23. Structura anatomică tulpini de monocotiledone erbacee și lemnoase.
24. Structura anatomică a tulpinilor plantelor erbacee dicotiledonate.
25. Structura anatomică a tulpinilor plantelor dicotiledonate lemnoase.
26. Structura anatomică a tulpinilor de conifere.
27. Structura anatomică a rizomilor plantelor mono și dicotiledonate.
31. Conceptul de tulpină, lăstar. Funcțiile și metamorfozele tulpinii. Caracteristicile morfologice ale tulpinii.
O tulpină este un organ suprateran, vegetativ, axial al unei plante, având simetrie radială și având o creștere apicală.
Tulpina cu muguri și frunze - evadare. Secțiuni ale tulpinii care poartă frunza - noduri ale tulpinii: între noduri - internoduri. În funcție de lungimea internodurilor, lăstarii pot fi alungiți (cereale) sau scurtați (păpădie).
Lăstarul se dezvoltă din mugure.
Apical mugure - vârful tulpinii, acoperit cu primordii de frunze tinere: un con de creștere, la baza acestuia se află tuberculi primari - din care se formează frunzele primordiale. În axilele tuberculilor primari sunt tuberculi secundari care dau lateral sau axilar rinichii Ramurile laterale cresc și ele la vârfuri și fiecare ramură se termină într-un mugure apical.
Odată cu debutul toamnei, toți mugurii intră în repaus sezonier. Se numesc hibernatori. Ele pot fi vegetativ(formând frunze sau lăstari) și vegetative – generative sau înflorire(înflorește în flori sau inflorescențe primăvara). Protejat de solzi tari acoperiți cu cuticulă. Au sclereide și uneori un dop. Înfloresc datorită rezervelor depuse în rădăcină, tulpină (sau în solzii de acoperire).
Mulți muguri axilari rămân latenți pentru o lungă perioadă de timpși cresc numai cu axa lor pe tulpină în conformitate cu îngroșarea acesteia - dormit rinichii Încep să crească dacă este îndepărtat mugurele apical. Uneori pot fi scufundați în lemn și se trezesc după spargerea sau tăierea trunchiului.
Propoziții subordonate mugurii se formează în noduri și pe internoduri, rădăcini, rizomi, frunze - o rezervă pentru înmulțirea vegetativă. Origine: din cambium, felogen, periciclu, scoarță. Aceștia sunt muguri de întinerire - produc lăstari asemănători cu puieții tineri.
Caracteristicile morfologice ale tulpinii.
eu. După poziție în spațiu tulpinile sunt: erecte, ascendente, târâtoare, cataratoare, crete etc.
II. Ramificarea lăstarilor.
1. Dihotomic - punctul de creștere este bifurcat și împărțit în 2 noi (inferioare, pteridofite, briofite).
2. Monopodial - axa principală crește cu vârful său, din muguri laterali se dezvoltă ramuri laterale, care se ramifică și monopodial (coniferele - trunchiurile sunt foarte apreciate în silvicultură și construcții navale).
3. Simpodial - mugurel apical moare, se dezvoltă în schimb un mugure lateral, deseori crescând în direcție verticală, parcă ar continua creșterea tulpinii principale etc. (mesteacăn, salcie, măr, roșie, cartof).
Cunoscut amestecat ramificare: mai întâi mono-, apoi simpodială (în fructe de pom, bumbac).
Înflorire intensivă număr mare mugurii face ca lăstarul să se scurteze, ceea ce asigură dezvoltarea unui număr mare de frunze pe coroană, adică. suprafață fotosintetică mare.
4. Falsă ramificare dihotomică - sub mugurel apical se formează doi muguri axilari situati opus; incepand sa creasca, formeaza o furculita (vâsc, dopa, cuisoare, liliac, castan de cal).
Studiul sistemelor de ramificare este de mare importanță practică. Recolta poate fi reglementată.
III. Aranjamentul frunzelor: frunzele sunt fixate pe nodurile tulpinii.
1. Următorul lucru este că nodul are o singură frunză - măr, mesteacăn, arțar.
2. Opus - două frunze într-un nod (Labiaceae, valeriană, hortensie).
3. Viculată - trei sau mai multe frunze - oleandru.
4. Rozeta bazala - primula.
Frunzele succesive sunt dispuse în spirală pe tulpină. Urmărindu-i revoluțiile, puteți găsi două foi situate vertical, una deasupra celeilalte. O linie care leagă frunzele în rânduri longitudinale pe o tulpină - poem ortostic. Între capetele ortosticului, frunzele rămase sunt aranjate în spirală numărul de spire ale spiralei între două frunze situate pe aceeași ortostică se numește -; cu frunze ciclu. O formulă pentru următorul aranjament al frunzelor este compilată sub forma unei fracții, unde numărătorul este numărul de rotații ale spiralei în ciclul frunzelor, iar numitorul este numărul de frunze din ciclu.
IV. Forma tulpinii în secțiune transversală.
Forma tulpinii în secțiune transversală poate fi foarte diversă: rotundă, plată, nervură, multifațetată etc.; poate fi gol și umplut, gol și pubescent.
Speranța de viață și grupele biomorfologice de plante. Chiparos - 3000 ani, castan - 2000 ani, stejar - 1200 ani, tei - 1000 ani, plop - 300-600 ani, măr - 200 ani, cireș - 40-100 ani, pin - 50 ani. În funcție de speranța de viață și de tipul de lăstari, plantele sunt împărțite în:
copaci- plante mari cu tulpini perene foarte dezvoltate. Există o creștere secundară.
tufișuri- trunchiul principal este absent sau slab exprimat, ramificarea începe aproape de la suprafața solului (4-6). Barberry, serviceberry, măceș. Există grupuri de arbuști nu mai mari de 1m - cimbru, saxaul.
subarbusti- lăstarii din partea inferioară rămân pereni, în partea superioară - lăstarii anuali îngheață sau se usucă - pelin, astragalus, sunătoare;
ierbos- moartea tulpinilor supraterane se observă la sfârșitul sezonului de vegetație. Majoritatea plantelor de câmp sunt cereale, mazăre, fasole, in, cânepă;
erbacee bienală- în primul an se dezvoltă doar o rozetă de frunze bazale, iar în al doilea se dezvoltă flori, fructe și o tulpină alungită (cicoare, găină, marshmallow, trifoi dulce);
perenă- părțile subterane durabile și moartea anuală a părților supraterane (valeriană, lacramioare, urzică).
Printre plantele erbacee există forme de iarnă și de primăvară:
a) dacă semințele germinează primăvara - primăvara;
b) culturi de iarnă - germinarea semințelor toamna (0-5 grade).
Funcțiile tulpinii:
1. Scheletul care leagă frunzele și rădăcinile între ele dă obiceiul plantei;
2. Conductiv;
3. Depozitare;
4.Sintetic;
5. Organ al reproducerii vegetative.
Metamorfoze:
Subteran: 1. Rizom - un lăstar subteran, frunze sub formă de solzi, dimensiunea rizomilor este determinată de funcții: dacă pentru depozitare - gros (calamus, coada calului, kupena), pentru reproducere - cu muguri de reînnoire (iarbă de grâu, iris ).
2. Tuberculi - formați la capetele tulpinilor subterane - stolloni, au muguri - ochi (3 sau mai mulți).
3. Bulbi - tulpina scurtata - fund, muguri si frunze.
Sol: 4. Spini - păducel, ghimpe.
5. Mustață - dovleac, castraveți, pepene galben.
6. Filocladia - tulpină în formă de frunză - mătură de măcelar.
Tulpina- organ suprateran, vegetativ, axial al plantei; are o creștere pe termen lung în lungime folosind meristemul apical din conul de creștere sau meristemul intercalar; poartă frunze care se formează într-o anumită ordine pe conul de creștere sub formă de tuberculi; se poate ramifica datorita mugurilor situati in frunzele axilare; are o structură radială și mai multe planuri de simetrie.
