7. Vacuoles. Composition and properties of cell sap. Osmotic pressure, turgor and plasmolysis.
Vacuoles- these are cavities bounded by a membrane; well expressed in plant cells and present in protozoa. They arise in different areas of the endoplasmic reticulum. And they gradually separate from it. Vacuoles maintain turgor pressure; cellular or vacuolar sap is concentrated in them, the molecules of which determine its osmotic concentration. It is believed that the initial products of synthesis - soluble carbohydrates, proteins, pectins, etc. - accumulate in the cisterns of the endoplasmic reticulum. These clusters represent the rudiments of future vacuoles. As a result of metabolism, vacuoles are formed in a plant cell - spaces filled with a solution of various substances - waste products of the protoplast. This solution is cell sap. In young cells there is little cell sap and the vacuoles look like very small bubbles of a viscous colloidal nature, but as the cell grows they liquefy, enlarge, and merge with each other. Eventually, one large vacuole is formed in the cell, and the cytoplasm encloses it in a thin layer and is located wall-to-wall. The vacuole is separated from the cytoplasm tonoplast.
Chemical composition of cell sap varies greatly depending on the type of plant. A huge variety of chemicals isolated from plants and having medicinal properties are found in cell sap. Cell sap is more likely to acid reaction.
Cell sap contains 2 groups of substances: Products of primary exchange : carbohydrates (mono- and disaccharides - glucose, fructose, sucrose), simple soluble proteins, fats in the form of glycerol and fatty acids.
Secondary products : Glycosides cell sap - compounds of certain sugars (usually glucose) with alcohols, aldehydes, phenols and other organic substances. When exposed to air under the influence of enzymes, they quickly disintegrate, often releasing a pleasant odor. This explains the smells of tea, coffee, cocoa, tobacco, mustard, and vanilla. Glycosides include the following substances: amygdalin (in almond and apricot seeds); saponins used as detergents (soapwort); coumarins - in sweet clover leaves, etc.; cardiac glycosides - in foxglove leaves.
Tannins(tannins) are complex organic nitrogen-free compounds with an astringent taste. They have antiseptic properties, which protects plants from damage by microorganisms. Widely distributed in flora: in oak bark 10-20%, in tea leaves 15-20%, willow bark 9-13%, in persimmon, quince, dogwood fruits. They are used in medicine as an astringent, in the textile industry for dyeing fabrics dark brown, and in the tanning industry for tanning leather.
Alkaloids- nitrogenous salts of organic acids: malic, tartaric, etc. are insoluble in alkalis, soluble in water. Formed in all parts of the plant: in the roots and leaves of belladonna (atropine), in the seeds and milky juice of poppy - papaverine, morphine, codeine; tobacco leaves contain nicotine; in potato tubers - solanine; in fly agaric - muscarine.
Widely used in medicine and agriculture; in small doses they have a stimulating effect on the nerve centers, in large doses they have a paralyzing effect. Quinine - against malaria, strychnine - stimulates muscle activity; cocaine is an analgesic; morphine is an analgesic and sleeping pill; papaverine - vasodilator; nicotine - used in agriculture to control insects. Plants containing alkaloids are poisonous and are not eaten by animals. In cells containing alkaloids, spores and germs of microorganisms do not develop, plants are not affected by fungal and bacterial flora (protective role).
Cell sap is rich in various organic acids: malic, tartaric, oxalic, citric, amber, etc. Functions are varied: they participate in the respiration process, partly act as phytoncides and antibiotics, protecting the plant from damage by fungi, viral, bacterial diseases, providing plant taste and smell due to volatile acids: formic , oil, vinegar.
Substances related to carbohydrates are widespread in cell sap - pectins. Easily gelatinous, used in the confectionery industry. In pharmacy - for the preparation of a number of dosage forms (as an emulsifier in emulsions, in pills - as a binding component).
Various coloring substances accumulate in the cell sap - pigments, specific for each plant species. Anthocyanins are the most common and are present in the form of glycosides. Depending on the reactions that it undergoes in the cell sap with salts, tannins, and acids, it gives different colors to the cell sap. The extraordinary variety of colors of flowers in plants, as well as leaves, is most often associated with anthocyanins. Red poppies, red clover heads, blue and blue hyacinths, blue cornflowers - all this is created by anthocyanin. Anthocyanin should not be mixed with chlorophyll, carotene, xanthophyll and other plastid pigments. In addition to attracting insects, anthocyanin has a protective effect against low temperatures and harmful short-wavelength waves. Of the yellow pigments, anthochlor is found in cell sap (in the flowers of yellow poppy, mullein, toadflax, and in citrus fruits).
