Sviluppo individuale(ontogenesi), periodizzazione dell'ontogenesi.
Tutti gli organismi hanno un ciclo di vita specifico. Per gli organismi che si sviluppano sessualmente, inizia dal momento in cui appare lo zigote e termina con la morte naturale dell'organismo.
La totalità dei processi che si verificano durante il ciclo di vita degli organismi è definita come sviluppo individuale o ontogenesi.
L'ontogenesi comprende 3 periodi:
1 periodo. Preembrionale o gametogenesi.
2 periodo. Embrionale.
3 periodo. Postembrionale.
1. Cicli vitali degli organismi. Lo sviluppo è larvale e diretto.
Lo sviluppo degli organismi è diretto e indiretto con la trasformazione.
sviluppo indiretto avviene attraverso lo stadio larvale. Nella larva si formano alcuni organi germinali o provvisori che forniscono l'attività vitale dell'organismo in questa fase di sviluppo.
Nei vertebrati superiori lo sviluppo è diretto, ma durante lo sviluppo embrionale si formano anche organi provvisori. Nei mammiferi, queste sono le membrane embrionali (amnion, corion, allantoide, placenta) e il sacco vitellino.
2. Periodo pre-embrionale (pre-embrionale, progenesi) di sviluppo. Fasi della gametogenesi. Cambiamenti nell'oogenesi associati allo sviluppo precoce dell'embrione (amplificazione genica, segregazione ooplasmatica, accumulo di nutrienti).
Il periodo preembrionale o gametogenesi comprende diverse fasi: isolamento, riproduzione, crescita, maturazione, formazione (quest'ultima solo negli spermatozoi).
Durante l'oogenesi si verificano gli eventi più importanti necessari per lo sviluppo del futuro organismo.
1 evento. Durante l'oogenesi si verifica l'amplificazione dei geni dell'rRNA o un aumento del numero di copie dei geni responsabili dell'rRNA. Questo processo si verifica nella profase della meiosi 1. Le copie dei geni dell'rRNA possono essere fino a un milione.
Quindi queste copie vengono separate dai cromosomi, fluttuano liberamente nel carioplasma, i nucleoli si formano attorno a loro e le subunità ribosomiche vengono sintetizzate nei nucleoli, che entrano nel citoplasma. Pertanto, il numero di ribosomi aumenta bruscamente in anticipo nell'uovo.
2 evento. Durante l'oogenesi, vari tipi di mRNA vengono sintetizzati durante la profase della meiosi 1. I processi di trascrizione avvengono su sezioni despiralizzate di cromosomi. I cromosomi allo stadio profase della meiosi 1 sono chiamati cromosomi lampbrush.
3 evento. I nutrienti sono immagazzinati nell'uovo sotto forma di tuorlo.
4 evento. L'uovo è caratterizzato dalla segregazione ooplasmatica, cioè dalla distribuzione di sostanze in tutto il citoplasma dell'uovo, che porta all'eterogeneità chimica del citoplasma. Si ritiene che ciò sia necessario per la differenziazione cellulare precoce.
5 evento. Le cellule sessuali sono cellule speciali del corpo, poiché hanno totipotenza, cioè pari ereditarietà. Solo le cellule germinali, così come i blastomeri nell'uomo, allo stadio di 2 blastomeri danno origine a tutti i tipi di cellule. Ad esempio, esperimenti di separazione, splicing o mescolamento di blastomeri allo stadio di clivaggio hanno dimostrato che in specie con clivaggio di tipo radiale, blastomeri di più generazioni, se isolati e posti in condizioni idonee, presentano totipotenza, cioè trasformarsi in un organismo completo. Per uguale ereditarietà e totipotenza delle cellule di embrioni umani fino allo stadio di 2-4 blastomeri, parlano i casi di nascita di due, tre, quattro gemelli identici.
3. Periodo di sviluppo embrionale, periodizzazione.
Il periodo embrionale dell'ontogenesi comprende diverse fasi:
1 fase di fecondazione.
2° stadio dello zigote.
3 Fase di frantumazione (formazione di un embrione monostrato).
4 Fase di gastrulazione (formazione di embrioni a due e tre strati).
5 Fase histo- e DI rganogenesis (la formazione di tessuti e organi).
4. Fecondazione e formazione dello zigote. Caratteristiche della fecondazione nei mammiferi e nell'uomo.
fase di fecondazione, questo è il processo di fusione dell'uovo e dello sperma, di conseguenza si forma uno zigote diploide, dal quale si sviluppa un organismo diploide. Ci sono 3 fasi in questo processo:
1 stadio- convergenza dei gameti. In questo, le sostanze secrete dall'uovo e dallo sperma giocano un ruolo importante. Sono chiamati gamoni (ormoni di gameti, ginogamoni e androgamoni, rispettivamente). Inoltre, ci sono una serie di fattori non specifici che aumentano la probabilità di incontro e interazione dello spermatozoo con l'uovo. Questi includono
coordinamento dell'inizio della prontezza alla fecondazione nel maschio e nella femmina,
il comportamento di maschi e femmine che assicura la copulazione e l'inseminazione,
eccesso di produzione di spermatozoi
grande ovulo,
la presenza di gamoni che contribuiscono alla convergenza e all'interazione dei gameti,
la presenza di organi copulatori che forniscono l'inseminazione interna.
Nei mammiferi Grande importanza ha una permanenza di spermatozoi nel tratto genitale della femmina, a seguito della quale le cellule germinali maschili acquisiscono una capacità fecondante, cioè capacità di reazione acrosomiale.
Fase 2 - l'attivazione dei gameti avviene dopo il loro contatto. L'attivazione dello spermatozoo è chiamata reazione acrosomiale. L'attivazione dell'uovo è una reazione corticale.
L'essenza della reazione acrosomiale : La permeabilità dello sperma nell'acrosoma cambia il ponte della membrana plasmatica e gli enzimi, le spermatolisine, vengono rilasciati dall'acrosoma. Questi enzimi rilassano i legami tra le cellule follicolari che circondano l'uovo. Lo spermatozoo passa attraverso lo strato di cellule follicolari, quindi la zona del pellucido viene distrutta e lo spermatozoo passa attraverso questa zona.
L'essenza della reazione corticale: Consiste in complessi cambiamenti strutturali e fisico-chimici. A causa del fatto che la membrana dello spermatozoo è permeabile agli ioni sodio, questi ultimi iniziano a entrare nell'uovo, cambiando potenziale di membrana cellule. Quindi, sotto forma di un'onda che si propaga dal punto di contatto dei gameti, si verifica un aumento del contenuto di ioni Ca 2+ (nell'ialoplasma lasciano il deposito - EPS, reticolo) e nell'uovo si innescano processi biochimici, dopodiché, anche i granuli corticali si dissolvono in un'onda. Gli enzimi specifici rilasciati contemporaneamente contribuiscono al distacco della membrana del tuorlo; lei lo indurisce guaina di fertilizzazione.
Uno dei significati della reazione corticale è la prevenzione della polispermia, cioè penetrazione di più di uno spermatozoo nell'ovulo. Nei mammiferi la reazione corticale non provoca la formazione della membrana di fecondazione, ma la sua essenza è la stessa.
L'attivazione dell'uovo termina con l'inizio della sintesi proteica a livello traduzionale, poiché mRNA, tRNA, ribosomi ed energia sono stati immagazzinati anche nell'oogenesi.
3 fasi- fusione di gameti, o sinagoga. In questo caso, si forma una membrana plasmatica comune nello sperma e nell'uovo. Il pronucleo femminile e quello maschile convergono e si fondono (syncaryon), formando una placca metafasica comune. Questo è il momento della fusione finale dei gameti: la sinagoga.
Caratteristiche della fecondazione in varie specie di organismi.
1 esempio. Nei mammiferi e nell'uomo, lo spermatozoo si lega all'uovo nell'area in cui è presente un recettore sulla zona pellucida. Dopo questa interazione, i restanti recettori vengono bloccati.
2 esempio. A riccio di mare dopo la fecondazione nell'uovo cambia radicalmente potenziale elettrico la membrana plasmatica, e quindi si forma una membrana di fertilizzazione che previene la polispermia.
stadio zigotico. Dopo la penetrazione, il nucleo maschile è chiamato pronucleo maschile. La cromatina si allenta in esso, si verifica la replicazione del DNA. Il nucleo femminile è chiamato pronucleo femminile. Gli stessi eventi si svolgono in esso. Nei mammiferi e nell'uomo non si verifica la fusione nucleare, ma si forma immediatamente una piastra metafasica.