La capătul de creștere, tulpina poartă un mugure apical - un con de creștere, îmbrăcat cu frunze tinere, modificate. Se termină cu un punct de creștere, în spatele căruia se află tuberculii primari - rudimentele frunzelor. Chiar mai jos, în axile lor, apar tuberculi secundari - rudimentele lăstarilor laterali.
Teoria histogenului lui Gapstein (1818): dermatogen, periblema, plerom - incorect pentru tulpină. La începutul anilor 20 ai secolului nostru, Schmidt a formulat teoria tunicii și a corpului, conform căreia conul de creștere la angiosperme și o parte a gimnospermelor este format din două forme histologice. diferite părți, având activitate meristemă diferită. Partea exterioară este tunica, celulele acesteia se împart perpendicular pe suprafața tulpinii și formează epiderma, uneori epiderma și mai multe straturi sau întreg cortexul primar; partea interioară este corpul - celulele se divid în toate direcțiile și formează straturile rămase ale cortexului primar și ale sistemului nervos central.
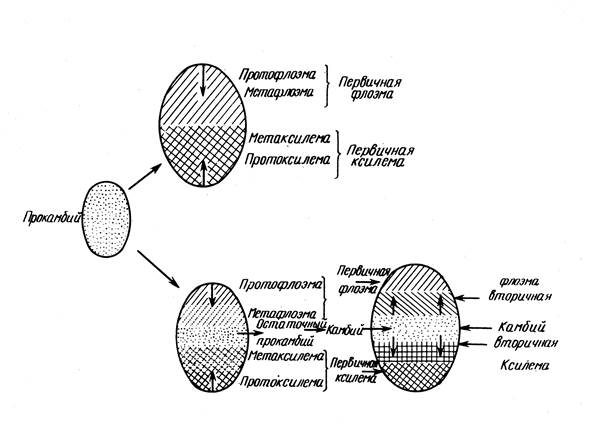
Structura primară a tulpinii se dezvoltă în timpul diferențierii celulelor meristemului apical.
La nivelul primordiilor primelor frunze, celulele tunicii și ale corpului încetează să se divizeze; partea lor periferică merge la formarea cortexului primar, partea interioară la formarea miezului. Dar între ele există mai multe rânduri de celule meristematice active dispuse într-un inel - inel educativ. Celulele sale de la baza primordiilor de frunze tinere dau naștere meristemului lateral primar - procambium.
Procambium reprezintă faza embrionară în dezvoltarea meristemului conducător primar, prin urmare localizarea acestuia determină în mare măsură localizarea ulterioară a sistemului conducător. Dacă este așezat într-un strat continuu (inel), atunci apar straturi continue de floem și xilem, dar dacă procambium este așezat sub formă de mănunchiuri (toroane), atunci țesuturile conductoare primare sunt aranjate sub formă de mănunchiuri. . Celulele straturilor periferice ale inelului educațional, care nu participă la formarea procambiului, formează un periciclu.
Procambium ia naștere la baza primordiului frunzei și de aici dezvoltarea sa se răspândește în două direcții - acropetal, adică. până la vârful primordiului frunzei și basipetal, adică. jos pe tulpină, unde se unește cu alte mănunchiuri care au apărut mai devreme.
Floemul se formează înaintea xilemului și se dezvoltă centripet, mai întâi protofloemul, apoi protoxilemul (vasele inelate și spiralate), apoi metaxilemul. În acest fel, se formează COC al structurii primare.
Pe o secțiune transversală a tulpinii, se disting trei grupuri de țesuturi: tegumentar, cortex primar și COC.
Țesutul tegumentar al tulpinii plantă erbacee dicotiledonată- epiderma. Celulele sunt ușor întortocheate, alungite în direcția tulpinii, cu puține sau deloc stomatele.
Scoarța primară este formată din colenchim sub formă de fire în colțurile tulpinii la Lamiaceae sau în coastele acesteia la Apiaceae, mai rar sub formă de inel la Cucurbitaceae și Solanaceae. La periferie, cortexul primar este format din benzi alternante de clorenchim si tesut mecanic. Partea interioară are parenchim incolor. Stratul cel mai interior este endodermul, în unele cazuri, este o înveliș purtător de amidon care conține granule mici de amidon protejat.
Partea periferică cilindru central- periciclu. Țesuturile conductoare sunt aranjate sub formă de mănunchiuri sau un inel continuu (în funcție de localizarea procambiului). Trecerea la structura secundară este asociată cu formarea cambiului, în funcție de care se disting următoarele tipuri de structură a tulpinii:
1. Procambiul este format într-un inel continuu, structura primară este nefasciculară; cambiul se formează din procambium; secundar - non-grindă.
2. Procambium este așezat prin corzi - structura primara grindă; Cambiumul se formează din procambium din fascicule și din țesutul de pământ al CoC dintre fascicule. Se formează un inel cambial continuu: structura secundară este nefasciculară.
3. Structura primară și secundară este grupată, deoarece nu se formează cambium interfascicular.
4. Structura primară și secundară este grupată, deoarece Procambiul este așezat în fire separate, iar cambiul interfascicular rezultat formează țesutul principal al C.O.C., și nu floemul și xilemul.
Tulpina plantelor erbacee monocotiledonate. Tulpina plantelor erbacee monocotiledonate se caracterizează printr-o structură de fascicul, smocuri sunt localizate aleatoriu și nu există o creștere secundară. Principalele caracteristici ale structurii anatomice a tulpinilor sunt determinate de sistemul de urme de frunze: de la baza frunzei, fasciculele intră în tulpină, situate de-a lungul întregii circumferințe, astfel toate fasciculele vasculare de monocotiledone sunt urme de frunze (tip palmier) . SVP-urile de monocotiledone au o formă caracteristică: două vase cu vârf mari (simetric) și 1-2 mai înguste cu îngroșări inelate și spiralate. Adiacent celei mai exterioare dintre ele este o cavitate de aer formată la locul distrugerii elementelor protoxilemului. Dimensiunea grinzilor crește de la periferie la centru.
Există diferențe în structura cortexului:
1. Monocotiledonee cu scoarța primară pronunțată - (sparaanghel): este format din mai multe straturi de celule purtătoare de clorofilă; colenchimul este absent.
2. La alte monocotiledone (palmier, cereale) nu se exprimă scoarța primară; imediat sub epidermă există zone de clorenchim înconjurate de sclerenchim, care se contopește cu sclerenchimul SVP.
În internodurile multor monocotiledone, se formează o cavitate centrală mare (paie de iarbă în aceste cazuri, fasciculele sunt deplasate la periferie, dar sunt și împrăștiate: cele mai mici - la periferie, cele mari - la cavitate).
Structura tulpinilor dicotiledonatelor lemnoase. Structura primară este non-bundle. Secundar este asociat cu formarea de felogen și cambium. Cambiumul formează floem și xilem secundar. Floem - floem - dur și moale, situat sub formă de trapeze, despărțiți de vârfurile razelor medulare primare; Funcția de conducere este îndeplinită de alburn - 1 mm la cambium. Lemn sub formă de inele de creștere; duramen - functie mecanica.
Miezul este un țesut liber, inițial viu, cu o cantitate de nutrienți. Conectat cu floem și scoarță prin razele medulare (primare și secundare)
Felogenul este format din epidermă și straturile cortexului primar, formează peridermul și crusta. Sub acesta, cortexul primar este păstrat - collenchim, clorenchim, țesut de depozitare - endodermul nu este exprimat (uneori diferențiat într-o teacă purtătoare de amidon).
