structura rădăcină primară a unei secțiuni transversale a unei rădăcini de iris. Într-o secțiune, chiar și la o mărire mică, se distinge clar o mică parte internă - cilindrul central și cortexul primar exterior, acoperit cu un singur strat de celule cu fire de păr de rădăcină - rizoderm (epiblem). Pe măsură ce firele de păr mor, pereții celulelor exterioare ale cortexului primar sunt acoperiți în interior cu un strat subțire de suberin. Suberinizarea face celulele impermeabile fie la deșeuri, fie la gaze. Aceste mai multe straturi de celule corticale sunt numite exodermă, efectuează f-yu tegumentarțesături. (Stratul exterior al cortexului primar, exodermul, este format din celule poligonale bine închise, ai căror pereți sunt ulterior suberizați și îndeplinesc o funcție de protecție). Sub exoderm există celule vii de parenchim (parenchim) ale cortexului primar, situate lax și formând spații intercelulare.
Stratul interior al cortexului primar, endodermul, este format dintr-un singur rând de celule, cu pereții radiali și interni îngroșați. Pereții celulelor endodermice devin mai mult sau mai puțin suberizați, iar la unele plante (iris) se lignifică. Dar nu toate celulele endodermice au un perete impenetrabil printre celulele cu pereți groși, în unele locuri există celule vii cu pereți subțiri, numite celule de trecere; Prin ele intră apa în cilindrul central. Stratul exterior cilindru central- periciclu - este format dintr-un rând de celule vii de parenchim. Partea centrală a cilindrului central este ocupată de un fascicul radial conducător. Xilemul este situat în centru și formează o serie de proiecții ascuțite sub formă de raze. Între proiecțiile xilemului există zone de floem. 72. Secțiune transversală a rădăcinii irisului (Iris germanica), 1 – rizoderm (epiblem), 2 – exoderm, 3 – parenchim principal (mezoderm), 4 – endoderm, 5 – celula de trecere a endodermului, 6 – pericic, 7 – raza xilemului primar, 8 - zona floemului primar (2-5 - cortexul primar, 6-8 - cilindrul central). .
Secundar structura anatomică rădăcină Rădăcini. 1. Folosind o secțiune transversală pregătită a unei rădăcini de dovleac, se studiază structura anatomică secundară a rădăcinii. La o mărire mică, în centrul cilindrului se găsește un xilem cu patru raze. Din razele xilemului primar încep razele radiale (nucleu) - secțiuni de parenchim viu cu pereți subțiri. Alternând cu razele radiale sunt zone largi de xilem secundar cu vase mari și parenchim lemnos cu celule mici. La marginea xilemului secundar, cambiul este clar vizibil. În afara cambiului, vizavi de fiecare secțiune a xilemului secundar, se află floemul secundar. Exteriorul rădăcinii este acoperit cu un strat relativ subțire de plută. Țesuturile situate în exteriorul cambiului (floem, parenchim de bază, feloderm și cambium plută) se numesc cortex secundar.
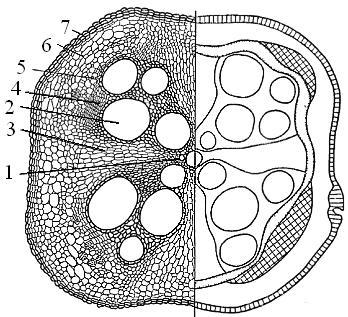 Orez. 73. Structura secundară a rădăcinii de dovleac (Cucurbita pepo):
Orez. 73. Structura secundară a rădăcinii de dovleac (Cucurbita pepo):
A - diagrama unei secțiuni transversale (în stânga - un desen detaliat, în dreapta - o schemă); B - fragment din desen.
1 - xilem primar, 2 - xilem secundar, 3 - rază radială, 4 - cambium, 5 - floem primar și secundar, 6 - parenchim principal al cortexului secundar, 7 - periderm (1-3 - xilem, 5-7 - secundar cortex) .