5. Inseminazione artificiale dell'uovo di animali e umani, aspetti biologici e medici.
La fecondazione artificiale delle uova degli animali è di grande importanza scientifica per la medicina, poiché nel processo del suo studio si stanno sviluppando modi e meccanismi per trattare l'infertilità nell'uomo.
L'inseminazione artificiale è usata per varie forme infertilità, sia maschile che femminile, che è difficile da trattare. Ad esempio, quando un uomo ha troppo pochi spermatozoi o sono praticamente immobili, quando una donna ha una ridotta pervietà tube di Falloppio o ci sono altri danni agli organi genitali interni, con incompatibilità immunologica dei partner.
6. caratteristiche generali schiacciamento. Tipi di schiacciamento caratteristici di varie specie animali. Scissione e formazione della blastula nei mammiferi placentari.
fase di frantumazione. Questo è lo stadio di formazione di un embrione a strato singolo: la blastula. All'interno della blastula c'è una cavità chiamata blastocele.
Caratteristiche di schiacciamento:
Le cellule si dividono per mitosi.
Alla vigilia di ogni divisione, si verifica la replicazione del DNA.
Le cellule in divisione non crescono.
Il tipo di frantumazione dipende dal tipo di uovo.
Frantumazione uniforme completa nella lancetta:
Il primo solco di scissione corre verticalmente, si formano due blastomeri. Anche il secondo solco corre verticalmente e si formano quattro blastomeri. Il terzo solco corre orizzontalmente, si formano otto blastomeri, quindi si alternano solchi verticali e orizzontali. Dopo 12 cicli la frantumazione diventa asincrona. Ad un certo stadio di sviluppo, l'embrione è un grumo di cellule o una morula. Quindi compaiono degli spazi tra le cellule e si forma una cavità: il blastocele. Nella lancetta, durante la frantumazione, si forma una blastula, chiamata coeloblastula, cioè una palla a strato singolo.
Frammentazione irregolare completa negli anfibi:
Le cellule anfibie sono moderatamente telolecitali. Al polo animale della cellula, la frammentazione procede più velocemente che al polo vegetativo. Di conseguenza, le cellule del polo animale sono più piccole: i micromeri. Al polo vegetativo, le cellule sono più grandi: i macromeri. La blastula anfibia è chiamata anfiblastula. Il blastocele si trova al polo animale.
Caratteristiche dello schiacciamento nei mammiferi e nell'uomo:
La scissione è completamente irregolare, asincrona fin dai primi stadi; a un certo stadio di sviluppo, l'embrione è una morula (grumo di cellule). Quindi, le cellule più grandi si separano verso la periferia, formando un trofoblasto, e le cellule più piccole verso il centro, formando un embrioblasto. La blastula è chiamata blastocisti. Il blastocele è molto piccolo. Il trofoblasto favorisce l'introduzione dell'embrione nella mucosa uterina. Questo processo è chiamato impianto. L'embrioblasto dà origine all'embrione stesso e ad alcuni organi provvisori.
7. Caratteristiche generali della gastrulazione. Caratteristiche della gastrulazione negli anfibi e negli uccelli. Gastrulazione nei mammiferi superiori (placentari).
Fase di gastrulazione, o lo stadio di formazione di un embrione a due strati, e poi uno a tre strati. L'embrione in questa fase è chiamato gastrula.
Metodi per la formazione di un embrione a due strati:
- Intussuscezione (invaginazione).
– Delaminazione (stratificazione).
– Immigrazione (insediamento).
- Epiboly (incrostazione).
Intussuscezione o invaginazione. Questo metodo è tipico per la lancetta. In una certa area, le cellule della blastula si invaginano nel blastocele, dando luogo alla formazione di un embrione a due strati. Lo strato esterno delle cellule è chiamato ectoderma, lo strato interno è chiamato endoderma. L'endoderma limita la cavità dell'intestino primario o gastrocele. L'ingresso a questa cavità è chiamato bocca primaria o blastopore. Il blastoporo è circondato da labbra.
Delaminazione o stratificazione. Questo metodo è tipico degli animali intestinali, in cui la blastula sembra una morula e il blastocele non è praticamente espresso.
immigrazione o insediamento. Alcune cellule di blastula vengono introdotte nel blastocele, quindi queste cellule si dividono intensamente. Di conseguenza, a causa di queste cellule si forma l'endoderma.
Epiboly (incrostazione). I micromeri sono divisi e, per così dire, stratificati sui macromeri. I micromeri formano l'ectoderma, mentre i macromeri formano l'endoderma. Nella sua forma pura, questi metodi non si trovano praticamente, di norma sono combinati. Gli anfibi combinano invaginazione ed epibolia. Uccelli e mammiferi combinano delaminazione e immigrazione.
A partire dai platelminti, il terzo strato germinale, il mesoderma, appare in evoluzione.
Metodi di formazione del mesoderma:
Il metodo teloblastico è caratteristico dei protostomi. Nella regione delle labbra del blastopore vengono secrete 2 cellule che si dividono e formano il mesoderma.
Il metodo enterocoel è caratteristico dei deuterostomi (cordati). 2 sezioni di cellule sotto forma di tasche sono separate simmetricamente dall'endoderma. Queste sono tasche mesodermiche. Le cellule delle tasche mesodermiche si dividono e danno origine al mesoderma. Il mesoderma è lo strato germinale.
8. Caratteristiche generali dell'isto- e dell'organogenesi (formazione di tessuti e organi).
Stadio dell'isto e dell'organogenesi(fase di formazione di tessuti e organi). È convenzionalmente diviso in due periodi.
1 periodo. Periodo di istruzione organi assiali nell'embrione, questa è la formazione del tubo neurale e della corda. Pertanto, questo periodo è chiamato periodo di neurulazione e l'embrione in questa fase è chiamato neurula.
2 periodo. È caratterizzato dalla formazione di altri tessuti e organi. Sul lato dorsale dell'embrione (dorsale), per tutta la sua lunghezza, una sezione di cellule si separa dall'ectoderma, che dà origine alla placca neurale. Quindi i bordi della placca neurale si alzano, si ispessiscono e si forma un solco neurale, che gradualmente affonda sotto l'ectoderma. Quindi i bordi del solco neurale si chiudono, formando un tubo neurale con una cavità all'interno, la cavità è chiamata neurocele. Nei vertebrati, la parte anteriore del tubo neurale si espande e dà origine al cervello, il resto al midollo spinale. Allo stesso tempo, una notocorda viene posta sotto il tubo neurale, è formata dall'endoderma e dal mesoderma adiacente. Inizialmente, il mesoderma è una massa cellulare omogenea, ma man mano che si sviluppa, si verifica la sua segmentazione. Si formano strutture chiamate somiti. Successivamente, danno origine al sistema muscolo-scheletrico.
Derivati degli strati germinali:
Ectoderma - smalto dei denti, sistema nervoso e organi sensoriali, epidermide della pelle e sue appendici, epitelio dell'intestino anteriore e posteriore.
Endoderma - epitelio dell'intestino medio, ghiandole digestive e apparato respiratorio.
Mesoderma - sistema muscolo-scheletrico, sistema genito-urinario, sistema circolatorio e linfatico, tutto il tessuto connettivo.
9. Caratteristiche degli organi provvisori di embrioni di vertebrati. Organi provvisori dei mammiferi superiori.
Gli organi provvisori funzionano nell'embrione e sono assenti nello stato adulto. Questi includono il sacco vitellino e le cosiddette membrane embrionali: amnion, corion e allantois.
Sacco vitellino. Il sacco vitellino svolge una serie di importanti funzioni: nutrizione, respirazione, escrezione, emopoiesi. Ma, a causa del basso contenuto di tuorlo nell'uovo, non svolge un ruolo significativo nella nutrizione dell'embrione.
Amnio. La membrana amniotica partecipa alla formazione dell'amnio, che limita la cavità amniotica piena di liquido amniotico, che ora circonda l'embrione da tutti i lati. A causa di ciò, l'embrione si sviluppa in un ambiente acquatico, che lo protegge dagli effetti traumatici meccanici e dall'adesione ai gusci.
