Based on their shape, all bacteria are divided into 3 groups:
- spherical or cocci
- rod-shaped or sticks
- convoluted forms of bacteria.
Cocci have a round, spherical, oval, candle-flame, lanceolate shape and are divided into 6 subgroups based on the connection method.
1 micrococci;
2 diplococci;
3 tetracocci;
4 streptococci;
5 staphylococci;
6 sarcinas.
All cocci are immobile and do not form spores.
Widely distributed in nature. Included in fermented milk starters. May be pathogenic (angina, gonorrhea, meningitis).
Rod-shaped bacteria have an elongated shape. Length is greater than width. They easily change their shape based on living conditions, ᴛ.ᴇ. have polymorphism. Rods are the most common group of all bacteria. They may not be pathogenic, but can cause various diseases (typhoid, dysentery).
Rods can be mobile or immobile, forming or not forming spores. Based on their ability to form spores, rods are divided into three groups:
- bacteria;
- bacilli;
- clostridia.
The convoluted forms of bacteria are divided into three groups:
1. vibrios;
2. spirilla;
3. spirochetes.
All convoluted forms are pathogenic.
Structure and functions of the cell membrane of bacteria.
Cell membrane covers the outside of the cell. It is a dense, elastic structure that can withstand differential pressure, consisting of two parts - an outer part called the cell wall and an inner part - the cytoplasmic membrane (CPM). Both the wall and the membrane have pores (holes) through which nutrients pass into the cell and waste products are removed. In this case, nutrients pass through the pores of the cell wall with a molecular weight of no more than 1000, ᴛ.ᴇ. During feeding, the wall acts as a mechanical sieve. Nutrients pass through the pores of the CPM not by mass, but as needed, ᴛ.ᴇ. it is semi-permeable.
The cell membrane performs a number of important functions:
1 – maintains body shape;
2 – protects the cell from external influences;
3 – participates in cell metabolism, ᴛ.ᴇ. allows nutrients to pass through and excretes waste products;
4 – participates in cell movement. Bacteria deprived of a cell membrane lose mobility;
5 – participate in the formation of the capsule.
According to their shape, all bacteria are divided into 3 groups: - spherical or cocci - rod-shaped or rods - convoluted forms of bacteria. Cocci have a round, spherical, oval, candle-flame, lanceolate shape and are divided into 6 subgroups depending on the method... [read more].
The microbes most commonly found during food preparation are bacteria, molds, yeasts and viruses. Most microbes are single-celled organisms, the size of which is measured in micrometers - microns (1/1000 mm) and nanometers - nm (1/1000 microns).
Bacteria are single-celled, most studied microorganisms with a size of 0.4-10 microns. According to their shape they are divided into cocci- spherical microbes (micrococci, diplococci, tetracocci, sarcins, streptococci, staphylococci), sticks(single, double, chains), vibrios, spirilla And spirochetes(curved and spirally crimped shapes). The size and shape of bacteria can change depending on various environmental factors (Fig. 3).
Rice. 3. Forms of bacteria:
1 - micrococci; 2 - streptococci; 3 - sarcins; 4 - sticks without spores;
5 - rods with spores (bacilli); 6 - vibrios; 7 - spirochetes;
8 - spirilla.
Bacteria are covered with a membrane, which is a compacted layer of cytoplasm that gives the cell its shape. The outer layer of the shell of many bacteria can mucus, forming a protective cover - a capsule. The main part of the cell is the cytoplasm - a transparent protein mass soaked in cell sap. The cytoplasm contains nuclear matter, reserve nutrients (starch grains, fat droplets, glycogen, protein) and other cellular structures. On the surface of some bacteria (rod-shaped) there are thread-like formations - flagella (single, in the form of a bundle or over the entire surface), with the help of which they move.
Some rod-shaped bacteria, under unfavorable conditions, form spores (condensed cytoplasm covered with a dense membrane). The spores do not require nutrition and are not able to reproduce, but remain viable at high temperatures, drying, freezing for several months (botulinus bacillus) or even many years (anthrax bacillus). Spores die during sterilization (heating to 120°C for
29 min). Under favorable conditions, they germinate into an ordinary (vegetative) bacterial cell. Spore-forming bacteria are called bacilli.
Bacteria reproduce by simple division. Under favorable conditions, reproduction of one cell occurs within 20 -
30 min. With the accumulation of harmful waste products of bacteria and the depletion of nutritional resources, the reproduction process stops.
Molds are unicellular or multicellular lower plant organisms that require ready-made food substances and access to air in their life. The cells of mold fungi have the form of elongated intertwining threads - hyphae 1-15 microns thick, forming the body of the mold - mycelium (mycelium), consisting of one or many cells. Fruiting bodies develop on the surface of the mycelium, in which spores ripen (Fig. 4).
In structure, mold cells differ from bacterial cells in that they have one or more nuclei and vacuoles (cavities filled with cellular fluid). Molds reproduce using hyphae and spores.
Molds are widespread in nature. Developing on food products, they form fluffy coatings of different colors. Molds produce substances that give food products a moldy smell and taste. They can develop at low humidity (15%), which explains the molding of dried fruits, crackers,

Rice. 4. Types of molds:
1 - penicillium; 2 - aspergillus; 3 - mukor..
at increased concentrations of salt and acids (on salty and sour foods), at low temperatures, affecting products stored in refrigerators.
Among the molds there are useful ones, used in the production of cheeses (Roquefort, Camembert), citric acid and medicines (penicillin).
Yeasts are single-celled, non-motile microorganisms. Yeast cells up to 15 microns in size come in different shapes: round, oval, rod-shaped (Fig. 5). They have a clearly defined large nucleus, vacuoles and various inclusions in the cytoplasm in the form of droplets of fat, glycogen, etc.
Yeast reproduces in favorable conditions within several hours by the following methods: budding, spores (1 - 112 pieces per cell), division. Yeasts are widespread in nature. They are able to break down (ferment) sugars into alcohol and carbon dioxide. Alcoholic fermentation is used in winemaking, baking and in the production of fermented milk products (kefir, kumiss). Some yeasts are distinguished by a high content of proteins, fats, B vitamins, and minerals, and therefore are used as a food and feed product.
 |
Classification of bacteria by shape
5. Yeast cell shapes:
1 - ovoid; 2 - ellipsoid; 3 - cylindrical (rod-shaped);
4 - spherical; 5 - lemon-shaped; 6 - yeast that reproduces by division and spores.
Viruses are particles that do not have a cellular structure and have a unique metabolism and the ability to reproduce. They come in round, rectangular and thread-like shapes, ranging in size from 8 to 150 nm. They can only be seen using electron microscopes.
⇐ Previous123456789Next ⇒
Date of publication: 2015-11-01; Read: 1474 | Page copyright infringement
Studopedia.org - Studopedia.Org - 2014-2018 (0.001 s)…
Characteristics of molds (part 1)
Molds, or molds, as they are commonly called, are ubiquitous. They belong to different classes of fungi. All of them are heterotrophs and, developing on food products (fruits, vegetables and other materials of plant or animal origin), cause their spoilage.
Classification of bacteria
A fluffy coating, initially white, appears on the damaged surface. This is the mycelium of the mushroom. Soon the plaque turns into various colors from light to dark shades. This coloring is formed by a mass of spores and helps to recognize mold.
The most common molds in grape must are Mucor, Penicillium and Aspergillus.
Mucor belongs to the mucoraceae family of the class of phycomycetes of the subclass of zygomycetes. This mold has a single-celled, highly branched mycelium; asexual reproduction is carried out using sporangiospores, and sexual reproduction is carried out by zygospores. In mucor, the sporangiophores are solitary, simple or branched (Fig. 21).

Fig.21. Phicomycetes:
a - Mucor; b—Rizopus.
The genus Rizopus (rhizopus) also belongs to the same family, differing from mucor by unbranched sporangiophores located in bushes on special hyphae - stolons.
Many mucor mushrooms are capable of causing alcoholic fermentation. Some mucor fungi (Mucor racemosus), developing in sugary liquids, form, when there is a lack of air, yeast-like cells that reproduce by budding, as a result of which they are called mucor yeast.
The molds Penicillium (Fig. 22) and Aspergillus (Fig. 23) belong to the Ascomycetes class. They have multicellular mycelium and reproduce mainly by conidiospores, colored in various colors and formed on characteristically shaped conidiophores. Thus, in Penicillium the conidiophore is multicellular, branched, and tassel-shaped, which is why it is also called a tassel.

Fig.22. Penicillium:
1 - hypha; 2 - conidiophore; 3 - sterigma; 4 - conidiospores.

Fig.23. Aspergillus niger (conidiophore):
1 - sterigma; 2 - conidia.
In Aspergillus, the conidiophore is single-celled, with a swollen apex, on the surface of which there are radially elongated cells - sterigmata with chains of conidiospores.
The fruiting bodies of these fungi are rarely formed and have the form of small balls, inside of which bags with spores are randomly located.
Penicillium and Aspergillus are pathogens that cause spoilage of food and organic materials. Developing on the surface of the wort, on barrels, and on the walls of cellars, they are dangerous enemies of wine production. They can penetrate into barrel staves to a depth of 2.5 cm. Containers contaminated with mold give wines an unpleasant and almost irremovable moldy tone.
Some species of these mushrooms are of technical importance. Thus, Penicillium notatum (penicillium notatum) is used to produce the antibiotic penicillin. Various species of Aspergillus, Penicillium, Botrytis and some other fungi are used to prepare enzyme preparations (nigrin, avamorin). The species Aspergillus niger (Aspergillus niger) is used to produce citric acid, and Aspergillus oryzae (Aspergillus oryzae) is used in the production of the Japanese national alcoholic drink from rice - sake. Both of these species have the ability to saccharify starch and can be used in the production of alcohol instead of malt.
part 1 >>> part 2 >>> part 3
1 2 3 4 5 6 7 8 9
GENERAL MICROBIOLOGY
1. Subject, tasks, sections of microbiology, its connection with other sciences.
Microbiology is the science of living organisms invisible to the naked eye (microorganisms): bacteria, archaebacteria, microscopic fungi and algae, this list is often extended by protozoa and viruses. The area of interest of microbiology includes their systematics, morphology, physiology, biochemistry, evolution, role in ecosystems, as well as possibilities for practical use.
The subjects of microbiology are bacteria, molds, yeasts, actinomycetes, rickettsia, mycoplasmas, and viruses. But since viruses absolutely cannot exist without a living organism, they are studied by an independent science called “virology.”
The purpose of medical microbiology is to study the structure and properties of pathogenic microbes, their relationship with the human body in certain conditions of the natural and social environment, improving microbiological diagnostic methods, developing new, more effective therapeutic and preventive drugs, solving such an important problem as the elimination and prevention of infectious diseases .
Sections microbiology: bacteriology, mycology, virology, etc.
- *General microbiology - studies the patterns of vital activity of all groups of microorganisms, clarifies the role and significance in the natural cycle.
- *Private microbiology – studies the taxonomy of bacteria, causative agents of certain diseases and methods of their laboratory diagnosis.
The broad science of microbiology includes sections:
- *Agricultural microbiology studies the role and formation of soil structure and its fertility, the role of bacteria in plant nutrition.
Develops methods and methods for using bacteria to fertilize soils and preserve feed.
- *Veterinary microbiology – studies microbes that cause diseases in domestic animals, develops methods for diagnosis, prevention and treatment of these diseases.
- *Technical (industrial) microbiology - studies microorganisms that can be used in production processes to obtain biologically active substances, biomass, etc. Many studies occur at the intersection of disciplines (for example, molecular biology, genetic engineering, biotechnology).
- *Sanitary microbiology studies bacteria living in environmental objects, both autochthonous and allochthonous, that can cause environmental pollution and play a certain role in the epidemiology of infections.
- *Environmental microbiology studies the role of microorganisms in natural ecosystems and food chains.
- *Population microbiology clarifies the nature of intercellular contacts and the interconnection of cells in a population.
- *Space microbiology characterizes the physiology of terrestrial microorganisms in space conditions, studies the influence of space on human symbiotic bacteria, and deals with issues of preventing the introduction of space microorganisms to Earth.
- *Medical microbiology - studies microbes that cause diseases in humans. Studies the pathogenesis and clinical picture of diseases, pathogenicity factors. Develops methods for the prevention, diagnosis and treatment of human infectious diseases.
During the existence of microbiology, general, technical, agricultural, veterinary, medical, and sanitary branches were formed.
General studies the most general patterns inherent in each group of listed microorganisms: structure, metabolism, genetics, ecology, etc.
Technical is developing biotechnology for the synthesis by microorganisms of biologically active substances: proteins, nucleic acids, antibiotics, alcohols, enzymes, as well as rare inorganic compounds.
Agricultural studies the role of microorganisms in the cycle of substances, uses them for the synthesis of fertilizers and pest control.
Veterinary studies the causative agents of animal diseases, methods of diagnosis, specific prevention and etiotropic treatment aimed at destroying the causative agent of infection in the body of a sick animal.
Medical microbiology studies pathogenic (pathogenic) and conditionally pathogenic microorganisms for humans, and also develops methods for microbiological diagnosis, specific prevention and etiotropic treatment of infectious diseases caused by them.
Sanitary microbiology studies the sanitary and microbiological state of environmental objects, food products and drinks, and develops sanitary microbiological standards and methods for indicating pathogenic microorganisms in various objects and products
The main stages of the development of microbiology.
The following are distinguished: 5 periods: heuristic, morphological, physiological, immunological, molecular genetic
- Heuristic: IV-III millennium BC. – empirical knowledge. Hippocrates: suggested the nature of the contagiousness of diseases. Facastoro: the idea of living contagion causing disease; recommended isolating sick people and wearing masks
- Morphological: Discovery in 1676 ^ Antony van Leeuwenhoek; production of lenses that magnify 200-300 times. He described and sketched many microorganisms found in various infusions, in well water, on meat and other objects. He called microbes “animalculi.”
- Physiological: Louis Pasteur(1822-1895) French chemist; the founder of microbiology, immunology, biotechnology but also the nature of life; they cause various chemical transformations in the substrates on which they develop; he studied various types of fermentation (alcoholic, butyric acid), proved the existence of anaerobic organisms
A significant contribution to microbiology was the research of the German scientist Robert Koch (1843-1910).He introduced dense nutrient media for growing microbes into practice; this made it possible to develop methods for isolating (isolating) microbes into “pure cultures,” that is, cultures of each species separately, developed in one cell. Introduced coloring with aniline dyes. Microphotographs. He studied the causative agents of anthrax, tuberculosis, cholera and other infectious diseases; Formulated the Koch-Henle triad: find, prove, destroy. In 1905 - Nobel Prize.
- Immunological: Numerous discoveries in the field of microbiology in the second half of the 19th century.
Classify bacteria according to their shape
contributed to the beginning of the rapid development of immunology.
^ I. I. Mechnikov(1845-1916) developed the phagocytic theory of immunity - the body's immunity to infectious diseases. He came up with the idea of using antagonistic relationships between microbes, which formed the basis of the modern doctrine of antibiotics; the development of microbiology in Russia is associated with it; he organized the first bacteriological laboratory in Russia (in Odessa). In 1903 - Nobel Prize. Paul Ehrlich: German chemist. Developed the theory of the body's humoral defense with antibodies. Received the Nobel Prize in 1908. - Molecular genetic: Stanley Prusiner: American biologist. Discovered prions, endogenous cellular formations associated with errors in protein biosynthesis, which are caused by gene mutation, translation errors, and proteolysis processes N. F. Gamaleya(1859 - 1949) studied issues of medical microbiology; opened a rabies vaccination station; described the phenomenon of bacteriophages
3. Classification of microorganisms. Differences between eukaryotes, prokaryotes and viruses.
Microbes or microorganisms(bacteria, fungi, protozoa, viruses), systematized according to their similarities, differences and relationships with each other. This is the subject of a special science - taxonomy of microorganisms. Systematics has three parts: classification, taxonomy and identification. The taxonomy of microorganisms is based on their morphological, physiological, biochemical and molecular biological properties. The following taxonomic categories are distinguished: kingdom, subkingdom, department, class, order, family, genus, species, subspecies, etc. Within a particular taxonomic category, taxa are distinguished - groups of organisms united by certain homogeneous properties.
Microorganisms are represented by precellular forms (viruses - kingdom Vira) and cellular forms (bacteria, archaebacteria, fungi and protozoa). There are 3 domains(or "empires"): "Bacteria", "Archaea" and "Eukarya":
domain “Bacteria” - prokaryotes, represented by real bacteria (eubacteria);
domain “Archaea” - prokaryotes, represented by archaebacteria;
“Eukarya” domain - eukaryotes, whose cells have a nucleus with a nuclear envelope and nucleolus, and the cytoplasm consists of highly organized organelles - mitochondria, Golgi apparatus, etc. The “Eukarya” domain includes: the kingdom Fungi (fungi); animal kingdom Animalia (includes protozoa - subkingdom Protozoa); plant kingdom Plante. Domains include kingdoms, phyla, classes, orders, families, genera, and species.
View. One of the main taxonomic categories is the species (species). A species is a collection of individuals united by similar properties, but differing from other representatives of the genus.
Pure culture. A set of homogeneous microorganisms isolated on a nutrient medium, characterized by similar morphological, tinctorial (relation to dyes), cultural, biochemical and antigenic properties, is called a pure culture.