Essential for cell life are vitamins. This group of organic compounds of diverse chemical nature, plant, and less often animal origin, is closely related to enzymes; grouped based on their physiological effects on the body. They are divided into two groups: water-soluble (C, B) and fat-soluble (A, D, E). The highest content of vitamins is observed in leaves, ripened fruits, and roots. Some (E) are contained in the embryos of seeds, others (D) - in germinating seeds.
Minimum doses are necessary for the normal functioning of the plants themselves (to maintain growth, regulate respiration, metabolism, etc.).
The protoplast of a plant cell also produces a special group of substances that have the property of enhancing physiological processes. Such substances are called phytohormones. Phytohormones have been identified that enhance growth, cell division, and sexual functions.
Growth hormones - auxins increase the access of oxygen, the flow of nutrients to embryonic tissues and create conditions for growth processes. Studied chemical composition auxins, first isolated and then synthesized artificial heteroauxin, which increases the yield of cucumbers, tomatoes, peppers, hemp and other industrial and vegetable crops.
Entry of substances into the plant cell. The vital activity of the body, all organs and cells is possible only with metabolic processes continuously occurring in them. The cell absorbs substances from the environment and at the same time transfers the products formed in it to neighboring cells or releases them into the external environment.
The ability of the protoplast to continuously exchange with the environment bears the features of selectivity. Of the large number of substances located outside the cell, under normal conditions only certain compounds penetrate into it in certain proportions. Accordingly, only certain waste products are released by the cell into the environment. In the phenomena of absorption and release of substances by a cell, the processes of diffusion and osmosis play an important role. As is known, the particles of the substances that make up protoplasm have a certain cellular energy, which is the reason for their continuous movement. The movement of dispersed matter from one part of a system to another is called diffusion. This is not a chaotic movement of molecules, but a directional one, the nature of which is determined by a number of factors: the activity of diffusing molecules, the gradient of concentrated solutions; the rate of diffusion is determined by the size and mass of the molecules, the viscosity of the medium, temperature and other conditions, the composition and properties of other compounds in the solution. The complexity and heterogeneity of the structure of protoplasm determines the unequal rate of diffusion in different parts of the same cell. If the diffusing substance encounters a membrane with different permeability for the solvent and solute on its way, the movement of substances in such a system becomes more difficult.
Being an obstacle to the free diffusion of electrolytes, it ensures a constant difference in concentration between the cell sap and the solution surrounding the cell. The penetration of liquid and soluble substances through semi-permeable partitions is called osmosis. The phenomena of adsorption and desorption are of primary importance in the process of osmosis. They are accompanied by electroosmotic processes. Osmotic pressure in a cell does not depend on protoplast colloids, but on solutions of various salts, sugars, and amino acids in cell sap. For any dissolved salts to penetrate into the cell from the outside, it is necessary that the osmotic pressure of the cell sap be higher than in the saline solution surrounding the cell. Salts (electrolytes) enter the cell not in the form of molecules, but as individual ions that are adsorbed on the surface of semi-permeable membranes due to its electrical potential. Ions also have their own charges, and the larger they are, the more difficult it is for them to penetrate into the cell. The adsorbed ions are then desorbed onto the inner wall of the plasmalemma and transferred to the mesoplasm. Sorption processes are of an exchange nature. The intensity of these phenomena depends on the respiration of cells. The energy released during the stepwise breakdown of substances during respiration is used to a large extent for the sorption functions of cells.
If living cell put into a highly dilute aqueous solution of saltpeter, an osmotic interaction immediately begins between the cell sap and the surrounding solution. Cell sap, which is a solution of different substances in different concentrations, will have a higher osmotic pressure than the external solution and will attract water from it. Cell sap, having increased in volume, will put pressure on the cytoplasm, the latter on the cell membrane, stretching it in all directions. Possessing elasticity, the shell will resist the pressure of cell sap. Since the shell has limited elongation, the resistance will increase as the pressure from the addition of water increases. At a certain moment, this resistance force will balance the osmotic pressure, although the concentration of both solutions will not yet be homogeneous. The state of tension in the cell membrane is called turgor, and the pressure is turgor.