2. folosind micropreparate și culturi de rădăcină vii de morcov, ridichi, sfeclă, studiază natura structurii anatomice și stabilesc tipul de rădăcină. Pe o secțiune transversală a unei rădăcini de morcov, puteți discerne un inel ușor care separă partea exterioară mai largă de cea interioară mai mică. Acesta este cambium. În afara ei este scoarța secundară, iar în interior este xilema. Două raze radiale ale parenchimului se extind din razele xilemului primar (fascicul diarh), iar între ele sunt situate două secțiuni în formă de evantai ale xilemului secundar. Cortexul secundar este format din parenchim și un număr mic de tuburi sită cu celule însoțitoare. În consecință, produsele de rezervă din rădăcină de morcov se depun în scoarța secundară.
Când se examinează o secțiune transversală a unei culturi de rădăcină de ridiche, în centru se găsește un xilem primar biradial. De la capete se extind două raze radiale de parenchim, apoi două secțiuni de xilem secundar, un strat de cambium și, în final, o scoarță secundară. Se observă că cea mai mare zonă a secțiunii transversale este ocupată de parenchim nelignificat, situat în interiorul inelului cambial, adică parenchimul xilemului și razelor radiale. Acest parenchim conține produse de rezervă. Vasele, înconjurate de un inel de parenchim lignificat, sunt situate în grupuri împrăștiate de-a lungul razelor printre țesutul de depozitare.
Când se examinează o secțiune transversală a unei rădăcini de sfeclă, în centrul este vizibil un xilem primar cu două raze, la care sunt adiacente două secțiuni de xilem secundar, separate de raze radiale ale parenchimului. Urmează cambiul cu zone adiacente de floem secundar. Apoi vine un strat de inele cambiale accesorii. se formează datorită diviziunii celulelor periciclului, în timpul căreia se formează un strat de celule parenchimoase. În acest strat, un rând de celule începe să se dividă prin septuri tangențiale și devine un nou strat de cambium. Stratul cambial depune xilemul spre interior și floemul spre exterior, care formează mici mănunchiuri vasculare colaterale separate unul de celălalt printr-un strat de parenchim cu pereți subțiri. În același timp, în stratul periferic al parenchimului se formează un nou inel de celule cambiale etc.
Că. straturile dure de pe secțiunea transversală a culturii de rădăcină sunt xilemul rândurilor concentrice de mănunchiuri vasculare, iar straturile largi suculente constau din cambium, floem și parenchim de depozitare. Cultiva de rădăcină de sfeclă este policambială.

Orez. 16. Structura secundară a rădăcinii morcovului (Daucus carota): 1 - periderm; 2 - parenchimul cortexului secundar; 3 - cambium; 4 - xilem secundar; 5 - xilem primar; 6 - raza radială 7 - floem primar și secundar.

Orez. 17. Structura secundară a rădăcinii de ridiche (Raphanus sativus): 1 - periderm; 2 - parenchimul cortexului secundar; 3 - xilem secundar 4 - floem primar și secundar; 5 - cambium; 6 - rază radială 7 - xilem primar

Orez. 18. Structura rădăcinii policambiale a sfeclei (Beta vulgaris): 1-periderm; 2 - straturi suplimentare de cambium; 3 - xilem secundar 4 - rază radială; 5 - xilem primar; 6 – cambium 7 – floem primar și secundar; 8 - fascicule vasculare colaterale
Stratul exterior al cortexului primar, care mărginește epiblema, numit exodermă, este format din celule strâns închise. Perețiile radiale longitudinale dintre ele în cea mai mare parte nu constituie o continuare a pereților radiali ale epiblemei, ci alternează cu acestea (Fig. 161, 2 , 162).
La multe plante, pereții celulari ai exodermului sunt de obicei supuși suberizării până la moartea epiblemei, iar apoi exodermul devine un strat tegumentar, protector al rădăcinii, asemănător într-o măsură mai mare sau mai mică cu pielea tulpinilor și frunze.
eu- strat purtător de păr (epiblema) cu fire de păr din rădăcină; II- cortexul primar cu exoderm ( 2 ) și endodermul ( 3 ), 4 - centura Casparian; III- cilindru axial, în el: 5 - periciclu, 6 - xilem primar, 7 - miez, 8 - cambium, 9 - floem primar (după Muromtsev).
În funcție de mediul în care se află rădăcina și de durata de funcționare a exodermei, aceasta atinge diferite grade de specializare.
Deci, la rădăcina unor acvatice monocotiledone(de exemplu, acuarelă) exodermul nu se diferențiază, deoarece peridermul se formează foarte devreme.