Strain. A pure culture of microorganisms isolated from a specific source and different from other members of the species is called a strain. A strain is a narrower concept than a species or subspecies.
Clone. Close to the concept of a strain is the concept of a clone. A clone is a collection of descendants grown from a single microbial cell.
To designate certain groups of microorganisms that differ in certain properties, the suffix is used var(variety) instead of the previously used type.
4. Classification of bacteria. Principles of modern taxonomy and nomenclature, basic taxonomic units. The concept of species, variant, culture, population, strain.
The best known phenotypic classification of bacteria is based on the structure of their cell wall.
The largest taxonomic groups in it were 4 divisions: Gracilicutes (gram negative), Firmicutes (gram-positive), Tenericutes (mycoplasma; single class department Mollicutes) And Mendosicutes (archaea) Mollicutes -Mycoplasmas - prokaryoticunicellular, gram-negative microorganisms, not having cell wall, which were discovered during the study pleuropneumonia at cows.
Mycoplasmas, apparently, are the simplest self-reproducing living organisms; the volume of their genetic information is 4 times less than that of Escherichia coli .
Numerous microorganisms (bacteria, fungi, protozoa, viruses) are strictly systematized in a certain order according to their similarities, differences and relationships with each other. This is the subject of a special science called taxonomy of microorganisms.
The branch of systematics that studies the principles of classification is called taxonomy (from the Greek.
taxis. location, order). Taxon. a group of organisms united by certain homogeneous properties within a particular taxonomic category. The largest taxonomic category is the kingdom, with smaller ones. subkingdom, department, class, order, family, genus, species, subspecies, etc. The formation of names of microorganisms is regulated by the International Code of Nomenclature (zoological, botanical, nomenclature of bacteria, viruses). The taxonomy of microorganisms is based on their morphological, physiological, biochemical, and molecular biological properties.
According to modern taxonomy, pathogenic (disease-causing) bacteria belong to the superkingdom of prokaryotae (Procaryotae), the kingdom of eukaryotes (Eucaryotae), fungi - to the kingdom of Mycota, protozoa - to the kingdom of Protozoa, viruses - to the kingdom of Vira.
View - a set of microorganisms that have a common root of origin and the closest possible phenotypic characteristics and properties. ( View - an evolutionarily established set of individuals that have a single type of organization, which under standard conditions is manifested by similar phenotypic characteristics: morphological, physiological, biochemical, etc.)
Population - a collection of individuals of the same species living within a biotope (a geographically limited area of the biosphere with relatively homogeneous living conditions).
Strain - pure cultures of microbes of the same species obtained from different sources or from the same source at different times.
Pure culture - a population consisting of individuals of one species. (from one microbial cell on an artificial nutrient medium).
5. Microscopy methods. Microscopic method for diagnosing infectious diseases.
Luminescent (or fluorescent) microscopy. Based on the phenomenon of photoluminescence.
Luminescence- glow of substances that occurs after exposure to any energy sources: light, electron rays, ionizing radiation. Photoluminescence- luminescence of an object under the influence of light. If you illuminate a luminescent object with blue light, it emits rays of red, orange, yellow or green. The result is a color image of the object.
Dark-field microscopy. Dark-field microscopy is based on the phenomenon of light diffraction under strong lateral illumination of tiny particles suspended in a liquid (Tyndall effect). The effect is achieved using a paraboloid or cardioid condenser, which replaces a conventional condenser in a biological microscope.
Phase contrast microscopy. A phase contrast device makes it possible to see transparent objects through a microscope. They acquire high image contrast, which can be positive or negative. Positive phase contrast is a dark image of an object in a bright field of view, negative phase contrast is a light image of an object on a dark background.
For phase-contrast microscopy, a conventional microscope and an additional phase-contrast device, as well as special illuminators, are used.
Electron microscopy. Allows you to observe objects whose dimensions lie beyond the resolution of a light microscope (0.2 microns). An electron microscope is used to study viruses, the fine structure of various microorganisms, macromolecular structures and other submicroscopic objects.
In everyday practice of a bacteriological laboratory, microscopic examination is usually used for rapid indicative diagnosis.
The main tasks of microscopy are: identification of the pathogen in clinical material, tentative identification based on the determination of characteristic morphological and tinctorial signs of microorganisms, as well as the study of stained smears from colonies of pure cultures. For some infectious diseases, the causative agents of which are characterized by specific morphology (protozoal diseases, helminthiasis, fungal diseases, spirochetosis), microscopic examination is the main or one of the main diagnostic methods.
Material for microscopic examination can include blood, bone marrow, CSF, punctate lymph nodes, feces, duodenal contents and bile, urine, sputum, genitourinary tract discharge, tissue biopsies, smears from mucous membranes (oral cavity, tonsils, nose, vagina and etc.).
6. Methods for staining microbes and their individual structures.
Coloring methods. The smear is stained using simple or complex methods. Simple ones involve coloring the preparation with one dye; complex methods (according to Gram, Ziehl-Nielsen, etc.) include the sequential use of several dyes and have differential diagnostic value. The relationship of microorganisms to dyes is regarded as tinctorial properties. There are special staining methods that are used to identify flagella, cell walls, nucleoids, and various cytoplasmic inclusions.
In simple methods, the smear is stained with any one dye, using aniline dyes (basic or acidic). If the coloring ion (chromophore) is a cation, then the dye has basic properties; if the chromophore is an anion, then the dye has acidic properties. Acid dyes - erythrosine, acid fuchsin, eosin. The main dyes are gentian violet, crystal violet, methylene blue, basic fuchsin. Mainly for staining microorganisms, basic dyes are used, which bind more intensely to the acidic components of the cell. Saturated alcohol solutions are prepared from dry dyes, sold in the form of powders, and aqueous-alcohol solutions are prepared from them, which are used to stain microbial cells. Microorganisms are stained by pouring dye onto the surface of a smear for a certain time. Staining with basic fuchsin is carried out for 2 minutes, with methylene blue for 5-7 minutes. The smear is then washed with water until the flowing streams of water become colorless, dried by careful blotting with filter paper and microscoped in an immersion system. If the smear is correctly stained and washed, the field of view is completely transparent and the cells are intensely colored.
Sophisticated staining techniques are used to study cell structure and differentiate microorganisms. Stained smears are examined microscopically in an immersion system. Consistently apply certain dyes, differing in chemical composition and color, mordants, alcohols, acids, etc. to the preparation.
1 2 3 4 5 6 7 8 9
Microbiology – a science that studies the structure, properties and vital functions of microorganisms. Food is a favorable breeding ground for the development of microbes, which through their actions can change the properties and quality of food, making it dangerous to human health.
Microbes - single-celled organisms - are widely distributed in soil, water, and air.
Some microbes play a positive role, while others play a negative role.
Morphology of microbes (bacteria, molds, yeasts, viruses)
|
Name of microbes |
Form |
Reproduction method |
|
Bacteria are single-celled microorganisms with a size of 0.4 - 10 microns. |
Divided into: 1) cocci – spherical in shape (micrococci, diplococci, tetracocci) 2) sticks (single, double, chains) 3. vibrios are curved and 4. spirilla spirally twisted 5. Spirochete shapes |
By simple division for 20-30 minutes. |
|
Mold fungi are unicellular or multicellular plant organisms that require food and air access. |
They have the form of elongated interlacing threads with a thickness of 1-15 microns. |
With the help of hyphae and spores. |
|
Yeasts are single-celled, non-motile microorganisms. |
They come in different shapes: round, oval, rod-shaped |
Under favorable conditions, within several hours, by the following methods: budding, spores and division. |
|
Viruses are particles that do not have a cellular structure, have a unique metabolism and the ability to reproduce. |
They come in round, rectangular and thread-like shapes with sizes ranging from 8 to 150 nm. |
Physiology of microbes
Microbes, like all living beings, consist of proteins (6-14%), fats (1-4%), carbohydrates, minerals, water (70-85%), and enzymes.
Water constitutes the bulk of the microorganism cell. Its quantity ranges from 70 to 85% - in vegetative cells and about 50% in spores. All important organic and mineral substances of the microbial cell are dissolved in water and the main biochemical processes occur (hydrolysis of proteins, carbohydrates, etc.).
Proteins - the basis of the life structures of microorganisms. They are part of the cytoplasm, nucleus, membranes and other structures of the cell. 1>Microbe trees are made up of amino acids.
Carbohydrates- are part of the membrane, mucous capsules, proto-plasma and in the form of glycogen grains - a reserve nutrient. Carbohydrates enter the microbial cell from the environment and are used by the cell as an energy source.
Classification and physiology of microorganisms
Cells contain both simple and complex carbohydrates (starch, glycogen, fiber).
Fats- in small quantities they are part of the cytoplasm and nucleus in the form of complex compounds with proteins. Fats serve as a source of energy for microorganisms.
Minerals play an important role in the construction of complex proteins, vitamins, and enzymes of microbial cells. Soluble minerals maintain normal levels of intracellular osmotic pressure (turgor).
Mineral substances of microbes are presented in the form of: phosphorus, sodium, magnesium, iron, sulfur, etc.
Enzymes- substances that accelerate (catalysts) biochemical processes and are located inside the microbial cell. Microbes contain various enzymes, some of which affect biochemical processes inside the cell, others are released outside, processing environmental substances, causing fermentation, rotting and other processes in food products.
Nutrition of microbes. Microbes feed on proteins, fats, carbohydrates, and minerals, which penetrate the cell in dissolved form through the membrane by osmosis (the process of diffusion through a semi-permeable membrane). Proteins and complex carbohydrates are digested by microbes only after they are broken down into simple components by enzymes secreted by microorganisms.
To ensure normal nutrition of microbes, a certain ratio of the concentration of substances is necessary both inside the cell of the microorganism and in the environment. The most favorable concentration is 0.5% sodium chloride in the environment. In an environment where the concentration of soluble substances is much higher (2-10%) than in the cell, water from the cell passes into the environment, dehydration and shrinkage of the cytoplasm occurs, which leads to the death of the microbe. This property of microorganisms is used when preserving foods with sugar (jam) or salt (curing meat, fish).
Respiration of microbes. Microbes need respiration to obtain energy that powers all life processes. According to the method of respiration, microbes are divided into aerobes, those requiring air oxygen (molds, acetic acid bacteria); anaerobes, living and developing in the absence of oxygen (botulinus, butyric acid bacteria), conditional(optional) anaerobes, developing both in the presence of oxygen and without it (lactic acid bacteria, yeast).
Yeast biology
5. Yeast morphology
Macromorphological characters are very variable and strongly depend on the composition of the medium and cultivation conditions, so they have very limited significance in the taxonomy of yeasts. . Yeast cultures growing on solid media...
Vegetative propagation of shrubs
1.2 Methods of propagation of shrubs
Shrubs reproduce by cuttings, seeds, and layering. Seed propagation of most conifers is often difficult due to the low quality and long germination of seeds, as well as the slow growth of seedlings...
Vegetative propagation of conifers
1.2 Methods of propagation of coniferous plants
Seed propagation of most conifers is often difficult due to the low quality and long germination of seeds, as well as the slow growth of seedlings...
Genetically modified organisms. Principles of obtaining, application
1.2.1 Methods for obtaining GM microorganisms
The ability of organisms to synthesize certain biomolecules, primarily proteins, is encoded in their genome. Therefore, it is enough to “add” the desired gene, taken from another organism, to the bacterium...
Microbiology
2. Energy metabolism of microbes. Ways to obtain energy are fermentation, respiration. Types of bacterial respiration
The vital functions of microorganisms: nutrition, respiration, growth and reproduction are studied by physiology. Physiological functions are based on continuous metabolism (metabolism). The essence of metabolism consists of two opposite...
Microbiology of drinking water
1.1 Patterns of quantitative and qualitative content of microorganisms in fresh water bodies depending on various factors
The microflora of various reservoirs contains a sufficient amount of nutrients, which is the main factor promoting the development of microorganisms. The richer it is in organic substances...
Morphology of the internal structure of fish
2.8 Reproductive system and methods of reproduction
The methods of fish reproduction are different. Some are viviparous - active young emerge from the mother's body. The rest are oviparous, i.e. lay eggs that are fertilized in the external environment. The reproductive behavior of some fish is very peculiar...
Morphology and classification of prokaryotes and eukaryotes. Genetics of microorganisms
4. Morphology and classification of eukaryotes (microscopic fungi and yeast)
Eukaryotes (mycelial and yeast fungi). Mushrooms. General characteristics. Mushrooms (Mycota) are a large and diverse group of plant organisms. They do not contain chlorophyll...
1.
Transfer of genetic material in actinomycetes
Transfer of genetic material and genetic mapping in actinomycetes
2. Genetic mapping of actinomycetes
The genetics of actinomycetes has been studied quite well. For the most studied species since the late 50s. Based on conjugation crosses, detailed genetic maps were compiled with many markers applied to them...
Molds
1. Methods of propagation of mold fungi. 2.2. Classification and morphology of bacteria
Methods of formation and reproduction of spores. The importance of asexual sporulation for the identification of fungal genus
Reproduction occurs by division in the transverse direction. When dividing, a bacterium splits into two equal or unequal parts. The resulting two cells are considered as mother and daughter...
Reproduction is one of the fundamental properties of living things. Methods and forms of reproduction of organisms
Section 2. Basic methods and forms of reproduction
The process of reproduction is extremely complex and is associated not only with the transfer of genetic information from parents to offspring, but also with the anatomical and physiological properties of organisms, with their behavior, hormonal control...
The role of microorganisms in the cycle of chemical elements in nature
6. The role of microorganisms in the phosphorus cycle. Different types of bacterial life based on the use of phosphorus compounds
The phosphorus cycle is somewhat different from the cycle of other elements. The release of phosphorus from organic compounds occurs as a result of decay processes. However, microorganisms have not yet been discovered...
Methods of reproduction in various microorganisms, the essence and chemistry of their respiration
2. Characteristics of aerobic and anaerobic microorganisms. The essence and chemistry of respiration in microorganisms
The need for energy is provided by energy metabolism processes, the essence of which is the oxidation of organic substances, accompanied by the release of energy...
Hydrocarbon-oxidizing microorganisms are promising objects of environmental biotechnology
1.3 Transformations carried out by spores of fungi and actinomycetes
Transformations carried out by disputes deserve special attention. They have a number of conveniences as technological processes. The unexpectedly high enzymatic activity demonstrated by the spores...
Bacteria are the oldest group of organisms currently existing on Earth. The first bacteria probably appeared more than 3.5 billion years ago and for almost a billion years they were the only living creatures on our planet. Since these were the first representatives of living nature, their body had a primitive structure.
Over time, their structure became more complex, but to this day bacteria are considered the most primitive single-celled organisms. It is interesting that some bacteria still retain the primitive features of their ancient ancestors. This is observed in bacteria living in hot sulfur springs and anoxic mud at the bottom of reservoirs.
Most bacteria are colorless. Only a few are purple or green. But the colonies of many bacteria have a bright color, which is caused by the release of a colored substance into the environment or pigmentation of cells.
The discoverer of the world of bacteria was Antony Leeuwenhoek, a Dutch naturalist of the 17th century, who first created a perfect magnifying microscope that magnifies objects 160-270 times.
Bacteria are classified as prokaryotes and are classified into a separate kingdom - Bacteria.
Body Shape
Bacteria are numerous and diverse organisms. They vary in shape.
| Name of the bacterium | Bacteria shape | Bacteria image |
| Cocci | Ball-shaped | |
| Bacillus |  | Rod-shaped |
| Vibrio | Comma-shaped | |
| Spirillum |  | Spiral |
| Streptococci | 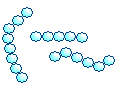 | Chain of cocci |
| Staphylococcus |  | Clusters of cocci |
| Diplococcus | Two round bacteria enclosed in one mucous capsule |
Methods of transportation
Among bacteria there are mobile and immobile forms. Motiles move due to wave-like contractions or with the help of flagella (twisted helical threads), which consist of a special protein called flagellin. There may be one or more flagella. In some bacteria they are located at one end of the cell, in others - at two or over the entire surface.

But movement is also inherent in many other bacteria that lack flagella. Thus, bacteria covered on the outside with mucus are capable of gliding movement.
Some aquatic and soil bacteria lacking flagella have gas vacuoles in the cytoplasm. There may be 40-60 vacuoles in a cell. Each of them is filled with gas (presumably nitrogen). By regulating the amount of gas in the vacuoles, aquatic bacteria can sink into the water column or rise to its surface, and soil bacteria can move in the soil capillaries.
Habitat
Due to their simplicity of organization and unpretentiousness, bacteria are widespread in nature. Bacteria are found everywhere: in a drop of even the purest spring water, in grains of soil, in the air, on rocks, in polar snow, desert sands, on the ocean floor, in oil extracted from great depths, and even in the water of hot springs with a temperature of about 80ºC. They live on plants, fruits, various animals and in humans in the intestines, oral cavity, limbs, and on the surface of the body.
Bacteria are the smallest and most numerous living creatures. Due to their small size, they easily penetrate into any cracks, crevices, or pores. Very hardy and adapted to various living conditions. They tolerate drying, extreme cold, and heating up to 90ºC without losing their viability.
There is practically no place on Earth where bacteria are not found, but in varying quantities. The living conditions of bacteria are varied. Some of them require atmospheric oxygen, others do not need it and are able to live in an oxygen-free environment.
