Стъблото. Морфология и функция на ствола.Стъблото е продълговата издънка на висши растения, служеща като механична ос, изпълнява и функцията на проводяща и поддържаща основа за листа, пъпки, цветя.Стъблата изпълняват две основни функции - носеща и водеща. Стъблата свързват всички части на издънката в една система. Благодарение на стъблото се осигурява увеличаване на повърхността на растението чрез неговото разклоняване, формиране и изгодно разположение на листата и генеративните органи. Стъблото осигурява движение нагоре и надолу на вода и вещества, разтворени в него. Младите издънки изпълняват функцията на фотосинтеза. Модифицираните стъбла изпълняват и допълнителни функции, например натрупване и съхраняване на хранителни вещества в многогодишни стъбла, както и съхраняване на вода в специализирани стъбла, закачане на други растения, защита и осигуряване на вегетативно размножаване на растенията.
Стъблата на растенията са много разнообразни и се различават по много начини. По консистенцията на стъблата са: тревисти или лигави, кухи (слама) или пълни (с сърцевина). В напречно сечение стъблата са кръгли или цилиндрични (при повечето растителни видове), тетраедрични (видове глухи, норничникови), триградни (видове осока) ), многопластови (видове киноа, киселец), сплескани (например, пресовани от синя трева, сегментирани носорози). По характера на растеж и разположение в пространството се разграничават следните видове стъбла: изправени (силни, най-вече ортотропни стъбла с добре изразени междувъзли, най-високи видове растения) пълзящи (главно плагиотропни стъбла, корен в възлите, с добре изразени междувъзлия; например, пълзящи живи ягоди и др. Пепеляшка пълзяща, сребриста гъска) къдрава (усукване около опората по посока на часовниковата стрелка или обратното; например, обикновен хмел, полеви връхчета, дондър, обикновен боб) катерене или шипове (стъблата с слабо развита механична тъкан са закачени с антени; например краставици, тиква, грах); пълзящи (стъблата с доста слабо развита механична тъкан, пълзящи над почвената повърхност: например, рогата е гола, койотът лежи, пълзящото стъбло, стъблото ) възходящо (долната част на стъблото се разпространява по почвената повърхност, горната е насочена вертикално; например мащерката, звездното кълнове, лекарството вероника) се скъсяват (стъблото е слабо изразено, с много близки възли, къси междувъзлия и Що се отнася до розетка листа, цветни форми на стрелки; например, трилистник, глухарче, лилии, кокиче). Изменения на стъблото.
Модифицираното стъбло е тяло, предназначено да акумулира резервните хранителни вещества, от които растението се нуждае, за да оцелее в периода на сън. В допълнение, модифицираните стъбла често участват в размножаването и разпространението на растенията. За да бъде причислен към тази група, модифицираното стъбло трябва да има всички признаци на нормално стъбло, тоест трябва да има характерна структура, което означава, че самото стъбло има горна точка на растеж и оставя спираловидно (редуващо или противоположно) и носещи пъпки в своите оси. , Разликата между модифицирано стъбло и нормално е, че не е задължително да расте над земята, растежът му често е повече или по-малко хоризонтален и обикновено е контейнер с резервни хранителни вещества.
Има шест основни вида модифицирани стъбла: коренището на грудката на кората на луковицата върху мустакалната мухика
Появата на първични тъкани и първичната анатомична структура.Анатомичната структура на стъблото на растенията поради основните му функции. Характеризира се с развитието на механична и проводима тъкан на растенията. Освен това стъблото се характеризира със сложна система от меристеми - апикална, латерална и интеркаларна, които определят растежа му за дълго време и появата на нови органи.