The degree of turgor depends on the difference in osmotic pressure inside and outside the cell and on the elasticity of the membrane. The combined turgor of the mass of cells in the plant’s body creates tension and elasticity of the entire plant, helps the stems maintain an upright position, maintain the mass of leaves, withstand wind, storms, showers, and orient the leaves in relation to the light. In a word, turgor ensures the normal physiological state of the plant.
The difference in osmotic pressure inside and outside the cell provides the suction force of the cells.
The opposite of turgor occurs if the cell is placed in a strong solution of table salt, more concentrated than cell sap. In this case, compression of the shell and protoplast will begin, but since the membrane is less elastic, its compression will soon stop, while the cytoplasm, continuing to contract, will move away from the cell wall and take the form of a lump inside the cell. This phenomenon is called plasmolysis. Plasmolysis in plant tissues makes them sluggish, organs become flabby. It can be curved (protoplast is round); concave (the protoplast in some places does not tear away from the shell, but is partially pulled inward); convulsive (without a specific pattern).
If you place a plasmolytic cell in clean water, the opposite phenomenon of plasmolysis is observed - deplasmolysis.
Under certain conditions, when cell turgor is lost, cytorhiz, when the entire cell (with the membrane) contracts. It is observed when plants wilt and is not a consequence of water loss by osmosis, but the result of water evaporation.
The protoplasm of the cell secretes a watery liquid that does not mix with the protoplasm. This fluid is called cell sap (see page 71). Cavities in the protoplasm filled with cell sap are called vacuoles. The layer of protoplasm bordering the vacuoles - the tonoplast - is sometimes called the vacuole shell.
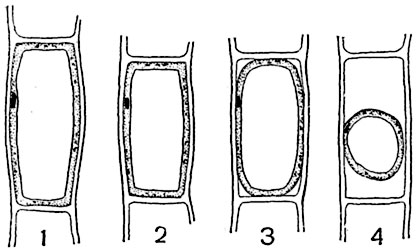
Usually, in a young cell that has just emerged from the embryonic state, filled with a protoplast, there are numerous but very small vacuoles of a round or elongated shape (Fig. 1, 3, 39), filled with water with substances dissolved in it and colloids in a gel state. Then the vacuoles enlarge and turn into a system of tubules filled with cell sap (Fig. 10).
Subsequently, the vacuoles partially separate, increase in size, decreasing in number, merge with one another and become rounded (Fig. 3, 4, 39), the amount of water in them increases, dissolved substances pass for the most part into a sol state, then real solutions appear. Later, the vacuoles usually merge into one large central vacuole; protoplasm at this time is located in the wall layer, where the nucleus, plastids, and chondriosomes are located (Fig. 1.39). In other cases, the nucleus takes place in the center of the cell; the protoplasm surrounding it is connected to the wall protoplasm by strands and plates passing through the cavity of the vacuole (Fig. 4, 12).
1 - embryonic stage: O- shell, P- protoplasm, I- nucleus with nucleolus; 2 - appearance of vacuoles ( V); 3 - vacuoles enlarge, their number increases; 4 - vacuoles merge, part of the protoplasm forms thin strands; 5 - vacuoles merged into one large central vacuole ( color), protoplasm with a nucleus and nucleolus formed a wall layer ( pm); 6 - only the shell remains of the cell ( O). The volume of the cell cavity increases from the first stage to the last.
It was already noted above that the protoplasm of a living cell has the so-called selective permeability or semi-permeability. This means that in some cases only water molecules can pass through the protoplasm into the vacuole or from the vacuole into the external environment, and for large molecules of organic and inorganic compounds living protoplasm turns out to be impenetrable. The intensity of water penetration through protoplasm depends on osmotic pressure fluid in the vacuole and in the external environment, or more correctly, from the difference in osmotic pressures inside and outside the cell. The higher the concentration of the solution (the degree of dissociation of ions), the higher its osmotic pressure, i.e., the greater the difference in the concentrations of solutions separated by a semi-permeable partition, the more intensely water will penetrate towards the solution with a higher concentration. The penetration of water through living protoplasm is mainly subject to the laws of osmosis. Osmotic pressure is measured in atmospheres.
The penetration of water through a semi-permeable partition (membrane) can occur until the osmotic pressure of liquids on both sides of it becomes equal, i.e., the concentration of both solutions becomes the same, or until the osmotic pressure on one side is balanced by the hydrostatic fluid pressure on the other side of the semipermeable partition. Thus, based on the hydrostatic pressure of the corresponding solution (in atmospheres), the osmotic pressure of the cell sap can also be measured.
The force with which water penetrates into the vacuole of any living cell is called suction force.