In the air: bacteria rise to the upper atmosphere up to 30 km. and more.
There are especially many of them in the soil. 1 g of soil can contain hundreds of millions of bacteria.
In water: in the surface layers of water in open reservoirs. Beneficial aquatic bacteria mineralize organic residues.
In living organisms: pathogenic bacteria enter the body from the external environment, but only under favorable conditions cause diseases. Symbiotic live in the digestive organs, helping to break down and absorb food, and synthesize vitamins.
External structure
The bacterial cell is covered with a special dense shell - a cell wall, which performs protective and supporting functions, and also gives the bacterium a permanent, characteristic shape. The cell wall of a bacterium resembles the wall of a plant cell. It is permeable: through it, nutrients freely pass into the cell, and metabolic products exit into the environment. Often, bacteria produce an additional protective layer of mucus on top of the cell wall - a capsule. The thickness of the capsule can be many times greater than the diameter of the cell itself, but it can also be very small. The capsule is not an essential part of the cell; it is formed depending on the conditions in which the bacteria find themselves. It protects the bacteria from drying out.
On the surface of some bacteria there are long flagella (one, two or many) or short thin villi. The length of the flagella can be many times greater than the size of the body of the bacterium. Bacteria move with the help of flagella and villi.

Internal structure
Inside the bacterial cell there is dense, immobile cytoplasm. It has a layered structure, there are no vacuoles, therefore various proteins (enzymes) and reserve nutrients are located in the substance of the cytoplasm itself. Bacterial cells do not have a nucleus. A substance carrying hereditary information is concentrated in the central part of their cell. Bacteria, - nucleic acid - DNA. But this substance is not formed into a nucleus.
The internal organization of a bacterial cell is complex and has its own specific characteristics. The cytoplasm is separated from the cell wall by the cytoplasmic membrane. In the cytoplasm there is a main substance, or matrix, ribosomes and a small number of membrane structures that perform a variety of functions (analogues of mitochondria, endoplasmic reticulum, Golgi apparatus). The cytoplasm of bacterial cells often contains granules of various shapes and sizes. The granules may be composed of compounds that serve as a source of energy and carbon. Droplets of fat are also found in the bacterial cell.
In the central part of the cell, the nuclear substance is localized - DNA, which is not delimited from the cytoplasm by a membrane. This is an analogue of the nucleus - a nucleoid. The nucleoid does not have a membrane, a nucleolus, or a set of chromosomes.
Eating methods
Bacteria have different feeding methods. Among them there are autotrophs and heterotrophs. Autotrophs are organisms that are capable of independently producing organic substances for their nutrition.
Plants need nitrogen, but cannot absorb nitrogen from the air themselves. Some bacteria combine nitrogen molecules in the air with other molecules, resulting in substances that are available to plants.
These bacteria settle in the cells of young roots, which leads to the formation of thickenings on the roots, called nodules. Such nodules form on the roots of plants of the legume family and some other plants.

The roots provide carbohydrates to the bacteria, and the bacteria to the roots provide nitrogen-containing substances that can be absorbed by the plant. Their cohabitation is mutually beneficial.
Plant roots secrete a lot of organic substances (sugars, amino acids and others) that bacteria feed on. Therefore, especially many bacteria settle in the soil layer surrounding the roots. These bacteria convert dead plant debris into plant-available substances. This layer of soil is called the rhizosphere.
There are several hypotheses about the penetration of nodule bacteria into root tissue:
- through damage to epidermal and cortex tissue;
- through root hairs;
- only through the young cell membrane;
- thanks to companion bacteria producing pectinolytic enzymes;
- due to stimulation of the synthesis of B-indoleacetic acid from tryptophan, always present in plant root secretions.
The process of introduction of nodule bacteria into root tissue consists of two phases:
- infection of root hairs;
- process of nodule formation.
In most cases, the invading cell actively multiplies, forms so-called infection threads and, in the form of such threads, moves into the plant tissue. Nodule bacteria emerging from the infection thread continue to multiply in the host tissue.

Plant cells filled with rapidly multiplying cells of nodule bacteria begin to rapidly divide. The connection of a young nodule with the root of a legume plant is carried out thanks to vascular-fibrous bundles. During the period of functioning, the nodules are usually dense. By the time optimal activity occurs, the nodules acquire a pink color (thanks to the leghemoglobin pigment). Only those bacteria that contain leghemoglobin are capable of fixing nitrogen.
Nodule bacteria create tens and hundreds of kilograms of nitrogen fertilizer per hectare of soil.
Metabolism
Bacteria differ from each other in their metabolism. In some it occurs with the participation of oxygen, in others - without it.
Most bacteria feed on ready-made organic substances. Only a few of them (blue-green, or cyanobacteria) are capable of creating organic substances from inorganic ones. They played an important role in the accumulation of oxygen in the Earth's atmosphere.
Bacteria absorb substances from the outside, tear their molecules into pieces, assemble their shell from these parts and replenish their contents (this is how they grow), and throw unnecessary molecules out. The shell and membrane of the bacterium allows it to absorb only the necessary substances.

If the shell and membrane of a bacterium were completely impermeable, no substances would enter the cell. If they were permeable to all substances, the contents of the cell would mix with the medium - the solution in which the bacterium lives. To survive, bacteria need a shell that allows necessary substances to pass through, but not unnecessary substances.
The bacterium absorbs nutrients located near it. What happens next? If it can move independently (by moving a flagellum or pushing mucus back), then it moves until it finds the necessary substances.

If it cannot move, then it waits until diffusion (the ability of molecules of one substance to penetrate into the thicket of molecules of another substance) brings the necessary molecules to it.
Bacteria, together with other groups of microorganisms, perform enormous chemical work. By converting various compounds, they receive the energy and nutrients necessary for their life. Metabolic processes, methods of obtaining energy and the need for materials for building the substances of their bodies are diverse in bacteria.
Other bacteria satisfy all their needs for carbon necessary for the synthesis of organic substances in the body at the expense of inorganic compounds. They are called autotrophs. Autotrophic bacteria are capable of synthesizing organic substances from inorganic ones. Among them are:
Chemosynthesis
The use of radiant energy is the most important, but not the only way to create organic matter from carbon dioxide and water. Bacteria are known that use not sunlight as an energy source for such synthesis, but the energy of chemical bonds occurring in the cells of organisms during the oxidation of certain inorganic compounds - hydrogen sulfide, sulfur, ammonia, hydrogen, nitric acid, ferrous compounds of iron and manganese. They use the organic matter formed using this chemical energy to build the cells of their body. Therefore, this process is called chemosynthesis.
The most important group of chemosynthetic microorganisms are nitrifying bacteria. These bacteria live in the soil and oxidize ammonia formed during the decay of organic residues to nitric acid. The latter reacts with mineral compounds of the soil, turning into salts of nitric acid. This process takes place in two phases.
Iron bacteria convert ferrous iron into oxide iron. The resulting iron hydroxide settles and forms the so-called bog iron ore.
Some microorganisms exist due to the oxidation of molecular hydrogen, thereby providing an autotrophic method of nutrition.
A characteristic feature of hydrogen bacteria is the ability to switch to a heterotrophic lifestyle when provided with organic compounds and the absence of hydrogen.
Thus, chemoautotrophs are typical autotrophs, since they independently synthesize the necessary organic compounds from inorganic substances, and do not take them ready-made from other organisms, like heterotrophs. Chemoautotrophic bacteria differ from phototrophic plants in their complete independence from light as an energy source.
Bacterial photosynthesis
Some pigment-containing sulfur bacteria (purple, green), containing specific pigments - bacteriochlorophylls, are able to absorb solar energy, with the help of which hydrogen sulfide in their bodies is broken down and releases hydrogen atoms to restore the corresponding compounds. This process has much in common with photosynthesis and differs only in that in purple and green bacteria the hydrogen donor is hydrogen sulfide (occasionally carboxylic acids), and in green plants it is water. In both of them, the separation and transfer of hydrogen is carried out due to the energy of absorbed solar rays.
This bacterial photosynthesis, which occurs without the release of oxygen, is called photoreduction. Photoreduction of carbon dioxide is associated with the transfer of hydrogen not from water, but from hydrogen sulfide:
6СО 2 +12Н 2 S+hv → С6Н 12 О 6 +12S=6Н 2 О
The biological significance of chemosynthesis and bacterial photosynthesis on a planetary scale is relatively small. Only chemosynthetic bacteria play a significant role in the process of sulfur cycling in nature. Absorbed by green plants in the form of sulfuric acid salts, sulfur is reduced and becomes part of protein molecules. Further, when dead plant and animal remains are destroyed by putrefactive bacteria, sulfur is released in the form of hydrogen sulfide, which is oxidized by sulfur bacteria to free sulfur (or sulfuric acid), forming sulfites in the soil that are accessible to plants. Chemo- and photoautotrophic bacteria are essential in the nitrogen and sulfur cycle.
Sporulation
Spores form inside the bacterial cell. During the process of sporulation, the bacterial cell undergoes a number of biochemical processes. The amount of free water in it decreases and enzymatic activity decreases. This ensures the resistance of the spores to unfavorable environmental conditions (high temperature, high salt concentration, drying, etc.). Sporulation is characteristic of only a small group of bacteria.

Spores are an optional stage in the life cycle of bacteria. Sporulation begins only with a lack of nutrients or accumulation of metabolic products. Bacteria in the form of spores can remain dormant for a long time. Bacterial spores can withstand prolonged boiling and very long freezing. When favorable conditions occur, the spore germinates and becomes viable. Bacterial spores are an adaptation to survive in unfavorable conditions.
Reproduction
Bacteria reproduce by dividing one cell into two. Having reached a certain size, the bacterium divides into two identical bacteria. Then each of them begins to feed, grows, divides, and so on.

After cell elongation, a transverse septum gradually forms, and then the daughter cells separate; In many bacteria, under certain conditions, after dividing, cells remain connected in characteristic groups. In this case, depending on the direction of the division plane and the number of divisions, different shapes arise. Reproduction by budding occurs as an exception in bacteria.
Under favorable conditions, cell division in many bacteria occurs every 20-30 minutes. With such rapid reproduction, the offspring of one bacterium in 5 days is capable of forming a mass that can fill all seas and oceans. A simple calculation shows that 72 generations (720,000,000,000,000,000,000 cells) can be formed per day. If converted into weight - 4720 tons. However, this does not happen in nature, since most bacteria quickly die under the influence of sunlight, drying, lack of food, heating to 65-100ºC, as a result of struggle between species, etc.
The bacterium (1), having absorbed enough food, increases in size (2) and begins to prepare for reproduction (cell division). Its DNA (in a bacterium the DNA molecule is closed in a ring) doubles (the bacterium produces a copy of this molecule). Both DNA molecules (3,4) find themselves attached to the wall of the bacterium and, as the bacterium elongates, move apart (5,6). First the nucleotide divides, then the cytoplasm.

After the divergence of two DNA molecules, a constriction appears on the bacterium, which gradually divides the body of the bacterium into two parts, each of which contains a DNA molecule (7).
It happens (in Bacillus subtilis) that two bacteria stick together and a bridge is formed between them (1,2).

The jumper transports DNA from one bacterium to another (3). Once in one bacterium, DNA molecules intertwine, stick together in some places (4), and then exchange sections (5).
The role of bacteria in nature
Gyre
Bacteria are the most important link in the general cycle of substances in nature. Plants create complex organic substances from carbon dioxide, water and mineral salts in the soil. These substances return to the soil with dead fungi, plants and animal corpses. Bacteria break down complex substances into simple ones, which are then used by plants.
Bacteria destroy complex organic substances of dead plants and animal corpses, excretions of living organisms and various wastes. Feeding on these organic substances, saprophytic bacteria of decay turn them into humus. These are a kind of orderlies of our planet. Thus, bacteria actively participate in the cycle of substances in nature.
Soil formation
Since bacteria are distributed almost everywhere and occur in huge numbers, they largely determine various processes occurring in nature. In autumn, the leaves of trees and shrubs fall, above-ground shoots of grasses die, old branches fall off, and from time to time the trunks of old trees fall. All this gradually turns into humus. In 1 cm3. The surface layer of forest soil contains hundreds of millions of saprophytic soil bacteria of several species. These bacteria convert humus into various minerals that can be absorbed from the soil by plant roots.
Some soil bacteria are able to absorb nitrogen from the air, using it in vital processes. These nitrogen-fixing bacteria live independently or settle in the roots of legume plants. Having penetrated the roots of legumes, these bacteria cause the growth of root cells and the formation of nodules on them.
These bacteria produce nitrogen compounds that plants use. Bacteria obtain carbohydrates and mineral salts from plants. Thus, there is a close relationship between the legume plant and the nodule bacteria, which is beneficial to both one and the other organism. This phenomenon is called symbiosis.
Thanks to symbiosis with nodule bacteria, leguminous plants enrich the soil with nitrogen, helping to increase yield.
Distribution in nature
Microorganisms are ubiquitous. The only exceptions are the craters of active volcanoes and small areas at the epicenters of exploded atomic bombs. Neither the low temperatures of Antarctica, nor the boiling streams of geysers, nor saturated salt solutions in salt pools, nor the strong insolation of mountain peaks, nor the harsh irradiation of nuclear reactors interfere with the existence and development of microflora. All living beings constantly interact with microorganisms, often being not only their repositories, but also their distributors. Microorganisms are natives of our planet, actively exploring the most incredible natural substrates.
Soil microflora
The number of bacteria in the soil is extremely large - hundreds of millions and billions of individuals per gram. There are much more of them in soil than in water and air. The total number of bacteria in soils changes. The number of bacteria depends on the type of soil, their condition, and the depth of the layers.
On the surface of soil particles, microorganisms are located in small microcolonies (20-100 cells each). They often develop in the thickness of clots of organic matter, on living and dying plant roots, in thin capillaries and inside lumps.
The soil microflora is very diverse. Here there are different physiological groups of bacteria: putrefaction bacteria, nitrifying bacteria, nitrogen-fixing bacteria, sulfur bacteria, etc. among them there are aerobes and anaerobes, spore and non-spore forms. Microflora is one of the factors in soil formation.
The area of development of microorganisms in the soil is the zone adjacent to the roots of living plants. It is called the rhizosphere, and the totality of microorganisms contained in it is called the rhizosphere microflora.
Microflora of reservoirs
Water is a natural environment where microorganisms develop in large numbers. The bulk of them enters the water from the soil. A factor that determines the number of bacteria in water and the presence of nutrients in it. The cleanest waters are from artesian wells and springs. Open reservoirs and rivers are very rich in bacteria. The largest number of bacteria is found in the surface layers of water, closer to the shore. As you move away from the shore and increase in depth, the number of bacteria decreases.
Clean water contains 100-200 bacteria per ml, and polluted water contains 100-300 thousand or more. There are many bacteria in the bottom sludge, especially in the surface layer, where the bacteria form a film. This film contains a lot of sulfur and iron bacteria, which oxidize hydrogen sulfide to sulfuric acid and thereby prevent fish from dying. There are more spore-bearing forms in silt, while non-spore-bearing forms predominate in water.
In terms of species composition, the microflora of water is similar to the microflora of soil, but there are also specific forms. By destroying various waste that gets into the water, microorganisms gradually carry out the so-called biological purification of water.
Air microflora
The microflora of the air is less numerous than the microflora of soil and water. Bacteria rise into the air with dust, can remain there for some time, and then settle on the surface of the earth and die from lack of nutrition or under the influence of ultraviolet rays. The number of microorganisms in the air depends on the geographical zone, terrain, time of year, dust pollution, etc. each speck of dust is a carrier of microorganisms. Most bacteria are in the air above industrial enterprises. The air in rural areas is cleaner. The cleanest air is over forests, mountains, and snowy areas. The upper layers of air contain fewer microbes. The air microflora contains many pigmented and spore-bearing bacteria, which are more resistant than others to ultraviolet rays.
Microflora of the human body
The human body, even a completely healthy one, is always a carrier of microflora. When the human body comes into contact with air and soil, various microorganisms, including pathogenic ones (tetanus bacilli, gas gangrene, etc.), settle on clothing and skin. The most frequently exposed parts of the human body are contaminated. E. coli and staphylococci are found on the hands. There are over 100 types of microbes in the oral cavity. The mouth, with its temperature, humidity, and nutrient residues, is an excellent environment for the development of microorganisms.
The stomach has an acidic reaction, so the majority of microorganisms in it die. Starting from the small intestine, the reaction becomes alkaline, i.e. favorable for microbes. The microflora in the large intestines is very diverse. Each adult excretes about 18 billion bacteria daily in excrement, i.e. more individuals than people on the globe.
Internal organs that are not connected to the external environment (brain, heart, liver, bladder, etc.) are usually free of microbes. Microbes enter these organs only during illness.
Bacteria in the cycle of substances
Microorganisms in general and bacteria in particular play a large role in the biologically important cycles of substances on Earth, carrying out chemical transformations that are completely inaccessible to either plants or animals. Different stages of the cycle of elements are carried out by organisms of different types. The existence of each individual group of organisms depends on the chemical transformation of elements carried out by other groups.
Nitrogen cycle
The cyclic transformation of nitrogenous compounds plays a primary role in supplying the necessary forms of nitrogen to organisms of the biosphere with different nutritional needs. Over 90% of total nitrogen fixation is due to the metabolic activity of certain bacteria.
