Книгата очертава текущ проблем съвременна естествена наука- произход на живота. Написана е въз основа на най-съвременни данни от геологията, палеонтологията, геохимията и космохимията, които опровергават много традиционни, но остарели представи за произхода и развитието на живота на нашата планета. Античностживота и биосферата, съизмерими с възрастта на самата планета, позволява на автора да заключи: възникването на Земята и живота е единен взаимосвързан процес.
За читатели, интересуващи се от геонауки.
книга:
| <<< Назад
|
Напред >>> |
Растенията, като типични представители на фотоавтотрофните организми на нашата планета, са възникнали по време на дълга еволюция, която произхожда от примитивните обитатели на осветената зона на морето - планктонни и бентосни прокариоти. Чрез сравняване на палеонтологични данни с данни за сравнителната морфология и физиология на живите растения е възможно общ изгледочертават следната хронологична последователност на тяхната поява и развитие:
1) бактерии и синьо-зелени водорасли (прокариоти);
2) циан, зелени, кафяви, червени и др. водорасли (еукариоти, както всички следващи организми);
3) мъхове и чернодробни червеи;
4) папрати, хвощове, мъхове, семенни папрати;
6) покритосеменни или цъфтящи растения.
Бактерии и синьо-зелени водорасли са открити в най-древните запазени отлагания от докамбрия; водораслите се появяват много по-късно и едва през фанерозоя се срещаме с буйно развитие на висши растения: ликофити, хвощове, голосеменни и покритосеменни.
През целия криптозойски период в първичните резервоари в еуфотичната зона на древните морета се развиват предимно едноклетъчни организми - различни видове водорасли.
Основните представители на прокариотите, открити в докамбрия, са имали автотрофно хранене - чрез фотосинтеза. Повечето благоприятни условияза фотосинтеза са създадени в осветената част на морето на дълбочина от повърхността до 10 m, което също съответства на условията на плитководния бентос.
Към днешна дата изучаването на докамбрийските микрофосили е напреднало и съответно е натрупан голям фактически материал. Като цяло интерпретацията на микроскопични проби е трудна задача, която не може да бъде решена еднозначно.
Най-добре се идентифицират и идентифицират трихомните бактерии, които се различават рязко от минерални образувания с подобна форма. Полученият емпиричен материал върху микрофосили ни позволява да заключим, че те могат да бъдат сравнени с живите цианобактерии.
Строматолитите, като биогенни структури от далечното минало на планетата, са се образували по време на натрупването на тънък седимент от калциев карбонат, уловен от фотосинтезиращи организми на микробиологични асоциации. Микрофосилите в строматолитите се състоят почти изключително от прокариотни микроорганизми, свързани главно със синьо-зелените водорасли - цианофити. При изучаването на останките от бентосни микроорганизми, съставляващи строматопитите, беше разкрита една интересна особеност от фундаментално значение. Микрофосилите от различни възрасти се променят малко в морфологията си и като цяло показват консерватизма на прокариотите. Микрофосилите, свързани с прокариотите, остават почти постоянни за доста време. за дълго време. Във всеки случай, пред нас е установен факт - еволюцията на прокариотите е била много по-бавна от тази на висшите организми.
И така, по време на геоложка историяПрокариотните бактерии проявяват максимална устойчивост. Устойчивите форми включват организми, които са се запазили непроменени в процеса на еволюцията. Както отбелязва G. A. Zavarzin, тъй като древните съобщества от микроорганизми показват значителни прилики със съвременните, развиващи се в хидротерми и в области на образуване на изпарители, това прави възможно по-задълбочено изследване на геохимичната активност на тези съобщества, като се използват съвременни природни и лабораторни модели, екстраполирайки ги към далечното докамбрийско време.
Първите еукариоти възникват в планктонни асоциации открити води. Краят на изключителното господство на прокариотите датира отпреди приблизително 1,4 милиарда години, въпреки че първите еукариоти са се появили много по-рано. Така, според най-новите данни, появата на изкопаеми органични останки от черни шисти и въглеродни образувания в района на Горното езеро показва появата на еукариотни микроорганизми преди 1,9 милиарда години.