Conform GF X1 (1987), scoarța în practica farmaceutică- aceasta este partea exterioară a trunchiurilor, ramurilor și rădăcinilor copacilor și arbuștilor, situată la periferia cambiumului. Prin semne exterioareÎntreaga scoarță arată ca bucăți tubulare, canelate sau plate de diferite dimensiuni. Suprafața exterioară a scoarței cu dopuri maro sau gri este de obicei netedă sau cu riduri longitudinale (sau transversale), uneori cu crăpături. Scoarța ramurilor și a trunchiurilor are linte rotundă sau alungită, uneori pot exista licheni de frunze pe ea (trebuie îndepărtate la recoltare). Suprafața interioară a scoarței este de obicei mai ușoară, netedă sau cu nervuri. Fractura transversală este de obicei neuniformă: așchiată, fibroasă sau granulară.
Pe un microdiapozitiv acordați atenție cortexului extern, situat la periferie de la capătul razelor medulare și format din periderm și resturi ale cortexului primar, iar cel intern, format din floem. Grosimea, culoarea și natura dopului, prezența colenchimului, raportul dintre grosimea cortexului primar și secundar, lățimea razelor medulare, numărul, locația și structura fibrelor libiene, celulele pietroase, precum și incluziunile de oxalat de calciu, laticifere, celule cu ulei esențial sunt de importanță diagnostică.
Structura tulpinilor de conifere. Cel primar este grupat, apoi se formează un inel cambial continuu și inele de floem și xilem. Fellogenul este depus.
În partea crustală: nu există colenchim, există canale de rășină.
2. Nu există celule însoțitoare, fibre libiene sau parenchim libien în floem.
3. Cambium multirând.
Lemnul este format din traheide, lipsesc fibrele mecanice și parenchimul lemnos.
Structura primară este fasciculară.
Structura tulpinilor de monocotiledone lemnoase. Caracteristică reprezentanților asemănătoare copacilor din familia crinului (dracaena, yucca, aloe). Tulpina tânără din apropierea conului de creștere este construită ca toate monocotiledonele: țesut tegumentar - epidermă; sub acesta se află cortexul primar, reprezentat de mai multe straturi de colenchim lamelar; COC începe cu periciclul și conține SVP-uri colaterale închise, aranjate aleatoriu. Îngroșarea are loc datorită formării unui inel de îngroșare care provine din periciclu sau din stratul interior al cortexului primar. Formează centrifug SVP concentrice centoxylem. Materialul de acoperire este din plută.
Structura rizomilor de monocotiledone. Similar cu tulpina unui copac. Țesutul tegumentar este exodermul lignificat; cortexul primar este întotdeauna dezvoltat, reprezentat de țesut de depozitare; endoderm tipic cu pete caspariene și îngroșări în formă de potcoavă. În COC există fascicule colaterale (V) și fascicule centrifloem concentrice. Format dintr-un inel de îngroșare.
Structura rizomilor dicotiledonați.Țesutul tegumentar este un dop, peridermul este format din felogen; cortexul primar - parenchim de stocare; endoderm cu pete caspariene sau în formă de potcoavă; în COC, aranjarea țesuturilor este similară cu tulpina de pământ; pentru tipul fasciculului - fascicul; pentru tranziție și fără grindă - fără grindă.
Evoluția stelei. Pe parcursul procesului de evoluție s-au observat modificări în structura și poziția relativă a diferitelor țesuturi vegetale, în special a țesuturilor conductoare. Au fost stabilite mai multe tipuri evolutive de structură a tulpinii, referitoare la COC sau stele (Van Tighem - botanist francez). Teoria stelară a primit ulterior o confirmare strălucitoare în studiul plantelor dispărute (rinofite). Cel mai vechi tip este protostele. Un fir de xilem acoperit cu un strat de floem, actinostele - xilemul are forma unei stele (mușchi de mușchi, coada-calului) înconjurat de floem
Sifonostele - apare miezul.
Dictyostele - sifonostele este disecat în mănunchiuri separate (ferigi).
Eustela - pachete colaterale deschise (bipartite).
Ataktostele - în monocotiledone, fasciculele colaterale închise sunt localizate aleatoriu.
Concluzie. Varietatea de tipuri de structură a tulpinii și a rizomilor trebuie cunoscută pentru diagnosticul macro și microscopic al materialelor plantelor medicinale.
Tulpina face parte din materialul vegetal medicinal – iarba (Herba). Pentru diagnosticul macroscopic plantele medicinale, în conformitate cu Farmacopeea de stat X1 (1987), notează următoarele caracteristici ale tulpinii: tipul de ramificare, forma secțiunii transversale, pubescența, dimensiunile (lungime și diametru la bază), aranjarea frunzelor. Pe micropreparate Se notează secțiunea transversală a tulpinii, caracteristicile structurale ale celulelor epidermice, localizarea fasciculelor vasculare, prezența și natura aranjamentului țesuturilor mecanice, incluziuni cristaline, recipiente, canale secretoare, laticifere și alte caracteristici ale planului de diagnostic.
Rizomi (Rhizomata) analizați prin semne externe diferă ca formă (simple sau ramificate, cilindrice sau ovale, limpede, drepte, curbate sau răsucite etc.), caracteristicile suprafeței exterioare (netede sau șifonate cu urme de rădăcini îndepărtate), natura fracturii (netedă, granulare, așchiate sau fibroase). Uneori, natura aranjamentului fasciculelor conductoare este examinată cu ochiul liber la o fractură. Pentru diagnosticul microscopic notați structura fără fascicule sau fascicule, caracteristici ale locației și tipuri de fascicule, structură țesut acoperit, depozitare și miez, prezența diferitelor recipiente, elemente mecanice, canale, lacticifere, cristale de oxalat de calciu, nutrienți de rezervă (amidon, mucus, inulină, ulei gras etc.).
Când se analizează bulbi (Bulbi), tuberculi (Tubera) și cormi (Bulbotubera) forma (sferică, ovoidă, ovală, alungită, turtită, fusiformă etc.), mărimea, natura suprafeței, caracteristicile structurale ale solzilor (uscate sau suculent) contează. La microscopic se notează diagnosticul, caracteristicile țesutului de depozitare și locația fasciculelor.
Structura tulpinii plantelor erbacee dicotiledonate
În stadiile incipiente de dezvoltare, tulpina dicotiledonelor are o structură primară. Ca urmare a activităților meristeme primare conurile de creștere formează epiderma, cortexul primar, cilindrul central și miezul.
Epidermă Tulpina are un număr mic de stomi, de obicei acoperite cu o cuticulă. Sub epidermă sunt țesuturi cortexul primar. Stratul său exterior (exodermul) este adesea format din colenchim, care este situat fie într-un inel continuu (floarea soarelui), fie în secțiuni separate. La plantele cu tulpini nervurate (dovleac), colenchimul umple proeminențele tulpinii. La tulpinile subțiri, colenchimul este de obicei unghiular, la tulpinile cu îngroșare secundară activă este lamelar. Partea principală a cortexului primar (mezoderm) este compusă din clorenchim și parenchim. Aici se pot dezvolta cavități de aer și recipiente de secreție. Celulele stratului interior al cortexului primar (endoderm) sunt de obicei umplute cu boabe mari de amidon (teaca de amidon).
Cilindru central(stela) constă dintr-un periciclu și un sistem conducător. Miez - din parenchim, care este adesea distrus, iar în locul lui se formează o cavitate de aer. Periciclul este reprezentat de unul sau mai multe rânduri de sclerenchim sau firele sale individuale. Sistemul conducător are forma unui cilindru gol care separă cortexul primar și miezul. Fasciculele conductoare care alcătuiesc acest cilindru sunt separate prin straturi de parenchim interfascicular - raze medulare care leagă miezul și cortexul primar. Cele mai multe dicotiledonate sunt caracterizate printr-un tip special de cilindru central - eustelle, într-o secțiune transversală a căruia fasciculele sunt dispuse într-un inel (Fig. 4).
Figura 4. Eustele dicotiledonate.