La unele monocotiledone cu rădăcini puternice (palmier de curmal, cana), dimpotrivă, celulele exodermice într-un stadiu incipient se divid tangențial
septuri: se formează exodermul, format din 2-20 straturi de celule. Exodermul multistrat este similar cu țesutul de plută, dar diferă de acesta nu numai prin metoda de formare, ci și prin aranjarea celulelor: în secțiunile radiale longitudinale ale rădăcinii, acestea nu formează rânduri regulate.
Exodermul este caracteristic mai ales plantelor monocotiledonate ale căror rădăcini nu au îngroșare secundară; cortexul primar persistă foarte mult timp și nu se formează periderm.
Sub exoderm se află restul cortexului primar; stratul cel mai interior, care mărginește cilindrul axial, se numește endoderm.
De obicei, cortexul primar este format din celule vii de parenchim cu pereți subțiri de celuloză. În regiunea exterioară, celulele sale sunt cu mai multe fațete, strâns închise, dar în cortexul interior, celulele sunt rotunjite într-un grad sau altul, iar între ele se formează spații intercelulare purtătoare de aer - foarte mici (în rădăcinile multor irisi) sau mai mare. Dimensiunea celulelor crește, de obicei, în direcția de la exoderm la partea mijlocie a cortexului primar, apoi scade spre endoderm (Fig. 162). La unele plante (în iris), pe lângă exoderm, alte 2-3 straturi exterioare ale cortexului primar sunt supuse suberizării. La multe cereale mari, pereții celulari ai stratului adiacent exodermului, în special pereții tangențiali exteriori, devin foarte groși, dopați și lemnos.
În rădăcinile care se dezvoltă în aer sau apă, celulele cortexului primar conțin plastide cu clorofilă.
Rezerve mari se acumulează în celulele parenchimatoase ale cortexului primar nutrienti nu numai în monocot, ci și în unele

Orez. 162. O parte a secțiunii transversale a rădăcinii irisului ( Iris germanica):
epb- epibleme; copie- exoderm cu trei straturi; prh- parenchimul de stocare al cortexului primar; Sfârşit- endoderm; p.kl. - cușcă de acces; pc- periciclu; p.k.s.. - xilem primar; p. fl. - floem primar; m. tk. - tesatura mecanica.
dicotiledonate în care scoarța primară este reținută pentru o lungă perioadă de timp, de exemplu, în ranuncul ( Ficaria ranunculoides), de la luptător ( Aconitum napellus). În aceste cazuri, cortexul primar este foarte puternic.
În zona cortexului primar, printre parenchim, se formează lacticifere - articulate (în flori de clopoțel) sau nesegmentate (în clopoțel grecesc). Periploca graeca), grupuri de celule pietroase (la anghinare), cordoane de sclerenchim (la curmal). La multe ierburi (inclusiv iarba Sudanului), două sau mai multe straturi ale cortexului primar exterior se diferențiază în sclerenchim, uneori cu pereți foarte groși.
La multe plante, în special cele acvatice (calamus) și cele care trăiesc pe soluri mlăștinoase (unele soiuri de orez cultivate în zone inundate cu apă), în scoarța primară se formează canale mari purtătoare de aer. Experimentele au arătat că puterea sistemului pneumatic este înăuntru relație inversă din aerarea substratului. Astfel, în experimentele cu porumb White Tooth și grâu Marquis în culturi acvatice, ambele plante au avut pasaje de aer puternice. La culturile cultivate pe nisip și sol, pasajele de aer nu s-au format deloc la grâu, în timp ce la porumb s-au dezvoltat, dar slab. La cultivarea porumbului în culturile acvatice, s-a dovedit că aerarea slăbește foarte mult procesul de formare a căilor de aer.
Stratul interior al cortexului primar, numit endoderm, este format din celule de parenchimat strâns, cu o structură caracteristică. În stare tânără, endodermul apare în secțiune transversală ca un inel de celule dreptunghiulare cu pereți subțiri de celuloză, cu cavități umplute cu conținut viu; Secțiunile longitudinale arată că endodermul este format din celule lungi și scurte; scurte sau împrăștiate printre lungi, sau dispuse în rânduri longitudinale. Mai târziu, endodermul trece printr-una, două sau trei etape de diferențiere.