От датата преди 1,4 милиарда години до наше време докамбрийските вкаменелости се разширяват значително. Към тази дата датира появата на сравнително големи форми, свързани с планктонните еукариоти и наречени „акритархи“ (в превод от гръцки „същества с неизвестен произход“). Трябва да се отбележи, че групата Acritarcha е предложена като неясна систематична категория, обозначаваща микрофосили от различен произход, но подобни на външен вид морфологични характеристики. В литературата са описани акритархи от докамбрия и долния палеозой. Повечето акритархи вероятно са били едноклетъчни фотосинтетични еукариоти - черупки на някои древни водорасли. Някои от тях все още могат да имат прокариотна организация. Планктонната природа на акритархите е показана от тяхното космополитно разпространение в седименти от същата възраст. Най-древните акритархи от раннорифейските отлагания на Южен Урал са открити от Т. В. Янкаускас.
С течение на геоложкото време размерът на акритархите се е увеличил. Според данните от наблюдения се оказа, че колкото по-млади са докамбрийските микрофосили, толкова по-големи са те. Предполага се, че значително увеличаване на размера на акритархите е свързано с увеличаване на размера на еукариотната клетъчна организация. Те биха могли да се появят като независими организми или, по-вероятно, в симбиоза с други. Л. Маргелис смята, че еукариотни клеткисъбрани от вече съществуващи прокариоти. Въпреки това, за оцеляването на еукариотите е необходимо местообитанието да бъде наситено с кислород и в резултат на това възниква аеробен метаболизъм. Първоначално свободният кислород, освободен по време на фотосинтезата на цианофитите, се натрупва в ограничени количества в плитки водни местообитания. Увеличаването на съдържанието му в биосферата предизвика реакция от страна на организмите: те започнаха да населяват безкислородни местообитания (по-специално анаеробни форми).
Данните от докамбрийската микропалеонтология показват, че в средния докамбрий, дори преди появата на еукариотите, цианофитите представляват относително малка част от планктона. Еукариотите се нуждаеха от свободен кислород и все повече се конкурираха с прокариотите в онези области на биосферата, където се появи свободен кислород. Въз основа на наличните микропалеонтологични данни може да се прецени, че преходът от прокариотна към еукариотна флора на древните морета е настъпил бавно и двете групи организми са съществували заедно дълго време. Това съжителство обаче се среща в различна пропорция в съвременната епоха. До началото на късния рифей вече са се разпространили много автотрофни и хетеротрофни форми на организми.
Докато се развиват, организмите се преместват за хранителни вещества в по-дълбоки и по-отдалечени райони на морето. Вкаменелостите отбелязват рязко увеличение на разнообразието от големи сфероидни форми на еукариотни акритархи в късните рифейски времена, преди 900-700 милиона години. Преди около 800 милиона години в Световния океан се появяват представители на нов клас планктонни организми - тела с форма на чаша с масивни черупки или външни обвивки, минерализирани с калциев карбонат или силициев диоксид. В началото на камбрийския период настъпват значителни промени в еволюцията на планктона - възникват различни микроорганизми със сложна изваяна повърхност и подобрена плаваемост. Те породиха истински бодливи акритархи.
Появата на еукариотите създава важна предпоставка за появата на многоклетъчни растения и животни в ранния Рифей (преди около 1,3 милиарда години). За серията Белта от докамбрия на западните щати Северна Америкате са описани от C. Walcott, но към какъв тип водорасли принадлежат (кафяви, зелени или червени) все още не е ясно. Така изключително дългата ера на господство на бактериите и свързаните с тях синьо-зелени водорасли беше заменена от ера на водорасли, които достигнаха значително разнообразие от форми и цветове във водите на древните океани. През късния рифей и венд многоклетъчните водорасли стават по-разнообразни с кафявите и червените водорасли.
Според академик Б.С.Соколов многоклетъчните растения и животни се появяват почти едновременно. Във вендските седименти се срещат различни представители на водните растения. Най-видно място заемат многоклетъчните водорасли, талиите на които често преливат слоевете на вендските седименти: калници, глини, пясъчници. Често се срещат макропланктонни водорасли, колониални водорасли, спираловидни нишковидни водорасли Volymella, филцови водорасли и други форми. Фитопланктонът е много разнообразен.