U plante dicotiledonate stratul de celule procambium dintre floemul primar și xilem păstrează capacitatea de a se diviza și se transformă în secundar țesătură educațională - cambium, care se divide, depunând elemente de xilem secundar în interior și floem secundar la periferie. Ciorchinii de plante dicotiledonate sunt deschiși datorită lucrului cambiumului, ciorchinul crește și diametrul acestuia crește. Activitatea cambiumului asigură apariția elementelor secundare în tulpină, adică trecerea de la structura anatomică primară la cea secundară.
Structura secundară este caracteristică tuturor plantelor dicotiledonate și conifere - ierburi și copaci. Varietatea tipurilor de structuri se datorează, în primul rând, aranjamentului țesuturilor conducătoare, care este determinată de formarea procambiumului și de activitatea cambiului.
În timpul procesului de diferențiere a conului de creștere, procambiul se formează sub formă de fire. Dacă firele sunt separate de șiruri suficient de largi de parenchim, atunci se formează un mănunchi sau o structură de tranziție a tulpinii; dacă firele de procambium sunt reunite atât de aproape încât se îmbină într-un cilindru, atunci se formează o structură negrupată (solidă).
1. Structura mănunchiului tulpinii intalnita la unele plante erbacee (marar, trifoi, ranuncul, mazare, lucerna). Sunt localizate cordoanele procambiale înglobate în conul de creștere într-un cerc în jurul periferiei cilindrului central. Fiecare cordon procambial se transformă într-un mănunchi colateral deschis format din xilem primar, floem primar și o bandă de cambium între ele. Celulele Cambium, divându-se, dau elemente noi (secundare) ale fasciculului vascular: spre interior - xilem spre periferie - floem. În acest caz, cele mai tinere secțiuni de floem și xilem sunt adiacente cambiului, iar cele mai vechi se deplasează la periferia mănunchiului. Cel mai mult pozitii extreme floemul primar şi xilemul vor ocupa. Mănunchiurile conducătoare sunt separate prin zone largi de interfase și înconjoară un miez puternic.
La majoritatea plantelor, cambiul funcționează activ, iar diametrul mănunchiului crește semnificativ. Meristemele fasciculare și interfasciculare se închid și formează un inel cambial continuu, a cărui activitate asigură îngroșarea uniformă a tulpinii. Dacă cambiul fascicular produce elemente de xilem secundar și floem, atunci cambiul interfascicular produce parenchim (în mărar, pui, etc.). La trifoi, lucernă și alte plante, cambiul interfascicular depune predominant celule asemănătoare sclerenchimului spre xilem. Aceste celule sunt alungite, cu pereți groși, lignificate. Prin urmare, întârzierea recoltării trifoiului reduce calitatea fânului. Structura fasciculului distinctă este menținută la aceste plante pe toată durata vieții tulpinii.
Structura mănunchiului are o tulpină trifoi. Sub epidermă se află cortexul primar, reprezentat de parenchim purtător de clorofilă. Rândul exterior al celulelor sale este colenchimul lamelar; rândul interior este celulele tecii de amidon (endoderm). Granița dintre cortexul primar și cilindrul central este clar vizibilă datorită sclerenchimului de origine periciclică.
Sclerenchimul formează semi-arce deasupra fasciculelor colaterale deschise dispuse în cerc. Pachetul conține floem primar adiacent sclerenchimului, floem secundar, cambium, xilem secundar și xilem primar. Cambiumul fascicular se închide cu cambiul interfascicular, formând un inel meristematic solid. Cambiumul interfascicular depune în interior parenchim lignificat, care leagă fasciculele vasculare. Apare un inel (cilindru) de țesuturi lignificate, oferind o rezistență ridicată tulpinii. Cu toate acestea, țesuturile conductoare au rămas separate sub formă de mănunchiuri. Miezul tulpinii este reprezentat de parenchim.
Structura grupată a tulpinii poate fi înlocuită odată cu vârsta cu una solidă fără smocuri. 2. Structura de tranziție a tulpinii tipic pentru floarea soarelui. La început, tulpina se formează în același mod ca la trifoi, dar cambiul interfascicular nu depune parenchim, ci xilem și floem. Se formează mănunchiuri conducătoare suplimentare. Treptat, toate pachetele se pot îmbina într-un cilindru continuu cu trei straturi (xilem, cambium, floem). În partea superioară (tânără) tulpina are o structură mănunchiată, la bază (în partea veche) are o structură fără grinzi..
3. Structură fără pachet caracteristic tulpinilor multor ierburi și copaci. În conul de creștere al acestor plante, firele procambiale sunt atât de apropiate între ele încât formează un cilindru aproape continuu (într-o secțiune transversală arată ca un inel). Cilindrul (inelul) procambial, diferențierea, dă un cilindru de xilem primar la centru, un cilindru de floem primar la periferie, iar între ele se formează un cilindru cambial.
Modificările secundare sunt asociate cu munca cambiului. Celulele Cambium se divid paralel cu suprafața tulpinii, în timp ce în interior sunt depuse de 10...20 de ori mai multe celule decât afară. Cambiumul formează elemente de xilem secundar (lemn) în interior, iar floemul secundar (bast) este depus în exterior. În unele locuri, cambiul depune celule parenchimoase ale razelor medulare pe ambele părți.
Caracteristicile structurii fără smocuri a tulpinilor de iarbă pot fi luate în considerare folosind exemplul unei tulpini in. Tulpina este acoperită cu epidermă, sub care există clorenchim liber al cortexului primar. Cantitate mare stomatele din epidermă și absența colenchimului în cortexul primar au o semnificație semnificație practică, deoarece în timpul curățării tehnice a inului, apa nu întâlnește obstacole și pătrunde ușor în interiorul tulpinii. Celulele tecii de amidon (endoderm) sunt mai mari și conțin boabe de amidon clar vizibile.
Cilindrul central adiacent endodermului începe cu grupuri de celule sclerenchimatoase - fibre libiene, între care se află celulele parenchimului. Fibrele libiene apar în periciclu și în floemul primar. Se remarcă prin pereți foarte groși, de obicei nelignificați, formați din celuloză 75...90%. Lungimea fibrelor de liben este de 4...60 mm, diametrul 12...37 microni. În timpul creșterii unei celule de sclerenchim, nucleul acesteia se divide, dar citokineza nu are loc, iar protoplastul se dovedește a fi multinucleat. Peretele celular este foarte îngroșat, cavitatea celulară seamănă cu un canal îngust. Peretele celular este format din multe straturi de celuloză și este aproape lipsit de pori. O celulă matură din fibre libiene este moartă. Protoplast după îngroșare peretele celular este distrus. Fibrele de bast au o rezistență neobișnuit de mare. Rezistența lor la tracțiune corespunde cu cea a oțelului, iar elasticitatea lor este mai mare. Celulele din fibre de bast sunt foarte strâns legate între ele. Fibra de in este o materie primă excelentă de filare folosită în industria textilă.
În continuare, cilindrii (inele) sunt amplasați în ordine secvențială: floem secundar, cambium, xilem secundar cu raze medulare clar vizibile. Xilemul primar este situat în zonele adiacente mudofului. Parenchimul miezului este distrus rapid (în procesul de alungire a tulpinii), formând o cavitate centrală.
Tulpinile plantelor erbacee dicotiledonate se caracterizează prin:
1. structura secundara, care apar foarte devreme după primar;
2. prezența țesutului tegumentar primar - epiderma;
3. cortexul primar dezvoltat, împărțit în colenchim (exoderm), parenchim (mezoderm) și înveliș de amidon (endoderm);
4. structură cu grinzi sau fără grinzi (solide);
5. aranjarea corectă a fasciculelor vasculare colaterale sau bicolaterale într-un cerc de-a lungul periferiei cilindrului central;
6. prezența cambiumului (mănunchiuri deschise),
7. nucleul este bine dezvoltat.
Structura anatomică primară a tulpinii.