As water is absorbed, the volume of the vacuole and the entire protoplast increases; Because of this, the protoplast produces pressure on the cell membrane - the so-called turgor pressure or turgor. Sucking force, turgor pressure and osmotic pressure depend on one another: suction force is equal to the difference between osmotic and turgor pressures. The greater the suction force at a given moment, the less turgor, and vice versa. The shell, straining under the influence of pressure on it from the inside, exerts pressure on the protoplast that is equal in magnitude and opposite in direction.
1 - a cell with a wall layer of protoplasm and a central vacuole; 2 - the beginning of plasmolysis, the cell size decreases slightly; 3 - the vacuole noticeably contracts, protoplasm moves away from the cell membrane in its corners; 4 - the protoplast takes on a spherical shape.
Osmotic phenomena have great importance in plant life. Water is transferred from cell to cell due to the difference in the suction forces of the cells. Experience has established that the suction force of the cells of the upper leaves of a plant is greater than that of the lower leaves, and that the concentration of salts in the root cells (root hairs) is greater than in the soil. The osmotic pressure of root hairs is 1.5 or more times higher than in the soil solution. Absolute value The osmotic pressure in the cells is quite high. U land plants it most often fluctuates from 5 to 10 atmospheres. In freshwater plants it is lower - only 1-3 atmospheres. In plants of saline soils it reaches 40, 80 and even 100 or more atmospheres. The sucking power of dry seeds is measured in hundreds of atmospheres.
Plasmolysis. When a living cell is exposed to hypertonic solutions, i.e., solutions with a suction force exceeding the suction force of cell sap, the protoplast departs from the cell walls as a result of the loss of part of the water by the vacuoles and protoplasm. This phenomenon is called plasmolysis. By placing a preparation (for example, a leaf of an aquatic plant Elodea, a piece of the skin of a juicy onion scale) in water on a glass slide, covering it with a cover glass, applying on the side a drop of a hypertonic solution that does not have a toxic effect on the protoplast (for example, a sugar solution or a 10% solution potassium nitrate), and suctioning the solution from the opposite side with pieces of filter paper, you can notice that soon the protoplasts, decreasing in volume, move away from the cell walls (Fig. 40). By placing cells in such a (hypertonic) solution, you can be sure that its concentration greatly exceeds the concentration of cell sap. Thanks to osmotic phenomena, water from the vacuoles will begin to pass through the protoplasm into the surrounding solution. The vacuoles will decrease in size. Due to the cohesion of liquid molecules and the elasticity of living protoplasm, it will follow the vacuole.
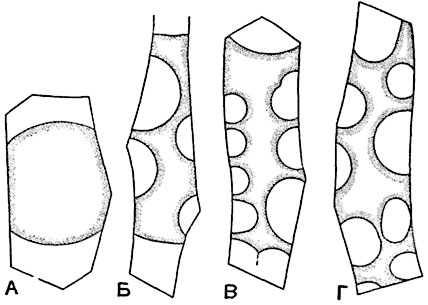
A- convex plasmolysis; B- concave plasmolysis; V, G- convulsive plasmolysis.
Following the contracting vacuole, the protoplasm will lag behind the cell walls, first in separate places, and then completely, and eventually in each cell there will be a round or oval (because it is slightly compressed by the side walls of the cell) protoplast with a very small vacuole inside. The space between the protoplast and the cell walls will be occupied by the solution that caused plasmolysis (plasmolytic).
Plasmolysis is observed especially clearly if you take an object with colored cell sap, for example, a cut from the root of a red beet, the skin from the leaves or stems of begonias that have a reddish color, from the scales of an onion of some variety that has strongly colored scales. Typically, the protoplast leaves the membrane primarily in the corners of the cell. Often in a cell there are certain areas of the inner surface of the shell, from which the protoplasm quickly retreats during plasmolysis, and other areas from which the protoplasm separates later or does not separate at all. Areas of the first kind are called positive areas, areas of the second type are called negative sites of plasmolysis. In wounded plant organs, in each of the cells located near the wounded area, the wall that is closest to the wound becomes a negative site of plasmolysis. After plasmolysis, the protoplast may have smooth convex outlines: so-called convex plasmolysis occurs (Fig. 41, A).
In highly elongated cells, the protoplast, after plasmolysis, breaks up into two or several rounded parts, often connected by thin plasma strands. With a high viscosity of the protoplasm and its strong adhesion to the cell walls, concave plasmolysis occurs: the protoplast in some areas of the surface remains in contact with the cell membrane, in other places it moves away from it, being turned with the concave side of the curved surface towards the walls (Fig. 41, B). Concave plasmolysis in a pronounced form is the so-called convulsive or spasmodic plasmolysis (Fig. 41, V, G).