Allantois formata come una conseguenza del hindgut. La funzione principale dell'allantoide è quella di essere l'organo germinale dell'escrezione. Accumula prodotti di decadimento che si formano durante il metabolismo nel corpo dell'embrione.
L'embriogenesi è un complesso processo olistico associato a determinati fenomeni e meccanismi. Molti di questi fenomeni non sono completamente compresi, sebbene per alcuni di essi siano stati ottenuti dati certi.
1. Cambiamenti genetici molecolari nelle prime fasi dello sviluppo.
2. Proliferazione cellulare (divisione cellulare).
3. Differenziazione cellulare.
4. Formazione o morfogenesi.
1. Cambiamenti genetici molecolari nello sviluppo iniziale (il periodo dello zigote e della scissione), il ruolo dei fattori citoplasmatici nell'uovo.
Lo sviluppo iniziale include le fasi dello zigote e della scissione.
Studiando queste fasi, gli scienziati hanno cercato di rispondere alle domande:
Primo, quando i geni dell'embrione iniziano a funzionare.
In secondo luogo, ci sono differenze qualitative e quantitative nell'mRNA e nelle molecole proteiche parti differenti embrione nelle prime fasi di sviluppo.
Nello zigote, l'attività genica è bassa, poiché il DNA è fortemente associato alle proteine istoniche. Le prime proteine che vengono sintetizzate nello zigote sono di origine materna, poiché i ribosomi e le molecole di mRNA si sono accumulate in anticipo nell'uovo. È stato stabilito che i geni propri dell'embrione nei mammiferi iniziano a funzionare allo stadio di 2-4 blastomeri. Negli anfibi è allo stadio di blastula. I primi a lavorare sono i geni responsabili della proliferazione e del metabolismo generale, successivamente iniziano a lavorare i geni responsabili della differenziazione delle cellule e dei tessuti. Ad esempio, quando il nucleo viene rimosso dallo zigote, si verifica la scissione e l'embrione nel suo sviluppo raggiunge quasi lo stadio di blastula, dopodiché l'ulteriore sviluppo si interrompe.
È stato stabilito che non ci sono differenze qualitative nell'mRNA e nelle molecole proteiche nelle diverse parti dell'embrione nelle prime fasi dello sviluppo. Ci sono solo differenze quantitative.
Un ruolo importante nella frantumazione è svolto dalla divisione del citoplasma - citotomia. Ha un particolare significato morfogenetico, poiché determina il tipo di frantumazione. I solchi di scissione corrono lungo i confini tra le singole sezioni dell'ooplasma, riflettendo il fenomeno della segregazione ooplasmatica. Pertanto, il citoplasma di diversi blastomeri differisce nella composizione chimica.
2. Proliferazione cellulare, crescita.
La proliferazione cellulare o la divisione cellulare avviene durante l'embriogenesi. Associato a questo è la crescita di tessuti e organi. La crescita dell'embrione nel suo insieme.
3. Differenziamento, meccanismi genetici molecolari del differenziamento.
La differenziazione cellulare è un insieme di processi, a seguito dei quali le cellule di origine comune acquisiscono differenze morfologiche, fisiologiche, biochimiche persistenti, che portano alla specializzazione cellulare. La specificità delle cellule è determinata dalle proteine che sono sintetizzate in esse e i geni corrispondenti sono responsabili delle proteine. Pertanto, possiamo concludere che alcuni geni funzionano in alcune cellule e altri in altre. Questa è l'essenza dell'ipotesi dell'attività differenziale dei geni.
Nelle fasi iniziali, la differenziazione cellulare è associata all'influenza delle sostanze citoplasmatiche sul lavoro dei geni corrispondenti: questo è il livello epigenetico di regolazione del lavoro dei geni. Il fenomeno della segregazione ooplasmatica avviene nell'uovo, di conseguenza, diverse parti del citoplasma dell'uovo contengono sostanze diverse. Durante la frantumazione compaiono i blastomeri, l'insieme di geni in essi contenuti è lo stesso e la composizione del citoplasma è diversa. Successivamente, queste sostanze del citoplasma, apparentemente, portano all'attività differenziale dei geni.
Quando si caratterizza la differenziazione cellulare, vengono utilizzati due concetti: determinazione e competenza.
determinazione significa che la differenziazione cellulare è geneticamente predeterminata e irreversibile.
Nel processo di differenziazione, il materiale cellulare degli anlages embrionali si trasforma in un certo elemento dell'organismo adulto. Consideriamo la differenziazione usando l'esempio del somite mesodermico, che è suddiviso in dermatomo, sclerotomo e miotomo. Cellule dermatome del derma, le seconde cellule cartilaginee, le terze fibre muscolari striate. Di conseguenza, il risultato finale dello sviluppo dei singoli anlage embrionali è predeterminato o determinato.
Competenza Questa è la capacità delle cellule di differenziarsi in direzioni diverse, sotto l'influenza di fattori ambientali. Ad esempio, la notocorda e il mesoderma adiacente agiscono sull'ectoderma, determinando la formazione del tubo neurale dall'ectoderma. Se non c'è tale effetto, allora l'ectoderma dà origine all'epidermide della pelle.
4. Morfogenesi (formatura), i suoi principali processi:
Formazione o morfogenesi. La morfogenesi è un insieme di processi, a seguito dei quali l'embrione acquisisce una caratteristica esterna e struttura interna. A sua volta, la morfogenesi è associata a:
a) movimento morfogenetico delle cellule
Durante l'embriogenesi, singole cellule o gruppi di cellule si muovono. Le cellule si muovono sulla superficie di altre cellule (a causa del meccanismo del movimento ameboide), dove sono presenti molecole speciali che indicano la direzione del movimento. Alcuni tipi di cellule si muovono lungo un gradiente di concentrazione chimica (chemiotassi), ma questo meccanismo è molto meno comune.
La violazione della migrazione cellulare durante l'embriogenesi porta al sottosviluppo degli organi oa un cambiamento nella sua normale localizzazione. Entrambe sono malformazioni congenite. Ad esempio, in violazione della migrazione delle cellule - i neuroblasti, nella sostanza bianca compaiono isole di materia grigia e allo stesso tempo le cellule perdono la capacità di differenziarsi.
Pertanto, la migrazione cellulare è sotto il controllo genetico, da un lato, e l'influenza delle cellule e dei tessuti circostanti, dall'altro.
b) induzione embrionale
Questo è l'impatto di un tessuto (induttore) su un altro tessuto, di conseguenza, lo sviluppo del tessuto indotto diventa qualitativamente nuovo. La prima e più significativa induzione è l'azione della notocorda e del mesoderma sull'ectoderma, con conseguente formazione del tubo neurale. Senza il tubo neurale, l'intero ectoderma si trasformerà nell'epidermide. Questa è l'induzione embrionale primaria, il primo passo in una catena di processi di induzione successivi (secondari, terziari) in ulteriore sviluppo.
È stato stabilito che esistono "induttori specifici", ad es. sostanze che esercitano un effetto inducente in concentrazioni trascurabili e si differenziano per il risultato finale della loro azione. Pertanto, un estratto dal fegato dei mammiferi induce principalmente lo sviluppo delle strutture cerebrali e un estratto del midollo osseo - quelle mesodermiche.
La capacità del germe embrionale di percepire lo stimolo induttivo si chiama competenza.
c) interazioni intercellulari
Questa è l'interazione di cellule o strati in contatto oa distanza. L'interazione a distanza va con la partecipazione di sostanze biologicamente attive (BAS).
Questi possono essere proteine, ormoni, ecc. Nelle prime fasi dello sviluppo embrionale, questi sono gli ormoni della madre, poiché l'embrione non ha le proprie ghiandole endocrine. Gli ormoni non provocano una nuova differenziazione, ma la rafforzano.
A causa delle interazioni intercellulari, vengono eseguiti fenomeni come il movimento cellulare morfogenetico, l'induzione embrionale e l'adesione cellulare.
d) adesione- la capacità delle cellule di restare unite. Nell'esperimento, le cellule dell'ectoderma, del mesoderma e dell'endoderma sono state separate e mescolate tra loro. Quindi si riuniscono nuovamente in gruppi separati, ciascuno dei quali è un aggregato cellulare di cellule omogenee. Si formano nuovamente tre strati germinali, posizionati normalmente l'uno rispetto all'altro.