Carbon cycle
The biological transformation of organic carbon into carbon dioxide, accompanied by the reduction of molecular oxygen, requires the joint metabolic activity of various microorganisms. Many aerobic bacteria carry out complete oxidation of organic substances. Under aerobic conditions, organic compounds are initially broken down by fermentation, and the organic end products of fermentation are further oxidized by anaerobic respiration if inorganic hydrogen acceptors (nitrate, sulfate, or CO 2 ) are present.
Sulfur cycle
Sulfur is available to living organisms mainly in the form of soluble sulfates or reduced organic sulfur compounds.
Iron cycle
Some freshwater bodies contain high concentrations of reduced iron salts. In such places, a specific bacterial microflora develops - iron bacteria, which oxidize reduced iron. They participate in the formation of bog iron ores and water sources rich in iron salts.
Bacteria are the most ancient organisms, appearing about 3.5 billion years ago in the Archean. For about 2.5 billion years they dominated the Earth, forming the biosphere, and participated in the formation of the oxygen atmosphere.
Bacteria are one of the most simply structured living organisms (except viruses). They are believed to be the first organisms to appear on Earth.
Bacteria are prokaryotic microorganisms with a cellular structure. Their sizes range from 0.1 to 30 microns. Germs are extremely common. They live in soil, air, water, snow and even hot springs, on the body of animals, as well as inside living organisms, including the human body.
The distribution of bacteria into species takes into account several criteria, among which the shape of microorganisms and their spatial distribution are most often taken into account. So, according to the shape of the cells, bacteria are divided into:
Coca - micro-, diplo-, strepto-, staphylococci, as well as sarcina;
Rod-shaped - monobacteria, diplobacteria and streptobacteria;
The convoluted species are vibrios and spirochetes.
Bergey's determinant systematizes all known bacteria according to the most widely used principles of bacterial identification in practical bacteriology, based on differences in the structure of the cell wall and the relationship to Gram staining. The description of bacteria is given by groups (sections), which include families, genera and species; in some cases, groups include classes and orders. Bacteria pathogenic to humans are included in a small number of groups.
The key identifies four main categories of bacteria -
Gracillicutes [from lat. gracilis, graceful, thin, + cutis, skin] - species with a thin cell wall, staining gram negative;
firmicutes [from lat. flrmus, strong, + cutis, skin] - bacteria with a thick cell wall, staining gram-positive;
Tenericutes [from Lat. tener, tender, + cutis, skin] - bacteria lacking a cell wall(mycoplasmas and other representatives of the class Mollicutes)
Mendosicutes [from Lat. mendosus, irregular, + cutis, skin] - archaebacteria (methane- and sulfate-reducing, halophilic, thermophilic and archaebacteria lacking a cell wall).
Group 2 of the Bergey determinant. Aerobic and microaerophilic motile convoluted and curved gram-negative bacteria. Species pathogenic to humans are included in the genera Campylobacter and Helicobacters Spirillum.
Group 3 of the Bergey determinant. Non-motile (rarely motile) gram-negative bacteria. Does not contain pathogenic species.
Group 4 of the Bergey determinant. Gram-negative aerobic and microaerophilic rods and cocci. Species pathogenic to humans are included in the families Legionellaceae, Neisseriaceae and Pseudomonada-ceae; the group also includes pathogenic and opportunistic bacteria of the genera Acinetobacter, Afipia, Alcaligenes, Bordetella, Brucella, Flavobacterium, Francisella, Kingella and Moraxella.
Group 5 of the Bergey determinant. Facultatively anaerobic gram-negative rods. The group is formed by three families - Enterobacteriaceae, Vibrionaceae and Pasteurellaceae, each of which includes pathogenic species, as well as pathogenic and opportunistic bacteria of the genera Calymmobaterium, Cardiobacterium, Eikenetta, Gardnerella and Streptobacillus.
Group 6 of the Bergey determinant. Gram-negative anaerobic straight, curved and spiral bacteria. Pathogenic and opportunistic species are included in the genera Bacteroides, Fusobacterium, Porphoromonas and Prevotelta.
Group 7 of the Bergey determinant. Bacteria performing dissimilatory reduction of sulfate or sulfur Does not include pathogenic species.
Group 8 of the Bergey determinant. Anaerobic gram-negative cocci. Includes opportunistic bacteria of the genus Veillonella.
Group 9 of the Bergey determinant. Rickettsia and chlamydia. Three families - Rickettsiaceae, Bartonellaceae and Chlamydiaceae, each of which contains species pathogenic to humans.
Groups 10 and 11 of Bergey's determinant include anoxy- and oxygenic phototrophic bacteria that are not pathogenic to humans.
Group 12 of the Bergey determinant. Aerobic chemolithotrophic bacteria and related organisms. Combines iron-sulphur- and manganese-oxidizing and nitrifying bacteria that do not cause damage to humans.
Groups 13 and 14 of Bergey's determinant include budding and/or protruding bacteria and sheath-forming bacteria. They are represented by free-living species that are not pathogenic to humans;
Groups 15 and 16 of Bergey's determinant combine gliding bacteria that do not form fruiting bodies and those that form them. The groups do not include species pathogenic to humans.
Group 17 of the Bergey determinant. Gram-positive cocci. Includes opportunistic species of the genera Enterococcus Leuconostoc, Peptococcus, Peptostreptococcus, Sarcina, Staphylococcus, Stomatococcus, Streptococcus.
Group 18 of the Bergey determinant. Spore-forming gram-positive rods and cocci. Includes pathogenic and opportunistic bacilli of the genera Clostridium and Bacillus.
Group 19 of the Bergey determinant. Spore-forming gram-positive rods of regular shape. Including opportunistic species of the genera Erysipelothrix and Listeria.
Group 20 of the Bergey determinant. Spore-forming gram-positive rods of irregular shape. The group includes pathogenic and opportunistic species of the genera Actinomyces, Corynebacterium Gardnerella, Mobiluncus, etc.
Group 21 of the Bergey determinant. Mycobacteria. Includes the only genus Mycobacterium, combining pathogenic and opportunistic species.
Groups 22-29. Actinomycetes. Among numerous species, only nocardioform actinomycetes (group 22) of the genera Gordona, Nocardia, Rhodococcus, Tsukamurella, Jonesia, Oerskovi and Terrabacter are capable of causing lesions in humans.
Group 30 of the Bergey determinant. Mycoplasmas. Species included in the genus Acholeplasma, Mycoplasma and Ureaplasma are pathogenic to humans.
The remaining groups of Bergey's determinant - methanogenic bacteria (31), sulfate-reducing bacteria (32 extremely halophilic aerobic archaebacteria (33), archaebacteria lacking cell walls (34), extreme thermophiles and hyperthermophiles that metabolize sulfur (35) - do not contain species pathogenic to humans.
2.1. Systematics and nomenclature of microbes
The microbial world can be divided into cellular and non-cellular forms. Cellular forms of microbes are represented by bacteria, fungi and protozoa. They can be called microorganisms. Non-cellular forms are represented by viruses, viroids and prions.
The new classification of cellular microbes includes the following taxonomic units: domains, kingdoms, types, classes, orders, families, genera, species. The classification of microorganisms is based on their genetic relationship, as well as morphological, physiological, antigenic and molecular biological properties.
Viruses are often considered not as organisms, but as autonomous genetic structures, so they will be considered separately.
The cellular forms of microbes are divided into three domains. Domains Bacteria And Archaebacteria include microbes with a prokaryotic type of cell structure. Domain representatives Eukarya are eukaryotes. It consists of 4 kingdoms:
Mushroom kingdoms (Fungi, Eumycota);
kingdoms of protozoa (Protozoa);
kingdoms Chromista(chrome plates);
Microbes with unspecified taxonomic position (Microspora, microsporidia).
Differences in the organization of prokaryotic and eukaryotic cells are presented in table. 2.1.
Table 2.1. Signs of a prokaryotic and eukaryotic cell
2.2. Classification and morphology of bacteria
The term "bacteria" comes from the word bacteria, what does stick mean? Bacteria are prokaryotes. They are divided into two domains: Bacteria And Archaebacteria. Bacteria included in the domain Archaebacteria, represent one of the oldest forms of life. They have structural features of the cell wall (they lack peptidoglycan) and ribosomal RNA. There are no pathogens of infectious diseases among them.
Within a domain, bacteria are divided into the following taxonomic categories: class, phylum, order, family, genus, species. One of the main taxonomic categories is species. A species is a collection of individuals having the same origin and genotype, united by similar properties that distinguish them from other representatives of the genus. The species name corresponds to binary nomenclature, i.e. consists of two words. For example, the causative agent of diphtheria is written as Corynebacterium diphtheriae. The first word is the name of the genus and is written with a capital letter, the second word denotes the species and is written with a lowercase letter.
When a species is mentioned again, the generic name is abbreviated to its initial letter, e.g. C. diphtheriae.
A set of homogeneous microorganisms isolated on a nutrient medium, characterized by similar morphological, tinctorial (relation to dyes), cultural, biochemical and antigenic properties is called pure culture. A pure culture of microorganisms isolated from a specific source and different from other members of the species is called strain. Close to the concept of “strain” is the concept of “clone”. A clone is a collection of descendants grown from a single microbial cell.
To designate certain sets of microorganisms that differ in certain properties, the suffix “var” (variety) is used, therefore microorganisms, depending on the nature of the differences, are designated as morphovars (difference in morphology), resistant products (difference in resistance, for example, to antibiotics), serovars (difference in antigens), phagovars (difference in sensitivity to bacteriophages), biovars (difference in biological properties), chemovars (difference in biochemical properties), etc.
Previously, the basis for the classification of bacteria was the structural feature of the cell wall. The division of bacteria according to the structural features of the cell wall is associated with the possible variability of their coloring in one color or another using the Gram method. According to this method, proposed in 1884 by the Danish scientist H. Gram, depending on the staining results, bacteria are divided into gram-positive, stained blue-violet, and gram-negative, stained red.
Currently, the classification is based on the degree of genetic relatedness, based on studying the structure of the genome of ribosomal RNA (rRNA) (see Chapter 5), determining the percentage of guanine cytosine pairs (GC pairs) in the genome, constructing a restriction map of the genome, and studying the degree of hybridization. Phenotypic indicators are also taken into account: attitude to Gram staining, morphological, cultural and biochemical properties, antigenic structure.
Domain Bacteria includes 23 types, of which the following are of medical importance.
Most gram-negative bacteria are grouped into the phylum Proteobacteria(named after the Greek god Proteus, capable of taking on different forms). Type Proteobacteria divided into 5 classes:
Class Alphaproteobacteria(birth Rickettsia, Orientia, Erlichia, Bartonella, Brucella);
Class Betaproteobacteria(birth Bordetella, Burholderia, Neisseria, Spirillum);
Class Gammaproteobacteria(family representatives Enterobacteriaceae childbirth Francisella, Legionella, Coxiella, Pseudomonas, Vibrio);
Class Deltaproteobacteria(genus Bilophila);
Class Epsilonproteobacteria(birth Campilobacter, Helicobacter). Gram-negative bacteria also include the following types:
type Chlamydiae(birth Chlamydia, Chlamydophila), type Spirochaetes(birth Spirocheta, Borrelia, Treponema, Leptospira); type Bacteroides(birth Bacteroides, Prevotella, Porphyromonas).
Gram-positive bacteria come in the following types:
Type Firmicutes includes class Clostridium(birth Clostridium, Peptococcus), Class Bacilli (Listeria, Staphylococcus, Lactobacillus, Streptococcus) and class Mollicutes(birth Mycoplasma, Ureaplasma), which are bacteria that do not have a cell wall;
type Actinobacteria(birth Actinomyces, Micrococcus, Corynebacterium, Mycobacterium, Gardnerella, Bifidobacterium, Propionibacterium, Mobiluncus).
2.2.1. Morphological forms of bacteria
There are several main forms of bacteria: coccoid, rod-shaped, convoluted and branching (Fig. 2.1).
Spherical forms, or cocci- spherical bacteria 0.5-1 microns in size, which, according to their relative position, are divided into micrococci, diplococci, streptococci, tetracocci, sarcina and staphylococci.
Micrococci (from Greek. micros- small) - separately located cells.
Diplococci (from Greek. diploos- double), or paired cocci, are located in pairs (pneumococcus, gonococcus, meningococcus), since the cells do not separate after division. Pneumococcus (the causative agent of pneumonia) has a lanceolate shape on opposite sides, and gonococcus (the causative agent of gonorrhea) and meningococcus (the causative agent of
 Rice. 2.1. Shapes of bacteria
Rice. 2.1. Shapes of bacteria
agent of epidemic meningitis) have the shape of coffee beans, with their concave surface facing each other.
Streptococci (from Greek. streptos- chain) - cells of a round or elongated shape, forming a chain due to cell division in the same plane and maintaining the connection between them at the site of division.
Sarcins (from lat. sarcina- bunch, bale) are arranged in the form of packets of 8 cocci or more, since they are formed during cell division in three mutually perpendicular planes.
Staphylococcus (from Greek. staphyle- grape bunch) - cocci located in the form of a bunch of grapes as a result of division in different planes.
Rod-shaped bacteria differ in size, shape of cell ends and relative position of cells. Cell length is 1-10 µm, thickness 0.5-2 µm. Sticks can be right
(Escherichia coli, etc.) and irregular club-shaped (Corynebacteria, etc.) shape. The smallest rod-shaped bacteria include rickettsia.
The ends of the rods can be cut off (anthrax bacillus), rounded (Escherichia coli), pointed (fusobacteria) or in the form of a thickening. In the latter case, the rod looks like a club (Corynebacterium diphtheria).
The slightly curved rods are called vibrios (Vibrio cholerae). Most rod-shaped bacteria are arranged randomly because the cells move apart after dividing. If, after division, the cells remain connected by common fragments of the cell wall and do not diverge, then they are located at an angle to each other (Corynebacterium diphtheria) or form a chain (anthrax bacillus).
Twisted Shapes- spiral-shaped bacteria, which come in two types: spirilla and spirochetes. Spirilla have the appearance of corkscrew-shaped convoluted cells with large curls. Pathogenic spirilla include the causative agent of sodoku (rat bite disease), as well as campylobacter and helicobacter, which have curves reminiscent of the wings of a flying seagull. Spirochetes are thin, long, convoluted bacteria that differ from spirilla in their smaller curls and pattern of movement. The peculiarities of their structure are described below.
Branching - rod-shaped bacteria, which can have Y-shaped branches found in bifidobacteria, can also be presented in the form of filamentous branched cells that can intertwine to form mycelium, as observed in actinomycetes.
2.2.2. Bacterial cell structure
The structure of bacteria has been well studied using electron microscopy of whole cells and their ultrathin sections, as well as other methods. The bacterial cell is surrounded by a membrane consisting of a cell wall and a cytoplasmic membrane. Under the shell there is protoplasm, consisting of cytoplasm with inclusions and a hereditary apparatus - an analogue of the nucleus, called the nucleoid (Fig. 2.2). There are additional structures: capsule, microcapsule, mucus, flagella, pili. Some bacteria are capable of forming spores under unfavorable conditions.
 Rice. 2.2. Structure of a bacterial cell: 1 - capsule; 2 - cell wall; 3 - cytoplasmic membrane; 4 - mesosomes; 5 - nucleoid; 6 - plasmid; 7 - ribosomes; 8 - inclusions; 9 - flagellum; 10 - pili (villi)
Rice. 2.2. Structure of a bacterial cell: 1 - capsule; 2 - cell wall; 3 - cytoplasmic membrane; 4 - mesosomes; 5 - nucleoid; 6 - plasmid; 7 - ribosomes; 8 - inclusions; 9 - flagellum; 10 - pili (villi)
Cell wall- a strong, elastic structure that gives the bacterium a certain shape and, together with the underlying cytoplasmic membrane, restrains high osmotic pressure in the bacterial cell. It is involved in the process of cell division and transport of metabolites, has receptors for bacteriophages, bacteriocins and various substances. The thickest cell wall is found in gram-positive bacteria (Fig. 2.3). So, if the thickness of the cell wall of gram-negative bacteria is about 15-20 nm, then in gram-positive bacteria it can reach 50 nm or more.
The basis of the bacterial cell wall is peptidoglycan. Peptidoglycan is a polymer. It is represented by parallel polysaccharide glycan chains consisting of repeating N-acetylglucosamine and N-acetylmuramic acid residues connected by a glycosidic bond. This bond is broken by lysozyme, which is an acetylmuramidase.
A tetrapeptide is attached to N-acetylmuramic acid by covalent bonds. The tetrapeptide consists of L-alanine, which is linked to N-acetylmuramic acid; D-glutamine, which in gram-positive bacteria is combined with L-lysine, and in gram-tri-
 Rice. 2.3. Scheme of the architecture of the bacterial cell wall
Rice. 2.3. Scheme of the architecture of the bacterial cell wall
beneficial bacteria - with diaminopimelic acid (DAP), which is a precursor of lysine in the process of bacterial biosynthesis of amino acids and is a unique compound present only in bacteria; The 4th amino acid is D-alanine (Fig. 2.4).