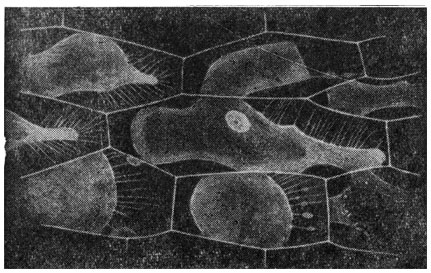
During plasmolysis, the protoplast in some cells remains connected to the walls by thin plasma filaments - Hecht filaments (Fig. 42). Some of these threads are broken in the further course of plasmolysis.
The ability to plasmolyze depends largely on the age stage of the cell: in young growing and developing cells, the protoplasm is more viscous and strongly adheres to the membrane; later, its viscosity and the adhesion of the plasmalemma to the cell membrane decrease, which manifests itself during plasmolysis.
Plasmolysis can also occur “autonomously” (on its own) in certain plant cells; As an example, one can cite the reduction in volume, rounding, and departure of the protoplast from the shell during the formation of a zoospore or egg in the green alga Oedogonia. Phenomena of this order usually occur in sporangia (receptacles for asexual reproductive cells - spores) and gametangia (receptacles for germ cells - gametes).
Protoplasm is not completely impenetrable to many of the substances soluble in water. Although slowly, much slower than water, these substances still penetrate through the protoplasm into the vacuoles, therefore, as a result of long-term interaction between the cell and the plasmolytic (a solution that causes plasmolysis), the sucking force of the cell sap and the plasmolytic can be equalized. The latter ceases to be hypertonic, and the protoplast, again sucking in water, restores its previous volume and previous outlines and comes into contact with the cell membrane with its entire outer surface. The restoration of the protoplast to its previous state, disturbed by plasmolysis, is called deplasmolysis. Deplasmolysis occurs much faster, and sometimes more completely, if plasmolyzed cells are placed in a solution of weaker concentration or in water. In experiments with artificial cells, this pressure can be measured with a mercury manometer.
In experiments of this kind, water from a tap or from a pond is suitable. In ordinary distilled water, cells quickly die: the traces of metals it contains, especially copper, and the pH value unfavorable for protoplasm act as a poison.
Through a series of systematic tests, the concentration (isosmotic) of the plasmolytic is found, which barely causes the first signs of plasmolysis. Having found the osmotic pressure of this plasmolytic concentration from the tables, one can almost accurately indicate the osmotic pressure of the cell sap in the cells under study.
Most mature plant cells are characterized by a large central vacuole, occupying up to 70-90% of the cell volume. In this case, the protoplast with all the organelles is located in the form of a very thin wall layer lining the cell wall. Small cytoplasmic vacuoles are usually found in the wall protoplast. Sometimes the nucleus is located in the center of the cell in nuclear pocket cytoplasm, which is connected to the wall layer by the thinnest cytoplasmic strands crossing the central vacuole.
Cell sap is an aqueous solution of various substances that are products of the vital activity of the protoplast, mainly reserve substances and waste. The reaction of cell sap is usually slightly acidic or neutral, less often alkaline. The substances that make up the cell sap are extremely diverse. These are carbohydrates, proteins, organic acids and their salts, amino acids, mineral ions, alkaloids, glycosides, tannins, pigments and other water-soluble compounds. Most of them belong to the group of ergastic substances - products of protoplast metabolism, which can appear and disappear at different periods of cell life. Many cell sap substances are formed only in plant cells.
Carbohydrates in plant cell sap are represented by monosaccharides (glucose, fructose), disaccharides (sucrose) and polysaccharides (mucus, inulin).
Glucose (grape sugar) and fructose (fruit sugar) accumulate in large quantities in juicy fruits. Sucrose (beet sugar) accumulates in large quantities in sugar beet roots and sugar cane stalks. A number of plant families (cactaceae, Crassulaceae, orchids) are characterized by the accumulation of mucus in the cell sap, which retains water. Inulin is a reserve polysaccharide, deposited as a colloidal solution in the cell sap of the underground organs of Asteraceae instead of starch.
Proteins accumulate in the form of a colloidal solution in the vacuoles of cells of ripening seeds. When seeds are dehydrated in the later stages of their development, water is removed from the vacuoles, the protein concentration in the cell sap increases, and it turns into a solid gel state. Dehydrated vacuoles of mature seeds are called aleurone grains.