Definiția și funcțiile tulpinii.
Tulpina este un organ axial, de obicei simetric radial, cu creștere apicală (apicală) pe termen lung.
Funcțiile tulpinii:
1. efectuează deplasarea în două sensuri a substanțelor între rădăcini și frunze,
2. susține coroana plantei,
3. ajuta la cresterea suprafetei totale de asimilare a plantei datorita ramificarii,
4. participă la depozitarea substanțelor de rezervă,
5. la o vârstă fragedă – realizează și fotosinteza.
Aceste funcții determină prezența în tulpină a țesuturilor mecanice tegumentare, conductoare bine dezvoltate și a parenchimului dezvoltat funcțional.
Structura conului de creștere.
În conul de creștere al tulpinii angiospermelor, sunt exprimate două zone: exterior - tunicăși intern - cadru, care diferă în planurile diviziunii celulare (teoria lui A. Schmidt și J. Buder) (Fig. 1).
Tunica poate fi formată din unul sau mai multe straturi de celule care se divid perpendicular de suprafață, la majoritatea plantelor dicotiledonate este în două straturi. Stratul exterior al tunicii dă naștere protoderma, din care se formează apoi epiderma, acoperind frunzele și tulpina. Dacă tunica este multistratificată, atunci stratul interior formează țesutul cortexului primar.
În prezența unei tunici cu un singur strat, întreg cortexul primar și stela sunt formate de corpus. Cadru format dintr-o masă de celule care sunt împărțite în toate planurile.
Primordiile frunzelor apar cel mai adesea în al doilea strat al tunicii, iar mugurii axilari se formează în corp.
Figura 1. b) vârful angiospermelor.
Ca urmare a activității meristemelor primare ale apexului, se formează structura anatomică primară a tulpinii: epiderma, cortexul primar, cilindrul central și miezul.
Îngroșarea secundară apare datorită activității cambiului.
Structura anatomică primară a tulpinii.
Structura tulpinii formată ca urmare a activității meristemelor primare ale apexului se numește primar. În tulpină, cu structura primară, se pot distinge trei zone anatomice și topografice:
1. țesut acoperit,
2. cortexul primar
3. cilindru central.
Epiderma protejează tulpina de evaporare.
Sub epidermă se află stratul exterior al cortexului primar - exoderma - reprezentată de clorenchim şi (în dicotiledonate) colenchim. În stratul interior care mărginește cilindrul central - endoderm - de obicei, boabele de amidon se acumulează, iar apoi se transformă în așa-numitul vagin purtător de amidon (strat purtător de amidon). Boabele de amidon se pot depune în celule sub influența gravitației. Din acest motiv, endodermul joacă un rol important în reacția geotropică ((din grecescul trpos - întoarcere, direcție), mișcări de creștere a tulpinilor plantelor cauzate de acțiunea direcțională a gravitației) a tulpinilor.
De-a lungul periferiei cilindrului central în periciclu se localizează sclerenchimul. Fibrele sclerenchimului formează un strat continuu sau sunt colectate în apropierea fasciculelor vasculare, în exteriorul floemului primar. Element principal cilindru central (stela) - fascicule conductoare. Sunt formate din fire procambiale și constau din xilem primar și floem. Diferențierea procambium are loc centripet: floemul se formează din partea exterioară a fasciculului spre centrul acestuia, iar xilemul se formează spre acesta (Fig. 2). Procambiul poate fi complet transformat în complexe conductoare primare, formând un mănunchi colateral închis (în monocotiledone), sau celulele rămase în centrul fasciculului formează cambium. Astfel de ciorchini deschisi sunt caracteristice plantelor dicotiledonate. Lucrarea cambiului asigură formarea xilemului și floemului secundar.
Figura 2.
Formarea procambiumului determină structura sistemului conducător:
1. Dacă procambium formează un cilindru gol, solid, atunci se formează din acesta cilindri imbricați de xilem și floem (în unele conifere și dicotiledone) - structură fără grinzi.
2. Dacă procambiul constă din fire separate (în majoritatea monocotiledonelor, multe dicotiledonate și conifere), atunci țesuturile conductoare primare emergente păstrează aceleași structura pachetului.
Procambiul ia naștere la baza primordiului frunzei, iar dezvoltarea sa se desfășoară în două direcții: până la vârful primordiului frunzei și în jos pe tulpină, unde se conectează cu firele care au apărut mai devreme. In aceeasi secvența mergeși diferențierea floemului și xilemului. De obicei, se formează mai întâi elementele floem, apoi xilemul. fasciculele conductoare sunt situate în parenchimul cilindrului central.
În centrul tulpinii se dezvoltă o midă clar vizibilă. Celulele parenchimatoase cu pereți subțiri ale miezului se slăbesc adesea, mor și se umplu cu aer, apoi întreg țesutul apare alb (în floarea soarelui, porumb, soc). Uneori, miezul moare foarte devreme, când alungirea internodurilor nu s-a terminat încă. Aceasta implică ruperea miezului și formarea unei tulpini goale.
Structura primară a tulpinii este păstrată în monocotiledone pe tot parcursul vieții, iar în dicotiledonate și gimnosperme - numai în fazele timpurii ale dezvoltării sale.
Structura anatomică a tulpinii unei plante dicotiledonate cu o structură de mănunchi
Tulpina reprezintă axa lăstarului. Împreună cu frunza, este partea structurală principală a lăstarului. Principalele funcții sunt suport și conducere. Prin tulpină există o legătură între rădăcini și frunze și schimbul de aer și produse nutritive minerale. În plus, nutrienții de rezervă sunt adesea depozitați în tulpină. Tulpina, la fel ca întregul lăstar în ansamblu, este un sistem de creștere „deschis”, adică. crește mult timp, iar pe ea apar frunze noi.
Tulpina, ca parte a lăstarului, are un sistem de meristeme care susțin creșterea țesutului în lungime și grosime. Creșterea în lungime se realizează datorită meristemelor apicale și intercalare, iar în grosime în dicotiledonate datorită meristemelor secundare laterale - cambium și felogen.
Structura primară a tulpinii se dezvoltă pe măsură ce celulele meristemului apical lăstarilor se diferențiază. Meristemul apical al unui lăstar dicotiledonat se diferențiază destul de devreme în mai multe grupuri de celule care diferă prin caracteristicile diviziunii și gradul de activitate meristematică. Straturile sale exterioare sunt transformate în protoderm, ale cărui celule formează ulterior țesutul tegumentar primar - epiderma.
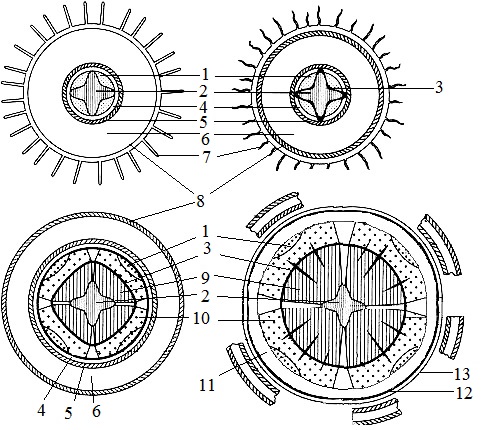
Orez. 1. Tip smoc cu cambium interfascicular în tulpina kirkazonei (Aristolochia clematitis) într-o secțiune transversală (I) și o diagramă a structurii tulpinii la diferite niveluri (II)
A - sectiune la nivelul aspectului procambium; B - la nivelul aspectului cambiului; B - la nivelul structurii formate. 1 - procambium, 2 - epidermă, 3 - colenchim, 4 - parenchim al cortexului, 5 - endoderm (3-5 - cortex primar), 6 - sclerenchim al periciclului, 7 - floem, 8 - xilem, 9 - cambium fascicular (7-9 - mănunchi colateral deschis), 10 - cambium interfascicular,11 - raza medulară, 12 - parenchim medular (6-12 - cilindru central)
La nivelul bazelor primordiilor prime frunze, celulele meristemului apical, situate spre periferie și în centrul apexului, încetează să se divizeze activ, cresc în dimensiune și se vacuolează. Din aceste celule se formează cortexul primar și miduful. În multe dicotiledonate, în inelul inițial se diferențiază un cerc de cordoane procambiale izolate unele de altele. Celulele inelului inițial situat între aceste catene se diferențiază ulterior în elemente parenchimatoase. În secțiuni transversale, aceste raze arată ca niște dungi radiale care leagă miezul cu cortexul primar.