The cell wall of gram-positive bacteria contains small amounts of polysaccharides, lipids and proteins. The main component of the cell wall of these bacteria is multilayer peptidoglycan (murein, mucopeptide), accounting for 40-90% of the mass of the cell wall. Tetrapeptides of different layers of peptidoglycan in gram-positive bacteria are connected to each other by polypeptide chains of 5 glycine residues (pentaglycine), which gives the peptidoglycan a rigid geometric structure (Fig. 2.4, b). Covalently linked to the peptidoglycan of the cell wall of gram-positive bacteria teichoic acids(from Greek tekhos- wall), the molecules of which are chains of 8-50 glycerol and ribitol residues connected by phosphate bridges. The shape and strength of bacteria is given by the rigid fibrous structure of the multilayer peptidoglycan, with cross-links of peptides.
 Rice. 2.4. Structure of peptidoglycan: a - gram-negative bacteria; b - gram-positive bacteria
Rice. 2.4. Structure of peptidoglycan: a - gram-negative bacteria; b - gram-positive bacteria
The ability of Gram-positive bacteria to retain gentian violet in combination with iodine when stained using Gram stain (blue-violet color of bacteria) is associated with the property of multilayer peptidoglycan to interact with the dye. In addition, subsequent treatment of a bacterial smear with alcohol causes a narrowing of the pores in the peptidoglycan and thereby retains the dye in the cell wall.
Gram-negative bacteria lose the dye after exposure to alcohol, which is due to a smaller amount of peptidoglycan (5-10% of the cell wall mass); they are discolored with alcohol, and when treated with fuchsin or safranin they become red. This is due to the structural features of the cell wall. Peptidoglycan in the cell wall of gram-negative bacteria is represented by 1-2 layers. The tetrapeptides of the layers are connected to each other by a direct peptide bond between the amino group of DAP of one tetrapeptide and the carboxyl group of D-alanine of the tetrapeptide of another layer (Fig. 2.4, a). Outside the peptidoglycan there is a layer lipoprotein, connected to peptidoglycan through DAP. Followed by outer membrane cell wall.
Outer membrane is a mosaic structure composed of lipopolysaccharides (LPS), phospholipids and proteins. Its inner layer is represented by phospholipids, and the outer layer contains LPS (Fig. 2.5). Thus, the outer mem-
 Rice. 2.5. Lipopolysaccharide structure
Rice. 2.5. Lipopolysaccharide structure
the brane is asymmetric. The outer membrane LPS consists of three fragments:
Lipid A has a conservative structure, almost the same in gram-negative bacteria. Lipid A consists of phosphorylated glucosamine disaccharide units to which long chains of fatty acids are attached (see Fig. 2.5);
Core, or core, crustal part (from lat. core- core), relatively conservative oligosaccharide structure;
A highly variable O-specific polysaccharide chain formed by repeating identical oligosaccharide sequences.
LPS is anchored in the outer membrane by lipid A, which causes LPS toxicity and is therefore identified with endotoxin. The destruction of bacteria by antibiotics leads to the release of large amounts of endotoxin, which can cause endotoxic shock in the patient. The core, or core part, of LPS extends from lipid A. The most constant part of the LPS core is ketodeoxyoctonic acid. O-specific polysaccharide chain extending from the core of the LPS molecule,
consisting of repeating oligosaccharide units, determines the serogroup, serovar (a type of bacteria detected using immune serum) of a particular strain of bacteria. Thus, the concept of LPS is associated with the concept of O-antigen, by which bacteria can be differentiated. Genetic changes can lead to defects, shortening of bacterial LPS and, as a result, the appearance of rough colonies of R-forms that lose O-antigen specificity.
Not all gram-negative bacteria have a complete O-specific polysaccharide chain, consisting of repeating oligosaccharide units. In particular, bacteria of the genus Neisseria have a short glycolipid called lipooligosaccharide (LOS). It is comparable to the R form, which has lost O-antigen specificity, observed in mutant rough strains E. coli. The structure of VOC resembles the structure of the glycosphingolipid of the human cytoplasmic membrane, so VOC mimics the microbe, allowing it to evade the host's immune response.
The matrix proteins of the outer membrane permeate it in such a way that protein molecules called porinami, border hydrophilic pores through which water and small hydrophilic molecules with a relative mass of up to 700 D pass.
Between the outer and cytoplasmic membrane is periplasmic space, or periplasm containing enzymes (proteases, lipases, phosphatases, nucleases, β-lactamases), as well as components of transport systems.
When the synthesis of the bacterial cell wall is disrupted under the influence of lysozyme, penicillin, protective factors of the body and other compounds, cells with an altered (often spherical) shape are formed: protoplasts- bacteria completely lacking a cell wall; spheroplasts- bacteria with a partially preserved cell wall. After removal of the cell wall inhibitor, such altered bacteria can reverse, i.e. acquire a full cell wall and restore its original shape.
Bacteria of the spheroid or protoplast type, which have lost the ability to synthesize peptidoglycan under the influence of antibiotics or other factors and are able to reproduce, are called L-shapes(from the name of the D. Lister Institute, where they first
have been studied). L-forms can also arise as a result of mutations. They are osmotically sensitive, spherical, flask-shaped cells of various sizes, including those passing through bacterial filters. Some L-forms (unstable), when the factor that led to changes in bacteria is removed, can reverse, returning to the original bacterial cell. L-forms can be produced by many pathogens of infectious diseases.
Cytoplasmic membrane in electron microscopy of ultrathin sections, it is a three-layer membrane (2 dark layers, each 2.5 nm thick, separated by a light intermediate layer). In structure, it is similar to the plasmalemma of animal cells and consists of a double layer of lipids, mainly phospholipids, with embedded surface and integral proteins that seem to penetrate through the structure of the membrane. Some of them are permeases involved in the transport of substances. Unlike eukaryotic cells, the cytoplasmic membrane of a bacterial cell lacks sterols (with the exception of mycoplasmas).
The cytoplasmic membrane is a dynamic structure with mobile components, so it is thought of as a mobile fluid structure. It surrounds the outer part of the cytoplasm of bacteria and is involved in the regulation of osmotic pressure, transport of substances and energy metabolism of the cell (due to enzymes of the electron transport chain, adenosine triphosphatase - ATPase, etc.). With excessive growth (compared to the growth of the cell wall), the cytoplasmic membrane forms invaginates - invaginations in the form of complexly twisted membrane structures, called mesosomes. Less complexly twisted structures are called intracytoplasmic membranes. The role of mesosomes and intracytoplasmic membranes is not fully understood. It is even suggested that they are an artifact that occurs after preparing (fixing) a specimen for electron microscopy. Nevertheless, it is believed that derivatives of the cytoplasmic membrane participate in cell division, providing energy for the synthesis of the cell wall, and take part in the secretion of substances, sporulation, i.e. in processes with high energy consumption. Cytoplasm occupies the main volume of bacteria
cell and consists of soluble proteins, ribonucleic acids, inclusions and numerous small granules - ribosomes, responsible for the synthesis (translation) of proteins.
Ribosomes bacteria have a size of about 20 nm and a sedimentation coefficient of 70S, in contrast to the 80S ribosomes characteristic of eukaryotic cells. Therefore, some antibiotics, by binding to bacterial ribosomes, inhibit bacterial protein synthesis without affecting protein synthesis in eukaryotic cells. Bacterial ribosomes can dissociate into two subunits: 50S and 30S. rRNA is a conserved element of bacteria (“molecular clock” of evolution). 16S rRNA is part of the small ribosomal subunit, and 23S rRNA is part of the large ribosomal subunit. The study of 16S rRNA is the basis of gene systematics, allowing one to assess the degree of relatedness of organisms.
The cytoplasm contains various inclusions in the form of glycogen granules, polysaccharides, β-hydroxybutyric acid and polyphosphates (volutin). They accumulate when there is an excess of nutrients in the environment and act as reserve substances for nutrition and energy needs.
Volyutin has an affinity for basic dyes and is easily detected using special staining methods (for example, according to Neisser) in the form of metachromatic granules. With toluidine blue or methylene blue, volutin is stained red-violet, and the cytoplasm of the bacterium is stained blue. The characteristic arrangement of volutin granules is revealed in the diphtheria bacillus in the form of intensely stained cell poles. The metachromatic coloration of volutin is associated with a high content of polymerized inorganic polyphosphate. Under electron microscopy, they look like electron-dense granules 0.1-1 microns in size.
Nucleoid- equivalent to the nucleus in bacteria. It is located in the central zone of bacteria in the form of double-stranded DNA, tightly packed like a ball. The nucleoid of bacteria, unlike eukaryotes, does not have a nuclear envelope, nucleolus and basic proteins (histones). Most bacteria contain one chromosome, represented by a DNA molecule closed in a ring. But some bacteria have two ring-shaped chromosomes (V. cholerae) and linear chromosomes (see section 5.1.1). The nucleoid is revealed in a light microscope after staining with DNA-specific stains
methods: according to Feulgen or according to Romanovsky-Giemsa. In electron diffraction patterns of ultrathin sections of bacteria, the nucleoid appears as light zones with fibrillar, thread-like structures of DNA bound in certain areas to the cytoplasmic membrane or mesosome involved in chromosome replication.
In addition to the nucleoid, the bacterial cell contains extrachromosomal heredity factors - plasmids (see section 5.1.2), which are covalently closed rings of DNA.
Capsule, microcapsule, mucus.Capsule - a mucous structure more than 0.2 microns thick, firmly associated with the bacterial cell wall and having clearly defined external boundaries. The capsule is visible in imprint smears from pathological material. In pure bacterial cultures, the capsule is formed less frequently. It is detected using special methods of staining a smear according to Burri-Gins, which creates a negative contrast of the substances of the capsule: ink creates a dark background around the capsule. The capsule consists of polysaccharides (exopolysaccharides), sometimes of polypeptides, for example, in the anthrax bacillus it consists of polymers of D-glutamic acid. The capsule is hydrophilic and contains a large amount of water. It prevents the phagocytosis of bacteria. The capsule is antigenic: antibodies to the capsule cause its enlargement (capsule swelling reaction).
Many bacteria form microcapsule- mucous formation less than 0.2 microns thick, detectable only by electron microscopy.
It should be distinguished from a capsule slime - mucoid exopolysaccharides that do not have clear external boundaries. Mucus is soluble in water.
Mucoid exopolysaccharides are characteristic of mucoid strains of Pseudomonas aeruginosa, often found in the sputum of patients with cystic fibrosis. Bacterial exopolysaccharides are involved in adhesion (sticking to substrates); they are also called glycocalyx.
The capsule and mucus protect bacteria from damage and drying out, since, being hydrophilic, they bind water well and prevent the action of the protective factors of the macroorganism and bacteriophages.
Flagella bacteria determine the mobility of the bacterial cell. Flagella are thin filaments that take on
They originate from the cytoplasmic membrane and are longer than the cell itself. The thickness of the flagella is 12-20 nm, length 3-15 µm. They consist of three parts: a spiral filament, a hook and a basal body containing a rod with special discs (one pair of discs in gram-positive bacteria and two pairs in gram-negative bacteria). Flagella are attached to the cytoplasmic membrane and cell wall by discs. This creates the effect of an electric motor with a rod - a rotor - rotating the flagellum. The proton potential difference on the cytoplasmic membrane is used as an energy source. The rotation mechanism is provided by proton ATP synthetase. The rotation speed of the flagellum can reach 100 rps. If a bacterium has several flagella, they begin to rotate synchronously, intertwining into a single bundle, forming a kind of propeller.
Flagella are made of a protein called flagellin. (flagellum- flagellum), which is an antigen - the so-called H-antigen. Flagellin subunits are twisted in a spiral.
The number of flagella in different species of bacteria varies from one (monotrichus) in Vibrio cholerae to tens and hundreds extending along the perimeter of the bacterium (peritrichus), in Escherichia coli, Proteus, etc. Lophotrichs have a bundle of flagella at one end of the cell. Amphitrichy has one flagellum or a bundle of flagella at opposite ends of the cell.
Flagella are detected using electron microscopy of preparations coated with heavy metals, or in a light microscope after treatment with special methods based on etching and adsorption of various substances leading to an increase in the thickness of the flagella (for example, after silvering).
Villi, or pili (fimbriae)- thread-like formations, thinner and shorter (3-10 nm * 0.3-10 µm) than flagella. The pili extend from the cell surface and are composed of the protein pilin. Several types of pili are known. General type pili are responsible for attachment to the substrate, nutrition, and water-salt metabolism. They are numerous - several hundred per cell. Sex pili (1-3 per cell) create contact between cells, transferring genetic information between them by conjugation (see Chapter 5). Of particular interest are type IV pili, in which the ends are hydrophobic, as a result of which they curl; these pili are also called curls. Location
They are located at the poles of the cell. These pili are found in pathogenic bacteria. They have antigenic properties, bring bacteria into contact with the host cell, and participate in the formation of biofilm (see Chapter 3). Many pili are receptors for bacteriophages.
Disputes - a peculiar form of resting bacteria with a gram-positive type of cell wall structure. Spore-forming bacteria of the genus Bacillus, in which the size of the spore does not exceed the diameter of the cell are called bacilli. Spore-forming bacteria in which the size of the spore exceeds the diameter of the cell, which is why they take the shape of a spindle, are called clostridia, for example bacteria of the genus Clostridium(from lat. Clostridium- spindle). The spores are acid-resistant, therefore they are stained red using the Aujeszky method or the Ziehl-Neelsen method, and the vegetative cell is stained blue.
Sporulation, the shape and location of spores in a cell (vegetative) are a species property of bacteria, which allows them to be distinguished from each other. The shape of the spores can be oval or spherical, the location in the cell is terminal, i.e. at the end of the stick (in the causative agent of tetanus), subterminal - closer to the end of the stick (in the causative agents of botulism, gas gangrene) and central (in the anthrax bacillus).
The process of sporulation (sporulation) goes through a number of stages, during which part of the cytoplasm and chromosome of the bacterial vegetative cell are separated, surrounded by an ingrowing cytoplasmic membrane - a prospore is formed.
The prospore protoplast contains a nucleoid, a protein synthesizing system, and an energy production system based on glycolysis. Cytochromes are absent even in aerobes. Does not contain ATP, energy for germination is stored in the form of 3-glycerol phosphate.
The prospore is surrounded by two cytoplasmic membranes. The layer surrounding the inner membrane of the spore is called wall of spores, it consists of peptidoglycan and is the main source of cell wall during spore germination.
Between the outer membrane and the spore wall, a thick layer is formed consisting of peptidoglycan, which has many cross-links - cortex.
Located outside the outer cytoplasmic membrane spore shell, consisting of keratin-like proteins, co-
holding multiple intramolecular disulfide bonds. This shell provides resistance to chemical agents. The spores of some bacteria have an additional covering - exosporium lipoprotein nature. In this way, a multilayer, poorly permeable shell is formed.
Sporulation is accompanied by intensive consumption by the prospore and then by the developing spore shell of dipicolinic acid and calcium ions. The spore acquires heat resistance, which is associated with the presence of calcium dipicolinate in it.
The spore can persist for a long time due to the presence of a multilayer shell, calcium dipicolinate, low water content and sluggish metabolic processes. In soil, for example, the pathogens of anthrax and tetanus can persist for decades.
Under favorable conditions, spores germinate, going through three successive stages: activation, initiation, growth. In this case, one bacterium is formed from one spore. Activation is readiness for germination. At a temperature of 60-80 °C, the spore is activated for germination. Germination initiation lasts several minutes. The outgrowth stage is characterized by rapid growth, accompanied by the destruction of the shell and the emergence of a seedling.
2.2.3. Structural features of spirochetes, rickettsia, chlamydia, actinomycetes and mycoplasmas
Spirochetes- thin long convoluted bacteria. They consist of an outer membranous cell wall that surrounds a cytoplasmic cylinder. On top of the outer membrane there is a transparent cover of glycosaminoglycan nature. Under the outer membrane of the cell wall are fibrils that twist around the cytoplasmic cylinder, giving the bacteria a helical shape. The fibrils are attached to the ends of the cell and directed towards each other. The number and arrangement of fibrils varies among species. Fibrils are involved in the movement of spirochetes, giving the cells rotational, bending and translational motion. In this case, spirochetes form loops, curls, and bends, which are called secondary curls. Spirochetes do not perceive dyes well. They are usually painted according to Romanovsky-Giemsa or silver plated. Live
The form of a spirochete is examined using phase-contrast or dark-field microscopy.
Spirochetes are represented by three genera that are pathogenic to humans: Treponema, Borrelia, Leptospira.
Treponema(genus Treponema) have the appearance of thin, corkscrew-twisted threads with 8-12 uniform small curls. Around the protoplast of the treponema there are 3-4 fibrils (flagella). The cytoplasm contains cytoplasmic filaments. Pathogenic representatives are T. pallidum- the causative agent of syphilis, T. pertenue- causative agent of the tropical disease yaws. There are also saprophytes - inhabitants of the human oral cavity and the silt of reservoirs.
Borrelia(genus Borrelia), unlike treponemas, they are longer, have 3-8 large curls and 7-20 fibrils. These include the causative agent of relapsing fever (B. recurrentis) and pathogens of Lyme disease (V. burgdorferi) and other diseases.
Leptospira(genus Leptospira) They have shallow and frequent curls in the form of a twisted rope. The ends of these spirochetes are curved like hooks with thickenings at the ends. Forming secondary curls, they take on the shape of the letters S or C; have two axial fibrils. Pathogenic representative L. interrogans causes leptospirosis when ingested through water or food, leading to hemorrhage and jaundice.