Of the organic acids in cell sap, the most common are citric, malic, succinic and oxalic. These acids are found in large quantities in the cell sap of unripe fruits, giving them a sour taste. When fruits ripen, organic acids can be used as respiration substrates, so the sour taste of the fruit usually disappears. Salts of organic acids, together with mineral ions, play an important role in osmotic processes.
Tannins (tannins) are polymeric phenolic compounds with an astringent taste. They have antiseptic properties and protect plant tissues from infections and decay. The cells of the bark of stems and roots (oak, willow), unripe fruits (walnuts), leaves (tea) and some pathological growths - galls are especially rich in tannins. Tannids are used in medicine, for tanning leather, and dyeing fabrics dark brown.
Alkaloids are chemically diverse nitrogen-containing organic substances that have a bitter taste. They have the properties of bases and are found in cell sap, usually in the form of salts. Many alkaloid-bearing plants are poisonous and are not eaten by herbivores. In cells containing alkaloids, spores and germs of microorganisms do not develop, and plants are not affected by fungal and bacterial diseases. Representatives of the families Solanaceae, Poppy, Rubiaceae, Ranunculaceae, etc. are especially rich in alkaloids.
Glycosides are a large group of natural substances, compounds of sugars with alcohols, aldehydes, phenols and other substances. A number of plant glycosides are used in medicine. Glycosides also include cell sap pigments - flavonoids. One of them - anthocyanins– give cell sap a red, blue or purple color; other - flavones- yellow. Anthocyanins are associated with the color of flowers of many plants. The color scheme is determined by the reaction of the cell sap: if it is acidic, then red tones predominate, if it is neutral, purple tones predominate, and with a slightly alkaline reaction, blue tones predominate. The appearance of shades is also influenced by the formation of anthocyanin complexes with various metals. Flavones are responsible for the yellow color of the petals of a number of plants.
The importance of organic acids, tannins, alkaloids and glycosides of cell sap in cell metabolism has not been sufficiently elucidated. Previously, they were considered as final products of exchange. It has now been shown that many of them can be re-involved in metabolic processes and therefore can be considered as reserve substances.
In addition to the function of accumulating reserve substances and waste, vacuoles in plant cells perform another important function - maintaining turgor. The concentration of ions and sugars in the cell sap of the central vacuole is usually higher than in cell wall; The tonoplast significantly slows down the diffusion of these substances from the vacuole and at the same time is easily permeable to water. Therefore, water will enter the vacuole. This unidirectional process of water diffusion through a selectively permeable membrane is called osmosis. Water entering the cell sap exerts pressure on the wall protoplast, and through it on the cell wall, causing its tense, elastic state, or turgor cells. Turgor ensures that non-lignified plant organs retain their shape and position in space, as well as their resistance to mechanical factors.
If a cell is placed in a hypertonic solution of some non-toxic salt or sugar (i.e., in a solution of a higher concentration than the concentration of cell sap), then an osmotic release of water from the vacuole occurs. As a result of this, its volume is reduced, the elastic wall protoplast moves away from the cell wall, turgor disappears, and plasmolysis cells( Fig.2.9).
Rice. 2.9. Plasmolysis scheme: 1 – cell in a state of turgor; 2 – beginning of plasmolysis; 3 – complete plasmolysis.
Plasmolysis is usually reversible. When a cell is placed in water or a hypotonic solution, water is again vigorously absorbed by the central vacuole, the protoplast is again pressed against the cell wall, and turgor is restored. Plasmolysis can serve as an indicator of the living state of a cell; a dead cell is not plasmolyzed, since it does not have selectively permeable membranes.
Loss of turgor causes the plant to wilt. When withering in air under conditions of insufficient water supply, the thin cell walls shrink simultaneously with the protoplast and become folded.
Turgor pressure not only maintains the shape of non-lignified plant parts, it is also one of the cell growth factors, providing height cells stretching, i.e. due to the absorption of water and an increase in the size of the vacuole. Animal cells do not have a central vacuole; their growth occurs mainly due to an increase in the amount of cytoplasm, therefore the size of animal cells is usually smaller than plant cells.
The central vacuole arises from the fusion of numerous small vacuoles that are present in meristematic (embryonic) cells. These cytoplasmic vacuoles are believed to be formed by the membranes of the endoplasmic reticulum or Golgi apparatus.