Стъблото на растението възниква от апикалната меристема, от която се разграничават три слоя тъкани: покривна, проводима и основна. Отначало тези тъкани са представени от така наречената първична меристема (proisomemem), състояща се от протодерма, прокамбий и основната меристема.Первичните проводими тъкани се развиват от прокамбиум. Първите елементи на флоемата са диференцирани от външните, разположени до периферията на прокамбиевите клетки. Първичната флоема е представена от тънкостенни краткотрайни удължени клетки и се нарича протофлоема, а външните й клетки могат да бъдат представени от механични влакна.Основните елементи на ксилемата са трахеиди, по-рядко съдове с пръстеновидни и спираловидни удебеления на стените - по-късно се появяват от вътрешните клетки на прокамбиума и обикновено се определят като протоксилем. В допълнение към проводящите елементи, съдържа паренхимни клетки.По-късно структура, която е повече или по-малко типична за флоема, се диференцира от протофлоема. Външно от протоксилема се образува метаксилен, състоящ се от трахеиди или трахеи с по-дебели лигнифицирани стени.
По този начин, поради активността на прокамбиума и останалата част от върховата меристема, възниква първичната структура на стъблото на растението. При едносемеделни растения целият прокамбиум се обособява в елементи от първични проводящи тъкани. Монокотиледоносни, особено тревисти (тревисти) стъбла имат по-проста структура в сравнение с диковите стъблови растения, те се характеризират най-вече с първична структура.При двудолни растения камбият се образува в средната част на прокамбиалния връв и започват да се образуват вторични проводими тъкани (метафлоем и метаксилем) , чийто обем се увеличава поради разделянето на клетките на камбий. Сноповете, състоящи се само от първични тъкани, като тези на монокотиледони, затворени, снопчета с камбий, характерни за двудолни, са отворени.За билки зърна едносемеделни характеризират с дифузно разпределение на съдови връзки. Провеждащите гроздове затворени, обезпечени, по-рядко концентрични. От механичните тъкани най-развит е склеренхимът, колеленхимът се намира в няколко растения. При тревисти монокотиледони няма вторично уплътняване.
Закръглени (например: слънчоглед)
тетраедални
триъгълна
Многопластови (оребрени) (например: тиква)
Крилати (имат кожени израстъци)
Образуване на стебла
Стъблото, подобно на корена, се формира от първичната апикална меристема. За разлика от корена, конусът на растежа е защитен от околната среда от млади развиващи се листа.Както в корена, клетките на меристемата делят митоза и се специализират в първични постоянни тъкани, но за разлика от корена, в стеблото няма ясно изразени зони на растеж. Специализацията на клетките се случва много бързо, както и в основата на тези клетки се специализират в три от едни и същи области.
От периферията слой от клетки се трансформира в първична тъкан на обвивката; в първичната кора; в центъра има централен аксиален цилиндър. В стъблото, за разлика от корена, централният цилиндър много пъти заема повече място от първичната кора. 
Първична цялостна тъканстеблов епидермис, 1 слой, с устни, с косми. Неговата функция е да защитава основните конструкции.
Първична корастъблото е отговорно за фотосинтезата и придава механична стабилност (издържа на огъване). Идентифицира стъблото в пространството. Състои се от:
1) хлоренхим (от 1 до няколко слоя на тези клетки)
2) коленхим (от 1 или повече слоя). В монокотите е рядкост.
3) ендодерма
Ендодермата на стъблото представлява слой от живи клетки, с целулозни черупки, съдържащи нишестени зърна. Те не са вещество за съхранение, те помагат за навигацията в пространството и затова се нарича стволова ендодерма krahmalonosnymвагина.Освен това при различни растения в първичната кора може да се образува резервоар от секрети, да се появят клетки, носещи кристали, това е пластмасовата част на стъблото.
Централен аксиален цилиндързапочва с рециклиращ склеренхим. Перикът в стъблото се преражда в склеренхим и частично в паренхима. Перицикличният склеренхим може да бъде в напречно сечение под формата на плътен пръстен или под формата на отделни шнурове.
Зад велосипеда са проводящи греди, обикновено колотрални; рядко бикололотрално заобиколено от клетки на основния паренхим.
Местоположението на проводящите лъчи е различно при монокотите и дикотите.
–– при монокотиледони, разположени дифузно (дифузно)
–– в дикоти в кръг
Подобно различно подреждане на проводими греди се дължи на естеството на съединението на проводимите греди на листа и стъблото.
В дикотите проводящите снопчета листа са свързани със стъблото в техния възел. ![]()
При едносемеделни снопове листа влизат в централния цилиндър и се огъват и се съединяват със стъблови снопчета, минаващи през 1-2 възли. 
При двудолни, 2-3 стълбови листа влизат в стъблото от листата, докато при еднодолни, във връзка с паралелно жилене, има много.
В едносемеделни растения всички проводими греди. затворен, т.е. нямат камбий и имат двудолни отворенснопове, т.е. има камбий.
Дебелината на стъблото на едносемеделни растения по цялата дължина остава една и съща, докато при двудолни горната част на стъблото е малко по-малка, отколкото в долната част.
Образуването на тъканите на стъблото на вторичната структура
Вторичната структура на стъблото е характерна за едногодишните и многогодишните тревисти, дървесните двудолни и гимнастиците. При двудолни растения първичната структура е много краткотрайна и с началото на активността на камбия се формира вторична структура. В зависимост от раздела на прокамбиума се формират няколко вида вторична структура на стъблото. Ако нишките на прокамбиума са разделени от широки редове паренхим, тогава се образува снопна структура, ако се обединят така, че да се слеят в цилиндър, се образува необразователна структура.
Фиг. 3.24.Тип лъч на структурата на стъблото на двудолно растение: А - детелина: 1 - епидермис; 2 - хлоренхим; 3 - перицикличен склеренхим с произход; 4 - флоема; 5 - сноп камбиум; 6 - ксилем; 7 - интерстициален камбий
Стрък структура на стеблатасреща се при растения като детелина, грах, пеперуда, копър (фиг. 3.24). Те имат прокамбиални нишки, положени в един кръг около периферията на централния цилиндър. Всеки прокамбиален шнур се превръща в колатерален сноп, състоящ се от първична флоема и първичен ксилем. Впоследствие между флоемата и ксилемата от прокамбиума се образува камбий, който образува елементите на вторичната флоема и вторичния ксилем. Флоемата се отлага към периферията на органа, а ксилемата - към центъра, а ксилемата се отлага повече. Първичната флоема и ксилем остават в периферията на лъча, а вторичните елементи са в съседство с камбия. За стъблата на двудолни растения е характерно образуването на отворени колатерални или биколотерални греди (фиг. 17, виж кол. Вкл.).
Също така за стъблата на двудолни растения, характеризиращи се с диференциране първична кора,която се състои от: коленхим (ъглов (фиг. 18, см. цвят вкл.) или ламеларен), паренхим, носещ хлорофил и вътрешния слой - ендодерма. Нишестето се натрупва в ендодермата; такъв а влагалище, съдържащо нишестеиграе важна роля в геотропната реакция на стъблата. На границата на първичната кора в централния аксиален цилиндър е разположен перицикличен склеренхим- плътен пръстен или парцели под формата на половин дъги над флоемата. Стъблото ядро е изразено и представено от паренхима. Понякога част от ядрото се разрушава с образуването на кухина (виж фиг. 3.24).
Необразователна структурахарактерно за дървесните растения (липа) (фиг. 19, виж кол. вкл.) и много билки (лен). В растежния конус промбамбиалните нишки се сливат и образуват непрекъснат цилиндър, видим в напречно сечение под формата на пръстен. Прокамбиевият пръстен странично образува пръстена на първичната флоема, а вътре е пръстенът на първичния ксилем, между който е положен камбиевият пръстен. Камбиевите клетки се разделят (успоредно на повърхността на органа) и излагат пръстена на вторичната флоема, а навътре - пръстена на вторичния ксилем в съотношение 1:20. Нека разгледаме необразователна структура на примера на многогодишно липово стъбло (фиг. 3.25).
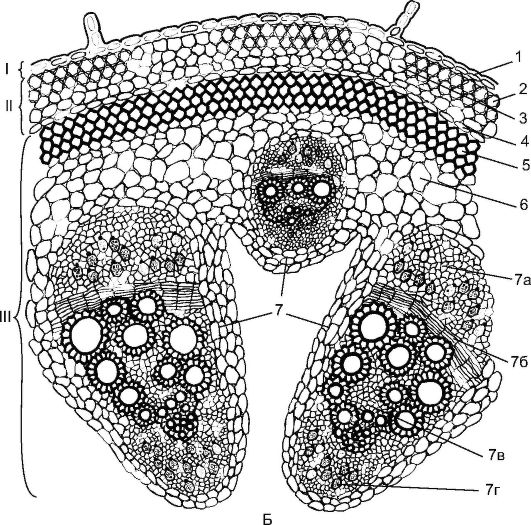
Фиг. 3.24.(продължение) Б - тиква: I - вътрешна тъкан; II - първична кора; III - централен аксиален цилиндър; 1 - епидермиса; 2 - ъглова колелхима; 3 - хлоренхим; 4 - ендодерма; 5 - склеренхим; 6 - основният паренхим; 7 - биколатерален съдово-фиброзен сноп: 7а - флоем; 7b - камбий; 7в - ксилем; 7g - вътрешна флоема
Млад кълнове от липа, образувано през пролетта от пъпка, е покрито с епидерма. Всички тъкани, разположени до камбия, се наричат кора. Кората е първична и вторична. Първична коратя е представена от ламеларен коленхим, разположен непосредствено под епидермиса от непрекъснат пръстен, паренхим, носещ хлорофил и влагалище, носещо единично нишесте. В този слой има зърна от „защитено“ нишесте, което растението не консумира. Смята се, че това нишесте участва в поддържането на растителния баланс.
Централният аксиален цилиндър в липа започва с перицикличен склеренхим над флоемите. В резултат на активността на камбий, вторична кора(от камбий до перидерма), представена от вторичната флоема, медуларните лъчи и паренхима на вторичната кора. Кората на липата се добива, отстранява се в камбия, особено лесно е да се направи това през пролетта, когато клетките на камбия активно се делят. Преди това липовата кора (бастун) се е използвала за тъкане на бастуни, изработка на кутии, перални и др.
Трапецовидната флоема е разделена от триъгълни лъчи на първично ядро, които проникват в дървесината до сърцевината. Съставът на флоемата в липата е разнороден. Той съдържа компонентите на твърдите дървесни дървесни влакна, а мекият лакът е представен от ситовидни тръби със сателитни клетки и голи паренхим. Lub губи способността да провежда органична материя обикновено за една година и се обновява с нови слоеве поради активността на камбия.
Камбият също образува вторични лъчи на ядрото, но те не достигат до сърцевината, като се губят във вторичната дървесина. Основните лъчи служат за радиално движение на водата и органичната материя. В паренхимните клетки на медуларните лъчи до есента се отлагат резервни хранителни вещества (нишесте, масла), които през пролетта се изразходват за растежа на младите леторасти.
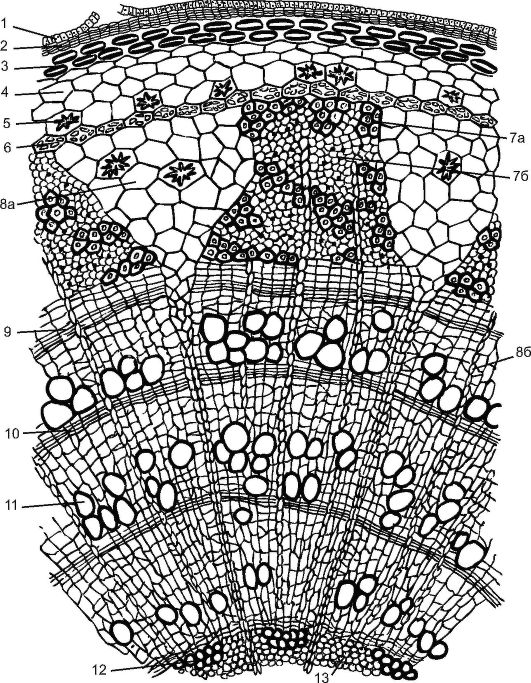
Фиг. 3.25.Напречен разрез на тригодишен клон на липа: 1 - останки от епидермиса; 2 - корк; 3 - ламеларен коленхим; 4 - хлоренхим; 5 - друзен; 6 - ендодерма; 7 - флоема: 7а - твърда лапа, (лакътни влакна); 7b - мека флоема - (ситови тръби със сателитни клетки и голи паренхим); 8а първичен първичен лъч; 8b - вторичен лъч на сърцевината; 9 - камбий; 10 - есенна дървесина; 11 - пролетно дърво; 12 - първичен ксилем; 13 - паренхим на сърцевината
Още през лятото фелогенът се полага под епидермиса и се образува вторична покривна тъкан, перидермата. До есента с образуването на перидерма клетките на епидермиса умират, но остатъците им остават 2-3 години. Наслояването на многогодишния перидерм образува кора.
Слоят ксилем, образуван от камбия от дървесни растения, е много по-широк от слоя флоеми. Дървесината функционира от няколко години. Клетките от мъртва дървесина не участват в поведението на вещества, но те са в състояние да поддържат огромната тежест на короната на растението.
Съставът на дървесината е разнороден, включва: tracheids(фиг. 20, виж цвета на), трахея, дървен паренхими libriform.Дървесината се характеризира с наличието на дървени пръстени.В началото на пролетта, когато в растението възниква активен соков поток, камбият в ксилемата образува широкопросветни и тънкостенни проводими елементи - съдове и трахеиди, и с наближаването на есента, когато тези процеси замръзват и активността на камбия отслабва, се появяват теснолюменни дебелостенни съдове, трахеиди и дървесни влакна. По този начин се образува едногодишен прираст или едногодишен пръстен (от една пружина до друга), ясно различим в напречното сечение. Чрез годишните пръстени можете да определите възрастта на растението (вижте фиг. 3.25).
Характеристики на структурата на стъблото на дикотите:
1) растежа на стъблото в дебелина (поради дейността на камбия);
2) добре диференцирана първична кора (коленхим, хлорофил-носещ паренхим, ендодерма, съдържаща нишесте);
3) двустранни и обезпечени гроздове само отворен тип (с камбий);
4) съдовите влакнести снопове са разположени в пръстен или се сливат (не-постепенна структура);
5) наличието на ядрото;
6) дървесните растения се характеризират с наличието на годишни пръстени в ксилема.
Характеристики на структурата на коренищата на двудолни.Епидермисът може да бъде покриващата тъкан на двудолни коренища, а при многогодишните коренища епидермисът се заменя с перидерма. Първичната кора е представена от паренхима и ендодермата за съхранение с петна от Caspari. Освен това ширината на първичния кортекс е близка до ширината на централния цилиндър. Структурата на централния аксиален цилиндър, сноповете на съдовите влакна и тяхното разположение в него имат същите характеристики, както при надземните стъбла.
В зависимост от условията на околната среда, в които се формира конкретен растителен вид, се създават различни характеристики на структурата на неговите органи, включително на стъблото.
В допълнение, стъблото в някои случаи изпълнява функциите на други органи. В този случай промените в структурата на стъблото са особено значими и обикновено се класифицират като аномалии. Особена структура на стъблото се характеризира с лозя. Лозята са особено богато представени в тропиците, където са най-разнообразни и мощно развити. Но в умерените ширини растат достатъчно лози. Хмел, клематис, бръшлян, лозарска грах, грах, боб, глициния, вратовръзка и др. - са истински лиани, не толкова луксозни, колкото в тропиците, но все пак имат характеристики в структурата на стъблото, присъщи на всички лиани.
Най-характерната особеност на стволовата структура на лозите е доста изразена дисекция на проводящата система и силно развитие на паренхимните ядрени лъчи. Дори пълзящи с добре изразени дървесни стъбла (лози), луковичните лъчи се състоят от паренхим, който се различава малко от паренхима на кората и кората, и тези луковици са много високи, простират се на голямо разстояние по дължината на стъблото, приличат на дълги, вертикално поставени по протежение на стъблото на лентата и др. разчленяване на масива на централния цилиндър на отделни сектори. При типичните дървесни растения основните лъчи поради незначителността на височината им върху тангенциални срезове на дървесина имат формата на къси вретеновидни струпвания от клетки. Още по-рязко разчленяването на централния цилиндър на стъблото се открива в тропическите лози. В някои тропически лози паренхимните клетки, разпръснати на различни места от дървесината на централния цилиндър, започват да се делят и растат, изтласквайки отделни участъци от съдовата система. В резултат на това старите стъбла на такива лози имат много погрешна и капризна структура. Като цяло структурата на стъблото на дървесните лози наподобява структурата на стъблото на някои тревисти растения.
Редица растения, които растат в пустинните райони на Африка и Америка, където вали много рядко и дори за кратко време на годината, формата и структурата на стъблото имат специфични особености. В такива растения стъблата са месести, зелени, приемат различна форма - топка, плоча или оборудвани с широки ламелни ребра. Тези стъбла функционират като листа. Листата често са много малки и понякога имат вид на сухи шипове. Наричат се растения с месести стъбла и намалени листа стволови сукуленти, Сред тях са различни видове бодливи круши, кактуси, еуфория.
Намаляването на листните остриета до сухи шипове води до значително намаляване на изпаряващата се повърхност на цялото растение, състояща се в надземната му част изцяло от особено модифицирано стъбло. Самото стъбло се трансформира в орган, пригоден за натрупване на водни запаси. Това прави възможно растението да съхранява влагата през дъждовни периоди и бавно да я изразходва по време на без дъжда.
В структурата на епителните тъкани на повечето видове сочни стебла има адаптиране към възможното намаляване на загубата на растителна вода чрез транспирация. В същото време системата за провеждане на тъканите трябва да бъде значително намалена. Механичните тъкани вътре в сочните стебла стават ненужни и почти липсват.
Клетъчният сок на сочните стволови клетки се състои от разтвори на различни киселини, захари и лигавични вещества. Тези вещества допринасят за намаляване на добива на вода от растението. Тъканта на сочните стебла е много хомогенна и се състои главно от тънкостенни паренхими, проникнати от слабо развити съдови снопове: има малко съдове във всеки сноп, всички те са с тесно-кухина и примитивни структури. Слабото развитие на съдовите снопове е напълно в съответствие с лекото развитие на листата, тъй като листът в развитието си стимулира образуването на съдовите снопове на стъблото. При типичните, добре изразени сочни стебла на вторичния ксилем, малко се образува поради слаба камбиална активност, съдовата система се състои главно от първични елементи, възникнали в съдовата меристема. Следователно, както физиологично, така и от гледна точка на историята на развитието, формирането на слаба проводяща система в стъблата на сочните растения е доста обяснимо.
Естествено, сред сукулентите няма дървесни форми, всички те са тревисти по своята структура.
В така наречените растения склеренхимен типРазраствайки се в засушливи области, също се развиват нормални листа и стъбла, проводната система се състои от мощно развити влакнести съдови снопове или техни продукти на сливане. Съдовите елементи в такива снопове са широкообхватни, многобройни и по-високо организирани от тези на сукулентите. Вторичните елементи на дървесните растения от тази група са ясно изразени, а в стъблата на тревистите растения се изразява лигнификация на клетъчните стени на главния паренхим. Механичната система е силно развита, анатомичните й елементи са дебелостенни.
Стъблата на растенията, които растат във вода, така наречените водни растения, с относително богато разнообразие от форми, имат една обща структурна особеност: те имат добре развита система от междуклетъчни пасажи и междуклетъчни пространства. Междуклетъчните пространства под формата на повече или по-малко големи кухини образуват система от въздушни проходи, проникващи както в ядрото, така и в кората на стъблото. В същото време ядрото, което заема значителен дял от общия диаметър на стъблото, е силно развито. В водните растения, особено сред представителите на класа от едносемеделни, механичната система понякога надделява над проводящата система. Групите съдови анатомични елементи, събрани в снопове, обикновено са с относително малък размер, докато масивите, състоящи се от механични елементи, концентрирани главно по периферията на стъблото, достигат в сравнение със съдовите снопчета с висока мощност.