Speciali molecole proteiche prendono parte al processo di adesione. Sono chiamate molecole di adesione cellulare (MAC) e ne esistono circa 100 tipi.
Un'altra ipotesi afferma che i contatti tra cellule simili sono più forti che tra cellule estranee.
L'adesione selettiva delle cellule di un certo strato germinale tra loro è una condizione necessaria per il normale sviluppo.
e) la morte cellulare è un processo necessario, perché la formazione di singole strutture (condotti, canali, fori, ecc.) richiede la distruzione di parte delle cellule.
Esistono due tipi fondamentalmente diversi di morte cellulare: apoptosi(tradotto dal greco "cadere") e necrosi.
L'apoptosi è una morte cellulare fisiologica, geneticamente predeterminata. Insieme ad altri meccanismi di morfogenesi, contribuisce al raggiungimento delle caratteristiche della sua organizzazione morfofunzionale caratteristica di una certa specie biologica. Pertanto, l'apoptosi è un meccanismo di morfogenesi naturale, evolutivamente determinato e geneticamente controllato.
Necrosi - morte cellulare non fisiologica, dovuta all'influenza di fattori avversi (meccanici, chimici, fisici, ecc.). La necrosi è solitamente accompagnata da infiammazione ed è un processo patologico.
5. Integrazione nello sviluppo, integrità dell'ontogenesi. Il ruolo degli ormoni nel coordinamento dei processi di sviluppo.
Attualmente sono note numerose sostanze che inducono le cellule a dividersi, ad esempio la fitoemoagglutinina, alcuni ormoni, nonché un complesso di sostanze rilasciate durante il danno tissutale. Sono stati scoperti anche inibitori tessuto-specifici della divisione cellulare: keylon. La loro azione è sopprimere o rallentare il tasso di divisione cellulare nei tessuti che li producono. Ad esempio, i kalon epidermici agiscono solo sull'epidermide. Essendo tessuto-specifici, i kalon mancano di specificità di specie. Pertanto, il kalon epidermico di merluzzo agisce anche sull'epidermide di un mammifero.
Gli ormoni sono composti organici prodotti da alcune cellule e progettati per controllare le funzioni del corpo, la loro regolazione e coordinazione.
L'azione fisiologica degli ormoni è finalizzata a:
1) fornendo umorale, cioè effettuato attraverso il sangue, la regolazione dei processi biologici;
2) mantenimento dell'integrità e della costanza dell'ambiente interno, interazione armoniosa tra i componenti cellulari del corpo;
3) regolazione dei processi di crescita, maturazione e riproduzione. Gli ormoni regolano l'attività di tutte le cellule del corpo. Influiscono sulla nitidezza del pensiero e sulla mobilità fisica, sul fisico e sulla crescita, determinano lo sviluppo di segni di dimorfismo sessuale e comportamento.
6. Il ruolo dell'ereditarietà e dell'ambiente nello sviluppo embrionale. Periodi critici dello sviluppo. fattori teratogeni. Anomalie e malformazioni.
In ogni stadio dell'ontogenesi, l'organismo esiste in unità con ambiente. L'embriogenesi non fa eccezione in questo senso. La gamma di condizioni necessarie per la vita di una specie può essere ampia. Tuttavia, per gli organismi di qualsiasi specie esiste un minimo, un ottimo e un massimo condizioni necessarie sviluppo. Lo sviluppo dell'embrione è influenzato dalle fluttuazioni di fattori naturali (temperatura, umidità, pressione atmosferica, radiazioni, composizione dei gas dell'ambiente).
Quindi, a seconda della temperatura, i processi di sviluppo rallentano o si intensificano. Ad esempio, le uova di rana della stessa covata si sviluppano più velocemente a temperature più elevate.
Nel nematode, quando l'accesso all'ossigeno all'embrione è terminato, lo sviluppo si interrompe.
Regola generaleè che sotto l'azione della luce dalla parte blu-viola dello spettro, lo sviluppo embrionale di molte specie animali viene accelerato e dal rosso rallenta.
Durante lo sviluppo intrauterino, i fattori ambientali sono di grande importanza. Se questi fattori portano alla formazione di anomalie o difetti dello sviluppo, vengono definiti teratogeni. I fattori teratogeni possono essere fisici (alta temperatura, Radiazione ionizzante, raggi X, ecc.), chimico (farmaci, sali di metalli pesanti, ecc.) e biologico (virus, batteri). I fattori teratogeni portano allo sviluppo di anomalie in determinati periodi dello sviluppo embrionale, che sono chiamati critici. Questi includono:
Il periodo di formazione delle cellule germinali (gametogenesi),
fase di fecondazione,
stadio zigotico,
Impianto dell'embrione nella parete dell'utero
Placentazione,
Il periodo di istogenesi e organogenesi,
Malformazioni.
Aplasia: l'assenza di un organo o parte di esso
Ipoplasia - sottosviluppo di un organo
Ipotrofia: una diminuzione del peso corporeo o degli organi
Ipertrofia: un aumento sproporzionato della massa degli organi
Gigantismo: un aumento della lunghezza del corpo
L'eterotopia è una localizzazione atipica di un gruppo di cellule o di un organo nel corpo.
Eteroplasia - violazione della differenziazione dei tessuti
Stenosi: restringimento di un canale o di un'apertura
Atresia - assenza di un canale o apertura
Persistenza - conservazione delle strutture embrionali
A seconda della causa, le malformazioni congenite si suddividono in:
ereditario causato da un cambiamento nei geni o nei cromosomi nei gameti dei genitori, a seguito del quale lo zigote fin dall'inizio porta una mutazione genica, cromosomica o genomica.
esogeno derivanti dall'influenza di fattori teratogeni: droghe (talidomide), additivi alimentari, virus, veleni industriali, ecc. Questi sono tutti fattori ambientali che, agendo durante l'embriogenesi, interrompono lo sviluppo di tessuti e organi.
Multifattoriale difetti che si sviluppano sotto l'influenza di fattori sia esogeni che genetici.
1. Ontogenesi postnatale, sua periodizzazione.
Lo sviluppo postembrionale (postnatale per una persona) inizia dal momento della nascita e termina con la morte naturale o la morte.
Lo sviluppo postembrionale comprende diversi periodi:
1. Pre-riproduttivo (giovanile).
2. Riproduttivo (periodo di maturità).
3. Post-riproduttivo (vecchiaia).
2. Periodo pre-riproduttivo, sue caratteristiche. La crescita corporea come caratteristica importante del periodo pre-riproduttivo.
Il periodo pre-riproduttivo inizia subito dopo la nascita. In questo momento, i processi di morfogenesi giungono al termine, quei sistemi che non funzionavano nell'embriogenesi (respiratorio, escretore e molti altri) iniziano a funzionare.
Una caratteristica importante del periodo pre-riproduttivo è la crescita dell'organismo. In questo caso si verifica un aumento delle dimensioni del corpo nel suo insieme, le sue dimensioni longitudinali aumentano; aumento delle dimensioni dei tessuti e degli organi.
3. La natura della crescita dell'organismo e delle sue singole parti.
Tre processi principali sono alla base della crescita di un organismo:
1. aumento del numero di celle.
2. aumento delle dimensioni delle cellule (ipertrofia).
3. accumulo di sostanza intercellulare.
Ci sono due opzioni di crescita: limitato E illimitato. La crescita illimitata continua per tutta l'ontogenesi, fino alla morte.
Esistono diversi tipi di crescita:
Ausiliario- crescita che va aumentando la dimensione delle cellule.
proliferativo- crescita procedendo per moltiplicazione cellulare: moltiplicativo E accrezione.
Moltiplicativo la crescita è caratterizzata dal fatto che entrambe le cellule che sono nate dalla divisione della cellula madre entrano nuovamente in divisione. La crescita moltiplicativa è molto efficace e quindi non si verifica quasi mai nella sua forma pura o termina molto rapidamente (ad esempio, nel periodo embrionale).
accrezione la crescita sta nel fatto che dopo ogni divisione successiva, solo una delle cellule figlie si divide nuovamente, mentre l'altra smette di dividersi. In questo caso, il numero di celle cresce linearmente. Tale crescita è tipica degli organi in cui avviene il rinnovamento della composizione cellulare.