През по-голямата част от историята на Земята еволюцията на растенията е протичала във водна среда. Тук водната растителност се заражда и преминава през различни етапи на развитие. Като цяло водораслите са голяма група нисши водни растения, които съдържат хлорофил и произвеждат органична материя чрез фотосинтеза. Тялото на водораслите все още не е диференцирано на корени, листа и други характерни части. Те са представени от едноклетъчни, многоклетъчни и колониални форми. Размножаването е безполово, вегетативно и полово. Водораслите са част от планктона и бентоса. Понастоящем те се класифицират като растително подцарство Thallophyta, в което тялото е съставено от относително еднаква тъкан - талус или талус. Талусът се състои от много клетки, които са сходни по външен вид и функция. В исторически план водораслите преминават през най-дългия етап от развитието на зелените растения и в общия геохимичен кръговрат на материята в биосферата играят ролята на гигантски генератор на свободен кислород. Появата и развитието на водораслите беше изключително неравномерно.
Зелените водорасли (Chlorophyta) са голяма и широко разпространена група от предимно зелени растения, която попада в пет класа. от външен видте са много различни един от друг. Зелените водорасли идват от зелени камшичести организми. Това се доказва от преходни форми - пирамидомони и хламидомони, подвижни едноклетъчни организми, които живеят във водите. Зелените водорасли се размножават по полов път. Някои групи зелени водорасли са постигнали голямо развитие през триаския период.
Камшичестите (Flagellata) са групирани в група микроскопични едноклетъчни организми. Някои изследователи ги приписват на растителното царство, други на животинското царство. Подобно на растенията, някои флагелати съдържат хлорофил. Въпреки това, за разлика от повечето растения, те нямат отделна клетъчна система и могат да усвояват храната с помощта на ензими, а също така живеят на тъмно, като животински организми. По всяка вероятност флагелатите са съществували в докамбрия, но техните безспорни представители са открити в юрските отлагания.
Кафявите водорасли (Phaeophyta) се отличават с наличието на кафяв пигмент в такива количества, че всъщност маскира хлорофила и придава на растенията подходящ цвят. Кафявите водорасли принадлежат към бентоса и планктона. Най-големите водорасли достигат до 30 m дължина. Почти всички те растат в солена вода, поради което се наричат морска трева. Кафявите водорасли включват саргасови водорасли - плаващи планктонни форми с голям брой мехурчета. Във фосилна форма те са известни от силура.
Червени водорасли(Rhodophyta) имат този цвят поради червения пигмент. Това са предимно морски растения, силно разклонени. Някои от тях имат варовит скелет. Тази група често се нарича cullipora. Те съществуват днес и са известни във вкаменелости от долната креда. Близките до тях сомипори с по-големи и широки клетки се появяват през ордовика.
Чаровая водорасли(Charophyta) са много уникална и доста добре организирана група от многоклетъчни растения, които се размножават полово. Те са толкова различни от другите водорасли, че някои ботаници ги класифицират като листно-стъблени водорасли поради появяващата се тъканна диференциация. Водораслите Charovaya са зелени на цвят и в момента живеят в сладки и солени води. Те избягват морска вода с нормална соленост, но може да се предположи, че през палеозоя са обитавали моретата. Някои харофити развиват спори, импрегнирани с калциев карбонат. Characeae са сред важните скалообразуващи организми на сладководни варовици.
Диатомови водорасли(Diatomeae) - типични представители на планктона. Имат продълговата форма и са покрити отвън с обвивка от силициев диоксид. Първите останки от диатомеи са открити в девонските седименти, но те може да са по-стари. Като цяло диатомените водорасли са сравнително млада група. Тяхната еволюция е проучена по-добре от другите водорасли, тъй като кремъчните черупки и клапите на диатомеите могат да се запазят във фосилно състояние за много дълго време. По всяка вероятност диатомените водорасли произлизат от флагелатите, които са жълти на цвят и са способни да отлагат малки количества силициев диоксид в черупките си. В съвремието диатомеите са широко разпространени в пресни и морски води, понякога се срещат във влажни почви. Останки от диатомеи са известни в юрските отлагания, но е възможно да са се появили много по-рано. Изкопаемите диатомеи от ранната креда са достигнали съвременната ера без прекъсване на седиментацията.