Rickettsia have a metabolism independent of the host cell, however, it is possible that they receive high-energy compounds from the host cell for their reproduction. In smears and tissues they are stained according to Romanovsky-Giemsa, according to Macchiavello-Zdrodovsky (rickettsia are red, and infected cells are blue).
In humans, rickettsiae cause epidemic typhus. (R. prowazekii), tick-borne rickettsiosis (R. sibirica), Rocky Mountain spotted fever (R. rickettsii) and other rickettsioses.
The structure of their cell wall resembles that of gram-negative bacteria, although there are differences. It does not contain typical peptidoglycan: it completely lacks N-acetylmuramic acid. The cell wall consists of a double outer membrane, which includes lipopolysaccharide and proteins. Despite the absence of peptidoglycan, the cell wall of chlamydia is rigid. The cytoplasm of the cell is limited by the inner cytoplasmic membrane.
The main method for detecting chlamydia is Romanovsky-Giemsa staining. The color depends on the stage of the life cycle: elementary bodies appear purple against the background of the blue cytoplasm of the cell, reticular bodies appear blue.
In humans, chlamydia causes damage to the eyes (trachoma, conjunctivitis), urogenital tract, lungs, etc.
Actinomycetes- branching, filamentous or rod-shaped gram-positive bacteria. Its name (from Greek. actis- Ray, mykes- fungus) they received due to the formation in the affected tissues of drusen - granules of tightly intertwined threads in the form
rays extending from the center and ending in flask-shaped thickenings. Actinomycetes, like fungi, form mycelium - thread-like intertwining cells (hyphae). They form substrate mycelium, which is formed as a result of cell ingrowth into the nutrient medium, and aerial mycelium, which grows on the surface of the medium. Actinomycetes can divide by fragmentation of the mycelium into cells similar to rod-shaped and coccoid bacteria. On the aerial hyphae of actinomycetes, spores are formed that serve for reproduction. Actinomycete spores are usually not heat-resistant.
A common phylogenetic branch with actinomycetes is formed by the so-called nocardiform (nocardioform) actinomycetes - a collective group of rod-shaped bacteria of irregular shape. Their individual representatives form branching forms. These include bacteria of the genera Corynebacterium, Mycobacterium, Nocardia etc. Nocardi-like actinomycetes are distinguished by the presence in the cell wall of the sugars arabinose, galactose, as well as mycolic acids and large amounts of fatty acids. Mycolic acids and cell wall lipids determine the acid resistance of bacteria, in particular mycobacteria tuberculosis and leprosy (when stained according to Ziehl-Neelsen, they are red, and non-acid-resistant bacteria and tissue elements, sputum are blue).
Pathogenic actinomycetes cause actinomycosis, nocardia - nocardiosis, mycobacteria - tuberculosis and leprosy, corynebacteria - diphtheria. Saprophytic forms of actinomycetes and nocardiform actinomycetes are widespread in the soil, many of them are producers of antibiotics.
Mycoplasmas- small bacteria (0.15-1 µm), surrounded only by a cytoplasmic membrane containing sterols. They belong to the class Mollicutes. Due to the absence of a cell wall, mycoplasmas are osmotically sensitive. They have a variety of shapes: coccoid, filamentous, flask-shaped. These forms are visible under phase-contrast microscopy of pure cultures of mycoplasmas. On a dense nutrient medium, mycoplasmas form colonies that resemble fried eggs: a central opaque part immersed in the medium and a translucent periphery in the form of a circle.
Mycoplasmas cause atypical pneumonia in humans (Mycoplasma pneumoniae) and lesions of the genitourinary tract
(M. hominis and etc.). Mycoplasmas cause diseases not only in animals, but also in plants. Non-pathogenic representatives are also quite widespread.
2.3. Structure and classification of mushrooms
Mushrooms belong to the domain Eukarya, kingdom Fungi (Mycota, Mycetes). Recently, fungi and protozoa have been divided into separate kingdoms: kingdom Eumycota(real mushrooms), kingdom Chromista and kingdom Protozoa. Some microorganisms previously considered fungi or protozoa have been moved into a new kingdom Chromista(chrome plates). Fungi are multicellular or unicellular non-photosynthetic (chlorophyll-free) eukaryotic microorganisms with a thick cell wall. They have a nucleus with a nuclear envelope, cytoplasm with organelles, a cytoplasmic membrane and a multilayered rigid cell wall consisting of several types of polysaccharides (mannans, glucans, cellulose, chitin), as well as protein, lipids, etc. Some fungi form a capsule. The cytoplasmic membrane contains glycoproteins, phospholipids and ergosterols (as opposed to cholesterol, the main sterol of mammalian tissues). Most fungi are obligate or facultative aerobes.
Fungi are widespread in nature, especially in soil. Some mushrooms contribute to the production of bread, cheese, lactic acid products and alcohol. Other fungi produce antimicrobial antibiotics (eg, penicillin) and immunosuppressive drugs (eg, cyclosporine). Fungi are used by geneticists and molecular biologists to model various processes. Phytopathogenic fungi cause significant damage to agriculture, causing fungal diseases of cereal plants and grain. Infections caused by fungi are called mycoses. There are hyphal and yeast fungi.
Hyphal (mold) fungi, or hyphomycetes, consist of thin threads 2-50 microns thick, called hyphae, which are woven into a mycelium or mycelium (mold). The body of the mushroom is called the thallus. There are demacium (pigmented - brown or black) and hyaline (non-pigmented) hyphomycetes. Hyphae that grow into the nutrient substrate are responsible for feeding the fungus and are called vegetative hyphae. Hyphae, ra-
standing above the surface of the substrate are called aerial or reproductive hyphae (responsible for reproduction). Colonies have a fluffy appearance due to aerial mycelium.
There are lower and higher fungi: the hyphae of higher fungi are separated by partitions, or septa with holes. The hyphae of lower fungi do not have partitions, being multinucleated cells called coenocytic (from the Greek. koenos- single, common).
Yeast fungi (yeast) are mainly represented by individual oval cells with a diameter of 3-15 microns, and their colonies, unlike hyphal fungi, have a compact appearance. According to the type of sexual reproduction, they are distributed among higher fungi - ascomycetes and basidiomycetes. When reproducing asexually, yeast buds or divides. They can form pseudohyphae and false mycelium (pseudomycelium) in the form of chains of elongated cells - “sausages”. Fungi that are similar to yeast, but do not have a sexual method of reproduction, are called yeast-like. They reproduce only asexually - by budding or fission. The concepts of “yeast-like fungi” are often identified with the concept of “yeast”.
Many fungi have dimorphism - the ability to grow hyphal (mycelial) or yeast-like, depending on the cultivation conditions. In an infected organism, they grow in the form of yeast-like cells (yeast phase), and on nutrient media they form hyphae and mycelium. Dimorphism is associated with the temperature factor: at room temperature mycelium is formed, and at 37 ° C (at human body temperature) yeast-like cells are formed.
Fungi reproduce sexually or asexually. Sexual reproduction of fungi occurs with the formation of gametes, sexual spores and other sexual forms. Sexual forms are called teleomorphs.
Asexual reproduction of fungi occurs with the formation of corresponding forms called anamorphs. Such reproduction occurs by budding, fragmentation of hyphae and asexual spores. Endogenous spores (sporangiospores) mature inside a round structure - a sporangium. Exogenous spores (conidia) are formed at the tips of fruiting hyphae, the so-called conidiophores.
There are a variety of conidia. Arthroconidia (arthrospores), or talloconidia, are formed by uniform septation and dismemberment of hyphae, and blastoconidia are formed as a result of budding. Small unicellular conidia are called microconidia, large multicellular conidia are called macroconidia. Asexual forms of fungi also include chlamydoconidia, or chlamydospores (thick-walled large resting cells or a complex of small cells).
There are perfect and imperfect mushrooms. Perfect mushrooms have a sexual method of reproduction; these include zygomycetes (Zygomycota), ascomycetes (Ascomycota) and basidiomycetes (Basidiomycota). Imperfect fungi have only asexual reproduction; These include the formal conventional type/group of fungi - deuteromycetes (Deiteromycota).
Zygomycetes belong to the lower fungi (nonseptate mycelium). They include representatives of the genera Mucor, Rhizopus, Rhizomucor, Absidia, Basidiobolus, Conidiobolus. Distributed in soil and air. They can cause zygomycosis (mucoromycosis) of the lungs, brain and other human organs.
During asexual reproduction of zygomycetes, a sporangium is formed on the fruiting hypha (sporangiophores) - a spherical thickening with a shell containing numerous sporangiospores (Fig. 2.6, 2.7). Sexual reproduction in zygomycetes occurs with the help of zygospores.
Ascomycetes (marsupial fungi) have septate mycelium (except for unicellular yeasts). They got their name from the main fruiting organ - the bursa, or ascus, containing 4 or 8 haploid sexual spores (ascospores).
Ascomycetes include individual representatives (teleomorphs) of the genera Aspergillus And Penicillium. Most fungi genera Aspergillus, Penicillium are anamorphic, i.e. they reproduce only helplessly
 Rice. 2.6. Mushrooms of the genus Mucor(drawing by A.S. Bykov)
Rice. 2.6. Mushrooms of the genus Mucor(drawing by A.S. Bykov)
 Rice. 2.7. Mushrooms of the genus Rhizopus. Development of sporangium, sporangiospores and rhizoids
Rice. 2.7. Mushrooms of the genus Rhizopus. Development of sporangium, sporangiospores and rhizoids
directly with the help of asexual spores - conidia (Fig. 2.8, 2.9) and should be classified according to this characteristic as imperfect fungi. In mushrooms of the genus Aspergillus at the ends of the fruiting hyphae, conidiophores, there are thickenings - sterigmata, phialides, on which chains of conidia are formed (“water mold”).
In mushrooms of the genus Penicillium(tassel) the fruiting hypha resembles a brush, since thickenings are formed from it (on the conidiophore), branching into smaller structures - sterigmata, phialides, on which there are chains of conidia. Some species of Aspergillus can cause aspergillosis and aflatoxicosis, and Penicillium can cause penicilliosis.
Representatives of ascomycetes are teleomorphs of the genera Trichophyton, Microsporum, Histoplasma, Blastomyces, as well as tremors
 Rice. 2.8. Mushrooms of the genus Penicillium. Chains of conidia extend from the phialids
Rice. 2.8. Mushrooms of the genus Penicillium. Chains of conidia extend from the phialids
 Rice. 2.9. Mushrooms of the genus Aspergillus fumigatus. Chains of conidia extend from the phialids
Rice. 2.9. Mushrooms of the genus Aspergillus fumigatus. Chains of conidia extend from the phialids
Basidiomycetes include cap mushrooms. They have septate mycelium and form sexual spores - basidiospores by detaching from the basidium - the terminal cell of the mycelium, homologous to the ascus. Basidiomycetes include some yeasts, such as teleomorphs Cryptococcus neoformans.
Deuteromycetes are imperfect fungi (Fungi imperfecti, anamorphic fungi, conidial fungi). This is a conditional, formal taxon of fungi, uniting fungi that do not have sexual reproduction. Recently, instead of the term “deuteromycetes”, the term “mitosporous fungi” was proposed - fungi that reproduce by non-sexual spores, i.e. by mitosis. When the fact of sexual reproduction of imperfect fungi is established, they are transferred to one of the known types - Ascomycota or Basidiomycota assigning the name to a teleomorphic form. Deuteromycetes have septate mycelium and reproduce only through the asexual formation of conidia. Deuteromycetes include imperfect yeasts (yeast-like fungi), for example, some fungi of the genus Candida, affecting the skin, mucous membranes and internal organs (candidiasis). They are oval in shape, 2-5 microns in diameter, divide by budding, form pseudohyphae (pseudomycelium) in the form of chains of elongated cells, and sometimes form hyphae. For Candida albicans the formation of chlamydospores is characteristic (Fig. 2.10). Deuteromycetes also include other fungi that do not have a sexual method of reproduction, belonging to the genera Epidermophyton, Coccidioides, Paracoccidioides, Sporothrix, Aspergillus, Phialophora, Fonsecaea, Exophiala, Cladophialophora, Bipolaris, Exerohilum, Wangiella, Alrernaria and etc.
 Rice. 2.10. Mushrooms of the genus Candida albicans(drawing by A.S. Bykov)
Rice. 2.10. Mushrooms of the genus Candida albicans(drawing by A.S. Bykov)
2.4. Structure and classification of protozoa
The simplest belong to the domain Eukarya, animal kingdom (Animalia), sub-kingdom Protozoa. It has recently been proposed to allocate protozoa to the rank of kingdom Protozoa.
The protozoan cell is surrounded by a membrane (pellicle) - an analogue of the cytoplasmic membrane of animal cells. It has a nucleus with a nuclear envelope and nucleolus, a cytoplasm containing the endoplasmic reticulum, mitochondria, lysosomes and ribosomes. The sizes of protozoa range from 2 to 100 microns. When stained according to Romanovsky-Giemsa, the nucleus of the protozoa is red, and the cytoplasm is blue. Protozoa move with the help of flagella, cilia or pseudopodia, some of them have digestive and contractile (excretory) vacuoles. They can feed as a result of phagocytosis or the formation of special structures. Based on the type of nutrition, they are divided into heterotrophs and autotrophs. Many protozoa (dysenteric amoeba, Giardia, Trichomonas, Leishmania, Balantidia) can grow on nutrient media containing native proteins and amino acids. Cell cultures, chicken embryos and laboratory animals are also used for their cultivation.
Protozoa reproduce asexually - by double or multiple (schizogony) division, and some also sexually (sporogony). Some protozoa reproduce extracellularly (Giardia), while others reproduce intracellularly (Plasmodium, Toxoplasma, Leishmania). The life cycle of protozoa is characterized by stages - the formation of the trophozoite stage and the cyst stage. Cysts are dormant stages, resistant to changes in temperature and humidity. Cysts are acid resistant Sarcocystis, Cryptosporidium And Isospora.
Previously, protozoa that cause diseases in humans were represented by 4 types 1 ( Sarcomastigophora, Apicomplexa, Ciliophora, Microspora). These types have recently been reclassified into larger numbers, and new kingdoms have emerged - Protozoa And Chromista(Table 2.2). To a new kingdom Chromista(chromovics) included some protozoa and fungi (blastocysts, oomycetes and Rhinosporidium seeberi). Kingdom Protozoa includes amoebas, flagellates, sporozoans and ciliates. They are divided into different types, among which are amoebas, flagellates, sporozoans and ciliates.
Table 2.2. Representatives of the kingdoms Protozoa And Chromista, of medical importance
 1 Type Sarcomastigophora consisted of subtypes Sarcodina And Mastigophora. Subtype Sarcodina(sarcodaceae) included the dysenteric amoeba, and the subtype Mastigophora(flagellates) - trypanosomes, leishmania, lamblia and trichomonas. Type Apicomplexa included class Sporozoa(spores), which included plasmodia of malaria, toxoplasma, cryptosporidium, etc. Type Ciliophora includes balantidia, and type Microspora- microsporidia.
1 Type Sarcomastigophora consisted of subtypes Sarcodina And Mastigophora. Subtype Sarcodina(sarcodaceae) included the dysenteric amoeba, and the subtype Mastigophora(flagellates) - trypanosomes, leishmania, lamblia and trichomonas. Type Apicomplexa included class Sporozoa(spores), which included plasmodia of malaria, toxoplasma, cryptosporidium, etc. Type Ciliophora includes balantidia, and type Microspora- microsporidia.
End of table. 2.2
 Amoebas include the causative agent of human amoebiasis - amoebic dysentery (Entamoeba histolytica), free-living and non-pathogenic amoebas (intestinal amoeba, etc.). Amoebas reproduce binary asexually. Their life cycle consists of a trophozoite stage (a growing, motile cell, weakly stable) and a cyst stage. Trophozoites move with the help of pseudopodia, which capture and immerse nutrients into the cytoplasm. From
Amoebas include the causative agent of human amoebiasis - amoebic dysentery (Entamoeba histolytica), free-living and non-pathogenic amoebas (intestinal amoeba, etc.). Amoebas reproduce binary asexually. Their life cycle consists of a trophozoite stage (a growing, motile cell, weakly stable) and a cyst stage. Trophozoites move with the help of pseudopodia, which capture and immerse nutrients into the cytoplasm. From
The trophozoite forms a cyst that is resistant to external factors. Once in the intestine, it turns into a trophozoite.
Flagellates are characterized by the presence of flagella: Leishmania has one flagellum, Trichomonas has 4 free flagella and one flagellum connected to a short undulating membrane. They are:
Flagellates of blood and tissues (Leishmania - the causative agents of leishmaniasis; Trypanosomes - the causative agents of sleeping sickness and Chagas disease);
Intestinal flagellates (giardia - the causative agent of giardiasis);
Flagellates of the genitourinary tract (Trichomonas vaginalis - the causative agent of trichomoniasis).
The ciliated ones are represented by balantidia, which affect the human colon (balantidiasis dysentery). Balantidia have a trophozoite and cyst stage. The trophozoite is mobile, has numerous cilia, thinner and shorter than flagella.