Va sottolineato che un aumento delle dimensioni longitudinali del corpo, che si verifica principalmente per la crescita delle ossa tubolari lunghe, è di particolare importanza nel caratterizzare la crescita. IN ossa tubolari al confine della diafisi e dell'epifisi, è isolata una zona di crescita. Ecco le cellule della cartilagine, durante la divisione delle quali l'osso cresce in lunghezza.
L'ossificazione finale di ciascun osso avviene in un determinato momento. Negli uomini, la crescita di solito termina all'età di 18-20 anni, nelle donne - entro 16-18 anni. In questo momento, le ultime zone di crescita scompaiono. È allora che la crescita delle ossa in lunghezza si interrompe.
Va sottolineato che fino a 30 anni una persona può crescere di 3 cm a causa di un aumento delle dimensioni delle vertebre.
L'aumento delle dimensioni lineari di una persona è descritto da una curva a forma di S. Subito dopo la nascita, c'è una maggiore crescita del corpo, quindi diminuisce e accelera bruscamente all'età di 13-14-15 anni. Questo è il cosiddetto scatto di crescita puberale (durante la pubertà). Inoltre, il tasso di crescita rallenta leggermente e all'età di 30-40-45 anni la crescita di una persona rimane costante. Secondo questo schema, ossa, muscoli e molti organi interni(fegato, reni, milza).
Con l'invecchiamento, c'è una leggera diminuzione della crescita.
Alcuni organi hanno un carattere completamente diverso:
Questi organi includono il cervello e il midollo spinale, gli organi linfoidi, gli organi riproduttivi.
Il peso del cervello di un neonato è il 25% del peso cerebrale finale (in età adulta), entro 5 anni - 90%, entro 10 anni - 95%.
La crescita del timo (timo) è l'organo centrale del sistema immunitario. Il peso relativo del timo (rispetto al peso corporeo) raggiunge un massimo a 12 anni di età. Il peso assoluto raggiunge il massimo all'età di 30 anni, quindi si verifica una forte diminuzione del peso del timo.
4. Controllo genetico della crescita. Il ruolo dei sistemi nervoso ed endocrino nella regolazione dei processi di crescita.
L'altezza si riferisce a tratti genetici ereditati come i capelli e il colore della pelle, la forma degli occhi, ecc. Ecco perché i genitori alti di solito hanno figli alti e viceversa. La crescita è un tratto poligenico; diversi geni sono responsabili della sua manifestazione nel fenotipo. I geni esercitano il loro controllo sulla crescita attraverso gli ormoni appropriati. L'ormone più importante è l'ormone della crescita o somatotropina prodotto dalla ghiandola pituitaria.
La somatotropina stimola la formazione di nuove cellule cartilaginee, e in parte la loro ossificazione, promuove la sintesi proteica in strutture cellulari ah e la formazione di nuovi capillari. Una grande quantità di questo ormone viene prodotta durante la notte. La somatotropina di un bambino viene prodotta da 3-4 anni.
La crescita è influenzata anche dagli ormoni tiroidei e dagli ormoni sessuali.
5. Interazione biologica e sociale durante l'infanzia e la giovinezza.
Il ruolo dell'eredità per la crescita è grande, ma questo non è l'unico fattore. L'ereditarietà dovrebbe essere considerata come un programma approssimativo, secondo il quale l'altezza di una persona può essere, ad esempio, compresa tra 160 e 180 cm. Ciò che sarà effettivamente dipende in gran parte dalle condizioni esterne che possono rallentare il programma ereditario o contribuire alla sua attuazione. Condizioni ambientali che influenzano la crescita umana: alimentazione, attività fisica, effetti psicologici del fumo, alcol.
Cioè, la popolazione sta crescendo (questa è l'accelerazione).
Una delle sue probabili cause è il miglioramento delle condizioni di vita (nutrizione). Si nota che durante gli anni di guerra e disastri naturali la crescita dei bambini diminuisce. La crescita è leggermente influenzata dal clima e dall'ambiente geografico.
6. Formazione di tipi costituzionali, tipi di corpo.
La formazione di tipi costituzionali di persone è collegata alla crescita di una persona. Questo dovrebbe essere inteso come le caratteristiche delle forme esterne del corpo, le caratteristiche delle funzioni del corpo, le caratteristiche del comportamento questa persona. A seconda della struttura del corpo, a seconda delle forme esterne del corpo, si distinguono alcuni tipi di fisico. Attualmente, ci sono parecchie classificazioni. Uno di questi è la classificazione di M.V. Chernorutsky. Secondo questo a
Ogni organismo, indipendentemente dal fatto che sia unicellulare o pluricellulare ea quale regno di vita appartenga, passa attraverso sviluppo individuale, O ontogenesi(dal greco. ontos- creatura e genesi- nascita). Il campo dello studio della biologia ontogenesi, è chiamato biologia dello sviluppo.
Ontogenesi - questo è il periodo di vita dell'organismo dallo zigote (cellula primaria) alla morte.
Negli organismi multicellulari, l'ontogenesi di solito inizia con la formazione di uno zigote e termina con la morte.
Allo stesso tempo, il corpo non solo cresce, aumentando di dimensioni, ma attraversa anche una serie di diverse fasi della vita, ognuna delle quali ha una struttura speciale, funziona in modo diverso e, in alcuni casi, uno stile di vita radicalmente diverso. .
Negli organismi unicellulari, l'inizio dell'ontogenesi è considerato il momento della separazione dalla cellula madre o sorella. Continua fino alla prossima divisione o morte. Allo stesso tempo, l'ontogenesi esterna degli organismi unicellulari di solito si manifesta solo come un leggero aumento delle dimensioni cellulari, sebbene in realtà questo nasconda periodi completamente diversi della sua vita. .
Ogni specie ha il proprio programma di ontogenesi. E questo non è solo un insieme e una sequenza di stadi di sviluppo che attraversa, ma anche la durata di ciascuno di essi. Allo stesso tempo, ogni individuo ha caratteristiche individuali di ontogenesi, che però non vanno oltre la specie. , e quelli, a loro volta, obbediscono alle leggi dell'ontogenesi a livello tribale, familiare, di distacco e persino di classe .
programma di ontogenesi non è altro che la realizzazione di informazioni ereditarie registrate nei geni. Pertanto, la specificità dell'ontogenesi a livello dei singoli individui è determinata da singole combinazioni di geni e, a livello di specie, generi, famiglie, da geni speciali che sono caratteristici solo di ciascun gruppo sistematico di organismi.
Il meccanismo per l'implementazione delle informazioni ereditarie è, prima di tutto, in differenziale(dall'inglese. differire- variare) attività genica. Ciò significa che in diversi periodi di sviluppo e in diversi tessuti organismo pluricellulare sono attivi i geni, sulle sequenze nucleotidiche di cui sono sintetizzati alcuni mRNA. Di conseguenza, vengono sintetizzate proteine ed enzimi strutturali specifici che, in ultima analisi, determinano le caratteristiche del funzionamento e del comportamento delle cellule in determinate fasi dello sviluppo del corpo, le caratteristiche del loro metabolismo.
Negli animali e nelle piante da fiore, l'ontogenesi è divisa in due periodi. Sviluppo embrionale, O embriogenesi(dal greco. embrione- feto e genesi) dura dalla formazione di uno zigote alla nascita o all'uscita dall'uovo, e postembrionale(dal greco. veloce- dopo ed embrione) lo sviluppo continua dalla nascita o dall'uscita dall'uovo e termina con la morte dell'organismo.
Il corso dell'embriogenesi. Lo schema generale dell'embriogenesi comprende le seguenti fasi: materiale dal sito
- successive divisioni dello zigote, culminanti nella formazione di un embrione multicellulare, costituito da centinaia e persino migliaia di cellule identiche;
- differenziazione (dall'inglese. differire- distinguere) cellule, portando alla formazione di tessuti;
- deposizione degli organi e crescita dell'embrione.
Durante ontogenesi l'organismo attraversa una serie di fasi - stati in cui differisce per struttura, funzionamento e stile di vita. Esistono due fasi dell'ontogenesi: embriogenesi - sviluppo embrionale e sviluppo postembrionale- il periodo di vita dell'organismo dalla nascita (uscita dall'uovo) alla morte.