Много важно събитие, което допринесе за рязкото ускоряване на скоростта на еволюция на цялото живо население на нашата планета, беше появата на растения от морската среда на сушата. Появата на растения на повърхността на континентите може да се счита за истинска революция в историята на биосферата. Развитието на сухоземната растителност създаде предпоставки животните да достигнат сушата. Масовият преход на растенията към сушата обаче беше предшестван от дълъг подготвителен период. Може да се предположи, че растителният живот на сушата се е появил много отдавна, поне локално - във влажен климат по бреговете на плитки заливи и лагуни, където промените в нивото на водата периодично довеждат водната растителност на сушата. Съветският натуралист Л. С. Берг пръв изрази идеята, че земната повърхност не е безжизнена пустиня нито в камбрия, нито в докамбрия. Изтъкнатият съветски палеонтолог Л. Ш. Давиташвили също допуска, че в докамбрия континентите вероятно вече са имали някакво население, състоящо се от нискоорганизирани растения и, вероятно, дори животни. Общата им биомаса обаче е незначителна.
За да живеят на сушата, растенията не трябваше да губят вода. Трябва да се има предвид, че при висшите растения - мъхове, птеридофити, голосеменни и цъфтящи растения, които в момента съставляват основната част от сухоземната растителност, само корените, кореновите власинки и ризоидите влизат в контакт с водата, докато останалите им органи са в атмосферата и изпарява водата по цялата повърхност.
Растителният живот процъфтява най-много по бреговете на лагуни езера и блата. Тук се появи вид растение, чиято долна част беше във вода, а горната част във водата въздушна среда, на пряка слънчева светлина. Малко по-късно, с навлизането на растенията в ненаводнена земя, се развиват първите им представители коренова системаи са били в състояние да консумират подземни води. Това допринесе за оцеляването им по време на сухи периоди. По този начин нови обстоятелства доведоха до разделянето на растителните клетки на тъкани и развитието на защитни устройства, които не съществуваха при предците, които живееха във вода.
Фиг. 14. Развитие и генетични връзки различни групиземни растения
Масовото завладяване на континентите от растения е настъпило през силурийския период на палеозойската ера. На първо място, това бяха псилофити - особени спорови растения, наподобяващи клубни мъхове. Някои от усукващите се стъбла на псилофитите бяха покрити с настръхнали листа. Псилофитите бяха лишени от корени и предимно листа. Те се състоят от разклонени зелени стъбла с височина до 23 см и коренища, простиращи се хоризонтално в почвата. Псилофитите, като първите надеждни растения за суши, създават цели зелени килими върху влажна почва.
Вероятно производството на органична материя от първата земна растителност е било незначително. Растителността на силурийския период несъмнено произхожда от водораслите на морето и сама по себе си е дала началото на растителността на следващия период.
След завладяването на земята развитието на растителността доведе до образуването на многобройни и разнообразни форми. Интензивното отделяне на растителни групи започва през девона и продължава в последващото геоложко време. Общото родословие на най-важните растителни групи е дадено на фиг. 14.
Мъховете произлизат от. морски водорасли Техният ранен етап на развитие е много подобен на някои зелени водорасли. Съществува обаче предположение, че мъховете са произлезли от по-прости представители на кафяви водорасли, адаптирани към живот на влажни скали или в почви като цяло.
На повърхността на ранните палеозойски континенти епохата на водораслите отстъпи място на епохата на псилофитите, което доведе до растителност, която напомняше по външен вид и размер на съвременни гъсталаци от големи мъхове. Доминирането на псилофитите беше заменено през карбоновия период от господството на папратовидни растения, които образуваха доста обширни гори върху блатисти почви. Развитието на тези растения допринесе за това, че съставът атмосферен въздухпроменен. Добавено е значително количество свободен кислород и е натрупана маса от хранителни вещества, необходими за появата и развитието на сухоземните гръбначни животни. В същото време бяха натрупани огромни маси въглища. Карбонният период се характеризира с изключителен разцвет на сухоземната растителност. Появиха се дървесни мъхове, достигащи до 30 м височина, започнаха да се появяват огромни хвощове, папрати, иглолистни дървета. През пермския период продължава развитието на сухоземната растителност, което значително разширява местообитанията му.