În prima etapă, în pereții radiali transversali și longitudinali ai celulelor endodermice, modificări biochimice, acoperind banda din mijloc a peretelui; în această zonă, peretele, în curs de suberizare și lignificare, se diferențiază într-un cadru sau brâu (beaua caspariană), parcurgând continuu pe cele patru laturi ale celulei. Centurile caspariene ale celulelor endodermice vecine sunt strâns adiacente una cu cealaltă (Fig. 163).
În endoderm, intrând în a doua fază de diferențiere, pe coajă se depune din interior un strat continuu de îngroșare, format din celuloză și suberină. Totuşi, unele celule situate vizavi de grupele de xilem ale cilindrului axial (celule de trecere) rămân în prima etapă (Fig. 162).
Nu toate plantele intră în a doua etapă: în coada-calului și

Orez. 163. Parte dintr-o secțiune transversală a rădăcinii ranuncul caustic ( Ranunculus acer):
şi- endoderm cu centuri caspariene ( PC); pc- periciclu; f- floem; ks- xilem.
La unele ferigi, dezvoltarea sa se limitează la prima etapă. La gimnosperme și dicotiledonate, endodermul trece de obicei prin a doua etapă. In unele dicotiledonate si in majoritatea monocotiledonatelor intra in stadiul al treilea. În această etapă, un strat de celuloză lignificată este depus în celulele endodermice în interiorul plăcii de suberină. Depunerea este de obicei neuniformă: este slabă pe peretele tangențial exterior, intensificându-se pe pereții radiali și transversali spre centrul rădăcinii; peretele tangenţial interior se îngroaşă în cea mai mare măsură. Îngroșarea descrisă este de obicei clar stratificată (Fig. 162).
În endodermul primei etape, celulele de trecere ale unor plante rămân de obicei cu pereți subțiri și vii pentru o perioadă de timp și trec prin ele însele în direcția razei rădăcinii apa cu substanțe dizolvate în ea (Fig. 162), dar apoi trec în a doua și a treia etapă. La unele cereale, celulele unuia sau două straturi ale cortexului primar adiacente endodermului devin foarte asemănătoare cu celulele endodermului din stadiul al treilea (Fig. 164). La unele plante (de exemplu, cereale), întreg cortexul primar al unor rădăcini suferă sclerificare în timp - îngroșare și lignificare pereții celulari. După ce endodermul intră în a treia etapă, cortexul primar moare uneori și se dezlipește. La dicotiledonate, moartea și descuamarea cortexului se produce ca urmare a unei îngroșări secundare puternice și a formării peridermului în periciclu (vezi mai jos) și cu endoderm în a doua sau chiar prima etapă.
Primarstructura rădăcină caracterizat prin prezența a trei sisteme tisulare - epiderma tegumentară cu fire de păr (epiblema), cortexul primar și cilindrul central Epiblema Efectuează aspirarea și funcții de protecție, constă dintr-un singur strat de celule cu pereți subțiri, strâns împachetate, cu peri de rădăcină, nu are stome, o cuticulă groasă și tricomi. Cortexul primar, Situat sub eliblema, este împărțit în exoderm, mezoderm și endoderm. Exoderm Este format din 3-4 straturi de celule mari, bine închise, cu membrane parțial suberizate. Îndeplinește funcții de protecție și de sprijin. mezodermul- parte de depozitare multistrat, alcătuiește cea mai mare parte a crustei primare. Mezodermul îndeplinește și funcții de transport și pneumatice. Endodermul- stratul cel mai interior, de obicei cu un singur rând al cortexului primar, mărginind cilindrul central, îndeplinind o funcție de susținere. Central, Sau Axial, cilindru Ocupă partea de mijloc a rădăcinii, înconjurată de un inel de periciclu din care se formează rădăcinile laterale. Nu există un miez real în rădăcină. În centru se pot forma vase, sclerenchim și uneori parenchim de depozitare Miez fals Structura rădăcină secundară se formează datorită apariţiei şi funcţionării meristemelor laterale secundare – cambiul şi felogenul. Cambiumul se formează din periciclul de deasupra razelor xilemului primar şi din procambium sau parenchim sub floem. Cambiumul de divizare depune floemul secundar spre exterior, iar xilemul secundar spre centru. Xilemul primar este împins spre axa