In questa pagina, materiale sugli argomenti:
Sintesi dell'ontogenesi
Ontogenesi dello sviluppo individuale abstract
Il concetto di ontogenesi in breve
-
Capitolo 7. PERIODIZZAZIONE DELL'ONTOGENESI
Capitolo 7. PERIODIZZAZIONE DELL'ONTOGENESI
7.1. stadi, periodi e stadi dell'ontogenesi
Lo sviluppo individuale è un processo olistico continuo in cui i singoli eventi sono interconnessi nello spazio e nel tempo. Esistono diverse varianti di periodizzazione dell'ontogenesi, ognuna delle quali è più adatta per risolvere specifici problemi scientifici o pratici.
CON biologico generale Punti di vista evento principale ontogenesi - riproduzione sessuale. Se correliamo diversi periodi di tempo dell'ontogenesi con la capacità di un individuo di svolgere la funzione di riproduzione, allora può essere suddiviso in tre periodi: pre-riproduttivo, riproduttivo attivo e post-riproduttivo.
IN periodo pre-riproduttivo individuo è incapace di riprodursi. Il suo contenuto principale risiede nello sviluppo di un fenotipo sessualmente maturo. Durante questo periodo si verificano le trasformazioni strutturali e funzionali più pronunciate, si realizza la maggior parte delle informazioni ereditarie, il corpo è altamente sensibile a tutti i tipi di influenze.
Inattivo periodo riproduttivo un individuo svolge la funzione di riproduzione sessuale, si distingue per il funzionamento più stabile di organi e sistemi, nonché per la relativa resistenza alle influenze.
Periodo post-riproduttivo associato all'invecchiamento del corpo ed è caratterizzato da un indebolimento o completa cessazione della partecipazione alla riproduzione. Ridotta capacità di adattamento e resistenza a una varietà di influenze. In relazione all'ontogenesi umana, questi periodi sono inoltre caratterizzati da fattori sociali specifici (istruzione, capacità di lavorare, creatività). Ogni periodo ha le sue caratteristiche di morbilità.
Il periodo pre-riproduttivo è suddiviso in altri quattro: embrionale, larvale, metamorfosi e giovanile.
Embrionale, O germinale, punto L'ontogenesi inizia dal momento della fecondazione e continua fino al rilascio dell'embrione dalle membrane dell'uovo. Questo periodo si distingue per la gravità dei processi di trasformazione dello zigote in un organismo capace di un'esistenza più o meno indipendente. Nella maggior parte dei vertebrati, include gli stadi (fasi) di zigote, scissione, gastrulazione, nonché isto e organogenesi. La sua durata varia. Nei mammiferi placentari, è particolarmente accorciato. Il singolo guscio d'uovo si dissolve prima dell'impianto della blastocisti nel rivestimento uterino. A questo punto, l'embrione ha solo il tempo di passare attraverso le fasi dello zigote e della frantumazione. Tutti gli ulteriori processi procedono sotto la protezione e con la partecipazione del corpo della madre. Il significato evolutivo di queste caratteristiche è considerato nella Sezione 13.2.
La scissione è preceduta dai processi di gametogenesi e fecondazione, che sono direttamente correlati allo sviluppo individuale e potrebbero anche non condurvi, ma che determinano in gran parte l'ulteriore sviluppo dell'embrione se avviene il concepimento. Questi processi sono chiamati progenesi, che precede l'ontogenesi propriamente detta. Citologicamente, i processi di gametogenesi e fecondazione rappresentano un anello intermedio che collega l'ontogenesi dei genitori con l'ontogenesi della loro prole.
Periodo larvale in una variante tipica, si osserva nello sviluppo di quei vertebrati i cui embrioni emergono dalle membrane delle uova e cominciano a condurre uno stile di vita autonomo senza raggiungere i caratteri definitivi (maturi) dell'organizzazione. Quindi, si trova in alcuni rappresentanti dei vertebrati inferiori: lamprede, la maggior parte dei pesci ossei e degli anfibi. I tratti più caratteristici della larva sono: la natura embrionale della sua organizzazione, la presenza di organi temporanei (provvisori), l'inizio precoce del funzionamento di un certo numero di organi, che rende possibile un'esistenza indipendente. Grazie all'alimentazione attiva, la larva ha l'opportunità di completare lo sviluppo e, grazie al movimento attivo, ha l'opportunità di scegliere condizioni ambientali ottimali per lo sviluppo, evitando così la competizione con i propri parenti adulti. Nei vertebrati la durata del periodo larvale è molto più lunga di quella del periodo embrionale.
Metamorfosi consiste nella trasformazione della larva in una forma giovanile. Nel processo di metamorfosi si verificano importanti trasformazioni morfogenetiche come la distruzione parziale, la ristrutturazione e la nuova formazione di organi. Il grado di trasformazione è tanto maggiore quanto maggiore è
differenze tra l'habitat della larva e l'organismo adulto, che è ben illustrato dall'esempio dello sviluppo di anfibi senza coda (confronta: girino e rana).
Giovanile il periodo inizia dal momento del completamento della metamorfosi (nei mammiferi placentari e nell'uomo - dalla nascita) e termina con la pubertà e l'inizio della riproduzione. Le caratteristiche del periodo giovanile si manifestano nell'originalità della nutrizione del giovane organismo, nel suo comportamento e nel grado di dipendenza dai genitori. Da un punto di vista morfologico, questo periodo è caratterizzato da una crescita intensiva, l'istituzione di proporzioni finali tra varie parti corpo, completamento dello sviluppo dello scheletro, della pelle, cambio dei denti, completamento dello sviluppo delle gonadi e regolazione ormonale. La durata del periodo giovanile nei vertebrati varia da un minimo di 13-18 giorni nei topi-arvicole a un massimo di 18-20 anni in beluga, coccodrillo, albatros ed elefante. In molti rappresentanti dei vertebrati, specialmente nell'uomo, il raggiungimento della pubertà e l'inizio della riproduzione possono essere separati da un periodo di tempo significativo.
Applicazione ecologico ed embriologico approccio ci permette di dividere l'ontogenesi in fasi che si verificano prima della nascita, durante E dopo la nascita individui. La nascita stessa, ad es. il rilascio di un individuo in via di sviluppo dai gusci dell'uovo o dal corpo della madre, in tipi diversi avviene in diversi stadi di maturità. Allo stesso tempo, in tutte le specie prima della nascita, il corpo è protetto dalle membrane delle uova o dal corpo della madre e non è in grado di nutrirsi e svolgere da solo altre importanti funzioni. La protezione dei primi stadi morfofunzionalmente immaturi assicura la sopravvivenza della specie. Dopo la nascita, l'individuo stabilisce connessioni con il nuovo ambiente, inizia a nutrirsi da solo, a muoversi ea svolgere tutte le altre funzioni.
Una presentazione più dettagliata della periodizzazione dell'ontogenesi umana, che ha valore pratico nella pratica ostetrica e pediatrica, vedere la clausola 7.6.1 e la tabella. 7.3.
7.2. caratteristiche morfofisiologiche ed evolutive delle uova di cordati
7.2.1. ORIGINE DELLE UOVA
Le uova cordate sono costituite da un ovulo, a volte chiamato anche uovo, e dalle membrane dell'uovo. Ovociti formata nel genitale femminile
ghiandola - ovaia. Attraversano un lungo percorso di sviluppo, che inizia nell'embrione e continua nel periodo riproduttivo attivo dell'ontogenesi degli individui femminili.
Cellule sessuali primarie molto presto nell'embriogenesi sono isolati dalle cellule somatiche. Quindi, negli anfibi senza coda, i gonociti sono isolati allo stadio di blastula, negli uccelli sono determinati durante la formazione dell'ipoblasto primario, nei mammiferi nell'epiblasto allo stadio di gastrula, negli anfibi caudati questo processo avviene più tardi, allo stadio di gastrula o anche neurula (nel mesoderma). Quindi queste cellule si muovono, raggiungendo infine i rudimenti delle gonadi. I gonociti dei mammiferi si trovano dapprima nella regione extraembrionale all'imboccatura del sacco vitellino, quindi migrano verso l'anlage delle gonadi (Fig. 7.1). I meccanismi di migrazione sono definiti nella clausola 6.5.3.