2.5. Structure and classification of viruses
Viruses are the smallest microbes belonging to the kingdom Virae(from lat. virus- I). They do not have a cellular structure and consist
The structure of viruses, due to their small size, is studied using electron microscopy of both virions and their ultrathin sections. The sizes of viruses (virions) are determined directly using electron microscopy or indirectly by ultrafiltration through filters with a known pore diameter, or by ultracentrifugation. The size of viruses ranges from 15 to 400 nm (1 nm is equal to 1/1000 μm): small viruses, the size of which is similar to the size of ribosomes, include parvoviruses and poliovirus, and the largest are variola virus (350 nm). Viruses differ in the shape of their virions, which have the form of rods (tobacco mosaic virus), bullets (rabies virus), spheres (poliomyelitis viruses, HIV), threads (filoviruses), sperm (many bacteriophages).
Viruses amaze the imagination with their diversity of structure and properties. Unlike cellular genomes, which contain uniform double-stranded DNA, viral genomes are extremely diverse. There are DNA and RNA viruses that are haploid, i.e. have one set of genes. Only retroviruses have a diploid genome. The genome of viruses contains from 6 to 200 genes and is represented by various types of nucleic acids: double-stranded, single-stranded, linear, circular, fragmented.
Among single-stranded RNA viruses, a distinction is made between genomic plus-strand RNA and minus-strand RNA (RNA polarity). The plus strand (positive strand) of the RNA of these viruses, in addition to the genomic (hereditary) function, performs the function of informational or messenger RNA (mRNA, or mRNA); it is a matrix for protein synthesis on the ribosomes of the infected cell. Plus-strand RNA is infectious: when introduced into sensitive cells, it can cause an infectious process.
cess. The minus strand (negative strand) of RNA viruses performs only a hereditary function; For protein synthesis, a complementary strand is synthesized on the minus strand of RNA. Some viruses have an ambipolar RNA genome (ambisense from Greek ambi- on both sides, double complementarity), i.e. contains plus and minus RNA segments.
There are simple viruses (for example, hepatitis A virus) and complex viruses (for example, influenza viruses, herpes, coronaviruses).
Simple, or non-enveloped, viruses have only nucleic acid associated with a protein structure called a capsid (from the Latin. capsa- case). Proteins associated with nucleic acid are known as nucleoproteins, and the association of viral capsid proteins with viral nucleic acid is called nucleocapsid. Some simple viruses can form crystals (eg foot-and-mouth disease virus).
The capsid includes repeating morphological subunits - capsomeres, composed of several polypeptides. The nucleic acid of the virion binds to the capsid to form a nucleocapsid. The capsid protects the nucleic acid from degradation. In simple viruses, the capsid is involved in attachment (adsorption) to the host cell. Simple viruses leave the cell as a result of its destruction (lysis).
Complex, or enveloped, viruses (Fig. 2.11), in addition to the capsid, have a membrane double lipoprotein envelope (synonym: supercapsid, or peplos), which is acquired by budding of the virion through the cell membrane, for example through the plasma membrane, nuclear membrane or endoplasmic reticulum membrane. The virus envelope contains glycoprotein spikes,
or spines, peplomers. Destruction of the shell with ether and other solvents inactivates complex viruses. Under the shell of some viruses is a matrix protein (M protein).
Virions have a helical, icosahedral (cubic) or complex type of capsid (nucleocapsid) symmetry. The helical type of symmetry is due to the helical structure of the nucleocapsid (for example, in influenza viruses, coronaviruses): capsomeres are arranged in a spiral along with the nucleic acid. The icosahedral type of symmetry is due to the formation of an isometrically hollow body from the capsid containing the viral nucleic acid (for example, in the herpes virus).
The capsid and shell (supercapsid) protect virions from environmental influences and determine selective interaction (adsorption) of their receptor proteins with certain
 Rice. 2.11. The structure of enveloped viruses with an icosahedral (a) and helical (b) capsid
Rice. 2.11. The structure of enveloped viruses with an icosahedral (a) and helical (b) capsid
cells, as well as antigenic and immunogenic properties of virions.
The internal structures of viruses are called the core. In adenoviruses, the core consists of histone-like proteins associated with DNA, in reoviruses - from proteins of the internal capsid.
Nobel Prize winner D. Baltimore proposed the Baltimore classification system based on the mechanism of mRNA synthesis. This classification places viruses in 7 groups (Table 2.3). International Committee on Taxonomy of Viruses (ICTV) adopted a universal classification system that uses taxonomic categories such as family (name ending in viridae), subfamily (name ends in virinae), genus (name ends in virus). The virus species did not receive a binomial name, like bacteria. Viruses are classified by the type of nucleic acid (DNA or RNA), its structure and the number of strands. They have double-stranded or single-stranded nucleic acids; positive (+), negative (-) nucleic acid polarity or mixed nucleic acid polarity, ambipolar (+, -); linear or circular nucleic acid; fragmented or non-fragmented nucleic acid. The size and morphology of virions, the number of capsomeres and the type of symmetry of the nucleocapsid, the presence of an envelope (supercapsid), sensitivity to ether and deoxycholate, place of reproduction in the cell, antigenic properties, etc. are also taken into account.
Table 2.3. Major viruses of medical importance
 Continuation of the table. 2.3
Continuation of the table. 2.3
 End of table. 2.3
End of table. 2.3
 Viruses infect animals, bacteria, fungi and plants. Being the main causative agents of human infectious diseases, viruses are also involved in the processes of carcinogenesis and can be transmitted in various ways, including through the placenta (rubella virus, cytomegalovirus, etc.), affecting the human fetus. They can also lead to post-infectious complications - the development of myocarditis, pancreatitis, immunodeficiency, etc.
Viruses infect animals, bacteria, fungi and plants. Being the main causative agents of human infectious diseases, viruses are also involved in the processes of carcinogenesis and can be transmitted in various ways, including through the placenta (rubella virus, cytomegalovirus, etc.), affecting the human fetus. They can also lead to post-infectious complications - the development of myocarditis, pancreatitis, immunodeficiency, etc.
In addition to viruses, non-cellular life forms include prions and viroids. Viroids are small molecules of circular, supercoiled RNA that do not contain protein and cause plant diseases. Pathological prions are infectious protein particles that cause special conformational diseases as a result of changes in the structure of normal cellular prion protein ( PrP c), which is present in the body of animals and humans. PrP with performs regulatory functions. It is encoded by the normal prion gene (PrP gene), located on the short arm of human chromosome 20. Prion diseases occur as transmissible spongiform encephalopathies (Creutzfeldt-Jakob disease, kuru, etc.). In this case, the prion protein acquires a different, infectious form, designated as PrP sc(sc from scrapie- scrapie is a prion infection of sheep and goats). This infectious prion protein has the appearance of fibrils and differs from normal prion protein in its tertiary or quaternary structure.
Tasks for self-preparation (self-control)
A. Label the microbes that are prokaryotes:
2. Viruses.
3. Bacteria.
4. Prions.
B. Note the distinctive features of a prokaryotic cell:
1. 70S ribosomes.
2. The presence of peptidoglycan in the cell wall.
3. Presence of mitochondria.
4. Diploid set of genes.
IN. Label the components of peptidoglycan:
1. Teichoic acids.
2. N-acetylglucosamine.
3. Lipopolysaride.
4. Tetrapeptide.
G. Note the structural features of the cell wall of gram-negative bacteria:
1. Mesodiaminopimelic acid.
2. Teichoic acids.
4. Porin proteins.
D. Name the functions of spores in bacteria:
1. Preservation of the species.
2. Heat resistance.
3. Dispersal of the substrate.
4. Reproduction.
1. Rickettsia.
2. Actinomycetes.
3. Spirochetes.
4. Chlamydia.
AND. Name the features of actinomycetes:
1. They have thermolabile spores.
2. Gram-positive bacteria.
3. There is no cell wall.
4. They have a convoluted shape.
Z. Name the features of spirochetes:
1. Gram-negative bacteria.
2. They have a motor fibrillary apparatus.
3. They have a convoluted shape.
AND. Name the protozoa that have an apical complex that allows them to penetrate into the cell:
1. Malarial plasmodium.
3. Toxoplasma.
4. Cryptosporidium.
TO. Name the distinctive feature of complex viruses:
1. Two types of nucleic acid.
2. The presence of a lipid membrane.
3. Double capsid.
4. Presence of non-structural proteins. L. Mark the higher mushrooms:
1. Mucor.
2. Candida.
3. Penicillium.
4. Aspergillus.
Nutrition of bacteria.
Nutrition.
Passive diffusion
Facilitated diffusion
Active transport
In the first case, the nutrient molecule forms a complex with a protein of the periplasmic space, which interacts with a specific permease of the cytoplasmic membrane. After energy-dependent penetration through the cytoplasmic membrane, the “substrate - periplasmic protein - permease” complex dissociates with the release of the substrate molecule.
During active transport with chemical modification of the transported substance, the chain of events includes: (1) phosphorylation of membrane enzyme-2 from the cytoplasm by phosphoenolpyruvate; (2) binding of substrate molecules by phosphorylated enzyme-2 to the surface of the cytoplasmic membrane; (3) energy-dependent transport of the substrate molecule into the cytoplasm; (4) transfer of a phosphate group to a substrate molecule; (5) dissociation of the “substrate-enzyme” complex in the cytoplasm. Due to phosphorylation, substrate molecules accumulate in the cytoplasm of cells and are not able to leave them.
Classification of bacteria by type of nutrition.
According to the method of nutrient intake substances bacteria are divided into holophytes And Holozoans. Holophytic bacteria (from Greek holos- complete and phyticos– related to plants) are unable to release enzymes into the environment that break down substrates, as a result of which they consume nutrients exclusively in dissolved, molecular form. Holozoan bacteria (from Greek holos- complete and zoikos- related to animals), on the contrary, have a complex of exoenzymes that provide external nutrition - the breakdown of substrates into molecules outside the bacterial cell. After this, the nutrient molecules enter the holozoan bacteria.
By carbon source among bacteria there are autotrophs And heterotrophs. Autotrophs (from Greek autos- myself, trophe– food) carbon dioxide (CO 2) is used as a carbon source, from which all carbon-containing substances are synthesized. For heterotrophs (from the Greek geteros - another and trophe– food) the source of carbon is various organic substances in molecular form (carbohydrates, polyhydric alcohols, amino acids, fatty acids). The greatest degree of heterotrophy is inherent in prokaryotes, which can live only inside other living cells (for example, rickettsia and chlamydia).
By nitrogen source Prokaryotes are divided into 3 groups: 1) nitrogen-fixing bacteria (assimilate molecular nitrogen from atmospheric air); 2) bacteria that consume inorganic nitrogen from ammonium salts, nitrites or nitrates; 3) bacteria that assimilate nitrogen contained in organic compounds (amino acids, purines, pyrimidines, etc.).
By energy source bacteria are divided into phototrophs And chemotrophs. Phototrophic bacteria , like plants, are able to use solar energy. Phototrophic prokaryotes do not cause diseases in humans. Chemotrophic bacteria obtain energy from redox reactions.
By the nature of electron donors lithotrophs(from Greek litos- stone) and organotrophs. U lithotrophs (chemolithotrophs ) inorganic substances (H 2, H 2 S, NH 3, sulfur, CO, Fe 2+, etc.) act as electron donors. Donors of electrons organotrophs (chemoorganotrophs ) are organic compounds - carbohydrates, amino acids, etc.
Most bacteria pathogenic to humans have a chemoorganotrophic (chemoheterotrophic) type of nutrition; The chemolithotrophic (chemoautotrophic) type is less common.
Based on the ability to synthesize organic compounds chemotrophic bacteria are divided into prototrophs, auxotrophs And hypotrophs. Prototrophic bacteria synthesize all necessary organic substances from glucose and ammonium salts. Bacteria are called auxotrophs , if they are unable to synthesize any organic matter from the specified compounds. The extreme degree of loss of metabolic activity is called malnutrition. Hypotrophic bacteria ensure their vital functions by reorganizing the structures or metabolites of the host.
In addition to carbon and nitrogen, bacteria need sulfur, phosphorus, and metal ions for full life. Sources of sulfur are amino acids (cysteine, methionine), vitamins, cofactors (biotin, lipoic acid, etc.), sulfates. Sources of phosphorus are nucleic acids, phospholipids, and phosphates. In sufficiently high concentrations, bacteria need magnesium, potassium, calcium, and iron; in much smaller ones - zinc, manganese, sodium, molybdenum, copper, nickel, cobalt.
Growth factors- these are substances that bacteria cannot synthesize on their own, but they desperately need them. Growth factors can include amino acids, nitrogenous bases, vitamins, fatty acids, iron porphyrins and other compounds. To create optimal conditions for the life of bacteria, growth factors must be added to the nutrient media.
Metabolism, energy conversion
A) Constructive metabolism.
An obligatory phase of bacterial nutrition is the assimilation of nutrients, that is, their inclusion in an altered or modified form in synthetic reactions to reproduce cellular components and structures.
Protein metabolism in bacteria it can occur in 3 phases: primary protein breakdown, secondary breakdown and protein synthesis. The primary breakdown of protein molecules into peptones is carried out by exoenzymes - exoproteases secreted by bacteria into the environment. Secondary breakdown occurs under the action of endoenzymes (endoproteases), which all bacteria have. This process takes place inside the bacterial cell and consists of the breakdown of peptides into their constituent amino acids. The latter can be used unchanged or subjected to chemical transformations (deamination, decarboxylation, etc.), as a result of which ammonia, indole, hydrogen sulfide, keto acids, alcohol, carbon dioxide, etc. appear. The detection of these compounds has diagnostic value in bacteriology.
Along with the reactions of protein breakdown, reactions of their synthesis occur. Some bacteria form proteins from ready-made amino acids obtained as a result of external nutrition, while other bacteria independently synthesize amino acids from simple compounds containing nitrogen and carbon. The synthesis of amino acids can be carried out in the reactions of amination, transamination, amidation, and carboxylation. Most prokaryotes are capable of synthesizing all the amino acids that make up cellular proteins. A feature of the biosynthesis of amino acids is the use of common biosynthetic pathways: the tricarboxylic acid cycle, glycolysis, the oxidative pentose-phosphate pathway, etc. The main starting compounds for the synthesis of amino acids are pyruvate and fumarate.
Carbohydrate metabolism autotrophs and heterotrophs have differences (Scheme 1). Autotrophic bacteria synthesize all necessary carbohydrates from carbon dioxide. The raw materials for the formation of carbohydrates in heterotrophic bacteria can be: (1) one-, two- and three-carbon compounds; and (2) polysaccharides (starch, glycogen, cellulose). To break down the latter, many heterotrophic bacteria have exoenzymes (amylase, pectinase, etc.), which hydrolyze polysaccharides to form glucose, maltose, fructose, etc.
In autotrophic bacteria, in the Calvin cycle, ribulose phosphate-phosphoric-glyceric acid is formed from carbon dioxide, which is included in glycolytic reactions going in the opposite direction. The final product of reverse synthesis is glucose.
Heterotrophic bacteria form glucose from one-, two- and three-carbon compounds, also including them in the reverse glycolysis reaction. Due to the fact that some reactions of glycolysis are irreversible, heterotrophs have developed special enzymatic reactions that allow them to bypass the irreversible reactions of the catabolic pathway.
When polysaccharides are broken down by heterotrophic bacteria, the resulting disaccharides enter the cells and, under the influence of maltose, sucrose, and lactose, undergo hydrolysis and decomposition into monosaccharides, which are then fermented or included in sugar interconversion reactions.
Lipid metabolism. Both exogenous lipids and amphibolites of interstitial metabolism can serve as starting materials for the formation of lipids in bacteria. Exogenous lipids are exposed to bacterial lipases and other lipolytic enzymes. Many types of bacteria metabolize glycerol, which serves as a source of plastic material and energy. Endogenous sources for lipid synthesis can be acetyl coenzyme A, propionyl-ACP, malonyl-ACP (ACP - acetyl transfer protein), phosphodioxyacetone, etc.
The starting substrate for the synthesis of fatty acids with an even number of carbon atoms is acetyl coenzyme A, for fatty acids with an odd number of carbon atoms - propionyl-ACP and malonyl-ACP. The formation of double bonds in an acid molecule in aerobic prokaryotes occurs with the participation of molecular oxygen and the enzyme desaturase. In anaerobic prokaryotes, double bonds are introduced early in synthesis as a result of a dehydration reaction. The starting substrate for the synthesis of phospholipids is phosphodioxyacetone (an intermediate in the glycolytic pathway), the reduction of which leads to the formation of 3-phosphoroglycerol. The latter is then joined by 2 fatty acid residues in the form of a complex with ACP. The product of the reaction is phosphatidic acid, the activation of which by CTP and the subsequent addition of serine, inositol, glycerol or another compound to the phosphate group leads to the synthesis of the corresponding phospholipids.
Microorganisms that are auxotrophic and hypotrophic in fatty acids (for example, mycoplasmas) obtain them in ready-made form from host cells or a nutrient medium.
Mononucleotide exchange. Purine and pyrimidine mononucleotides are essential components of DNA and RNA. Many prokaryotes are capable of both using ready-made purine and pyrimidine bases, their nucleosides and nucleotides contained in the nutrient medium, and synthesizing them from low molecular weight substances. Bacteria have enzymes that catalyze the following stages of interconversion of exogenous purine and pyrimidine derivatives: nitrogenous base – nucleoside – nucleotide (mono- – di- – triphosphate).
Synthesis of purine and pyrimidine mononucleotides de novo carried out in independent ways. During the synthesis of purine nucleotides, as a result of successive enzymatic reactions, inosinic acid is formed, from which adenyl (AMP) and guanylic (GMP) acids are synthesized through chemical modifications of the purine ring. The synthesis of pyrimidine nucleotides begins with the formation of orotidylic acid, the decarboxylation of which produces uridylic acid (UMA). From the latter, UTP is formed, the acylation of which leads to the formation of CTP.
Deoxyribonucleotides are formed as a result of the reduction of the corresponding ribonucleotides at the level of diphosphates or triphosphates. The synthesis of a DNA-specific nucleotide, thymidylic acid, occurs through enzymatic methylation of deoxyuridylic acid.
Ion exchange. Mineral compounds - ions, NH 3 +, K +, Mg 2+, Fe 2+, SO 4 2-, PO 4 3- and other bacteria are obtained from the environment both in a free state and in a state associated with other organic substances. Cations and anions are transported into the bacterial cell in various ways, described in § 3. The rate of penetration of ions into the bacterial cell is influenced by the pH of the environment and the physiological activity of the microorganisms themselves.
B) Respiration of bacteria (energy metabolism).
All life processes are energy dependent, so energy production is an extremely important aspect of prokaryotic metabolism. They obtain energy through anaerobic and aerobic respiration.
Breath, or biological oxidation is a catabolic process of electron transfer from a donor substance to an acceptor substance, accompanied by the accumulation of energy in high-energy compounds . Respiration occurs through the process of catabolic reactions, as a result of which complex organic substances, when broken down, release energy and are converted into simple compounds. The energy accumulated in macroergic substances (ATP, GTP, UTP, etc.) is used in anabolic reactions.
According to the method of respiration, microorganisms are divided into obligate (strict) aerobes, obligate anaerobes And facultative anaerobes.
Obligate aerobes need free oxygen. The electron donors in chemoorganotrophic aerobes that are pathogenic for humans are organic compounds (carbohydrates, fats, proteins), and the electron acceptor is molecular oxygen. Energy storage in the form of ATP in chemoorganotrophic aerobes occurs during oxidative phosphorylation of electron donors. Aerobes have cytochromes (participated in electron transfer), as well as enzymes (catalase, superoxide dismutase, peroxidase), which inactivate toxic oxygen radicals formed during respiration. Superoxide dismutase inactivates the most toxic metabolite, the superoxide radical in H2O2. The enzyme catalase converts H 2 O 2 into H 2 O and O 2 .
A special group of aerobes are microaerophilic bacteria which, although they require oxygen to obtain energy, grow better with increased CO 2 content, for example, bacteria of the genera Campylobacter And Helicobacter.
Obligate anaerobes do not need free oxygen; on the contrary, even in small quantities oxygen has a toxic effect on them. Various organic compounds (mainly carbohydrates) serve as electron donors for chemoorganotrophic anaerobes that are pathogenic to humans. The electron acceptor in chemoorganotrophic anaerobes are organic oxygen-containing compounds - acids or ketones, that is, the electron acceptor is oxygen bound to an organic fragment. Energy storage in these prokaryotes occurs through substrate phosphorylation. Obligate anaerobes, as a rule, do not have cytochromes and enzymes that inactivate oxygen radicals (catalase- and superoxide dismutase-negative).
In chemolithotrophic anaerobes, which are non-pathogenic to humans, the electron acceptor is inorganic oxygen-containing compounds - nitrates, sulfates, carbonates.
A special group of anaerobes consists of aerotolerant bacteria, which are able to grow in the presence of atmospheric oxygen, but do not use it as an electron acceptor (for example, lactic acid bacteria). Aerotolerant prokaryotes are catalase- and superoxide dismutase-positive.
Facultative anaerobes are able to exist in both oxygen and oxygen-free environments. Their electron donors are organic substances; electron acceptors, depending on environmental conditions - molecular oxygen or oxygen bound in organic and inorganic compounds. Energy can be accumulated by facultative anaerobes through both oxidative and substrate phosphorylation. Like aerobes, this group of bacteria has cytochromes and antioxidant enzymes.
The main substrate for energy production is carbohydrates, which in chemoheterotrophic prokaryotes of different types of respiration can be catabolized to acetyl coenzyme A (“activated acetic acid”). Lipids and proteins can act as energy substrates, since acetyl coenzyme A is also one of the intermediate products of their metabolism (Scheme 2).
Carbohydrate catabolism in chemoorganotrophic prokaryotes includes: (a) anaerobic processes - glycolysis, pentose phosphate pathway and ketodeoxyphosphogluconate pathway; (b) aerobic process - tricarboxylic acid cycle (Krebs cycle). Anaerobic processes occur in all prokaryotes, while the aerobic process is characteristic only of obligate aerobes and facultative anaerobes. Energy production by anaerobic pathways is based on substrate phosphorylation, while the aerobic process is based on oxidative phosphorylation.
Definition of concepts.
Sterilization, disinfection and antiseptics are integral parts of modern medical and especially surgical practice. Understanding the principles and practical application of these methods is necessary because many potentially pathogenic microorganisms can remain viable outside the host for long periods of time, exhibit high resistance to physical and chemical disinfectants, and are relatively easily transmitted from one person to another.
Antiseptics- destruction or prevention of the growth of pathogenic or opportunistic microorganisms by chemical methods. This term is usually used to refer to the external application of a chemical drug to living tissue.
Antiseptic- a substance that inhibits the growth or destroys a microorganism (without affecting bacterial spores). The term is specific for substances that are used for local action on living tissues.
Asepsis means the absence of sepsis, but in general this term is used to emphasize the absence of any living organisms. Aseptic techniques means any procedure designed to eliminate living organisms and prevent recontamination by them. Modern surgical and microbiological techniques are based on aseptic procedures.
Biocide- a substance that kills all living microorganisms, both pathogenic and non-pathogenic, including spores.
Biostat- an agent that prevents the growth of microorganisms, but does not necessarily kill them.
Decontamination- removal of microorganisms without quantitative determination. This term is relative; The final removal of germs can be accomplished by sterilization or disinfection.
Disinfection- a process that reduces the number or completely destroys all pathogenic microorganisms except spores.
Germicide- a substance that destroys microorganisms, especially pathogenic ones. The germicide does not destroy spores.
Sanitation- a method by which microbial contamination is reduced to a “safe” level. This method has previously been used to “purify” inanimate objects.
Sterilization- the use of physical factors and (or) chemicals for the complete destruction or destruction of all forms of microbial life.
Sterilization.
Sterilization is defined as the destruction or removal (by filtration) of all microorganisms and their spores. Sterilization is usually done using heat. Sterilization, being one of the routine procedures in the operation of a microbiology laboratory, is a necessary method to ensure that the cultures, equipment, glassware and media support the growth of only the necessary microorganisms, while other microbes are destroyed. There are the following types of sterilization: calcination in a burner flame, boiling, action with flowing steam, steam under pressure in an autoclave, dry heat, pasteurization, tindialization, chemical, cold (mechanical) sterilization.
Selection of sterilization methods.
When choosing sterilization methods, the following requirements must be taken into account:
1. Activity: bactericidal, sporicidal, tuberculocidal, fungicidal and viruscidal.
2. Speed of procedure: sterilization should be carried out as quickly as possible.
3. Permeability: sterilant substances must penetrate through the packaging and into the internal parts of the instrument.
4. Compatibility: There should be no change in the structure or function of materials that are sterilized several times.
5. Non-toxic: there should be no threat to human health or the environment.
6. Stability of organic material: The effectiveness of sterilization should not be impaired in the presence of organic material.
7. Adaptability: ability to use for large and small volumes of sterilized material.
8. Control over time: The processing cycle must be easily and accurately controlled.
9. Price: reasonable cost of equipment, installation and operation.
 Physical sterilizers
Physical sterilizers
Wet warm, which is formed during the steam autoclaving process, is the main sterilizing agent used in clinical microbiology laboratories. Autoclaves are used to sterilize culture media, heat-resistant materials, and treat infectious waste. A steam sterilizer, or autoclave, is an insulated, pressurized chamber that uses saturated steam to create high temperatures (Figure 1). Air is removed from the chamber by mass displacement or by creating a vacuum. The most commonly used autoclaves are those with mass substitution. Lighter steam is released into the chamber to displace heavier air. Brief pressure steam treatment can destroy bacterial spores. For routine sterilization of culture media and other materials, the exposure time is 15 minutes at 121ºC and the pressure is 1.5 kg per 1 square centimeter. For infectious waste, exposure time increases to 30-60 minutes. In addition to the correct time and temperature, direct contact with steam is very important during sterilization. When processing infectious material, maximum steam penetration into the waste should be ensured. Such material must be processed at a temperature of 132ºС. Antineoplastic drugs, toxic chemicals and radioisotopes that may not degrade, and unstable chemicals that may evaporate and spread throughout the chamber due to heat should not be autoclaved.
 Dry heat sterilization used for materials that cannot be sterilized with steam due to the possibility of damage or due to the material being impermeable to steam. Dry heat is less effective than moist heat and requires longer exposure times and higher temperatures. Dry heat sterilization is usually carried out in a dry heat oven (Fig. 2). The mechanism of dry heat sterilization is an oxidative process. Examples of materials that use dry heat sterilization include oils, powders, sharp instruments, and glassware. Dry heat or thermal inactivation-sterilization are used as alternative methods for treating infectious waste.
Dry heat sterilization used for materials that cannot be sterilized with steam due to the possibility of damage or due to the material being impermeable to steam. Dry heat is less effective than moist heat and requires longer exposure times and higher temperatures. Dry heat sterilization is usually carried out in a dry heat oven (Fig. 2). The mechanism of dry heat sterilization is an oxidative process. Examples of materials that use dry heat sterilization include oils, powders, sharp instruments, and glassware. Dry heat or thermal inactivation-sterilization are used as alternative methods for treating infectious waste.
Pasteurization destroys pathogenic microorganisms by rapidly heating the substance to 71.1ºC for 15 seconds, followed by rapid cooling. Pasteurization is not sterilization because not all microorganisms are sensitive to it. This method eliminated the food route of transmission of diseases such as tuberculosis of the digestive tract and Q-fever.
Tyndalization is an intermittent heat sterilization method that can be used to kill all bacteria in solutions. Since growing bacteria are easily killed by short boiling (5 times for 1 hour for 5 minutes), all that needs to be done is to allow the solution to sit for a certain time before the heat disrupts the maturation of the spores with a significant loss of their resistance to heat.
Filtration is a process used to remove germs and microscopic parts from solutions, air and other gases. The most common uses of sterilization by filtration in the laboratory are for the processing of diagnostic preparations, culture media, tissue culture media, serums, and solutions that contain serum components. Another common application of filtration is the sterilization of air and gases. Plastic or paper membrane filters, which are differentiated by pore diameter (approximately 12 to 0.22 μm) and used for mechanical separation, also serve to collect microbes from liquids for microscopic examination or cultivation directly on the filter when it is placed on a surface impregnated nutrient medium.
Ultraviolet irradiation is a type of electromagnetic wave radiation that acts on cellular nucleic acid. Microorganisms are highly sensitive to ultraviolet rays with a wavelength of 254 nm. Ultraviolet light is most widely used to destroy microorganisms in the air or on any surfaces. Other applications are cold sterilization of certain chemicals and plastics for pharmaceutical purposes, serum sterilization for cell cultures, and water disinfection. A significant disadvantage of ultraviolet irradiation as a sterilizer is its inability to penetrate materials.
Ionizing radiation in the electromagnetic spectrum has a lethal effect on microorganisms. This spectrum includes microwaves, gamma rays, x-rays and electron flow. The lethal effect of ionizing radiation occurs due to a direct effect on the target molecule, resulting in the transfer of energy into the molecule; and due to indirect action - diffusion of radicals.
Ultrasonic energy inactivates microorganisms in aqueous solutions with low frequency. The physical effect of sonication occurs due to cavitation. Ultrasonic cleaners and other devices are often used to clean instruments, but are not considered sterilants. However, combining ultrasound with chemical treatment kills microorganisms.
Chemical sterilizers
2 % glutaraldehyde As a liquid chemical sterilant, it was previously widely used to process medical and surgical materials that cannot be sterilized by heat or radiation. Glutaraldehyde is also used in the preparation of vaccines.
Disinfection.
Disinfection can be carried out using chemical methods or boiling. Boiling is an effective method of disinfecting equipment such as needles and syringes if an autoclave is not available. Pre-cleaned medical instruments should be boiled for 20 minutes. Chemical disinfection is used for heat-sensitive equipment that can be damaged by high temperatures. Chemical disinfectants such as chlorine components, ethyl and isopropyl alcohol, quaternary ammonium components and glutaraldehyde are widely used.
Chemical disinfectants.
Alcohol (ethyl and isopropyl), dissolved in water to a concentration of 60-85%, is very effective in disinfection. Alcohols have bactericidal, fungicidal and tuberculocidal effects, but do not affect spores. Ethyl alcohol has a wider spectrum of viruscidal activity than isopropyl alcohol, so it is more effective against lipophilic and hydrophilic viruses.
Solution 37% formaldeide, which is called formalin, can be used as a sterilizer, while its concentrations of 3-8% can be used as disinfectants.
Phenol in its pure form is not used as a disinfectant due to its toxicity, ability to induce the development of tumors and corrosion. Phenol derivatives, in which a functional group (chlorine, bromine, alkyl, benzyl, phenyl, amyl) replaces one of the hydrogen atoms in the aromatic ring, are widely used as disinfectants. Such substitution reduces the disadvantages of phenol. Phenol components kill microbes through inactivation of enzyme systems, precipitation of proteins and disruption of the cell wall and membrane. Concentrations of 2-5% are typically used, with lower concentrations requiring longer exposure times.
Halogens. Only chlorine and iodine are used for disinfection in laboratory practice. Because chlorine is a powerful oxidizing agent, it is believed to kill microbes through oxidation. Iodine is believed to kill microorganisms by reacting with the N-H and S-H groups of amino acids, as well as with the phenolic group of the amino acid tyrosine and the carbon-carbon double bonds of unsaturated fatty acids. Conventional treatment involves spraying a 2-5% formaldehyde solution in the presence of steam at a temperature of 60-80ºC.
Antiseptic.
Antiseptics can be found in microbiology laboratories, primarily in substances used to wash hands. When medical personnel provide emergency care to patients using substances containing antibacterial agents, this reduces the number of hospital-acquired infections. The most common chemicals found in hand washes are alcohols, chlorhexidine gluconate, iodophors, chloroxylenol and triclosan.
Traditional methods for treating waste and garbage are incineration and steam sterilization.
Burning is the method of choice for processing waste and debris. This method makes waste non-infectious and also changes its shape and size. Sterilization is an effective method of treating waste, but it does not change its form. Steam sterilization in an autoclave at 121ºC for a minimum of 15 minutes destroys all forms of microbial life, including large numbers of bacterial spores. This type of complete sterilization can also be carried out using dry heat at a temperature of 160-170ºC for 2-4 hours. However, you should ensure that dry heat is in contact with the material being sterilized. Therefore, bottles that contain liquid must be loosely sealed with stoppers or cotton swabs so that steam and heat can exchange with the air in the bottles. Biohazard containers containing waste should be sealed tightly. Sterilized biohazardous material should be sealed in appropriate labeled containers.
Steam sterilization (autoclave). Infectious debris is considered decontaminated when the number of vegetative bacteria, fungi, mycobacteria and viruses containing lipids is reduced by 6 lg times, and bacterial endospores are reduced by 4 lg times.
Nutrition of bacteria.
Nutrition. Nutrition of a bacterial cell should be understood as the process of absorption and assimilation of plastic material and energy as a result of transformative reactions . The nutritional types of prokaryotes are complex and varied. They differ depending on the method of entry of nutrients into the bacterial cell, sources of carbon and nitrogen, method of obtaining energy, and the nature of electron donors.
Transport of nutrients into the cell can be carried out by 3 mechanisms: passive diffusion, facilitated diffusion and active transport.
Passive diffusion is a nonspecific energy-dependent process carried out along a concentration gradient of substances (a substance from a medium with a higher concentration passively, according to the laws of osmosis, enters a medium with a lower concentration). By passive diffusion, a limited amount of substances, some ions, and monosaccharides, enter the bacterial cell. The rate of substance transfer during passive diffusion is insignificant and depends on the lipophilicity and size of the transported molecules.
Facilitated diffusion is an energy-independent transport of substances along a concentration gradient using permease enzymes. Permeases are specific membrane proteins that facilitate the passage of substances through the cytoplasmic membrane. The permease fixes on itself a molecule of the transported substance, together with which it crosses the cytoplasmic membrane, after which the “substance-permease” complex dissociates. The released permease is used to carry other molecules. In prokaryotes, only glycerol enters the cell by facilitated diffusion. In this case, the intracellular concentration of glycerol corresponds to that outside the cell. Facilitated diffusion is most characteristic of eukaryotic microorganisms.
Active transport- This is the energy-dependent transfer of substances into the cell against a concentration gradient using specific enzymes. The vast majority of substances (ions, carbohydrates, amino acids, lipids, etc.) enter the bacterial cell by active transport. Active transport can occur: (1) without chemical modification of the transported substance; (2) with chemical modification.