Периодът на господство на папратите отстъпи място на периода на конусовидни иглолистни дървета. Повърхността на континентите започна да придобива модерен вид. В началото Мезозойска ераШироко разпространение получават иглолистните дървета и цикадите, които се появяват през периода Креда цъфтящи растения. В самото начало на ранната креда все още съществуват юрски форми на растения, но след това съставът на растителността се промени значително. В края на епохата ранна креда се срещат много покритосеменни растения. От самото начало на късната креда те изтласкаха голосеменните растения и заеха господстващо положение на сушата. Като цяло в сухоземната флора се наблюдава постепенна замяна на мезозойската растителност от голосеменни (иглолистни, цикасови, гинго) с растителност от кайнозойски вид. Растителността от късната креда вече се характеризира с наличието на значителен брой съвременни цъфтящи растения като бук, върба, бреза, чинар, лавровишня и магнолия. Това преструктуриране на растителността подготви добра хранителна база за развитието на висшите сухоземни гръбначни животни - бозайници и птици. Развитието на цъфтящите растения е свързано с разцвета на множество насекоми, които играят важна роля в опрашването.
Началото на нов период в развитието на растенията не доведе до пълното унищожаване на древните растителни форми. Някои организми от биосферата бяха запазени. С появата на цъфтящи растения бактериите не само не изчезнаха, но продължиха да съществуват, намирайки нови източници на хранене в почвата и в органичната материя на растенията и животните. водорасли различни груписе променя и развива заедно с висшите растения.
Иглолистните гори, появили се през мезозоя, растат и днес заедно с широколистните. Те осигуряват подслон на папратовидни растения, тъй като тези древни обитатели на мъгливия и влажен климат на периода на карбон се страхуват от открити места, осветени от слънцето.
И накрая, трябва да се отбележи, че в съвременната флора има устойчиви форми. Най-устойчиви са определени групи бактерии, които остават практически непроменени от ранния докамбрий. Но от по-високо организирани форми на растения са се образували и родове и видове, които са се променили малко до днес.

Трябва да се отбележи, че в съвременната флора има несъмнено присъствие на относително високо организирани многоклетъчни растителни родове. Късните палеозойски и мезозойски форми на растения, които са живели без промени в продължение на десетки и стотици милиони години, разбира се, са устойчиви. Така понастоящем сред растителния свят са запазени „живи вкаменелости” (фиг. 15) от групи папрати, голосеменни и клубни мъхове. Терминът „жива вкаменелост“ е използван за първи път от Чарлз Дарвин, цитирайки източноазиатското голосеменно дърво Ginkgo biloba като пример. От света на земните растения живите вкаменелости включват най-известните папратови палми, дърво гинко, араукария, мамутово дърво или секвоя.
Както отбелязва експертът по изкопаеми флори А. Н. Кршптофович, много родове растения, господари на древни гори, също са съществували изключително дълго време, особено в палеозоя; например Сигилария, Лепидодендрон, Каламити - поне 100-130 милиона години. Същият брой - мезозойски папрати 11 иглолистни Metasequoia. Родът Ginkgo датира отпреди повече от 150 милиона години и модерен видГинко билоба, ако включите по същество неразличимата форма Ginkgo adiantoides, е на около 100 милиона години.
Живите вкаменелости от съвременния растителен свят иначе могат да се нарекат филогенетично запазени типове. Растенията, които са добре проучени от палеоботаническа гледна точка и класифицирани като живи вкаменелости, са консервативни групи. Те не са се променили изобщо или са се променили много малко в сравнение със сродните форми на геоложкото минало.
Естествено, наличието на живи вкаменелости в съвременната флора повдига проблема за тяхното формиране в историята на биосферата. Консервативните организации присъстват във всички основни филогенетични клонове и съществуват в повечето различни условия: в дълбоки и плитки морски зони, в древни тропически гори, в открити степни пространства и във всички водни тела без изключение. Най-важното условиеза съществуването на еволюционно консервативни организми - наличие на местообитания с постоянна жизнена среда. Стабилните условия на живот обаче не са решаващи. Само присъствие отделни форми, а не всички съобщества от флора и фауна, показва други фактори в запазването на живите вкаменелости. Проучването на географското им разпространение показва, че те са ограничени до строго определени територии, докато географска изолация. Така Австралия, островите Мадагаскар и Нова Зеландия са типични райони на разпространение на земни живи вкаменелости.
В своята еволюция растителният свят създава общия облик на древните ландшафти, в които е протичало развитието на животинския свят. Следователно разделянето на геоложкото време може да се извърши въз основа на последователността на различни растителни форми. Немският палеоботаник В. Цимерман още през 1930 г. разделя цялото геоложко минало от гледна точка на развитието на растителния свят на шест епохи. Той им даде буквено обозначение и ги подреди в последователност от древни епохи към по-млади.
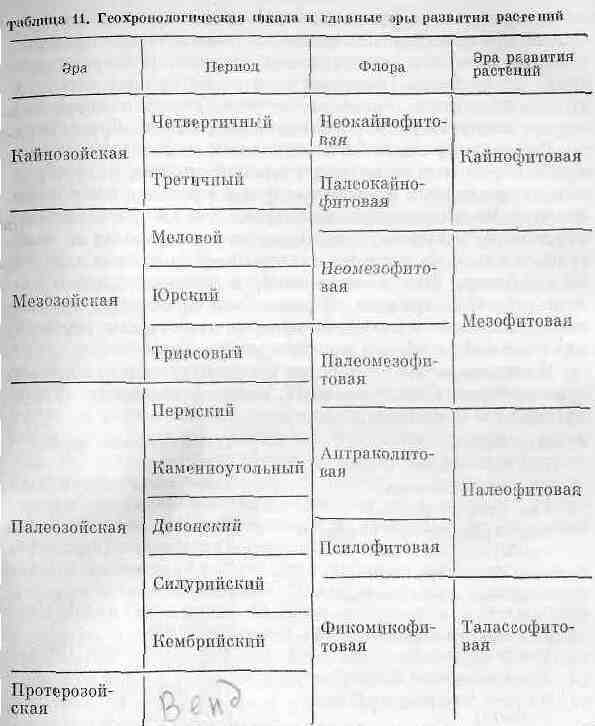
Сравнение на обичайната геоложка времева скала, изградена предимно от палеозоложки данни, със скалата на развитие на растенията е представена в табл. 11.
| <<< Назад
|
Напред >>> |
Първите растителни организми са възникнали във волята в много далечни времена. Първите живи същества са били микроскопично малки бучки слуз. Много по-късно някои от тях придобиват зелен цвят и тези живи организми започват да приличат на едноклетъчни водорасли. Едноклетъчните създания породиха многоклетъчни организми, които подобно на едноклетъчните организми са възникнали във водата. От едноклетъчните водорасли са се развили различни многоклетъчни водорасли.
Повърхността на континентите и океанското дъно са се променили с времето. Новите континенти се издигнаха и съществуващите преди това потънаха. Поради колебание земна кораНа мястото на моретата възникнала земя. Изследването на фосилни останки показва, че растителният свят на Земята също постепенно се е променил.
Преходът на растенията към сухоземен начин на живот, според учените, е свързан със съществуването на земни площи, които периодично се наводняват и почистват от вода. Отдръпналата се вода се задържа във вдлъбнатините. Те или изсъхнаха, или се напълниха отново с вода. Отводняването на тези площи става постепенно. Някои водорасли са развили адаптации за живот извън водата.
Климатът по това време на земното кълбо е влажен и топъл. Започва преходът на някои растения от воден към сухоземен начин на живот. Структурата на тези растения постепенно се усложнява. Те са дали началото на първите сухоземни растения. Най-старата група известни сухоземни растения са псилофитите.
Развитието на растителния свят на Земята е дългосрочен процес, който се основава на прехода на растенията от воден към сухоземен начин на живот.
Псилофитите вече са съществували преди 420-400 милиона години, а по-късно са изчезнали. Псилофитите растяха по бреговете на резервоари и бяха малки многоклетъчни зелени растения. Те нямаха корени, стъбла или листа. Ролята на корените се играе от ризоиди. Псилофитите, за разлика от водораслите, имат по-сложен вътрешна структура- наличие на покривни и проводими тъкани. Те се възпроизвеждат чрез спори.
От псилофитите произлезли бриофитите и папратите, които вече имали стъбла, листа и корени. Разцветът на папратите е преди около 300 милиона години по време на карбонския период. Климатът по това време беше топъл и влажен. В края на карбоновия период климатът на Земята стана забележимо по-сух и по-студен. Дървесните папрати, хвощовете и клубните мъхове започнаха да измират, но по това време се появиха примитивни голосеменни растения - потомци на някои древни папрати. Според учените първите голосеменни са били семенни папрати, които по-късно са напълно изчезнали. Техните семена се развиха върху листата: тези растения нямаха шишарки. Семенните папрати са били дървовидни, лианоподобни и тревисти растения. От тях са произлезли голосеменните.
Условията на живот продължиха да се променят. Там, където климатът беше по-тежък, древните голосеменни растения постепенно измряха и бяха заменени от по-напреднали растения - древни иглолистни дървета, след което бяха заменени от съвременни иглолистни дървета: бор, смърч, лиственица и др.
Преходът на растенията към сушата е тясно свързан не само с появата на органи като стъбла, листа и корени, но главно с появата на семена, специален метод за размножаване на тези растения. Растенията, които се размножават чрез семена, са по-добре адаптирани към живота на сушата, отколкото растенията, които се размножават чрез спори. Това стана особено ясно, когато климатът стана по-малко влажен.
На израстъците, развиващи се от спори (в мъхове, мъхове, папрати), се образуват женски и мъжки гамети (полови клетки) - яйца и сперматозоиди. За да се осъществи оплождането (след сливането на гамети), са необходими атмосферни или подпочвени води, в които сперматозоидите се придвижват към яйцата.
Голосеменните не се нуждаят от безплатна вода за оплождане, тъй като това се случва вътре в яйцеклетките. При тях мъжките гамети (сперма) се приближават до женските гамети (яйцата) чрез поленови тръбички, растящи вътре в яйцеклетките. По този начин оплождането при споровите растения зависи напълно от наличието на вода; при растенията, които се размножават чрез семена, тази зависимост не е налице.
Покритосеменните - потомци на древни голосеменни - се появяват на Земята преди повече от 130-120 милиона години. Те се оказаха най-адаптирани за живот на сушата, тъй като само те имат специални репродуктивни органи - цветя, а семената им се развиват вътре в плода и са добре защитени от перикарпа.
Благодарение на това покритосеменните растения бързо се разпространяват по цялата земя и заемат голямо разнообразие от местообитания. Повече от 60 милиона години покритосеменните са доминирали на Земята. На фиг. 67 показва не само последователността на появата на определени растителни отдели, но и техния количествен състав, където покритосеменните имат значително място.
Въздушно-земното местообитание се различава от водното. Плътността на водата е по-голяма от плътността на въздуха. Следователно водата изглежда поддържа телата в дебелината си. На сушата въздухът не може да „задържи“ растенията и гравитацията би ги накарала да паднат, ако сухоземните растения нямаха здрави механични тъкани. Няма недостиг на вода във водната среда; растението може да я абсорбира в цялото тяло. Във въздуха има много малко вода и тя е там под формата на пара. Растението вече не може да го абсорбира с цялото си тяло. Следователно беше необходимо да се развие коренова система, която абсорбира вода от почвата. За да се достави вода до други органи, е необходима проводна система.
Така появата на растенията на сушата постепенно доведе до образуването на различни тъкани и органи. Имаше специализация на частите на тялото. Корените на растението осъществяват храненето на почвата чрез абсорбиране на вода с разтворени в нея минерали. Листата извършват въздушно хранене, синтезирайки органични вещества на светлина. Стъблото осигурява движението на вода и вещества.
За дълго време земни растенияимаше само спорови растения - мъхове, папрати, хвощ и мъхове. Те се нуждаят от вода, за да се размножават, така че не могат да живеят навсякъде на сушата.
Преди около 250 милиона години климатът на Земята е станал по-студен и сух. Това доведе до появата на голосеменни растения, които можеха да понасят такива условия. Голосеменните са развили пъпки, които са били защитени от неблагоприятни условия с пъпкови люспи. Възпроизвеждането чрез семена се появява, когато водата вече не е необходима по време на оплождането. Сперматозоидите се доставят до яйцеклетката с помощта на поленова тръба. Също така семената имат доставка хранителни вещества. Голосеменните успяха да колонизират сухи и студени местообитания и там, където живееха спорови растения, те постепенно ги замениха.
Покритосеменните се появяват преди около 150 милиона години. По това време климатът на Земята е станал още по-сух и топъл. Покритосеменните са по-приспособени за живот в сложни и разнообразни условия поради по-сложната си структура. Растителните тъкани се подобриха и се появи разлистване. образуване на цветя, различни начиниопрашването, двойното оплождане и образуването на плодове дадоха предимства на цъфтящите растения. Разпространението на вегетативното размножаване сред цъфтящите растения и различните му методи направиха възможно бързото заемане на пространство.
Цъфтящите растения и техните животински опрашители (предимно насекоми) са тясно свързани. Тяхната еволюция протичаше заедно, взаимно обусловена. Растенията и насекомите са се приспособили към жизнени циклиедин друг.
Много растения, съществували в по-ранни времена, не са оцелели до днес. Повечето от тях са изчезнали, защото са се оказали неадаптирани към променените условия на живот или са били заменени от по-адаптирани видове. Някои растения са унищожени от хората. Изчезналите растения се изучават от палеоботаниката. Чрез изучаване на растителни вкаменелости учените могат да ги проследят еволюционен пътвъв времето.
Преди около милиард години много зелени, кафяви и други водорасли вече са живели на дъното на Световния океан. Прогресивната им еволюция доведе до факта, че някои от тях достигнаха гигантски размери, но останаха водни създания. Това се обяснява не само с липсата на специална защита от изсушаване в телата им, но и с особеностите на сексуалното размножаване. Както е известно, във водораслите има редуване на поколения: диплоиден спорофит, който се възпроизвежда чрез спори, и хаплоиден гаметофит, на етапа на който възпроизвеждането става с помощта на гамети, копулацията на които изисква водна среда. Именно това се превърна в основната пречка водораслите да достигнат сушата.
Смята се, че необходимостта от адаптиране към сухоземен начин на живот се е превърнала в основен фокус еволюция на растенията. Това се постига чрез увеличаване на продължителността на спорофитния стадий и постепенно намаляване на гаметофитната фаза и с течение на времето нейното намаляване като цяло. Именно ароморфозата, свързана с намаляването на гаметофита в семенните растения, им позволи да развият ново адаптивно пространство - земя и да създадат тук разнообразие от видове. ФлораСега има около половин милион вида, от които цъфтящите растения представляват повече от половината - около 300 хиляди вида.
Бриофитите се считат за най-примитивните висши растения, запазени на Земята. Те вече имат някакво разделение на тялото на стъбло и ризоиди, въпреки че все още нямат проводяща система. Мъховете са се развили от водорасли в самото начало на фанерозоя преди около 600 милиона години. От спорите развиват т.нар предварително бягство,много подобен на водораслите, а от него - тялото на мъха - гаметофита. Копулацията на гамети става само във вода, която те натрупват в пазвите на листата. Следователно мъховете могат да живеят само на влажни, сенчести места. От зиготата се образува спорофит, който се развива директно върху гаметофита (фиг. 223). За разлика от гаметофита, спорофитът е устойчив на суша, което позволява на мъховете да достигнат сушата. Следователно те не са истински сухоземни растения, а земноводни. Може би затова те се оказаха сляпо разклонение в еволюцията на растенията.
Всички останали висши растенияпроизлиза от риниофити(Фиг. 224, 225), което е дало началото на зелени или кафяви водорасли в силурийския период преди 400-500 милиона години. Мопсите, хвощовете, папратите и голосеменните се отделиха от тях като отделни еволюционни клонове. Тази еволюционна експлозия се е случила в девонския период преди около 300-400 милиона години. Папратите станаха първите растения, които наистина завладяха земята, образувайки истински гори през карбоновия период.
Поради необходимостта да живеят на сушата, растенията развиват проводяща система, подобряват се покривните и механичните тъкани. От голосеменни на границата на мезозоя и кайнозоя са се образували покритосеменни растения. Появата на покритосеменните се характеризира с редица ароморфози: появата на цвете, образуването на вътрешно и двойно оплождане, защита на ембриона от неблагоприятни условия и осигуряването му на храна в ранните етапи на развитие. Материал от сайта
По време на еволюцията на покритосеменните цветето е претърпяло най-големи промени. Това до голяма степен се дължи на адаптацията на растението към опрашване от вятър или насекоми. В последния случай цветята обикновено са големи, ярки, с изобилен прашец и ароматен нектар и много често имат специализиран опрашител ( запомнете: цветята на детелината могат да се опрашват само от земни пчели).Тази специализация от своя страна стимулира еволюцията на насекомите. Неслучайно началото на кайнозоя е белязано от избухване на разнообразие в цъфтящи растения и насекоми. Този процес на взаимна еволюция се нарича коеволюция(от лат. ко- заедно).
Основен начин еволюция на растениятаимаше намаляване на значението и продължителността на гаметофита, което допринесе за появата на растения на сушата и последващото образуване на цъфтящи растения.
На тази страница има материали по следните теми:
Модели на доклад за биологичната еволюция
Доклад по темата за еволюцията на висшите растения
Доклад по темата за еволюцията на растенията
Коеволюция на висшите растения