchyma or thin-walled parenchyma. Characteristic feature The root, which distinguishes it in anatomical structure from the stem, is the alternation of primary xylem and primary phloem along the periphery of the stele.
This primary root structure is characteristic of young roots in all groups of higher plants. In ferns, horsetails, mosses and representatives of the class of monocotyledonous flowering plants primary structure the roots will remain throughout his life.
Secondary structure of the root.
In gymnosperms and dicotyledonous angiosperms, the primary structure of the root is preserved only until the process of its thickening begins. This process is the result of the activity of secondary lateral meristems - cambium And phellogen (or cork cambium).
The beginning of the process of secondary changes is the appearance of layers of cambium under areas of the primary phloem, directed inward from it. The cambium arises from the poorly differentiated parenchyma of the central cylinder. It deposits elements of secondary phloem (or bast) on the outside, and elements of secondary xylem (or wood) on the inside. At the beginning of this process, the cambium layers are separated, later they close and a continuous layer is formed. This occurs due to the fact that the pericycle cells divide intensively opposite the xylem rays. From the cambial areas that arose from the pericycle, only parenchyma cells, the so-called medullary rays, are formed. But the remaining cells of the cambium form conducting elements: xylem and phloem.