Una volta nelle gonadi, le cellule germinali primarie iniziano a proliferare. Si dividono per mitosi e sono chiamati ovo(oo)gonia. Nella maggior parte dei vertebrati inferiori, l'ovo(oo)gonia conserva la capacità di dividersi durante l'intero periodo riproduttivo, ad esempio i pesci rilasciano migliaia di uova in una deposizione delle uova, gli anfibi - centinaia. Nei vertebrati superiori, il numero di uova che maturano contemporaneamente raramente raggiunge le 15, di solito sono meno, a volte una, il che può anche spiegare le caratteristiche dell'ovo(oo)genesi.
Negli embrioni femminili umani, l'ovo(oo)gonia si riproduce più intensamente tra il 2° e il 5° mese di vita intrauterina
Riso. 7.1. Localizzazione delle cellule germinali primarie in un embrione umano allo stadio di 16 somiti (a) e loro migrazione all'anlage delle gonadi (b): 1 - intestino anteriore; 2 - posteriore; 3 - allantoide; 4 - cellule germinali primarie; 5 - sacco vitellino; 6 - cuore; 7 - gonade in via di sviluppo (tubercolo genitale); 8 - rene primario; 9 - cloaca
esimo periodo di sviluppo, quando il loro numero raggiunge circa 7 milioni Entro il 7 ° mese, molti ovo(oo)gonia muoiono e i restanti entrano nella profase della prima divisione della meiosi e si fermano allo stadio della diacinesi. Con l'inizio della pubertà, un ovocita ovula ogni mese, raggiungendo lo stadio della metafase della seconda divisione della meiosi. Per alcuni ovuli ciò si verifica al momento della pubertà e per altri appena prima della menopausa. L'ovocita completa la meiosi solo se avviene la fecondazione.
7.2.2. SPECIFICITÀ E SIGNIFICATO DELLA COMPOSIZIONE CHIMICA DEL CITOPLASMA DELL'UOVO
Un uovo maturo, di regola, è più grande di un ovo(oo)gonia e di qualsiasi altra cellula somatica. Durante l'ovo(oo)genesi nel citoplasma dell'uovo, un gran numero di sostanze necessarie per la sua maturazione e l'embriogenesi precoce. Pertanto, durante il periodo di crescita, il volume di un ovocita di topo aumenta di circa 40 volte e quello di un ovocita di rana aumenta di oltre 400. Il tasso di sintesi delle sostanze negli ovociti aumenta in modo significativo, a causa di una serie di caratteristiche. Nelle uova in maturazione si verifica l'amplificazione (vedi Sezione 2.4.3.4) di singoli geni (vedi Sezione 4), ad esempio i geni rRNA. Anfibi (Xenopo laevis) circa 450 geni rRNA si trovano in una cellula diploide e più di un milione in un ovo(oo)cito. Inoltre, l'attività trascrizionale del genoma di queste cellule aumenta. Pertanto, il tasso di trascrizione negli ovociti di topo in crescita è 10 volte superiore a quello delle cellule somatiche.
Il ruolo funzionale delle sostanze immagazzinate è diverso.
In primo luogo, questi sono i componenti necessari per i processi di replicazione, trascrizione e traduzione, come i corrispondenti enzimi, ribosomi, mRNA, tRNA e loro precursori.
In secondo luogo,è un insieme di sostanze normative specifiche che assicurano il funzionamento coordinato di tutti i componenti immagazzinati. Queste sostanze includono il fattore di disintegrazione della membrana nucleare (la prometafase della 1a divisione della meiosi inizia con la distruzione della membrana nucleare); un fattore che provoca la condensazione dei cromosomi; un fattore che converte il nucleo dello sperma in un pronucleo e attiva la sintesi del DNA in esso prima della scissione; fattore citostatico csf, responsabile del blocco della meiosi allo stadio della metafase II (in molti vertebrati la rimozione di questo blocco avviene solo a seguito della fecondazione); sistema di microfilamenti citoscheletrici coinvolti nel mantenimento del blocco
meiosi; nucleotidi ciclici (p. es., cAMP), protein chinasi e altre sostanze necessarie per la segnalazione dalle cellule ovo(oo)cyt circostanti e dall'ambiente extracellulare; componenti del cosiddetto sistema inositolo fosfato, che assicurano l'attivazione dell'uovo dopo la fecondazione; fattore responsabile della citotomia durante la scissione. Alcuni di loro, trovandosi nell'ovocita già al momento della fecondazione, iniziano ad agire solo nella fase di gastrulazione.
Terzo,è il tuorlo, che comprende proteine, fosfolipidi, grassi neutri, carboidrati, sali minerali. Il tuorlo è un deposito di nutrienti e risorse energetiche necessarie per garantire il periodo embrionale. Molte sostanze prodotte dal fegato entrano nell'ovo(oo)gonia durante la loro crescita attraverso le cellule follicolari dell'ovaio. Ciò richiede un grande dispendio energetico da parte della femmina.
Il quarto, Queste sono sostanze specifiche chiamate determinanti morfogenetici, determinare piuttosto rigidamente il destino delle cellule (blastomeri) in cui cadono durante la scissione. Sostanze simili non si trovano nel citoplasma delle uova in tutti gli animali.
A causa delle caratteristiche descritte della composizione chimica del citoplasma dell'uovo, l'embrione durante il periodo di schiacciamento in alcuni casi non utilizza le informazioni genetiche dei nuclei dello zigote e dei blastomeri per lo sviluppo. Specifica Composizione chimica e la regolare distribuzione delle sostanze nel citoplasma dell'uovo sono di grande importanza per le fasi iniziali dell'embriogenesi. I nutrienti immagazzinati e le sostanze energetiche assicurano lo sviluppo embrionale senza il loro ulteriore apporto dall'esterno.
7.2.3. LA DIMENSIONE DELLE UOVA E IL LORO RUOLO NELL'EVOLUZIONE. TIPI DI UOVA
Nel processo di sviluppo si rivela una regolarità, che consiste nel fatto che più lungo è il periodo embrionale, più tuorlo dovrebbe accumularsi nell'uovo. La durata del periodo embrionale dipende dallo stadio in cui l'embrione passa all'esistenza indipendente nell'ambiente esterno. Se lo sviluppo postembrionale è diretto, cioè senza larva e metamorfosi, allora dovrebbe esserci più tuorlo nell'uovo. In base al numero di tuorlo, le uova cordate (Tabella 7.1) sono suddivise in alecitale, oligo-, meso- E poliletrico, quelli. con una trascurabile quantità di tuorlo piccola, piccola, media e grande (dal greco. lecithos- tuorlo) (Tabella 7.2).
Tabella 7.1.Sistematica del tipo Chordata
Tabella 7.2.Tipi di ovociti trovati nei cordati

A lancetta, rappresentante dei cordati inferiori, uovo oligolecitale. Nella maggior parte dei vertebrati, le uova contengono una quantità significativa di tuorlo. Tra i vertebrati inferiori (Anamnia) le uova più grandi si trovano nelle missine (classe Ciclostomi), negli squali e nelle chimere (classe Pesci cartilaginei) e in alcuni anfibi. Il riposo anfibio, così come le uova di storione mesolecitale, quelli. avere una quantità media di tuorlo. Nei vertebrati superiori (Amniota) come rettili, uccelli e mammiferi ovipari, poliletrico, quelli. c'è molto tuorlo nell'uovo. Il loro sviluppo embrionale richiede un tempo particolarmente lungo.
Questo schema è interrotto nei marsupiali e placentare mammiferi che hanno oligo- E alecithal uova, rispettivamente. Nei marsupiali, l'embrione emerge dalle membrane dell'uovo e dall'utero con organogenesi incompleta, viene trasferito nella sacca, dove continua lo sviluppo. Nei placentali, compresi gli esseri umani, esce l'embrione
dalle membrane dell'uovo anche prima, allo stadio di blastocisti, ma poi passa all'esistenza intrauterina, dove compie tutti i principali periodi di sviluppo che lo preparano alla nascita. Diminuire si può chiamare la quantità di tuorlo nelle uova dei mammiferi secondario, perché i loro antenati, che hanno dominato ambiente terrestre, aveva, come tutti gli amnioti, uova polilecitali.