Procambiul dintr-un număr de dicotiledonate se poate forma și sub forma unui inel continuu. Se poate dezvolta pe toată grosimea inelului inițial sau se poate forma dintr-o parte a acestuia. Procambium este precursorul țesuturilor conductoare primare: floem primar și xilem primar. Floemul începe să se formeze mai devreme și se formează în părțile exterioare ale cordurilor procambiale sau inelului procambial. Floemul se dezvoltă centripet, adică. chiar primele elemente ocupă poziția exterioară, iar cele mai recente ocupă poziția interioară. Xilemul se formează în părțile interne ale procambiumului și se dezvoltă centrifug. Astfel, floemul și xilemul se formează unul față de celălalt. Primele elemente ale xilemului sunt proxilemele, vase înguste, cu pereți relativ subțiri sau traheide cu îngroșări secundare spiralate sau inelate. Metaxilemul se formează ceva mai târziu decât protoxilemul și este format din vase scalariforme și poroase. Principalul factor care controlează diferențierea atât a floemului, cât și a xilemului este fitohormonul auxina, produs de primordiile frunzelor și care se deplasează de-a lungul procambiumului de la vârf la bază. Floemul primar și xilemul formați din procambium formează baza cilindrului axial sau stelei.
Stela, care ocupă partea centrală a tulpinii, este formată din țesuturi conducătoare, miezul, periciclul și acele țesuturi permanente care iau naștere din aceasta. În exteriorul reciclării se află cortexul primar, care constă din parenchim, adesea colenchim și uneori elemente secretoare. Miezul constă de obicei din celule de parenchimat cu pereți relativ subțiri. Nutrienții de rezervă sunt adesea depozitați în miez. Idioblastele se găsesc adesea aici, de exemplu. celule individuale umplute cu taninuri, cristale, mucus etc. Uneori, o parte a miezului este distrusă și se formează o cavitate. Partea periferică a miduvei adiacentă xilemului se numește zonă perimedulară.
Îngroșarea secundară a tulpinii la majoritatea dicotiledonatelor are loc destul de devreme, ceea ce duce la formarea unui corp secundar al plantei. Aceste modificări sunt asociate în principal cu activitatea meristemului secundar lateral - cambiul și parțial a unui alt meristem secundar - felogenul. Datorită apariției țesuturilor secundare, plantele dicotiledonate cresc în grosime. Modificările secundare ale cilindrului central încep cu formarea cambiului. Cambiumul ia naștere din resturile procambiului, la limita xilemului primar și a floemului.
Celulele Cambium sunt foarte vacuolate și alungite pe direcție verticală sau orizontală. Primele - inițialele fuziforme - dau naștere la elemente conductoare ale țesuturilor conductoare, a doua - inițialele de rază - formează celule de raze orientate orizontal ale razelor medulare. Celulele inițiale ale cambiului sunt capabile de două tipuri de diviziune - periclinală și anticlinală. În primul caz, placa celulară este așezată paralel cu suprafața tulpinii, în al doilea - perpendicular. Ca urmare, apare o serie continuă de celule derivate, extinzându-se de la cambium radial spre exterior și spre interior. Celulele depuse spre suprafața exterioară și tulpina se diferențiază treptat în floem secundar, iar spre miez - în xilem secundar.
Activitatea cambiumului este activata de fitohormonii giberelina si auxina, proveniti din muguri si frunze tinere. În cele din urmă, în structura tulpinii unui lăstar anual al unei plante dicotiledonate, este posibil să se distingă un cilindru central modificat, care include țesuturi permanente care provin din periciclu, rămășițe de floem primar și secundar, cambium, secundar și resturi de primar. xilema si midula. Cilindrul central modificat este înconjurat de resturi ale cortexului primar.
Structura secundară a rădăciniiRădăcina este organul principal planta superioara. Îndeplinește funcția de nutriție cu minerale și apă. O altă funcție importantă este asigurarea, „ancorarea” plantei în sol. Prin rădăcini, plantele absorb apa și ionii minerali dizolvați în ea din sol. Biosinteza unui număr de metaboliți secundari, în special alcaloizi, se realizează și în rădăcini. Rădăcina este capabilă de metamorfoză. Cel mai adesea este un loc pentru depozitarea pieselor de schimb. nutrienti. Uneori acționează ca un organ respirator și poate servi ca organ de reproducere vegetativă.
Orez. 2. Trecerea de la structura primară a rădăcinii la cea secundară
Educaţie structura secundara Creșterea rădăcinii este asociată în primul rând cu activitatea cambiului, care asigură creșterea rădăcinii în grosime. Cambiumul apare mai întâi din celulele parenchimului cu pereți subțiri sub formă de zone deconectate pe partea interioară a firelor floemului dintre razele xilemului primar. Unele secțiuni ale periciclului, situate în exteriorul razelor xilemului primar, capătă în curând activitate cambială. Ca rezultat, se formează un strat cambial continuu.
Cambiumul depune celule xilemice secundare spre centru, iar celulele floemului secundar spre periferie. Celulele cambiului, care se formează în periciclu, formează raze de lumină radiale largi de parenchim, situate între firele de țesut conducător secundar. Aceste raze, numite uneori raze medulare primare, asigură legătura fiziologică dintre partea centrală a rădăcinii și cortexul primar. Mai târziu, se pot forma și raze medulare secundare, „conectând” xilemul secundar și floemul.
Ca urmare a activității cambiului, floemul primar este împins spre exterior, iar „steaua” xilemului primar rămâne în centrul rădăcinii. „Razele” sale persistă mult timp, uneori până la sfârșitul vieții rădăcinii. Pe lângă modificările secundare care apar în cilindrul central, apar modificări semnificative și în cortexul primar. Datorită creșterii rapide a țesuturilor secundare din interior, provocând o îngroșare puternică a rădăcinii, cortexul primar este adesea rupt. Până în acest moment, celulele periciclului, împărțindu-se de-a lungul întregii circumferințe a cilindrului axial, formează o zonă largă de celule parenchim, în partea exterioară a căreia este așezat felogenul, depunând un dop în exterior și feloderm în interior. Pluta izolează cortexul primar de țesuturile conductoare, cortexul moare și se scurge. Celulele felodermice și parenchimul format de periciclu cresc ulterior și formează zona de parenchim care înconjoară țesuturile conductoare. Această zonă este uneori numită cortexul secundar. La exterior, rădăcinile dicotiledonatelor, care au o structură secundară, sunt acoperite cu periderm. Crusta se formează rar, doar pe rădăcinile copacilor bătrâni.
În stadiile incipiente de dezvoltare, tulpina dicotiledonelor are o structură primară. Ca urmare a activității meristemelor primare ale conului de creștere, se formează epiderma, cortexul primar, cilindrul central și miezul.
Epidermă Tulpina are un număr mic de stomi, de obicei acoperite cu o cuticulă.
Sub epidermă sunt țesuturi cortexul primar. Stratul său exterior este adesea format din colenchim, care este situat fie într-un inel continuu (floarea soarelui), fie în secțiuni separate. Partea principală a cortexului primar este compusă din clorenchim și parenchim. Aici se pot dezvolta cavități de aer și recipiente de secreție. Celulele stratului interior al cortexului primar sunt de obicei umplute cu boabe mari de amidon (teaca de amidon).
Cilindru central constă dintr-un periciclu și un sistem conducător. Miez - din parenchim, care este adesea distrus, iar în locul lui se formează o cavitate de aer. Periciclul este reprezentat de unul sau mai multe rânduri de sclerenchim sau firele sale individuale. Sistemul conducător are forma unui cilindru gol care separă cortexul primar și miezul. Fasciculele conductoare care alcătuiesc acest cilindru sunt separate prin straturi de parenchim interfascicular - raze medulare care leagă miezul și cortexul primar. Majoritatea dicotiledonelor sunt caracterizate printr-un tip special de cilindru central, într-o secțiune transversală a căruia fasciculele sunt dispuse într-un inel (Fig.).
Desen. Tulpina plantelor dicotiledonate.
La plantele dicotiledonate, stratul de celule procambium dintre floemul primar și xilem își păstrează capacitatea de a se diviza și se transformă în țesut educațional secundar - cambium, care se divide, depunând elemente de xilem secundar în interior și floem secundar la periferie. Ciorchinii de plante dicotiledonate sunt deschiși datorită lucrului cambiumului, ciorchinul crește și diametrul acestuia crește. Activitatea cambiumului asigură apariția elementelor secundare în tulpină, adică trecerea de la structura anatomică primară la cea secundară.
Structura secundară este caracteristică tuturor plantelor dicotiledonate și conifere - ierburi și copaci. Varietatea tipurilor de structuri se datorează, în primul rând, aranjamentului țesuturilor conducătoare, care este determinată de formarea procambiumului și de activitatea cambiului.
În timpul procesului de diferențiere a conului de creștere, procambiul se formează sub formă de fire. Dacă firele sunt separate de rânduri suficient de largi de parenchim, atunci structură de tulpină grupată sau de tranziție; dacă firele de procambium sunt adunate atât de aproape încât se contopesc într-un cilindru, atunci structură fără grinzi (solide)..
Structura mănunchiului tulpinii intalnita la unele plante erbacee (marar, trifoi, ranuncul, mazare, lucerna). Sunt localizate cordoanele procambiale înglobate în conul de creștere într-un cerc în jurul periferiei cilindrului central. Fiecare cordon procambial se transformă într-un mănunchi colateral deschis format din xilem primar, floem primar și o bandă de cambium între ele. Celulele Cambium, divându-se, dau elemente noi (secundare) ale fasciculului vascular: spre interior - xilem spre periferie - floem. Mănunchiurile conducătoare sunt separate prin zone largi de interfase și înconjoară un miez puternic.
La majoritatea plantelor, cambiul funcționează activ, iar diametrul mănunchiului crește semnificativ. Meristemele fasciculare și interfasciculare se închid și formează un inel cambial continuu., a cărui activitate asigură îngroșarea uniformă a tulpinii. Dacă cambiul fascicular produce elemente de xilem secundar și floem, atunci cambiul interfascicular produce parenchim (în mărar, pui, etc.). La trifoi, lucernă și alte plante, cambiul interfascicular depune predominant celule asemănătoare sclerenchimului spre xilem. Aceste celule sunt alungite, cu pereți groși, lignificate. Prin urmare, întârzierea recoltării trifoiului reduce calitatea fânului. Structura fasciculului distinctă este menținută la aceste plante pe toată durata vieții tulpinii.
Structura grupată a tulpinii poate fi înlocuită odată cu vârsta cu una solidă fără smocuri. Structura de tranziție a tulpinii tipic pentru floarea soarelui. La început, tulpina se formează în același mod ca la trifoi, dar cambiul interfascicular nu depune parenchim, ci xilem și floem. Se formează mănunchiuri conducătoare suplimentare. Treptat, toate pachetele se pot îmbina într-un cilindru continuu cu trei straturi (xilem, cambium, floem). În partea superioară (tânără) tulpina are o structură mănunchiată, la bază (în partea veche) are o structură fără grinzi. .
Structură fără pachet caracteristic tulpinilor multor ierburi și copaci. În conul de creștere al acestor plante, firele procambiale sunt atât de apropiate între ele încât formează un cilindru aproape continuu (într-o secțiune transversală arată ca un inel). Cilindrul (inelul) procambial, diferențierea, dă un cilindru de xilem primar la centru, un cilindru de floem primar la periferie, iar între ele se formează un cilindru cambial.
Modificările secundare sunt asociate cu munca cambiului. Celulele Cambium se divid paralel cu suprafața tulpinii, cu de 10-20 de ori mai multe celule depuse în interior decât în exterior. Cambiumul formează elemente de xilem secundar (lemn) în interior, iar floemul secundar (bast) este depus în exterior. În unele locuri, cambiul depune celule parenchimoase ale razelor medulare pe ambele părți.
Caracteristicile structurii fără smocuri a tulpinilor de iarbă pot fi luate în considerare folosind exemplul unei tulpini in. Tulpina este acoperită cu epidermă, sub care există clorenchim liber al cortexului primar. Un număr mare de stomi în epidermă și absența colenchimului în scoarța primară au o importanță practică semnificativă, deoarece în timpul tăierii tehnice a inului, apa nu întâmpină obstacole și pătrunde ușor în tulpină. Celulele tecii de amidon (endoderm) sunt mai mari și conțin boabe de amidon clar vizibile.
Cilindrul central adiacent endodermului începe cu grupuri de celule sclerenchimatoase - fibre libiene, între care se află celulele parenchimului. Fibrele libiene apar în periciclu și în floemul primar. Se remarcă prin pereți foarte groși, de obicei nelignificați, formați din celuloză 75...90%. O celulă matură din fibre libiene este moartă. Protoplastul este distrus după îngroșarea peretelui celular. Fibrele de bast au o rezistență neobișnuit de mare. Rezistența lor la tracțiune corespunde cu cea a oțelului, iar elasticitatea lor este mai mare. Celulele din fibre de bast sunt foarte strâns legate între ele. Fibra de in este o materie primă excelentă de filare folosită în industria textilă.
În continuare, cilindrii (inele) sunt amplasați în ordine secvențială: floem secundar, cambium, xilem secundar cu raze medulare clar vizibile. Xilemul primar este situat în zonele adiacente mudofului. Parenchimul miezului este distrus rapid (în procesul de alungire a tulpinii), formând o cavitate centrală.
Tulpinile plantelor erbacee dicotiledonate se caracterizează prin:
1. structura secundara, care apare foarte devreme dupa cea primara;
2. prezența țesutului tegumentar primar - epiderma;
3. cortexul primar dezvoltat, împărțit în colenchim (exoderm), parenchim (mezoderm) și înveliș de amidon (endoderm);
4. structură cu grinzi sau fără grinzi (solide);
5. aranjarea corectă a fasciculelor vasculare colaterale sau bicolaterale într-un cerc de-a lungul periferiei cilindrului central;
6. prezența cambiumului (mănunchiuri deschise).
Metamorfozele de evadare.
Dacă funcțiile suplimentare încep să prevaleze asupra celor principale sau un organ dobândește unele funcții noi în procesul de dezvoltare, se schimbă foarte mult atât morfologic, cât și anatomic. Astfel de modificări ale organelor sunt numite metamorfoze(din grecescul metamorfoză - transformare, transformare). Organe care au aceeași origine, dar funcționează diferite funcții numit omolog . Organele omoloage sunt rizomii, tuberculii, bulbii, tepii de păducel, cladodele, filocladiile, cârcile de struguri și dovleci, deoarece toate s-au dezvoltat istoric din lăstari cu frunze.
Se numesc organe care îndeplinesc aceleași funcții și sunt similare morfologic, dar au origini diferite asemănătoare . Asemănătoare sunt conurile de rădăcină și tuberculii de lăstari, țepii de lăstari de păducel și țepii de frunze de arpaș, ramurile de frunze de mazăre și țepii de lăstari de dovleac.