rădăcinii, iar floemul primar este împins spre periferia cilindrului central. Structura secundară a rădăcinii poate fi cu smocuri sau fără smocuri Tipul fasciculului Se formează dacă cambiul interfascicular, format deasupra razelor xilemului primar, produce parenchimul razelor medulare, iar cambiul fascicular aflat sub floem produce floemul secundar și xilemul fasciculului vascular colateral sau bicolateral. La Tip fără fascicule Structura inelului cambium formează inele continue de xilem și floem. Transformările secundare din cortexul radicular primar sunt asociate cu activitatea felogenului. Dacă apare din periciclu, atunci dopul format spre exterior izolează cortexul primar și este rupt și desprins. Dacă felogenul se formează din parenchimul mezodermului, ceea ce este rar, atunci partea cortexului primar se află sub felogen. se păstrează. La plantele lemnoase, rădăcina este de obicei de tip fără smocuri, iar în lemn se formează inele de creștere anuală. Anual Inel este o colecție de țesuturi formată de cambium în timpul unui sezon de vegetație. material de acoperire Rădăcinile copacilor pot avea periderm sau crustă. Prezența în centrul organului a unui fascicul vascular radial, a sclerenchimului sau a vaselor de xilem primar distinge rădăcina de tulpină și rizom, în care partea centrală este ocupată de miezul adevărat.
Această structură, când în cilindrul central există un fascicul radial compus din xilem și floem, se numește primar.
La plantele monocotiledonate aceasta structura primara Rădăcina durează toată viața. Pe măsură ce rădăcina se dezvoltă și crește, are loc lignificarea celulelor endodermice și periciclice. Rădăcinile monocotiledonelor nu sunt capabile să se îngroașe, deoarece fasciculele lor vasculare sunt închise și nu au un cambium.
La plantele cu flori dicotiledonate, precum și la conifere și alte gimnosperme, rădăcinile sunt capabile să se îngroașe. În mănunchiul lor, se formează un cambium, care depune elemente de xilem și floem. Depunerea de noi elemente secundare duce la modificări în compoziția și structura anatomică a secțiunilor ulterioare ale rădăcinii. Ca rezultat al activității cambiumului, o structură radiculară numită secundar. Cambiumul apare în țesutul parenchimatos sub fasciculele floemului și deasupra razelor xilemului ( "arcuri cambiale"), sau mai degrabă, între xilem și floem se formează o panglică continuă de celule cambiale. Celulele parenchimului capătă un caracter meristematic - se întind și devin tabulare. După ceva timp, se formează un inel solid cambium, având inițial contururi neuniforme, sinuoase (Fig. 63, 2). Celulele Cambium încep să depună elemente ale xilemului secundar spre interior și elemente ale floemului secundar spre exterior. Depunerea de xilem are loc mai rapid; secțiunile floemului primar sunt adesea împinse la periferie, datorită cărora inelul este aliniat. La plantele lemnoase, se depun ulterior inele continue de xilem și floem, iar structura rădăcinii devine destul de asemănătoare cu structura tulpinii. Cu toate acestea, întotdeauna în rădăcină, în partea sa centrală, se păstrează resturi ale xilemului primar - un mănunchi radial sub formă de raze cruciforme. Prin această caracteristică este ușor să distingem rădăcina de tulpină la microscop. Floem primar, în mod natural, este împins în partea cea mai periferică și, datorită volumului său mai mic, nu se distinge la o rădăcină adultă.
Pe măsură ce rădăcina crește, are loc o schimbare în scoarța acesteia. Epiderma este vărsată și înlocuită periderm (plută), care provin din periciclu. Crusta primară este, de asemenea, vărsată. Rădăcinile copacilor foarte vechi dezvoltă o crustă. În acest moment, endodermul, precum și periciclul, moare, iar formarea de noi straturi de periderm are loc în straturile mai profunde ale rădăcinii - în floemul secundar. Astfel, cu o structură secundară, rădăcina nu are cortex, endoderm și periciclu și este acoperită la exterior cu periderm sau crustă (Fig. 63, 3 ).Material de pe site
Pe această pagină există material pe următoarele subiecte: