











Клетки эпидермиса почти непроницаемы для воды и газов благодаря своеоб-разному устройству их наружной стенки. Как же осуществляются газообмен между растением и наружной средой и испарение воды — процессы, не-обходимые для нормальной жизнедеятельности растения? Среди клеток эпидермиса встречаются характерные образования, называемые устьи-цами .
Устьице — щелевидное отверстие, окаймленное с двух сторон двумя замыкающими клетками, имеющими большей частью полулун-ную форму.
Клетки эти живые и со-держат хлорофилловые зерна и кру-пинки крахмала, отсутствующие в других клетках эпидермиса. Особен-но много устьиц на листе. Вид усть-ица с поверхности и в разрезе изоб-ражен на рисунке 40. На поперечном разрезе видно, что непосредственно под устьицем внутри ткани листа находится полость, называемая дыхательной . В пределах щели за-мыкающие клетки более сближены в средней части клеток, а выше и ниже они дальше отступают друг от друга, образуя пространства, называемые передним и задним дво-риком .
Замыкающие клетки способны увеличивать и сокращать свои разме-ры, благодаря чему устьичная щель то широко раскрывается, то сужива-ется или даже совсем бывает закрыта.
Таким образом, замыкающие клетки являются аппаратом, регулирующим процесс открывания и закрывания устьиц.
Как же осуществляется этот процесс?
Стенки замыкающих клеток, обращенные к щели, утолщены значи-тельно сильнее, чем стенки, обращенные к соседним клеткам эпидер-миса. Когда растение освещено и имеет избыток влаги, в хлорофилловых зернах замыкающих клеток происходит накопление крахмала, часть ко-торого превращается в сахар. Сахар, растворенный в клеточном соке, при-тягивает воду из соседних клеток эпидермиса, вследствие чего в замы-кающих клетках повышается тургор. Сильное давление приводит к выпячиванию стенок клеток, примыкающих к эпидермальным, а про-тивоположные, сильно утолщенные стенки выпрямляются. Вследствие этого устьичная щель раскрывается, и газообмен, а также испарение воды увеличиваются. В темноте или при недостатке влаги тур-горное давление уменьшается, замыкающие клетки принимают прежнее по-ложение и утолщенные стенки смыкаются. Щель устьица закрывается.
Устьица расположены на всех молодых неодревесневших наземных ор-ганах растения. Особенно много их на листьях, причем здесь они располо-жены, главным образом, на нижней поверхности. Если лист расположен вертикально, то устьица развиваются с обеих его сторон. У плавающих на поверхности воды листьев некоторых водных растений (например, кувшинки, кубышки) устьица расположены только на верхней стороне листа. Материал с сайта
Число устьиц на 1 кв. мм листовой поверхности в среднем равно 300, однако иногда достигает 600 и более. У рогоза (Typha ) насчитывают свыше 1300 устьиц на 1 кв. мм. Листья, погруженные в воду, устьиц не имеют. Расположены устьица чаще всего равномерно по всей поверхности кожи-цы, но у некоторых растений собраны группами. У однодольных растений, а также на хвоинках многих хвойных они расположены продольными рядами. У растений засушливых областей неред-ко устьица бывают погружены в ткань листа.
Развитие устьиц обычно происхо-дит следующим образом. В отдельных клетках эпидермиса образуются дуго-образные стенки, разделяющие клетку на несколько более мелких так, что центральная из них становится родо-начальницей устьиц. Эта клетка раз-деляется продольной (по оси клетки) перегородкой. Затем эта перегородка расщепляется, и образуется щель. Ог-раничивающие ее клетки становятся замыкающими клетками устьица. У не-которых печеночных мхов имеются свое-образные устьица, лишенные замыкаю-щих клеток.
Замыкающие и побочные клетки устьиц.
Устьица, или устьичный аппарат – это две замыкающих клетки бобовидной формы с разноутолщенными стенками и с расположенным между ними межклетником - устьичной щелью (внутренние стенки устьичных клеток, обращённые к щели толстые, а наружные тонкие).
Реже рядом с ними находятся побочные клетки эпидермы, отличающиеся от ее основных клеток.
Под устьицем расположена подустьичная воздушная полость, окруженная клетками паренхимы и связанная с системой межклетников органа.
Характерно:
1) Есть хлоропласты (фотосинтез углеводов) и митохондрии (синтез АТФ). Необходимы, т.к. раскрытие устьичной щели происходит активно с затратой энергии АТФ, а смыкание пассивно.
2) За счёт разноутолщенности стенок замыкающих клеток образуется щель.
Следовательно, осн. функции :
1 - транспирация (регулируемое испарение воды)
(Самостоятельно, изучить механизм работы устьиц).
Механизм открывания и закрывания устьичной щели основан на осмотических явлениях, а также принимают участие все органоиды протопласта замыкающих клеток.
- - K-Na – насос (механизм быстрого реагирования). При высоком содержании в растении воды в плазмалемме замыкающих клеток работают белки-переносчики К + , активно, с затратой энергии АТФ перекачивающие ионы К + из окружающих клеток в цитоплазму, и далее они поступают в вакуоли замыкающих клеток устьиц. При дальнейшем повышении концентрации ионов К + , вслед за ними, пассивно, по градиенту концентрации, в замыкающие клетки устьиц из окружающих клеток поступает вода. Объём вакуолей увеличивается, осмотическое давление в замыкающих клетках повышается, оболочки клеток растягиваются. Из-за того, что оболочки замыкающих клеток утолщены неравномерно, наружная оболочки растягиваются сильнее, а внутренние (обращённые друг к другу) не могут растянуться и расходятся, между ними образуется щель – устьица открываются. При недостатке воды в растении белки переносчики ионов К + прекращают свою работу. К-Nа – насос останавливается. При этом ионы К + по градиенту концентраций пассивно покидают устьичные клетки и уходят в окружающие клетки, вслед за ними уходит вода. Замыкающие клетки теряют тургор и пассивно смыкаются – устьица закрываются.
Т.о., открытие устьичной щели происходит активно, с затратой энергии, источником которой являются углеводы, накапливаемые хлоропластами. Закрытие щели происходит пассивно, без затрат энергии.
- – Суточное изменение концентрации сахаров (днем много, к концу ночи – мало) (т.к. есть хлоропласты). В течение дня в хлоропластах замыкающих клеток идёт фотосинтез. Следовательно, на свету в них повышается концентрации глюкозы и по мере её накопления, к вечеру, по градиенту концентрации внутрь клеток поступает вода. Замыкающие клетки приобретают упругое тургорное состояние, устьица раскрываются. Ночью фотосинтез прекращается, глюкоза расходуется на дыхание замыкающих клеток, её концентрация снижается. Из-за этого по градиенту концентрации вода покидает замыкающие клетки, они теряют тургор и спадаются. Устьичная щель закрывается. Утром процесс повторяется. Сейчас считается, что этот механизм ограничен, основным является работа К-Nа – насоса.
Разные растения выработали определённый ритм работы устьичного аппарата. У большинства растений устьица открыты и днём и ночью, закрываясь лишь при уменьшении содержания воды. В жаркие часы устьица обычно закрыты, но у растений пустынь они открыты (с помощью испарения они охлаждают поверхность листьев и активнее поглощают воду). У некоторых растений, например у каланхоэ, устьица открываются ночью и закрываются днём.
Типы устьиц:
Важны при установлении родства таксонов растений.
1.- анамоцитные устьица – имеют побочные клетки, не отличающиеся от осн. клеток эпидермы (хар-ны для всех групп растений, кроме хвощей).
2.- диацитные устьица – имеют две побочные клетки, общая стенка которых перпендикулярна устьичной щели (есть у ряда цветковых, у сем. Губоцветных и Гвоздичных).
3. – парацитные устьица - побочные клетки располагаются параллельно замыкающим клеткам и устьичной щели (есть папоротников, хвощей и ряда цветковых растений).
4. – анизоцитные устьица – замыкающие клетки окружены тремя побочными, одна из которых или крупнее, или меньше остальных (только у цветковых растений).
5. – тетрацитные устьица - имеют 4 побочные клетки (есть у однодольных).
6. – энциклоцитные устьица – побочные клетки образуют узкое кольцо вокруг замыкающих клеток (есть у папоротников, голосеменных и ряда цветковых).
Если устьица открыты, испарение идет так, будто эпидермы нет совсем.
Число и расположение устьиц варьирует у разных растений. В среднем, у растения 100-700 устьиц на 1 мм 2 . Обычно, у наземных растений большая часть устьиц расположена на нижней стороне листьев, сверху их очень мало или нет совсем. Это объясняется следующими причинами: 1) сверху листья сильнее нагреваются, следовательно, испарение будет идти интенсивнее и потери влаги будут больше; 2) углекислый газ образуется в почве и поднимается вверх, попадая сразу в устьица, к тому же, он тяжелее воздуха и скапливается обычно в нижних воздушных слоях.
У растений с вертикально стоящими листьями (ребром), например, у эвкалипта, устьица распределены по обеим сторонам листа. У водных растений с плавающими листьями устьица расположены на верхней стороне.
- Трихомы – волоски, выросты и производные эпидермы.
Форма, строение – систематический признак для определения видов и родов растений.
Бывают: железистые и кроющие.
Железистые трихомы образуют и накапливают экскреты и секреты. Это эфирные масла (герань, мята и др.) или защитные в-ва (крапива). Капля масла выделяется под кутикулу, накапливается там, затем при разрыве кутикулы выходит наружу. Кутикула восстанавливается и под ней накапливается новая капля эфирного масла.
Функции:
1-защитная
2-терморегуляция
3- бактерицидная (эфирные масла – фитонциды)
Кроющие трихомы состоят:
1- часто из мертвых клеток заполненных воздухом (одно- или многоклеточные мертвые трихомы)
Функции: отражение солнечных лучей, благоприятные микроклиматические условия для работы устьиц, уменьшение нагрева. Следовательно, снижение транспирации и мех. защита (растения пустынь и степей).
2- редко из живых клеток.
Функции: увеличивают общую испар. поверхность. Следовательно, увеличение испарения (растения влажных тропиков и тропических болот).
Т.о., эпидерма как многофункциональная сложная ткань выполняет функции только в живом виде.
1. Регуляция газообмена и транспирации (между растением и окр. средой)
2. Защитная (мех., бактерицидная, от излишнего испарения воды).
3. Выделительная (выделяет соли, воду, эфирные масла).
4. Поглотительная (всасывающая) (вода с мин. в-вами----внекорневые подкормки).
5. Биосинтетическая (синтез орг. в-в: кутинов, восков и т.п.)
Устьица - это отверстия в эпидермисе, через которые происходит газообмен. Они находятся в основном на листьях, но имеются также и на стеблях. Каждое устьице окружено двумя замыкающими клетками, которые в отличие от обычных эпидермальных клеток содержат хлоропла-сты. Замыкающие клетки контролируют величину отверстия устьица за счет изменения своей тургесцентности. Внешний вид устьиц и замыкающих клеток хорошо видны на микрофотографиях, полученных с помощью сканирующего электронного микроскопа.
В статье мы уже говорили, как выглядят клетки эпидермиса, замыкающие клетки и устьица , если их рассматривать сверху в световом микроскопе. На рисунке приведено схематическое изображение устьица в разрезе. Видно, что стенки замыкающих клеток неравномерно утолщены: стенка, которая ближе к отверстию устьица, называемая вентральной, толще, чем противоположная, называемая дорсальной. Кроме того, целлюлозные микрофибриллы в стенке ориентированы таким образом, что вентральная стенка менее эластична, чем дорсальная. Некоторые микрофибриллы образуют как бы обручи вокруг замыкающих клеток, похожих на сардельки.
Эти обручи не эластичны, и по мере заполнения клетки водой,т. е. роста ее тургора, они не дают увеличиваться ее диаметру, позволяя растягиваться только в длину. Но поскольку замыкающие клетки соединены своими концами, а тонкие дорсальные стенки растягиваются легче, чем толстые вентральные, клетки приобретают полукруглую форму. В результате между двумя соседними замыкающими клетками появляется зазор, называемый устьичной щелью. Такой же эффект наблюдается, если надувать два скрепленных концами продолговатых воздушных шарика, наклеив вдоль их соприкасающихся сторон липкую ленту (имитация нерастяжимой вентральной стенки). Для полноты картины можно неплотно обмотать их такой же лентой по спирали, имитировав целлюлозные обручи.
Когда замыкающие клетки теряют воду и тургор, устьичная щель закрывается. Каким образом происходит изменение тургесцентности клеток, пока не ясно.
Согласно классической, так называемой «сахаро-крахмально й» гипотезе, в светлое время суток в замыкающих клетках повышается концентрация водорастворимых Сахаров, а следовательно, осмотический потенциал их становится более отрицательным, что стимулирует поступление в них воды путем осмоса. Однако никому еще не удалось показать, что в замыкающих клетках накапливается достаточное количество сахара, чтобы вызвать наблюдаемые изменения осмотического потенциала.
Недавно было установлено, что днем на свету в замыкающих клетках интенсивно накапливаются катионы калия и сопутствующие им анионы: они и играют роль, отводившуюся раньше сахару. До сих пор неясно, уравновешиваются ли при этом их заряды. У некоторых изученных растений отмечалось накопление на свету большого количества анионов органических кислот, в частности малата. Вместе с тем размеры крахмальных зерен, появляющихся в темноте в хлоропластах замыкающих клеток, уменьшаются. Дело в том, что крахмал на свету (необходимы синие лучи спектра) превращается в малат, возможно, по следующей схеме:
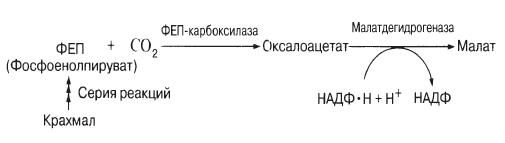
У некоторых видов, например у лука, крахмала в замыкающих клетках нет. Поэтому при раскрытых устьицах малат не накапливается, а катионы, по-видимому, поглощаются вместе с неорганическими ионами типа хлорид-ионов.
В темноте калий (К+) выходит из замыкающих клеток в окружающие эпидермальные клетки . В результате водный потенциал замыкающих клеток растет, и вода из них устремляется туда, где он ниже. Тургор замыкающих клеток падает, они изменяют форму, и устьичная щель закрывается.
Некоторые вопросы пока остаются без ответа. Например, почему на ceeiy в замыкающие клетки поступает калий? Какова роль хлоропластов, кроме накопления крахмала? Возможно, калий поступает внутрь благодаря «включению» АТФ-азы, локализованной в плазмалемме. Судя по некоторым данным, этот фермент активируется синим светом. Возможно, АТФаза нужна для откачивания из клетки протонов (Н+), а катионы калия движутся в клетку для уравновешивания заряда (аналогичный, обсуждаемый в разд. 13.8.4 насос работает во флоэме). Действительно, как и предполагает эта гипотеза, внутри замыкающих клеток рН на свету падает. В 1979 г. было показано, что в хлоропластах замыкающих клеток конских бобов (Vtcia faba) нет ферментов цикла Кальвина, а тилакоидная система развита слабо, хотя хлорофилл там имеется. Следовательно, обычный Сз-фотосинтез не работает, крахмала таким путем не образуется. Это, вероятно, объясняет, почему крахмал образуется не днем, как в обычных фотосинтезирующих клетках, а ночью.
Для соприкосновения листа с атмосферой имеются поры - устьица. Устьице - это отверстие (щель), ограниченная двумя замыкающими клетками. Устьица встречаются у всех наземных органов растения, но больше всего у листьев. Каждая замыкающая клетка устьица в отличие от клеток эпидермиса имеет хлоропласта. В них происходит фотосинтез, хотя с меньшей интенсивностью, чем в клетках мезофилла. Устьица - одно из оригинальных приспособлений, обладающих способностью открываться и закрываться в зависимости от насыщенности замыкающих клеток водой . Обычно устьичные отверстия ограничены двумя замыкающими клетками , стенки которых неравномерно утолщены. У двудольных растений замыкающие клетки бобовидной, или полулунной, формы, при этом их внутренние прилегающие друг к другу клеточные стенки более толстые, а внешние - более тонкие. Протопласты замыкающих клеток связаны в единое целое перфорациями в основании граничащих общих стенок. Когда воды мало, замыкающие клетки плотно прилегают друг к другу и устьичная щель закрыта. Когда воды в замыкающих клетках много, то она давит на клеточные стенки, и более тонкие стенки растягиваются сильнее, а более толстые втягиваются внутрь, между замыкающими клетками появляется щель.
В последнее время доказано, что для движения устьиц большое значение имеет также расположение микрофибрилл целлюлозы . Если обычно в клетках листьев целлюлозные фибриллы ориентированы в длину и в этом направлении утолщены, то в замыкающих клетках устьиц микрофибриллы организованы радиально, что усиливает устойчивость к процессу растяжения.
У злаков строение замыкающих клеток несколько иное. Они представлены двумя удлиненными клетками, на концах которых стенки более тонкие. При насыщении водой более тонкие стенки на концах растягиваются и раздвигают замыкающие клетки, благодаря чему образуется щель.
Число устьичных отверстий колеблется в зависимости от вида растений от 10 до 600 на 1 мм2 листа. У многих растений (75% видов), в том числе для большинства древесных, устьица расположены на нижней стороне листа. Диаметр устьичных щелей составляет всего 3-12 мкм. Устьица соединяют внутренние пространства листа с внешней средой. Вода поступает в лист через сеть жилок, в которых расположены сосудистые элементы. Возможны три пути испарения:
- через устьица - устьичная,
- кутикулу - кутикулярная,
- через чечевички - лентикулярная транспирация.
Впервые разграничение на кутикулярную и устьичную транспирацию было введено в 1877 г.
Основные типы устьичного аппарата листа растений.
- аномоцитный (у всех высших растений, кроме хвощей),
- диацитный (у папоротникови цветковых),
- парацитный (у папоротников, хвощей, цветковых и гнетовых),
- анизоцитный (только у цветковых),
- тетрацитный (главным образом у однодольных),
- энциклоцитный (у папоротников, голосеменныхи цветковых).
Устьица представляют собой высокоспециализированные образования эпидермы, состоящие из двух замыкающих клеток, между которыми имеется своеобразный межклетник, или устьичная щель.
Щель может расширяться и сужаться, регулируя транспирацию и газообмен. Под щелью располагается дыхательная, или воздушная, полость, окруженная клетками мякоти листа. Клетки эпидермы, примыкающие к замыкающим, получили название побочных, или околоустьичных . Они участвуют в движении замыкающих клеток. Замыкающие и побочные клетки образуют устьичный аппарат .
Число и распределение устьиц на листе или побеге варьируют в зависимости от вида растений и условий жизни. Число их обычно колеблется от нескольких десятков до нескольких сотен на 1 кв.мм поверхности.
Механизм движения замыкающих клеток весьма сложен и неодинаков у разных видов. У большинства растений при недостаточном водоснабжении в ночные часы, а иногда и днем тургор в замыкающих клетках понижается и щель замыкается, снижая тем самым уровень транспирации. С повышением тургора устьица открываются. Считают, что главная роль в этих изменениях принадлежит ионам калия. Существенное значение в регуляции тургора имеет присутствие в замыкающих клетках хлоропластов. Первичный крахмал хлоропластов, превращаясь в сахар, повышает концентрацию клеточного сока. Это способствует притоку воды из соседних клеток и переходу замыкающих клеток в упругое состояние.
Общая площадь устьичных отверстий составляет лишь 1-2% площади листа. Несмотря на это, транспирация при открытых устьичных щелях достигает 50-70% испарения, равного по площади открытой водной поверхности.