Modificări ale lăstarilor subterani.
Lăstarii subterani sunt răspândiți în natură: rizomi, tuberculi, cormi, bulbi. Sunt implicați în aprovizionarea cu substanțe și în propagarea vegetativă a plantelor.
Rizom Este un lăstar cu frunze asemănătoare solzilor, muguri și rădăcini adventive. Cicatricile de la frunzele inferioare și lăstarii morți de anul trecut sunt clar vizibile pe suprafața rizomului. Rizomul are multe rădăcini adventive. De obicei, rizomii sunt sub pământ. Muguri care se dezvoltă în lăstari supraterane. Rizomii sunt caracteristici pentru iarba de grâu, iris și nuferi.
Stolon - un lăstar subțire alungit cu frunze subdezvoltate. Spre deosebire de rizom, este de obicei de scurtă durată. În iarba de mosc și măcrișul de lemn, stolonii participă la acumularea de nutrienți depuși în frunzele inferioare puternic îngroșate, dar funcția principală a stolonului este de a participa la propagarea vegetativă.
Tubercul spre deosebire de rizom și stolon, este foarte scurtat și îngroșat. Substanțele de rezervă din acesta sunt localizate în parenchimul tulpinii.
Tuberculii se pot dezvolta pe rizomi, stoloni, lăstari principali și alte părți ale plantelor. Ele pot fi subterane sau supraterane.
Tuberculii rizomatoși puternici cu o cantitate mare de amidon sunt caracteristici taro, sau colocasia comestibilă din familia aroid, care este cultivată în țările tropicale și subtropicale.
U cartofi tuberculii se formează la capătul stolonilor subterani - lăstari fragili albici, cu frunze asemănătoare solzilor, care se dezvoltă din muguri axilari aflați la baza lăstarilor cu frunze. Pe suprafața tuberculului există muguri axilari - ochi stând în gropi mici încadrate pe o parte sprancene - resturi de baze ale frunzelor.
Tuberculi topinambur, sau para de pământ, se formează și pe stoloni. Țesuturile conductoare din ele sunt slab dezvoltate. Tuberculii sunt bogați în inulină, care se depune în principal în parenchimul tulpinii.
Tuberculii formați pe stoloni și rizomi sunt de scurtă durată. Durează doar un an, între două sezoane de creștere, murind după ce mugurii lor se transformă în lăstari cu frunze care consumă substanțe nutritive până când se formează rădăcini și plantele devin autohrănite.
Bec, ca un tubercul, este un lăstar scurtat, specializat, modificat, care servește nu numai la stocarea nutrienților, ci și pentru a suporta perioade nefavorabile ale anului, reluarea vegetativă și reproducerea plantelor. Bulbii sunt caracteristici plantelor monocotiledonate din familiile Liliaceae, Amaryllis și alte familii. Prin excepție, ele se găsesc în dicotiledone: unele specii de măcriș și măcriș.
Tulpina numit fund, in bulb este mult redus si mai mult sau mai putin turtit. Poartă frunze sub formă de solzi, iar în partea inferioară sunt rădăcini adventive. Spre deosebire de rizom și tubercul, organele de depozitare ale bulbului sunt solzi cărnoase. Bulbii se formează în lalele, cocoși de alun, narcise, zambile, scilla, ghiocel și alte plante.
In functie de dezvoltarea mugurilor laterali, bulbii sunt simplu când mugurii sunt mici (ceapă, crini etc.) și complex– mugurii din axilele solzilor uscati sunt mari, foarte dezvoltati (usturoi).
Corm- o modificare subterană a lăstarilor, combinând caracteristicile unui rizom și ale unui bulb. Se dezvoltă dintr-un lăstar cu o tulpină foarte scurtată, din partea inferioară a cărei rădăcini adventive se extind. În vârful cormului sau la baza acestuia se află un mugure din care se formează un lăstar înflorit. Tulpina cormului este acoperită cu bazele frunzelor moarte de anul trecut, care arată ca niște solzi peliculoase uscate. Din mugurii localizați în sinusurile lor se dezvoltă noi cormi - copii. Cormii includ șofran, gladiole și colchicum.
Modificări ale lăstarilor supraterani
Kochan varza este în esență un mugur gigantic: partea de mijloc a capului de varză este ocupată de o tulpină cărnoasă ( poker), de-a lungul cărora sunt frunze suculente, cărnoase. Cea mai mică și cea mai tânără dintre ele se află în interiorul capului de varză, în jurul conului de creștere cu care se termină tulpina. Varza de Bruxelles formează mici capete de varză din mugurii laterali.
În capul de varză, ambele organe ale lăstarilor sunt implicate în acumularea de nutrienți: zaharuri solubile în apă și alți compuși valoroși din punct de vedere biologic, inclusiv vitamine, dar rolul principal revine frunzelor.
Tuberculi supraterani, care se dezvoltă din partea inferioară a lăstarului principal, sunt caracteristice varzei de guli-rabe.
În zona inflorescenței se pot dezvolta și noduli mici. La troscotul vivipari, de exemplu, ele se formează în axilele frunzelor inferioare de acoperire ale inflorescenței. Mugurii lor încep să înmugurească pe planta mamă. După cădere, ei prind rădăcini, formând noi plante.
Lăstarii metamorfozați pot servi nu numai pentru stocarea nutrienților, reproducerea și propagarea plantelor, ci și pentru alte funcții.
La unele plante, lăstarii pierd frunzele în timpul dezvoltării, iar tulpina devine un organ fotosintetic, numit în acest caz cladodiu . De obicei este turtită și are capacitatea de a crește timp îndelungat (în planta tropicală Mühlenbeckia, sau Homalocladus, din familia hrișcăi, pere). Nodurile au constricții bine definite, astfel încât cladodiul pare segmentat. În condiții umede, cladodele pot fi de foioase, dar pe vreme uscată frunzele cad. Ca organ al fotosintezei, cladodele au țesut bine dezvoltat, purtător de clorofilă, situat sub epidermă.
Cladodia, conform aspect asemănătoare cu frunzele se numesc filocladiile . Se dezvoltă din muguri laterali, așa că se găsesc întotdeauna în axila unei mici frunze peliculoase sau sub formă de solzi. Filocladiile sunt foarte diverse. La sparanghelul pinnat sau cu peri, sunt mici, în formă de ac, la sparanghelul lui Sprenger, sau cu flori dense, sunt turtite și liniare. În linii mari lanceolate sau ovoid, filocladiile piele sunt caracteristice speciilor de mături de măcelar găsite în Marea Mediterană, Transcaucazia și Crimeea. Vara, pe partea inferioară a filocladiului, în partea mijlocie, se dezvoltă o inflorescență, situată în axila frunzei mici de acoperire toamna și iarna, înflorește mătura de măcelar, iar primăvara se coc fructele de portocal; filocladia.
Un tip destul de comun de schimbare a lăstarilor este ghimpe . Acesta este un lăstar foarte lemnos, fără frunze, scurtat, cu un vârf ascuțit. La păducel, lăstarul care se dezvoltă la axila frunzei primăvara are inițial frunze mici și un mugure apical. Curând creșterea apicală se oprește, frunzele cad, tulpina devine foarte lignificată și se transformă într-un ghimpe. Spinii se dezvoltă în același mod.
Sunt echipate plante agatatoare antene , care, ca şi spinii, sunt lăstari laterali modificaţi la unele plante. Pasiflora are virici simple, neramificați, cu vârful ondulat. Se găsesc în axilele frunzelor, la fel ca și florile. Vricile ramificate de struguri sunt inflorescențe modificate. La strugurii de fetiță, capetele vârcilor ramificați au formă de disc și joacă rolul de ventuze, ajutând planta să urce pe un suport. În dovleac, pepene verde, castraveți și alți membri ai familiei dovleac se formează șiruri de origine lăstară.