Структурата на подземните стъбла, които изпълняват функцията на запаси, коренища и грудки, се отклонява още по-рязко от нормалния тип. Коренището е многогодишно и обикновено достига много по-голяма дебелина от надземните издънки, където и да растат, във вода или друга среда. Във връзка с основната цел на коренището - да служи като хранилище на резервни вещества - паренхиматизацията на нейните тъкани е най-характерната характеристика на структурата. Ядрото в тъканите на типичните коренища е ядро, състоящо се от кръгли паренхимни клетки, тънкостенни, с малки междуклетъчни пространства между тях. Със своята масивност ядрото в коренищата преобладава над другите тъкани. Механичната и проводима тъкан, подчинявайки се на силата на ядрото и кората, са по-слабо изразени в коренището, отколкото в надземните издънки.
Структурата на коренището като дългосрочен изстрел е съставена под влияние на различни формиращи фактори на външната среда. Влиянието на тези фактори може да бъде както директно, така и чрез взаимовръзка в развитието на растителните органи: например, степента на развитие на листата, образуването на цветни клонки, освобождаването на резервни вещества влияе върху структурата на ризомите. Особено забележима е зависимостта на структурата на коренището от степента на развитие на листата върху тях.
По време на подземния растеж, когато върху коренището се намират само люспести подобни малки листовки, се образува много слаба проводяща система, в ксилемовата част, състояща се от редки съдове с тясна кухина с примитивна структура. След повишаване на плодоносната издънка с нормални големи листа, активността на камбия се увеличава и в резултат се развива дървесина, както и броят и лумена на кръвоносните съдове, които имат по-съвършена структура в сравнение с образуваните преди растежа на въздушните издънки.
Особена морфологична особеност на коренището са много късите междувъзлия. Очевидно, незначителността на дължината на междувъзлия е в тясна връзка със слабата диференциация на листата на коренището.
На коренището, според неговата морфология, в някои отношения, друга модификация на подземния изстрел - клубена - изглежда подобна. Основната разлика между коренището и клубена е в продължителността на съществуването: коренището е дългосрочно кълнове, клубените обикновено съществуват от една до две години. Следователно, само тези структурни особености, които коренищата имат на първия етап от образуването на техните тъкани, се разкриват в клубена. Преобладаването на паренхима в клубена е дори по-ясно изразено, отколкото в коренището.
По степента на паренхимация тъканта на клубените прилича на тъкан на стволови на сукулентни растения. Точно както в стъблата на сукулентни растения, цялата маса на клубеновата тъкан, с изключение на обвивката (перидерма), е тъкан за съхранение, проникната от недоразвити и примитивно конструирани снопове на проводящата система. Наред с намаляването на проводящата тъкан в нормален клубен, механичната тъкан обикновено не се образува. Тъканите, съставящи клубени, са конструирани еднакво и адаптирани да изпълняват само една функция - отлагане на запаси.

Типичен пример за структурата на клубена може да бъде картофен клубен. На фигурата е показан фрагмент от напречно сечение на клубен от един от сортовете картофи (“Voltman”), от централната му част. За удобство на демонстрация на тъканна структура, нишестето, което обилно запълва всички клетки, не е изобразено. Паренхимът на клубена само на едно място се разрязва от тясна ивица от по-малки клетки, състоящи се от четири групи. Във всяка група има 1-3 съдове с тясна кухина, останалата част от тъканта се състои от меристемни клетки, сред които има няколко ситообразни тръби. Съдовете с недоразвита структура са примитивни, някои от които са в състояние на унищожаване и унищожаване.
Тази тъкан, състояща се от примитивни и недоразвити съдови елементи, се намира в пръстен около клубена, представляващ централен цилиндър. Външният ъгъл е основната кора, вътре в ядрото. В грудки, които имат само мащабна прилика на листа, вторичните тъкани не се образуват в резултат на активност на камбий.
Ако откриете грешка, моля, маркирайте част от текста и кликнете Ctrl + Enter.