Ţesut conjunctiv numită și țesutul mediului intern. Face parte din fiecare organ și formează straturi între organe, ca și cum le-ar conecta. Țesutul conjunctiv acoperă vasele de sânge și nervii, participă la formarea scheletului uman și a scheletului organelor sale individuale și la formarea sângelui și a limfei.
Țesutul conjunctiv funcționează următoarele funcții: trofic, protector, de sustinere (mecanic) si plastic.
Trofic sau nutritiv, funcţie este acel sânge, care aparține țesutului conjunctiv, transportă nutrienți în tot corpul. În plus, prin îmbrăcarea vaselor, țesutul conjunctiv împreună cu acestea pătrunde în toate țesuturile și organele.
Funcție de protecțiețesutul conjunctiv este asociat nu numai cu proprietățile sale mecanice (oasele - formațiuni dense - protejează organele), ci și cu faptul că celulele sale au capacitatea de a fagocitoza: absorb și digeră substanțele nocive. Țesutul conjunctiv este, de asemenea, implicat în formarea corpurilor de protecție care creează imunitate (imunitate la boli).
Funcția de suportȚesutul conjunctiv este determinat în principal de substanța intercelulară.
Funcție plasticățesutul conjunctiv se exprimă prin capacitatea sa ridicată de a se regenera și de a se adapta la condițiile de mediu. Acest țesut este format din stratul germinal mijlociu al mezodermului, din așa-numitul țesut conjunctiv embrionar (mezenchim).
Țesutul conjunctiv este format din celule și substanță intercelulară, în care sunt secretate substanța fundamentală și fibrele. Spre deosebire de alte tipuri de țesut, substanța intercelulară predomină în el, în timp ce există puține celule. În diferite tipuri de țesut conjunctiv, raportul cantitativ dintre substanța intercelulară și celule este diferit.
Substanța fundamentală a țesutului conjunctiv conține multe fibre. Unele dintre ele, situate sub formă de panglici groase drepte sau oarecum contorte, nu se ramifică, constau dintr-o substanță adezivă specială și se numesc fibre de colagen sau adezive. Sunt slab extensibile, foarte durabile. Un alt tip de fibra este elastica. Sunt mai subțiri și ramificate. Aceste fibre sunt mai puțin puternice decât colagenul, dar au o elasticitate și o elasticitate mai mari (ca și cauciucul, se pot întinde și apoi reveni la forma lor inițială).
Principalele celule ale țesutului sunt fibroblastele, fibrocitele, macrofagele, mastocitele și celulele plasmatice. Poate conține celule adipoase, celule pigmentare și chiar celule albe din sânge.
Fibroblastele- tipul principal de celule ale țesutului conjunctiv. Au o formă neregulată sau în formă de fus (alungită). Miezul lor este destul de mare și de formă ovală. Fibroblastele participă la formarea substanței și fibrelor intercelulare, la vindecarea rănilor și la dezvoltarea țesutului cicatricial. Fibroblastele care și-au terminat ciclu de viață, se numesc fibrocite.
Macrofage poate exista diverse forme: rotund, alungit, neregulat. Cochilia lor este pliată, cu un număr mare de microvilozități, cu ajutorul cărora captează substanțe străine. Aceste celule au de obicei un singur nucleu, de dimensiuni mici, oval sau în formă de fasole. Macrofagele sunt principalii apărători ai corpului uman. Ei distrug microbii și neutralizează substanțele toxice (otrăvitoare).
Mastocitele Au o formă neregulată, procese scurte largi și un miez mic. Există multe boabe în citoplasmă. Mastocitele au o capacitate bine dezvoltată pentru mișcările amiboide. Ele participă la formarea substanței intercelulare și la reglarea compoziției acesteia, produc substanțe care previn coagularea sângelui și depunerea de săruri în pereții vaselor de sânge.
Celule plasmatice de formă ovală sau rotundă participă la formarea corpurilor de protecție, mai ales reacționează atunci când o proteină străină este introdusă în organism.
Celulele grase conțin grăsime în citoplasmă, împingând nucleul la periferie. Numărul lor în țesutul conjunctiv lax este variabil. Odată cu o nutriție sporită, numărul de celule adipoase crește brusc.
Celulele pigmentare- acestea sunt aceleași fibroblaste sau fibrocite, în a căror citoplasmă se află multă materie colorantă - pigment.
În funcție de funcțiile îndeplinite, care sunt determinate în mare măsură de caracteristicile fizico-chimice ale substanței intercelulare (poate fi lichid, dens și foarte dur), țesutul conjunctiv se împarte în protector-trofic și de susținere. Țesutul conjunctiv protector-trofic include: sânge, limfa, reticular sau reticular, țesut, țesut fibros lax și endoteliu. Țesutul conjunctiv de susținere include: țesut fibros dens, cartilaj și țesut osos. Pe măsură ce substanța intercelulară devine mai densă, funcția trofică a țesutului scade și funcția de susținere crește.
Sânge- Acesta este un tip de țesut conjunctiv cu o substanță intercelulară lichidă și celule specifice. Substanța intercelulară a sângelui este partea sa lichidă - plasma, care conține elementele formate (celulele) din sânge. În volum, plasma reprezintă 55-60%, iar elementele formate 40-45% din tot sângele În corpul unui adult există 4,5-5 litri de sânge.
Plasma. Plasma sanguină este formată din substanțe anorganice și organice. Substante anorganice conține aproximativ 91% (90% este apă și 1% minerale), și organic aproximativ 9%. Partea principală a materiei organice sunt proteinele - 7%. Există 3 tipuri de ele - fibrinogen, albumină și globuline. Fibrinogenul participă la coagularea sângelui, albuminele transportă substanțe slab solubile în apă (inclusiv medicamente), iar globulinele asigură formarea de corpuri de protecție. Cantitatea de globuline crește brusc în timpul bolilor infecțioase. Plasma sanguină lipsită de fibrinogen se numește ser sanguin. Este folosit în scopuri terapeutice sau profilactice pentru a crea imunitate (imunizare pasivă), iar seruri terapeutice sunt preparate. Plasma sanguină conține și substanțe organice de natură neproteică (uree, grăsimi, aminoacizi etc.), deși în cantități foarte mici.
Elemente formate din sânge. Există trei tipuri de ele: globule roșii - eritrocite, globule albe - leucocite și trombocite - trombocite.
Globule roșii(eritros - roșu, citos - celulă) sunt celule specifice foarte diferențiate care, în timpul dezvoltării, și-au pierdut nucleul, mitocondriile, aparatul reticular și centrul celular (Fig. 6). La broaște, pești și păsări, celulele roșii din sânge conțin nuclei (Fig. 7). În citoplasma eritrocitelor există proteine complexe- hemoglobina, cu ajutorul căreia se realizează schimbul de gaze în organism: oxigenul este transferat de la plămâni la țesuturi și de la țesuturi la plămâni - dioxid de carbon. Membrana celulelor roșii din sânge este foarte subțire, iar prin ea se fac schimb de gaze. Globulul roșu are forma unui disc biconcav, care își mărește suprafața, facilitând un contact mai bun al hemoglobinei cu gazele transportate. Absența unui nucleu în celulă pare să promoveze, de asemenea, o absorbție mai mare de oxigen.
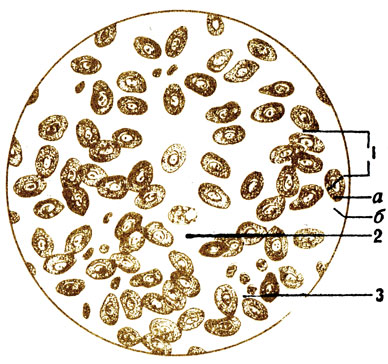
Dimensiunea globulelor roșii este mică, doar 7-8 microni, așa că trec destul de ușor prin cele mai subțiri vase de sânge - capilare. 1 mm 3 de sânge conține 4,5-5,0 milioane, iar în total sunt 25-28 de trilioane de globule roșii. Dacă ar fi posibil să le plasăm una lângă alta, s-ar crea un lanț care ar fi suficient pentru a înconjura globul de 3 ori de-a lungul ecuatorului. Suprafata totala globule roșii care circulă în sânge, mai mult de 1/4 hectar. La bărbați, numărul de globule roșii este puțin mai mare decât la femei; la copii - mai mult decât la adulți; Locuitorii din zonele montane înalte, unde există mai puțin oxigen în aer, au mai mult decât locuitorii de câmpie. Chiar și cu o ședere de scurtă durată (1-2 luni) în zonele muntoase, numărul de celule roșii din sânge crește, ceea ce este important pentru desfășurarea antrenamentului sportiv acolo. Odată cu creșterea activității musculare, există și mai mulți dintre ei din cauza unei cereri crescute de oxigen. Globulele roșii nu au capacitatea de a mișcări independente, se deplasează prin vasele de sânge odată cu fluxul de sânge. Cu toate acestea, sunt foarte elastice atunci când se mișcă în capilare, se vede clar cum se alungesc, se aplatizează, schimbându-și forma. Durata de viață a globulelor roșii este de 80-120 de zile. Celulele roșii din sânge se descompun în splină și se formează în măduva osoasă roșie. Conform datelor disponibile, 1/100 dintre globulele roșii sunt distruse în fiecare zi, adică în puțin mai mult de 3 luni, toate globulele roșii sunt reînnoite.
Leucocite- acestea sunt celule cu nucleu. Ele sunt mai mari decât globulele roșii (până la 10 microni), capabile de mișcare independentă a ameboidului și pot ieși din capilare în țesutul subiacent.
În funcție de natura citoplasmei, prezența incluziunilor în ea sub formă de boabe de proteine, pigment și, de asemenea, în funcție de forma nucleului, leucocitele sunt împărțite în granulare și negranulare. Primele au granularitate în citoplasmă și un nucleu segmentat, împărțit în părți separate. În funcție de tipul de granularitate și de relația acestuia cu coloranții, se disting neutrofile, bazofile și eozinofile, care se găsesc în anumite rapoarte cantitative în sânge. Modificarea acestui raport în diferite boli determină nu numai natura lor, ci și rezultatul lor.
Formele negranulare ale leucocitelor nu conțin incluziuni în citoplasmă; nucleul lor nu este împărțit în părți, are o formă rotundă și este cel mai adesea situat în centrul celulei. Leucocitele non-granulare includ limfocitele și monocitele.
1 mm 3 de sânge conține 6 - 8 mii de leucocite. Numărul acestora poate crește după masă, în timpul bolilor infecțioase și mai ales după o activitate musculară intensă. Durata de viață a leucocitelor variază: de la câteva zile până la 2 - 3 luni. Când o infecție intră în organism, aceștia mor în număr semnificativ în lupta împotriva ei. Formele granulare de leucocite sunt produse în măduva osoasă roșie, iar limfocitele sunt produse în splină și ganglionii limfatici.
Funcția principală a leucocitelor este de protecție. Păzesc sănătatea organismului, ajutându-l să lupte împotriva diferitelor boli. Ei îndeplinesc o funcție de protecție participând la fagocitoză și la formarea corpurilor de protecție. În plus, leucocitele produc enzime care reglează coagularea sângelui și permeabilitatea vasculară. In sfarsit, forme separate limfocitele pot forma celule diverse tipurițesut conjunctiv (fibroblaste, macrofage, celule musculare netede), care este important în procesele de recuperare.
Trombocitele, sau trombocitele din sânge, sunt corpuri rotunde sau ovale, care măsoară doar 1 - 2 microni. Nu conțin un nucleu. Există 200 - 300 mii dintre ele în 1 mm 3 de sânge Durata de viață a trombocitelor este de 5 - 8 zile. Trombocitele din sânge iau parte la coagularea sângelui.
Limfa, ca și sângele, este format dintr-o parte lichidă - limfoplasmă - și elemente formate. Spre deosebire de plasma sanguină, conține mai puține proteine, dar mai multe produse metabolice. Dintre elementele formate, predomină limfocitele, eritrocitele sunt absente.
Țesut reticular constă din celule de formă neregulată. În contact unul cu altul, formează un fel de rețea. În buclele acestei rețele există o substanță intercelulară care conține un număr mare de fibre de reticulină care împletesc suprafața celulelor. Organele hematopoietice (măduvă osoasă, splină, ganglioni limfatici) sunt construite din țesut reticular.
Țesut conjunctiv fibros lax- acesta este țesutul în care sunt exprimate cel mai clar toate elementele structurale ale țesutului conjunctiv: substanță intercelulară, fibre și celule (Fig. 8). Acoperă vasele de sânge și nervii, formează țesut subcutanat și participă la structura aproape tuturor organelor.
Țesut endotelial(endoteliul) are legătură cu țesutul conjunctiv doar la origine, în timp ce ca structură seamănă cu țesutul epitelial. Celulele sale sunt plate și situate pe membrana bazală. Există puțină substanță intercelulară în acest țesut. Endoteliul căptușește suprafața interioară a vaselor de sânge, oferindu-i un aspect neted, strălucitor; Metabolismul are loc prin celulele endoteliale ale capilarelor; Ele îndeplinesc și o funcție de protecție.
Țesut conjunctiv fibros dens are trăsătură caracteristică- este dominata de fibrele de colagen, care sunt colectate in fascicule orientate in functie de directia fortelor de tractiune. Există puține celule aici (în principal fibroblaste) și sunt situate între fasciculele de fibre. Din acest tesut sunt construite ligamentele, tendoanele, fascia, septurile intermusculare, periostul, pericondrul etc. (Fig. 9).

În tendoane și ligamente, mănunchiurile de fibre de colagen sunt situate în paralel, în fascie, aponevroze, septuri intermusculare - în straturi unul deasupra celuilalt (cu cât fascia este mai groasă, cu atât mai multe straturi), iar direcția fibrelor în diferite straturi este diferită: în unele - în unghi drept, în altele - sub ascuțit, ceea ce conferă acestor formațiuni o rezistență deosebită. Dacă țesutul conjunctiv fibros dens este dominat de fibre elastice, se numește țesut conjunctiv elastic. Prezența fibrelor elastice ajută un organ sau o parte a corpului să revină la poziția inițială după schimbarea formei.
Țesutul cartilajului(cartilajul) prin proprietăți fizice și chimice și caracteristici funcționale diferă puternic de alte tipuri de țesut conjunctiv. Substanță intercelulară Este destul de dens și, prin urmare, îndeplinește în principal funcții de susținere și de protecție (mecanice). Există trei tipuri de cartilaj: hialin, sau vitros, colagen-fibros și elastic. Țesutul cartilajului nu are vase de sânge. Metabolizarea (nutriția și îndepărtarea deșeurilor) se realizează prin vasele membranei țesutului conjunctiv care acoperă exteriorul cartilajului (pericondru). Nutrienții din vasele pericondului pătrund în substanța intercelulară a cartilajului. Cartilajul care acoperă suprafețele articulare ale oaselor primește nutrienți din lichidul sinovial care umple cavitatea articulară sau din vasele osoase din apropiere. Creșterea cartilajului are loc datorită pericondului.
Cartilajul hialin are cea mai mare distribuție în corpul uman. Substanța sa intercelulară este translucidă, alb-albăstruie. Celulele cartilajului sunt situate în cavități speciale înconjurate de o capsulă, care este mai densă decât substanța intercelulară. Cartilajul hialin formează capetele anterioare ale coastelor, cartilajul traheei, bronhiile, majoritatea cartilajul laringelui și acoperă suprafețele articulare ale oaselor. În perioada embrionară, o parte semnificativă a scheletului este formată din cartilaj hialin. La bătrânețe, varul se poate depune în cartilajul hialin (Fig. 10).

Fibrocartilajul de colagen mai puțin elastic, dar mai durabil. Substanța sa intercelulară conține un număr mare de mănunchiuri de fibre de colagen situate mai mult sau mai puțin paralel. Celulele sunt situate între fasciculele de fibre. Discurile intervertebrale și cartilajul care leagă oasele pubiene sunt construite din acest cartilaj (Fig. 11).
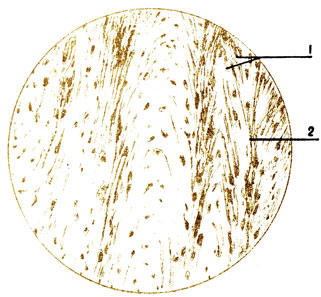
Cartilaj elastic mai puțin durabilă, dar foarte elastică, calcificarea nu apare niciodată în ea. În substanța intercelulară a cartilajului există multe fibre elastice care se împletesc între ele, formând o rețea densă. Celulele sale seamănă cu forma unei flăcări de lumânare și sunt situate la 2-3 în capsule între fibre. Cartilajul elastic este situat acolo unde nu este nevoie de multă rezistență fortele curente. Din el sunt construite auriculul, epiglota, peretele canalului auditiv extern și tubul auditiv (Fig. 12).

Țesut osos este cel mai dens dintre toate tipurile de țesut conjunctiv. Substanța sa intercelulară constă din fibre, care sunt adesea conectate în mănunchiuri, și substanța fundamentală, în care un procent mare compuși anorganici, predominant saruri de calciu, deci functia de sustinere a osului este cea mai pronuntata. Cu toate acestea, în ciuda densității țesut osos- un sistem viu, suferă modificări de-a lungul vieții unei persoane, însoțite de reînnoirea elementelor sale constitutive, ceea ce îi asigură adaptabilitatea la condiții mediu(Fig. 13).

Restructurarea țesutului osos depinde de vârstă, nutriție, funcția organelor de secreție internă și alți factori. Cele mai pronunțate modificări ale țesutului osos apar în timpul activității musculare: nu numai structura internă a țesutului osos se modifică, ci și forma organelor - oasele pe care le formează.
Există trei tipuri de elemente celulare în țesutul osos: osteocite, osteoblaste și osteoclaste.
Osteocitul(oss - os, cytos - celula) - celula principală a țesutului osos - are o formă neregulată, un număr mare de procese lungi cu care contactează celulele învecinate. Aceste celule osoase se află în cavități speciale.
Osteoblastele- creatori, creatori de tesut osos. Ele sunt situate acolo unde are loc procesul de formare a oaselor. Forma lor poate fi cubică, piramidală sau unghiulară. Pe măsură ce se formează țesutul osos, osteoblastele se transformă în osteocite.
Osteoclaste- celule multinucleate. Sunt mai mari decât osteocitele și osteoblastele. Fiecare osteoclast poate avea până la 50 de nuclei. O mică depresiune se formează în punctul de contact al osteoclastei cu substanța osoasă. Osteoclastele se află în astfel de depresiuni, golfuri. Aceste celule distrug țesutul osos, în locul lui se formează unul nou. Ambele procese au loc continuu în țesutul osos - atât procesul de distrugere, cât și procesul de creație, asigurând reconstrucția osului.
Există două tipuri de țesut osos: cu fibre grosiere și cu fibre fine sau lamelare.
Țesut osos fibros aspru Apare într-o măsură mai mare la făt; la adulți se găsește numai în locurile de atașare a tendoanelor musculare la oase, în suturile dintre oasele craniului. În substanța intercelulară a țesutului osos grosier fibros, fasciculele de fibre sunt groase, situate paralel, în unghi sau sub formă de rețea. Osteocitele au o formă aplatizată.
Fibră fină sau lamelar, țesut osos cel mai diferenţiat. Unitatea sa structurală și funcțională este placa osoasă. În substanța intercelulară a plăcii, fibrele sunt subțiri și orientate în anumite direcții paralele între ele. Osteocitele se află între lamine sau în interiorul laminelor. Plăcile sunt aranjate astfel încât fibrele din două plăci adiacente să curgă aproape în unghi drept, ceea ce asigură o rezistență și o elasticitate deosebite țesutului osos. Aproape toate oasele scheletului uman adult sunt construite din țesut osos cu fibre fine.
Am scris deja despre termenii de bază și componentele generale ale CT într-un articol anterior despre caracteristicile țesutului conjunctiv. Acum să caracterizăm câteva grupe de țesut conjunctiv(SF).
Loose ST- acesta este țesutul principal și principal când vine vorba de țesut conjunctiv (Fig. 10). Fibrele elastice (1), colagen (2), precum și unele celule sunt incluse în componenta sa amorfă. Celula cea mai de bază este fibroblastul (latina fibra - fibre, greacă blastos - germen sau germen). Fibroblastul este capabil să sintetizeze elementele constitutive ale componentei amorfe și să formeze fibre. Adică, funcția reală a unei celule fibroblastice este capacitatea de a sintetiza substanță intercelulară. Fibroblastele (3) cu un nucleu mare (a) în endoplasma lor (b) și ectoplasma (c) conțin un reticul endoplasmatic destul de impresionant, în care sunt sintetizate proteine precum colagenul și elastina. Aceste proteine sunt constructorii fibrelor corespunzătoare. O altă celulă importantă a CT liber este histiocitul (4). Microorganismele ar trebui să se ferească de aceste celule, deoarece odată ce intră în substanța intercelulară, le fagocită sau, pur și simplu, le mănâncă. În sfârșit, în imaginea color I puteți vedea o altă celulă importantă a CT-ului - aceasta este un mastocit care stochează doi compuși biologic activi: heparină și histamina; Heparina este o substanță care previne coagularea sângelui. Histamina este o substanță care participă la diferite reacții alergice și procese inflamatorii. Datorită eliberării histaminei din mastocite, se observă simptome precum înroșirea pielii, urticarie, mâncărime, vezicule, arsuri și șoc anafilactic. 
Imaginea I. Țesut conjunctiv lax
CT liber însoțește toate vasele. Aorta este acoperită cu o pernă întreagă - adventiția, iar cele mai mici capilare sunt înconjurate de o rețea foarte subțire de fibre și celule. Vasele sunt protejate, întărite și, parcă, bazate pe acest tip de ST. Aceasta înseamnă că CT liber este localizat oriunde sunt prezente vase. Din acest motiv, merită evidențiat ca țesut conjunctiv principal și principal.
Un medic practic în munca sa zilnică întâlnește foarte des o manifestare a țesutului conjunctiv lax - edem. Glicozaminoglicanii, care formează o componentă amorfă, sunt capabili să rețină apa, ceea ce o fac ori de câte ori este posibil. Și această oportunitate apare în anumite procese patologice: insuficiență cardiacă, stagnare limfatică, boli de rinichi, inflamație etc. În acest caz, lichidul se acumulează în țesutul conjunctiv, care se umflă, făcând pielea umflată. Uneori, umflarea sub ochi poate fi simptomul inițial al unei boli precum glomerulonefrita - o inflamație imună a rinichilor. ![]()
Dens ST conține un număr foarte mic de componente celulare și o componentă amorfă a substanței intercelulare cea mai mare parte a țesutului conjunctiv dens este alcătuită din fibre; Există două forme de CT dens. ST dens neformat(Fig. 11) prezintă o dezordine completă a fibrelor (4). Fibrele sale se împletesc după bunul plac; fibroblastele (5) pot fi orientate în orice direcție. Acest tip de ST este implicat în formarea pielii, este situat sub epidermă (1) și stratul de ST liber (2) care înconjoară vasele (3), și conferă dermului o anumită rezistență. Dar în asta nu se poate compara cu puterea dens decorat ST(Fig. 12), care constă din mănunchiuri strict ordonate (5), care au, la rândul lor, o anumită direcție a fibrelor de colagen (3) și/sau elastice (4). Țesutul conjunctiv format face parte din tendoane, ligamente, tunica albuginea globului ocular, fascia, durei mater, aponevroze și alte formațiuni anatomice. Fibrele sunt înfășurate (1) și „stratificate” (7) cu CT liber care conține vase (2) și alte elemente (6). Datorită paralelismului fibrelor tendonului, acestea capătă rezistență și rigiditate ridicate. 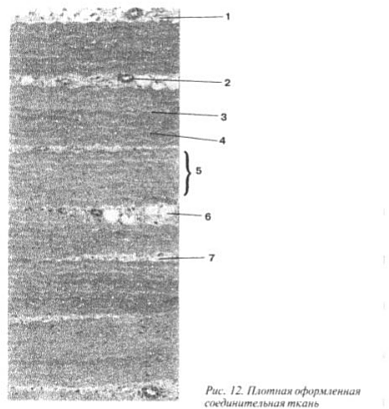
Țesut adipos(Fig. 13) este distribuit aproape peste tot în piele, retroperitoneu, epiploon și mezenter. Celulele țesutului adipos sunt numite lipocite (1 și imaginea II). Ele sunt foarte dens localizate, permițând doar vaselor mici, cum ar fi capilarele (2) să treacă între ele, iar odată cu ele fibroblastele omniprezente cu fibre individuale (3). Lipocitele sunt aproape complet lipsite de citoplasmă și pline cu picături mari de grăsime solide. Nucleul se dovedește a fi deplasat în lateral, în ciuda faptului că este un regulator al celulei. 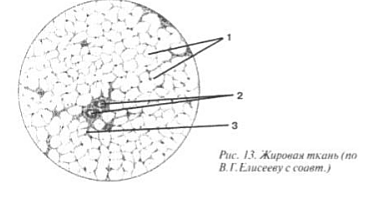

Poza II. Țesut adipos
Țesutul adipos este cea mai importantă sursă de energie a organismului. La urma urmei, atunci când grăsimea este descompusă, aceasta este eliberată mult mai mult decât atunci când se utilizează carbohidrați și proteine. În plus, în acest caz se formează o cantitate semnificativă de apă, astfel încât țesutul adipos se dovedește simultan a fi un rezervor de rezervă de apă legată (nu degeaba această variantă specială de ST se găsește în cocoașele cămilelor, care încet. descompune grăsimea când traversezi deșerturile fierbinți). Mai este o funcție. La copiii nou-născuți, un subtip special a fost găsit în piele - țesutul adipos maro. Conține un număr foarte mare de mitocondrii și datorită acestui fapt este cea mai importantă sursă de căldură pentru copilul care se naște. 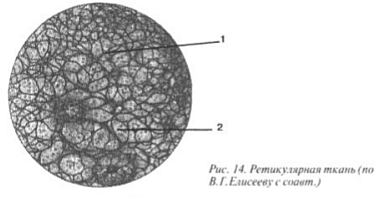
Țesut reticular, situat în organele sistemului limfatic: în măduva osoasă roșie, ganglionii limfatici, timusul (glanda timus), splina, este format din celule multiprocesate numite reticulocite. Cuvântul latin reticulum înseamnă „rețea”, care se potrivește perfect cu această țesătură (Fig. 14). Reticulocitele, ca și fibroblastele, sintetizează fibre (1), numite reticulare (o variantă a colagenului). Acest tip de CT asigură hematopoieza, adică aproape toate celulele sanguine (2) suferă dezvoltare, într-un fel de hamac format din țesut reticular(figura III). 
Imaginea III. Țesut reticular
Ultima subspecie a ST în sine este țesut pigmentar(Fig. 15) se găsește în aproape tot ceea ce este intens colorat. Exemplele includ părul, retina globului ocular și pielea bronzată. Țesătură pigmentată reprezentate de melanocite, celule umplute cu granule ale principalului pigment animal - melanina (1). Au formă de stea: din nucleul situat în centru, citoplasma diverge în petale (2). 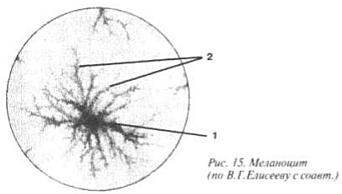
Aceste celule pot da naștere unei tumori maligne - melanom. Boala in în ultima vreme a devenit mult mai comună decât înainte. În ultimul deceniu, incidența cancerului de piele a crescut foarte puternic, se crede că acest lucru se datorează modificărilor grosimii stratului de ozon, care protejează planeta noastră cu un strat gros de efectele mortale ale radiațiilor ultraviolete. Peste poli a scăzut cu 40-60% oamenii de știință chiar vorbesc despre „; găuri de ozon" Și, ca urmare, la oamenii care se prăjesc sub soare, melanocitele semnelor de naștere sunt primele care răspund la efectul mutagen al razelor ultraviolete. Divându-se continuu, ele dau naștere la creșterea tumorii. Din păcate, melanomul progresează rapid și de obicei metastazează devreme.
Țesutul cartilajului(Fig. 16) - țesut care are o componentă amorfă concentrată foarte „de înaltă calitate” în substanța sa intercelulară. Glicozamina și proteoglicanii îl fac dens și elastic, ca jeleul. De această dată, atât componentele amorfe, cât și cele fibroase ale substanței intercelulare sunt sintetizate nu de fibroblaste, ci de celule tinere ale țesutului cartilaginos, care se numesc condroblasti (2). Cartilajul nu are vase de sânge. Nutriția sa provine din capilarele stratului cel mai superficial - pericondrul (1), unde se află de fapt condroblastele. Numai că s-au „maturat”, ele sunt acoperite cu o capsulă specială (5) și trec în substanța amorfă a cartilajului propriu-zis (3), după care se numesc condrocite (4). Mai mult, substanța intercelulară este atât de densă încât atunci când un condrocit se divide (6), celulele sale fiice nu se pot separa, rămânând împreună în cavități mici (7). 
Țesutul cartilaginos formează trei variante de cartilaj. Primul, cartilajul hialin, are foarte puține fibre și se găsește la joncțiunea coastelor cu sternul, în trahee, în bronhii și laringe, pe suprafețele articulare ale oaselor. Al doilea tip de cartilaj este elastic (figura IV), conținând multe fibre elastice, este situat în auriculă și laringe. Cartilajul fibros, care conține în principal fibre de colagen, formează simfiza pubiană și discurile intervertebrale. 
Imaginea IV. Cartilaj elastic
Țesut osos contine trei tipuri de celule. Osteoblastele tinere sunt similare ca funcție cu fibroblastele și condroblastele. Ele formează substanța intercelulară a osului, situată în stratul cel mai superficial bogat în vase de sânge - periostul. Pe măsură ce îmbătrânesc, osteoblastele sunt încorporate în osul însuși, devenind osteocite. În perioada embrionară, corpul uman nu are oase ca atare. Embrionul are „blankuri” cartilaginoase, modele ale viitoarelor oase. Dar începe treptat osificarea, necesitând distrugerea cartilajului și formarea țesutului osos real. Distrugătorii de aici sunt celulele - osteoclaste. Ei zdrobesc cartilajul, făcând loc osteoblastelor și muncii lor. Apropo, osul îmbătrânit este înlocuit în mod constant cu alții noi și, din nou, osteoclastele sunt responsabile pentru distrugerea osului uzat.
Substanța intercelulară a țesutului osos conține o cantitate mică de substanțe organice (30%), în special fibre de colagen, care sunt strict orientate în substanța compactă a osului (figura V) și aleatoare în cea spongioasă. Componenta amorfă, după ce „și-a dat seama” că este „de prisos la această sărbătoare a vieții”, este practic absentă. În schimb, există diverse săruri anorganice, citrați, cristale de hidroxiapatită și mai mult de 30 de microelemente. Dacă încălzești un os în foc, tot colagenul va arde; forma se va păstra, dar doar atingeți-o cu degetul și osul se va sfărâma. Și după o noapte într-o soluție de niște acid, în care se dizolvă toate sărurile anorganice, osul poate fi tăiat ca untul cu un cuțit, adică își va pierde rezistența, dar pe gât (mulțumită fibrelor rămase) se va tăia. fii legat ca o cravată de pionier. 
Imaginea V. Țesutul osos
Ultimul, dar nu cel din urmă grup de țesut conjunctiv, este sânge. Studierea lui necesită o cantitate imensă de informații. Prin urmare, nu vom diminua semnificația sângelui prin descrierea lui aici, ci vom lăsa acest subiect pentru o analiză separată.
Țesuturile conjunctive sunt țesuturi larg răspândite de origine mezenchimală. Ele asigură menținerea integrității altor țesuturi, formează stroma organelor, conțin vase de sânge și limfatice și participă la furnizarea trofică a tuturor țesuturilor și organelor. Printre țesuturile conjunctive există țesături fibroase(laxe și dense) și țesuturi cu proprietăți speciale (de exemplu, adipoase, reticulare). Diferite țesuturi conjunctive diferă în ceea ce privește proprietățile fizice. În majoritatea organelor și între ele, țesutul conjunctiv lax acționează ca un pachet de parenchim, organizând un pat pentru elementele sale histologice (stroma de organ). Pentru multe tipuri de celule, organe și țesuturi, mediul țesutului conjunctiv joacă nu numai un rol de susținere pasiv, ci are și o semnificație organizatorică și informațională. Țesuturile conjunctive dense oferă rezistență pielii, formează capsule de organe și le permit să reziste la sarcini mecanice semnificative (de exemplu, ligamente și tendoane). Țesutul conjunctiv are o mare importanță metabolică nu numai pentru că conține vasele de sânge. Astfel, țesutul adipos alb stochează grăsimea ca sursă de energie, iar țesutul adipos maro este implicat în reglarea temperaturii corpului la nou-născuți. Un număr de celule de țesut conjunctiv
aparține funcției de protecție împotriva microorganismelor patogene. Țesutul conjunctiv este o trambulină pentru reacțiile inflamatorii și imune.
MATRICE EXTRACELULARĂ
Toate țesuturile conjunctive fibroase conțin o cantitate semnificativă de substanță intercelulară. Compoziția chimică, structura și volumul matricei extracelulare determină proprietăți fizice fiecare tip de țesut conjunctiv. Matricea extracelulară constă dintr-o substanță fundamentală și diferite tipuri de fibre încorporate în ea.
Substanța principală
Substanța principală este un material amorf cu proprietăți de gel. Lichidul tisular se leagă de componentele substanței principale, formând un mediu pentru trecerea moleculelor prin țesutul conjunctiv și pentru schimbul de substanțe cu sângele. Substanța principală conține glicozaminoglicani, proteoglicani și glicoproteine. Moleculele substanței principale sunt strâns legate de fibrele matricei extracelulare și interacționează cu diferite celule.
Glicozaminoglicani
Glicozaminoglicanii sunt polizaharide construite din unități repetate de dizaharide, dintre care una este de obicei acid uronic, iar cealaltă este un amino zahăr (N-acetilglucozamină, N-acetilgalactozamină). Glicozaminoglicanii sunt compuși acizi (încărcați negativ), deoarece unitățile lor dizaharidice conțin multe grupări hidroxil, carboxil și sulfat. Moleculele de glicozaminoglican sunt hidrofile și leagă un număr mare de molecule de apă și ioni (în special Na +) care intră în lichidul extracelular. Moleculele mari de glicozaminoglican legate între ele formează un gel prin care difuzează metaboliții. Sunt cunoscute următoarele tipuri de glicozaminoglicani: acid hialuronic, sulfat de condroitină, sulfat de dermatan, sulfat de keratan, sulfat de heparan și heparină (sulfat de heparină).
Proteoglicani
Proteoglicanul este o proteină centrală fibroasă cu glicozaminoglicani atașați covalent. Așa se formează macromoleculele mari, care conțin 90-95% carbohidrați. Proteoglicanii se pot lega necovalent de lanțurile de acid hialuronic, formând complexe și mai mari (de exemplu, proteoglicanii cartilajului).
Glicoproteine
Glicoproteinele constau din lanțuri polipeptidice conectate la polizaharide ramificate și conectează celulele la matricea extracelulară. Exista glicoproteine care formeaza structuri fibroase (fibronectina si fibrilina), precum si o serie de proteine nefibroase (laminina, tenascina si entactina).
Fibronectină găsit în membrana bazală, în jurul fibrelor și celulelor de colagen. O cantitate mare de fibronectină se găsește în plasma sanguină. Fibronectina este implicată în adeziunea celulară prin legarea acestora de componente ale matricei extracelulare, în special de colagen și glicozaminoglicani. În timpul embriogenezei și vindecării rănilor, fibronectina formează căi pentru migrarea celulelor. Receptorul de fibronectină este o integrină încorporată în membrana celulara. În interiorul celulei, integrina interacționează cu microfilamentele de actină ale citoscheletului perimembranar, iar în exterior se conectează cu fibronectina. La rândul său, fibronectina formează legături cu colagenul și glicozaminoglicanii. Aceasta stabilește continuitatea structurală între citoschelet și matricea extracelulară (Fig. 6-18 ).
Fibrilina(M r ≈350 kD) formează microfibrile cu diametrul de 8–12 μm, întărind legăturile dintre componentele extracelulare; face parte din fibre elastice, asigurându-le structura ordonată. Mutațiile genei fibrilinei duc la dezvoltarea sindromului Marfana.
În matricea extracelulară sunt scufundate diferite tipuri de fibre: fibre de colagen și reticulină construite din colagen și fibre elastice (incluzând elastina și fibrilina).
Fibre de colagen
Fibrele de colagen sunt componenta principală a majorității țesuturilor conjunctive, iar colagenul este proteina cea mai abundentă.
Colageni. Molecula de colagen este o spirală cu trei lanțuri pro-α. Lungimea unei astfel de spirale este de 300 nm, diametrul este de 1,5 nm. Toate tipurile de colagen conțin regiuni cu o secvență repetată de trei aminoacizi cu glicina în poziția a treia. Primul aminoacid din această secvență poate fi oricare, al doilea poate fi prolină, hidroxiprolină sau lizină. Datorită abundenței de legături încrucișate între reziduurile de lizină, fibrele de colagen au o rezistență ridicată. Sunt cunoscute cel puțin 13 tipuri de colagen (Tabel. 6-2 ).
Orez. 6-18. Comunicarea celulară mediată de fibronectină cu fibrele de colagen. Domeniul citoplasmatic al integrinei interacționează cu microfilamentele din apropierea membranei (actina), iar domeniul extracelular interacționează cu fibronectina, care la rândul ei este asociată cu fibrele de colagen. Molecula de fibronectină conține, de asemenea, domenii pentru legarea la heparină și fibrină.
Cele mai frecvente colageni sunt primele cinci tipuri, restul sunt relativ rare și în cantități mici. Colagenii de tip I, II și III formează fibrile cu o periodicitate de 65 nm. Colagenul de tip I este unul dintre cele mai comune, sintetizat de fibroblaste, osteoblaste și SMC. Colagenul de tip II este sintetizat de condroblaste, condrocite, neuroni retinieni și este prezent în corpul vitros. Colagenul de tip III face parte din fibrele reticulinei, care formează o rețea delicată de susținere în unele organe și țesuturi (de exemplu, în ficat, măduvă osoasă, organe limfoide) și este sintetizat de fibroblaste și SMC. Colagenul de tip IV formează baza reticulata a membranelor bazale și este sintetizat de majoritatea celulelor epiteliale. Colagenul de tip V este prezent în vasele de sânge, în jurul SMC și în matricea osoasă și este sintetizat de către SMC și osteoblaste. Mutațiile genelor de colagen duc la dezvoltarea diferitelor boli (osteogeneză imperfectă, acondrogeneza și achondroplazie, sindroame Ehlers–Danlos).
Formarea fibrelor de colagen(orez. 6-19 ). În stadiul intracelular, sinteza și asamblarea lanțurilor pro-α are loc în reticulul endoplasmatic granular. Mai departe în rezervoarele complexului Golgi Are loc hidroxilarea și glicozilarea polipeptidelor, ducând la formarea de elice a trei lanțuri pro-α (procolagen). Fiecare lanț pro-α conține secțiuni terminale - peptide de înregistrare. Aceste peptide previn asocierea intracelulară a elicelor de procolagen în fibrile de colagen. Moleculele de procolagen se acumulează în granule secretoare și sunt eliberate în spațiul extracelular. În afara celulei, peptidele de înregistrare ale moleculei de procolagen sunt scindate folosind procolagen peptidază (molecula rezultată este uneori numită tropocolagen). Această pierdere nu are loc la tipurile de colagen IV și V, ceea ce explică absența fibrilelor construite din colagen de acest tip. În continuare, are loc asamblarea fibrilelor de colagen (Fig. 6-20 ). Moleculele de tropocolagen au capete și coadă și se combină spontan în lanțuri paralele, aliniându-se secvenţial: „cap la cap” și „parte la alta”. În acest caz, între capetele capului și coada rămân spații de 35 nm lățime.
Tabelul 6-2. Distribuția colagenului în țesuturi și organe
| Tip | Țesuturi și organe |
| eu | Piele, tendoane, oase, cornee, placenta, artere, ficat, dentina, tumori |
| II | Cartilaj, discuri intervertebrale, corp vitros, cornee |
| III | Plămâni, artere, uter, fibre de reticulină în ficat și organe hematopoietice |
| IV | Membrane bazale |
| V | Placentă, piele, vase de sânge, țesut muscular neted, rabdomiosarcom |
| VI | Vase de sânge, ligamente, piele, uter, plămâni, rinichi |
| VII | Amnion, piele, esofag, cornee |
| IX | |
| X | Cartilaj |
| XI | Cartilaj, discuri intervertebrale, vitros |
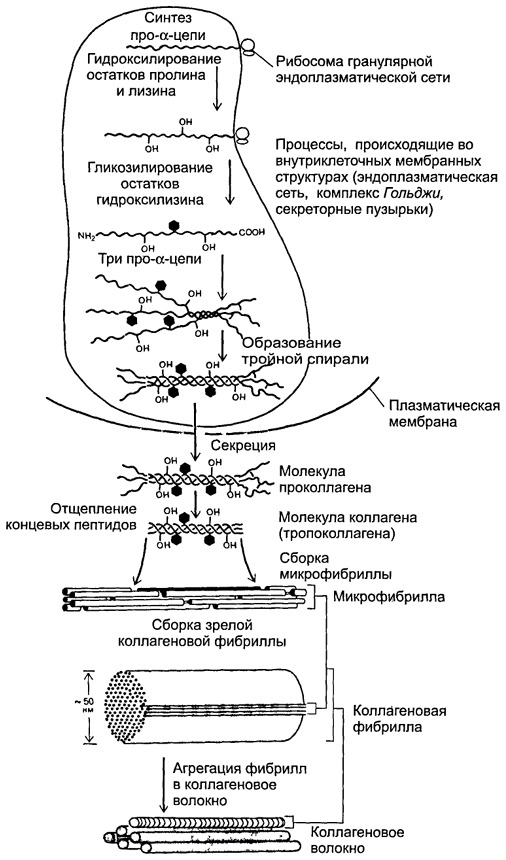
Orez. 6-19. Formarea fibrei de colagen. Sinteza lanțurilor α polipeptidice, hidroxilarea și glicozilarea acestora, precum și formarea unui helix de trei lanțuri α au loc în reticulul endoplasmatic granular și complex. Golgi. Moleculele de procolagen se acumulează în veziculele secretoare și sunt eliberate în spațiul extracelular. În afara celulei, peptidele terminale sunt separate de polipeptide și se formează fibrile de colagen.
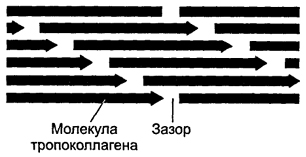
Orez. 6-20. Ambalarea moleculelor de tropocolagen polarizate în timpul formării fibrei de colagen. Atunci când moleculele sunt combinate secvențial într-un lanț, se menține un spațiu de 35 nm lățime între coada și capetele moleculelor învecinate. Fiecare moleculă a unui lanț este deplasată față de molecula lanțului vecin cu un sfert din lungimea sa. Așa se formează o fibrilă de colagen, care are o striație transversală cu o periodicitate de 65 nm.
Fiecare moleculă de tropocolagen din lanț este deplasată față de molecula din lanțul vecin cu un sfert din lungimea sa, motiv pentru alternarea dungilor întunecate și deschise cu o periodicitate de 65 nm. Fibrilele mai groase se formează prin atașarea unor noi molecule de tropocolagen. Enzima lisil oxidaza care conține cupru este implicată în formarea legăturilor încrucișate între moleculele de tropocolagen. Interacțiunea colagenului cu glicozaminoglicanii și fibronectina reglează asamblarea fibrilelor. Fibra de colagen este formată din multe fibrile de colagen conectate în paralel prin glicoproteine. Mănunchiurile de fibre de colagen sunt o colecție de fibre de colagen scufundate în matricea intercelulară.
- Vitamina C– unul dintre cofactorii necesari pentru formarea legăturilor încrucișate între lanțurile polipeptidice. Deficitul de vitamina C duce la formarea de lanțuri pro-α slab hidroxilate, care nu sunt capabile să formeze elice triple și fibrile de colagen. Ca urmare, apar defecte ale oaselor, dinților și pereților vasculari.
Interacțiunea colagenului cu celulele(Masă 6-3 ). Fibrele de colagen înconjoară multe celule. Unele celule (de exemplu, hepatocite și SMC) au receptori de colagen și se leagă direct de ei. Alte celule se leagă de colagen prin proteine intermediare de legare speciale (de obicei fibronectina, mai rar condronectină și laminină).
Fibre elastice
Structuri elastice(fibre și membrane) au proprietăți elastice pronunțate. Fibrele elastice sunt prezente în cartilajele elastice, piele, plămâni și vasele de sânge. Membranele elastice fenestrate sunt caracteristice arterelor mari.
Tabelul 6-3. Legarea moleculelor dintre colagen și diferite tipuri de celule
Fibră elastică(fir de ramificare cu un diametru de 0,2–1,0 μm) constă din fibrilină și elastină amorfă (Fig. 6-21 ). Elastina glicoproteină, ca și colagenul, conține multă glicină și prolină. Fibroblastele și SMC-urile sintetizează elastina și fibrilina în cisternele reticulului endoplasmatic granular. În complex Golgi polipeptidele sunt ambalate în
 |
| Orez. 6-21. Fibră elastică. O. Datorită stabilirii legăturilor intermoleculare între polipeptidele de elastină, se formează o rețea elastică de molecule care îi reface forma după deformare. B. Microfibrilele de fibrilină, care organizează elastina, sunt situate atât în exteriorul, cât și în interiorul fibrei elastice. |
Fibre de reticulină
Fibrele de reticulină sunt filamente subțiri cu un diametru de 0,5–2,0 μm, constând din colagen de tip III asociat cu glicoproteine și proteoglicani. Cu defecte ale genei care codifică lanțul α1 al colagenului de tip III, formarea fibrelor de reticulină este întreruptă, ceea ce duce la apariția unor rupturi în peretele intestinal și/sau arterele mari, unde fibrele de reticulină formează o teacă în jurul SMC (sindromul). Ehlers–Danlos tipul IV).
membrana bazala
Membrana bazală este un tip special de matrice extracelulară, care arată ca o foaie sau o placă cu o grosime de 20-200 nm (vezi Fig. 16-2 ) și constă din proteine speciale care servesc la conectarea celulelor de diferite tipuri
(epitelial, muscular, Schwannian) cu țesutul conjunctiv înconjurător. Membrana bazală este formată din două straturi - lamina lucida(strat ușor) 10–50 nm grosime, adiacent plasmalemei și lamina densa(strat dens de electroni care variază în grosime). Urzeală lamina lucidaŞi lamina densa - o retea tridimensionala formata din fibre de colagen de tip IV, la care sunt asociate laminina, entactina si sulfatul de heparan.
Fibre de ancorare
Majoritatea membranelor bazale sunt ferm atașate de țesutul conjunctiv lax. Membranele bazale ale epiteliului stratificat al pielii, esofagului, corneei și amnionului sunt conectate prin fibre scurte de ancorare la plăci de ancorare în țesutul conjunctiv subiacent. Fibrele de ancorare constau în principal din colagen de tip VII, plăcile de ancorare conțin colagen de tip IV. Fibrele de ancorare conectează, de asemenea, plăcile de ancorare în stromă. Rezultatul este o rețea densă de fibre întrețesute care stabilizează membrana bazală în țesuturile supuse unor forțe de forfecare semnificative.
Membrana bazală unește celulele, facilitând organizarea lor într-un strat; mentine fenotipul celular si influenteaza diferite tipuri activitate celulară: aderență, diferențiere, polarizare, migrare. Membrana bazală împiedică celulele epiteliale să invadeze țesutul conjunctiv subiacent. În neoplasmele maligne, întreruperea acestei funcții duce la germinarea celulelor tumorale și formarea de metastaze. Membrana bazală filtrează macromoleculele (de exemplu, în rinichi lamina densa captează molecule cu M r mai mare de 50 kDa).
Celulele țesutului conjunctiv sunt diverse. Acestea sunt fibroblaste, fibrocite, condroblaste, condrocite, osteoblaste, osteocite, macrofage, mastocite, leucocite, plasmocite, pericite, adipocite. Celulele țesutului conjunctiv sunt împărțite în două grupuri - rezidenți și imigranți. Rezidenți – fibroblaste și fibrocite (țesut conjunctiv fibros), condroblaste și condrocite (țesut cartilaginos), osteoblaste și osteocite (țesut osos), mastocite, adipocite, macrofage. Imigranții sunt leucocite (neutrofile, eozinofile, bazofile, monocite, limfocite). Când apar focare de inflamație, aceste celule părăsesc fluxul sanguin în țesutul conjunctiv. Limfocitele, care efectuează supraveghere imunologică, circulă constant între sânge, țesuturi conjunctive și limfă.
Grupuri funcționale.În funcție de funcția pe care o îndeplinesc, celulele țesutului conjunctiv pot fi împărțite în trei grupuri principale. Celulele responsabile de sinteza moleculelor de substanță extracelulară și de menținerea integrității structurale a țesutului sunt fibroblastele și fibrocitele, condroblastele și condrocitele, osteoblastele și osteocitele, odontoblastele, celulele reticulare.
Celulele responsabile de acumularea și metabolismul grăsimilor sunt adipocitele; aceste celule formează țesut adipos. Celulele cu funcții de protecție(inclusiv imunologic) - grăsime, macrofage și toate tipurile de leucocite.
Fibroblaste și fibrocite
Fibroblast
Fibroblast (Fig. 6-22 , vezi si fig. 3 inset) – cel mai comun tip de celule de țesut conjunctiv; secretă componente ale matricei extracelulare, participă la vindecarea rănilor și este capabil de proliferare și migrare. Fibroblastul este o celulă aplatizată, în formă de stelat, care formează procese largi în formă de pană; conţine un nucleu oval mare cu mai mulţi nucleoli. Dimensiunea celulei este variabilă. Fibroblastul sintetizează intens proteine, astfel încât citoplasma acestuia conține cantități mari de cisterne ale reticulului endoplasmatic granular, un complex bine definit. Golgi, multe mitocondrii. Există lizozomi și granule secretoare, glicogen, numeroase microfilamente și microtubuli.
Funcții. Fibroblastele sintetizează colagenul, elastina, fibronectina, glicozaminoglicanii, proteoglicanii și alte componente ale matricei extracelulare. Fibroblastele produc diverse citokine - factor de stimulare a coloniilor de granulocite-macrofage (GM-CSF), factor de stimulare a coloniilor de granulocite (G-CSF) și factor de stimulare a coloniilor de macrofage (M-CSF). Fibroblastele măduvei osoase secretă ILZ și IL7. În timpul inflamației și vindecării rănilor, fibroblastele sunt activate de factorii de creștere
Orez. 6-22. Fibroblast ( O) și fibrocite ( B). Fibroblastul (forma activă a celulei) conține organite bine definite: reticul endoplasmatic granular, complex Golgi, mitocondriile. Fibroblastul formează procese mari alungite. Într-un fibrocit, există semnificativ mai puține organele, celula este lipsită de procese și are o formă în formă de fus.
bFGF și PDGF proliferează și migrează la locul leziunii. Fibroblastele conțin diverse enzime, inclusiv colagenaze, care descompun colagenul. Prin distrugerea deteriorate și sintetizând noi macromolecule ale matricei extracelulare, fibroblastul promovează restructurarea acestuia și formarea de cicatrici la locul leziunii (inflamație).
fibrocit
Fibrocite (fig. 6-22 ) este o formă matură de fibroblast prezentă în țesutul conjunctiv dens, format. Fibrocitul are o formă de fus. Nucleul compactat este alungit și situat de-a lungul celulei. Există cisterne împrăștiate ale reticulului endoplasmatic granular și un număr mic de mitocondrii. Complex Golgi slab dezvoltat. Celula conține relativ puține granule secretoare. Funcția fibrocitelor este de a menține structura țesutului prin reînnoirea continuă (deși lentă) a componentelor matricei extracelulare. În timpul vindecării rănilor, fibrocitele pot fi stimulate la activitate sintetică. Un fibrocit activat capătă caracteristicile unui fibroblast: nucleul este rotunjit, numărul de cisterne reticulului endoplasmatic și mitocondriile crește; complex Golgi devine mai pronunțată.
Fibroblaste pericriptale
Mucoasa intestinală conține fibroblaste pericriptale, celule stromale care prezintă caracteristicile morfologice ale SMC. Se crede că ele reglează creșterea și diferențierea celulelor epiteliale. Aceste fibroblaste exprimă actina musculară netedă.
Miofibroblaste
Miofibroblastele sunt celule contractile care au caracteristici comune cu MMC (vezi capitolul 7 ).
Macrofage
Macrofagul este o formă diferențiată de monocite. Macrofagele sunt fagocite profesionale; se găsesc în toate țesuturile și organele. Aceasta este o populație foarte mobilă de celule care se poate mișca rapid. Speranța de viață este de luni. Macrofagele tisulare păstrează o anumită capacitate de a se diviza (de exemplu, macrofagele alveolare în procesele inflamatorii cronice). În focarul inflamației, ca urmare a fuziunii mai multor macrofage, se formează celule gigantice multinucleate de corpuri străine. Macrofagele sunt împărțite în rezidente și mobile. Macrofagele rezidente sunt prezente în țesuturi în mod normal, în absența inflamației. Printre acestea, există macrofage libere, rotunde și fixe - celule în formă de stea, atașate cu procesele lor de matricea extracelulară sau de alte celule. Macrofagele mobile sunt o populație de macrofage migratoare (numite).
Sistemul fagocitar mononuclear. Macrofagele țesutului conjunctiv fac parte din sistemul fagocitar mononuclear. Celulele sistemului fagocitar mononuclear diferă de alte celule fagocitare în funcție de trei criterii:
| Orez. 6-23. Macrofag. Miezul este de formă neregulată, cu crestături. Citoplasma conține ribozomi, mitocondrii, corpuri multiveziculoase Complex bine dezvoltat Golgiși reticulul endoplasmatic granular. Există numeroși lizozomi. Prezența fagozomilor, fagolizozomilor și a corpurilor reziduale este caracteristică. Celula formează procese citoplasmatice implicate în migrație și fagocitoză. | 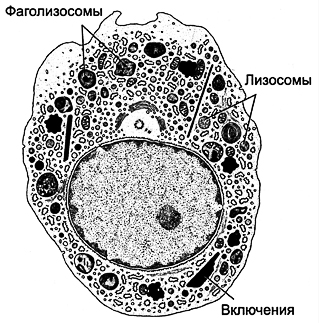 |
au morfologia macrofagelor, provin din măduva osoasă, activitatea fagocitară este modulată de componente Ig și complement. Sistemul fagocitar mononuclear include histiocite (macrofage tisulare), macrofage alveolare, osteoclaste, celule von Kupffer, celule Langerhans, celule Hofbauer, celule gigantice cu corpuri străine și probabil celule microgliale ale sistemului nervos central.
Morfologie
Structura (Fig. 6-23 ) a unui macrofag depinde de activitatea și localizarea acestuia. Diametrul celulei este de aproximativ 20 de microni. Miezul are formă neregulată, cu crestaturi. Citoplasma conține mitocondrii, ribozomi liberi și un complex bine definit Golgi, corpi multiveziculare, reticul endoplasmatic granular, lizozomi, fagolizozomi și corpuri reziduale, al căror material poate fi eliberat din macrofag prin exocitoză. Lizozomii conțin agenți bacterici: mieloperoxidază, lizozim, proteinaze, hidrolaze acide, proteine cationice, lactoferină, superoxid dismutază - enzimă care favorizează formarea H 2 O 2, OH -, O - 2. Sub plasmalemă, microfilamentele de actină, microtubulii și filamentele intermediare necesare migrării și fagocitozei sunt prezente în cantități mari. Macrofagele migrează de-a lungul unui gradient de concentrație al multor substanțe care provin din diverse surse. Macrofagele activate formează pseudopode citoplasmatice de formă neregulată care participă la mișcarea ameboidului și la fagocitoză.
Macrofagele sunt fagocite profesionale. Ele captează din sânge proteine denaturate și celule roșii îmbătrânite (macrofage fixe ale ficatului, splinei, măduvei osoase). Macrofagele fagocitară resturile celulare și matricea tisulară. Fagocitoza nespecifică este caracteristică macrofagelor alveolare, captând particule de praf de diferite naturi, funingine etc. Fagocitoza specifică apare atunci când un macrofag interacționează cu o bacterie opsonizată. Un macrofag activat secretă mai mult de 60 de factori. Macrofagele prezintă efecte antibacteriene
activitate, eliberând lizozimă, hidrolaze acide, proteine cationice, lactoferină, H 2 O 2, OH -, O 2 -. Activitatea antitumorală constă în efectul citotoxic direct al H 2 O 2, arginazei, proteinazei citolitice, factorului de necroză tumorală (TNF) al macrofagului. Un macrofag este o celulă prezentatoare de antigen: prelucrează Ag și îl prezintă limfocitelor, ceea ce duce la stimularea limfocitelor și declanșarea reacțiilor imune. IL1 din macrofage activează limfocitele T și, într-o măsură mai mică, limfocitele B. Macrofagul produce mediatori lipidici - prostaglandina PGE 2 și leucotriene, factorul de activare a trombocitelor (PAF). Celula secretă, de asemenea, α-IFN, care blochează replicarea virală. Un macrofag activat secretă enzime care distrug matricea extracelulară (elastaza, hialuronidază, colagenază). Pe de altă parte, factorii de creștere sintetizați de macrofage stimulează eficient proliferarea celulelor epiteliale (factorul de creștere transformator TGFα, bFGF), proliferarea și activarea fibroblastelor (factorul de creștere derivat din trombocite PDGF), sinteza colagenului de către fibroblasti (factorul de creștere transformator TGFβ) și formarea de noi vase de sânge (factorul de creștere a fibroblastelor bFGF). Astfel, principalele procese care stau la baza vindecării rănilor (reepitelializarea, formarea matricei extracelulare, repararea vaselor deteriorate) sunt mediate de factorii de creștere produși de macrofage. Prin producerea unui număr de factori de stimulare a coloniilor (macrofage - M-CSF, granulocite - G-CSF), macrofagele influențează diferențierea celulelor sanguine.
Mastocit
Mastocitele sunt similare din punct de vedere morfologic și funcțional cu bazofilele din sânge, dar sunt tipuri diferite de celule. Mastocitul, ca și bazofilul, provine dintr-un precursor din măduva osoasă, dar diferențierea finală are loc în țesutul conjunctiv. Mastocitele sunt celule rezidente ale țesutului conjunctiv. Există mai ales multe în piele, în membrana mucoasă a sistemelor respirator și digestiv și în jurul vaselor de sânge. Mastocit (Fig. 6-24 ) conține numeroase granule metacromatice mari (lizozomi modificați). Diferiți receptori sunt încorporați în membrana celulară, inclusiv receptori pentru fragmentul Fc al IgE.
Mastocitele sintetizează și acumulează în granule o varietate de biologic substanțe active, mediatori și enzime: heparină (sulfat de heparină), histamină, triptază, chimază, elastază, dipeptidază, activator de plasminogen, hidrolaze acide, factor de chemotaxie a eozinofilelor (ECF), factor de chemotaxie a neutrofilelor (NCF). Componenta principală a granulelor de mastocite este heparina glicozaminoglican sulfatată încărcată negativ, sintetizată și stocată exclusiv de mastocite. Heparina secretată de celulă leagă antitrombina III care circulă în sânge, sporind brusc activitatea sa anticoagulantă. Histamina determină contracția SMC, hipersecreția de mucus și creșterea permeabilității vasculare odată cu dezvoltarea edemului. Triptaza favorizează descompunerea fibrinogenului,
conversia C3 în anafilatoxină C3a, activarea colagenazei, degradarea fibronectinei. Triptaza, chimaza, carboxipeptidaza B, alte proteaze și hidrolaze acide, eliberate din celula degranulată, provoacă distrugerea matricei tisulare. Când mastocitele sunt activate (împreună cu secreția de conținut de granule), se formează metaboliți ai acidului arahidonic - prostaglandine, tromboxan TXA 2 și leucotriene. Acești mediatori au proprietăți vaso- și bronhoactive. Factorul de activare a trombocitelor (PAF), care este unul dintre cei mai puternici spasmogeni, este, de asemenea, format din fosfolipidele membranare.
Mastocitele sunt implicate în reacții inflamatorii și alergice. Activarea și degranularea mastocitelor, precum bazofilele, este mediată de IgE (vezi Fig. 11-4 ). Mastocitele au receptori de suprafață de mare afinitate pentru fragmentele Fc ale IgE. Legarea Ag (alergen) de molecula IgE de pe suprafața mastocitelor este însoțită de exocitoza conținutului de granule și formarea metaboliților acidului arahidonic.
Celula plasmatică
Celulele plasmatice (Fig. 6-25 ) se diferențiază de limfocitele B activate imunologic, sintetizează și secretă Ig (AT). Nucleul mare, rotund sau oval, este situat excentric și conține cromatina dispersată. În citoplasmă există un complex bine dezvoltat Golgiși o masă de cisterne mari de reticul endoplasmatic granular, situate concentric în jurul nucleului.
Pericitele
Pericitele sunt celule de proces adiacente la exterior arteriolelor, venulelor și capilarelor. Cele mai numeroase în venule postcapilare.
Morfologie
Pericitele au un nucleu în formă de disc cu depresiuni mici, conțin setul obișnuit de organite, corpuri multiveziculoase, microtubuli și glicogen. Zona care se confruntă cu peretele vasului conține bule. Proteinele contractile sunt prezente în apropierea nucleului și în procese, inclusiv. actina si miozina. Pericitele sunt acoperite de o membrană bazală, dar sunt strâns asociate cu
celula endotelială, deoarece membrana bazală dintre ele poate fi absentă. S-au identificat goluri și contacte adezive în aceste locuri.
Pericitele sintetizează componente ale membranei bazale capilare; controlează proliferarea celulelor endoteliale atât în timpul creșterii vasculare normale, cât și în timpul regenerării lor; modulează funcția celulelor endoteliale prin reglarea transportului macromoleculelor de la capilare la țesuturi. În timpul vindecării rănilor și al restaurării vasculare, pericitele se diferențiază în SMC.
Adipocite
Dintre adipocite se disting celulele adipoase albe și maro.
Celulă de grăsime albă
În timpul diferențierii, picăturile de grăsime apar în citoplasma celulei mezenchimale, contopindu-se pe măsură ce numărul lor crește. Un adipocit diferențiat este o celulă rotundă mare, cu un diametru de 120 de microni, care conține o picătură mare de grăsime, împingând citoplasma și toate organitele spre periferie. În citoplasmă, o margine îngustă care înconjoară o picătură de grăsime, există un nucleu turtit, ribozomi liberi, reticul endoplasmatic neted și granular, un complex Golgiși mitocondriile. Adipocitele secretă hormonul leptina, care interacționează cu receptorii celulele nervoase nucleii ventromediali și laterali ai hipotalamusului duce la activarea mecanismelor centrale ale comportamentului alimentar; leptina contracarează acumularea de țesut adipos.
Sinteza și depozitarea grăsimilor. Adipocitele sintetizează lipoprotein lipaza, care difuzează pe suprafața celulelor endoteliale (în principal în capilarele țesutului adipos). Lipidele alimentare sunt transportate în țesutul adipos sub formă de VLDL și sub formă de chilomicroni. Lipoprotein lipaza hidrolizează aceste lipide în acizi grași. Acizii grași liberi pătrund în adipocite și sunt stocați în picături de grăsime sub formă de trigliceride. Celulele grase sunt capabile să sintetizeze acizi grași
tot din glucoză şi aminoacizi. Insulina stimulează sinteza lipoprotein lipazei, depunerea de grăsime și absorbția de glucoză de către adipocite. În același timp, insulina încetinește mobilizarea grăsimilor din adipocite, suprimând activitatea enzimelor care descompun grăsimea. Trigliceridele stocate sunt hidrolizate de lipaza sensibilă la hormoni activată de cAMP. Acizii grași liberi rezultați intră în lumenul capilarelor, unde se leagă necovalent de albumină și sunt transportați la ficat.
Celulă de grăsime brună
Celula de grăsime brună conține multe picături mici de grăsime și mitocondrii mari. În adipocitele brune, un mecanism natural pentru decuplarea funcțiilor de fosforilare oxidativă, care este util din punct de vedere biologic ca modalitate de a genera căldură. Culoarea maro a celulei și a țesutului în ansamblu se datorează prezenței pigmenților care conțin fier în mitocondrii. Lipaza activată sensibilă la hormoni hidrolizează trigliceridele în acizi grași și glicerol. Acizii grași eliberați sunt metabolizați pentru a produce căldură. Creșterea generării de căldură se explică prin prezența proteinei transmembranare termogenină în membrana interioară a mitocondriilor. Termogenina permite fluxul invers al protonilor transportați anterior în spațiul intermembranar fără a trece prin sistemul ATP sintetazei. Astfel, energia generată de curentul de protoni nu este folosită pentru sinteza ATP, ci este disipată sub formă de căldură.
TIPURI DE ȚESUT CONECTIV
Există țesuturi conjunctive fibroase și țesuturi conjunctive cu proprietăți speciale. Țesuturile conjunctive fibroase (în funcție de numărul de fibre și de substanța principală din matricea extracelulară) sunt împărțite în laxe și dense. La rândul său, țesutul conjunctiv dens este împărțit în format și neformat, care este determinat de organizarea fibrelor în matricea extracelulară. Țesuturile conjunctive cu proprietăți speciale includ embrionare (mezenchim), adipos și reticular. Aceste țesuturi se caracterizează printr-o predominanță a celulelor de același tip.
Țesut conjunctiv fibros
Țesut conjunctiv lax
Țesut conjunctiv lax (neformat) (Fig. 6-26 ) se găsește în toate organele, le formează stroma și însoțește vasele. Acest țesut conține relativ puține fibre de colagen și elastice distribuite aleator. Între fibre există o cantitate mare de substanță fundamentală cu diferite celule scufundate în ea: fibroblaste, macrofage migratoare și rezidente, mastocite, pericite, adipocite, plasmocite, leucocite. Moleculele de glicozaminoglican, împletindu-se, formează o rețea, în celulele și canalele căreia se reține o cantitate mare de lichid tisular cu substanțe dizolvate.

Figura 6-26Țesut conjunctiv lax Substanța intercelulară constă de cele mai multe ori din substanța principală cu fibre distribuite aleator, colagen și elastic. Diversitatea celulară caracteristică (fibroblaste, mastocite, diverse leucocite, adipocite, macrofage, plasmocite)
substante. Leucocitele, macrofagele, mastocitele și celulele plasmatice au un rol activ în reacțiile de protecție.
Țesuturi conjunctive dense
Țesutul conjunctiv dens conține un număr mare de fibre dens aranjate. Cantitatea de substanță principală este relativ mică.
Țesut conjunctiv dens, neformat consta din cantitate mare fibre dens dar dispuse aleatoriu. Între fibre sunt prezente fibroblaste (fibrocite), macrofage și mastocite. Caracteristic pielii în sine, periostul.
Țesut conjunctiv format dens. Fibrele sunt aranjate strâns, formând mănunchiuri paralele. În spațiile înguste dintre fibre, fibrocitele se aliniază în lanțuri. Din acest țesut se formează ligamente, tendoane și membrane fibroase.
- Elastic buchet(De exemplu, ligamenta flavaŞi ligamentum nuchae) este format din elastina, care formeaza fibre groase. Între ele se află fibre subțiri de colagen și fibrocite.
- Tendon(orez. 6-27 ) constă din fibre de colagen care formează fascicule tendinoase de ordine I, II și III. Între fasciculele de ordinul întâi se află șiruri de fibrocite (celule tendinoase) cu procese lamelare. Grupuri de fascicule de ordinul întâi, înconjurate de țesut conjunctiv lax cu vase și nervi, formează mănunchiuri de ordinul doi. Mai multe fascicule de ordinul doi sunt unite prin țesut conjunctiv lax în fascicule ordinul III. Când un tendon este deteriorat, fibrocitele și fibroblastele activate sintetizează colagen pentru noi fibre.
- Membrane fibroase- un tip de țesut conjunctiv dens, de formă. Mănunchiuri de fibre de colagen și fibrocite aflate între ele sunt aranjate în straturi. În fiecare strat, mănunchiuri ondulate de fibre de colagen sunt paralele într-o direcție, diferită de direcțiile din straturile adiacente. Mănunchiuri individuale de fibre trec de la un strat la altul, conectându-le între ele. Membranele fibroase includ fascia, aponevroze, centrul tendonului diafragmei, capsule organele interne, dura mater, sclera.
Țesuturi conjunctive cu proprietăți speciale
Țesuturile conjunctive cu proprietăți speciale includ mezenchimul, țesutul reticular și adipos.
Mezenchim
Țesut conjunctiv embrionar - mezenchim (Fig. 6-28 ) este sursa de origine a celulelor tuturor țesuturilor conjunctive. Celulele mezenchimale sunt stelate sau în formă de fus, cu procese delicate de ramificare care formează o rețea. În miezul oval – dispersat

Figura 6-27 Tendon O– secțiune transversală B– secţiune longitudinală ÎNŞi G– schema structurii tendonului în secțiuni transversale și longitudinale
cromatină și nucleoli clar vizibili. Materialul extracelular asemănător gelului constă aproape exclusiv din substanță fundamentală și o cantitate minimă de fibre împrăștiate (în principal reticulină).
Țesut reticular
Țesutul reticular (Fig. 6-29 ) are o structură asemănătoare rețelei și constă din fibre de reticulină și celule reticulare cu procese lungi. Celulele reticulare împreună cu fibrele de reticulină formează o rețea liberă. Țesutul reticular creează un micromediu specific pentru dezvoltarea celulelor sanguine în plus, celulele reticulare sintetizează diverși factori de creștere hematopoietică (ILZ, IL7, GM-CSF, G-CSF, M-CSF).
Țesut adipos
Țesutul adipos se găsește în multe organe. Există țesut adipos alb și maro.
Grăsime albă alcătuiește aproape tot țesutul adipos al corpului. Participă la absorbția din sânge, sinteza, depozitarea și mobilizarea lipidelor neutre (trigliceride). Distribuția țesutului adipos în organism este influențată de hormonii sexuali și hormonii cortexului suprarenal. Celulele adipoase (adipocitele) formează grupuri (lobuli), separate prin partiții de țesut conjunctiv lax. În acesta din urmă, vasele de sânge și nervii trec în țesutul adipos. Celulele adipoase individuale sunt înconjurate de o rețea de reticulină și fibre de colagen. Fibroblastele și mastocitele sunt prezente în septurile țesutului conjunctiv.
Grăsime brună la un nou-născut este implicat în termoreglare. La adulți, grăsimea brună se găsește în cantități mici în mediastin, de-a lungul aortei și sub piele între omoplați. Țesutul adipos brun este alimentat din abundență cu capilare sanguine, formând o rețea în jurul fiecărui adipocit și are o inervație simpatică pronunțată.