7.2.4. POLARITÀ DELL'OVA
Polarità degli ovocitiè pianificato anche nella fase di accumulo del tuorlo negli ovo(oo)citi durante la loro rapida (grande) crescita ed è fissato durante l'isolamento dei corpi polari (riduzione). Dopo l'isolamento del secondo corpo di riduzione, la polarità diventa stabile e irreversibile, come dimostrano gli esperimenti di Guerrier sulla centrifugazione delle uova a diversi stadi della loro maturazione. Viene chiamato il polo in cui spiccano i corpi di riduzione animale, ed è il contrario vegetativo. I poli della cellula uovo differiscono in molti modi: concentrazione varie sostanze, il numero di organelli, l'attività dei processi intracellulari e molti altri. Sì, esperimenti anni recenti utilizzando elettrodi vibranti rivelato campi elettrici intorno agli ovociti e alle uova di un certo numero di animali e piante e il flusso di correnti elettriche attraverso il loro citoplasma. Si ritiene che ciò sia dovuto a diverse concentrazioni di canali ionici e pompe ai poli opposti degli ovociti. Nell'uovo della rana artigliata, la concentrazione dei canali ionici è maggiore al polo animale e la concentrazione delle pompe è maggiore al polo vegetativo.
Accumulo di tuorlo da parte dell'uovo- la prima manifestazione della sua polarizzazione. Con una piccola quantità di tuorlo nell'uovo, di solito è distribuito uniformemente nel citoplasma e il nucleo si trova approssimativamente al centro. Queste uova sono chiamate isolecitale(dal greco. isos- uguale). La maggior parte dei vertebrati ha molto tuorlo ed è distribuito in modo non uniforme nel citoplasma dell'uovo. Questo anisolecitale cellule. La maggior parte del tuorlo si accumula polo vegetativo. Queste uova sono chiamate telolecita(dal greco. telos- FINE). Verso il contrario animale il citoplasma attivo libero dal tuorlo viene spinto da parte verso il polo. Se il tuorlo è ancora immerso nel citoplasma e non ne è isolato come frazione separata, come negli storioni e negli anfibi, le uova sono chiamate moderatamente telolecitale. Se il tuorlo è completamente separato dal citoplasma, come negli amnioti, allora questo nettamente telolecitale uova.
Le caratteristiche della dimensione e della polarità delle uova cordate sono raggruppate schematicamente nella Tabella. 7.2.
Nei processi di polarizzazione dell'uovo, a quanto pare, prende parte e corteccia- Questo è lo strato superficiale del citoplasma dell'uovo, situato direttamente sotto la membrana plasmatica. Contiene microfilamenti e granuli corticali. Questi ultimi contengono un'intera gamma di sostanze coinvolte nella formazione della membrana di fertilizzazione e nella prevenzione della polispermia. L'assemblaggio e la ridistribuzione degli elementi del citoscheletro dello strato corticale osservati dopo la fecondazione assicurano l'acquisizione della contrattilità da parte sua, necessaria per le divisioni della scissione dello zigote. Nella maggior parte degli animali, i primi due solchi di scissione corrono lungo piani animale-vegetativo reciprocamente perpendicolari (meridiani che collegano i poli animale e vegetativo).
In generale, al polo animale dell'uovo, di regola, c'è più citoplasma libero, organelli, RNA immagazzinato, di solito anche il nucleo si trova al polo animale o più vicino ad esso. Il polo vegetativo è caratterizzato dalla predominanza dei granuli di tuorlo.
La polarizzazione dell'uovo è accompagnata dall'aspetto segregazione ovo(oo)-plasmica uova, ad es. creazione di eterogeneità interna delle sezioni del citoplasma dell'uovo.
La polarizzazione animale-vegetativa dell'uovo è di importanza decisiva per tutti i successivi processi di embriogenesi, poiché determina la futura organizzazione spaziale dell'embrione. Negli animali adulti, l'asse antero-posteriore del corpo coincide con l'asse animale-vegetativo dell'uovo (ad esempio, nei vertebrati) o è perpendicolare ad esso (ad esempio, negli oligocheti e in alcuni artropodi).
7.2.5. GUSCI D'UOVO
Le uova sono ricoperte all'esterno da una o più membrane, che successivamente svolgono, tra l'altro, la funzione di proteggere l'embrione in via di sviluppo.
Distinguere guscio primario, formata dall'ovulo stesso guscio secondario- un prodotto dell'attività delle cellule follicolari ovariche, e conchiglie terziarie, da cui l'uovo è circondato durante il passaggio attraverso l'ovidotto.
guscio primario, a volte chiamato tuorlo, presente nelle uova di tutti gli animali. Nei vertebrati, compresi i mammiferi,
ing, il guscio primario fa parte del guscio denso, formandone la parte interna. La parte esterna del guscio denso produrre cellule follicolari guscio secondario. Il guscio denso è permeato dall'interno dai microvilli dell'uovo e dall'esterno dai microvilli delle cellule follicolari. Per il loro proprietà ottiche nei mammiferi si chiama guscio lucido(zona pellucida). Quindi questo guscio combina primario e secondario. In cima alla zona pellucida c'è corona radiosa(corona radiata), formato da cellule follicolari che aderiscono all'uovo mentre si trova nel follicolo ovarico (Fig. 7.2).
La zona pellucida è un complesso complesso di glicoproteine extracellulari, che nei mammiferi sono designate ZP. Nei topi, è costituito da tre diverse glicoproteine solfatate: ZP1, ZP2, ZP3. Usando ZP3 gli spermatozoi si legano alla zona pellucida e, dopo la penetrazione di uno spermatozoo, questa glicoproteina viene modificata e la penetrazione di un altro
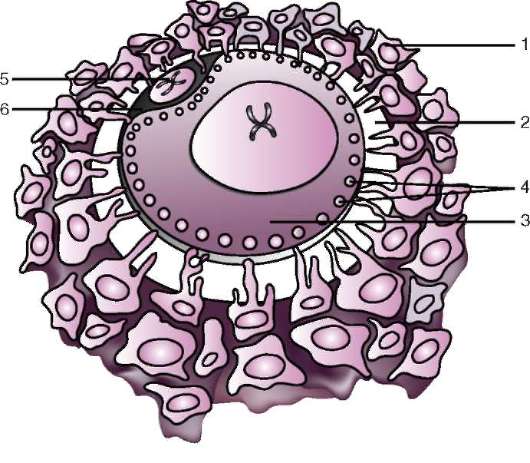
Riso. 7.2. Ovum (ovocita del primo ordine) di mammiferi: 1 - corona radiante; 2 - guscio lucido; 3 - citoplasma; 4 - granuli corticali; 5 - corpo polare; 6 - spazio perituorlo
gih spermatozoi diventa impossibile. Inoltre, le glicoproteine sono specie-specifiche, il che impedisce la fecondazione interspecifica. In altri cordati, la specificità della specie della fecondazione è determinata dall'interazione delle proteine dello sperma con i recettori. membrana di tuorlo. La zona pellucida non confina direttamente con la membrana dell'uovo, ma si separa spazio perivitellino (periyotellino). Dopo la penetrazione del primo spermatozoo nell'uovo, il granulo corticale ovoperossidasi entra in questo spazio. Si ritiene che l'azione di questo enzima modifichi ZP3 E Zp2, che porta all'indurimento della zona pellucida. Persiste attorno all'embrione per tutto il periodo preimpianto o per una parte significativa di questo periodo. Il guscio lucido impedisce agli embrioni adiacenti di attaccarsi tra loro e agli embrioni di attaccarsi alle pareti dell'ovidotto e dell'utero. È noto che nelle fasi iniziali della frantumazione fino alla blastocisti, i blastomeri hanno un'elevata adesività. Se un embrione privo di zona pellucida viene trapiantato nell'ovidotto, i blastomeri aderiranno inevitabilmente alla parete dell'ovidotto e l'embrione morirà. Inoltre, a causa della zona pellucida, i blastomeri sono compatti e ordinati, il che favorisce la formazione di contatti e l'interazione tra di loro e garantisce il normale sviluppo dell'embrione in questa fase. Se il guscio viene rimosso, la frammentazione continuerà, ma i blastomeri saranno disposti in una catena e la loro compattazione sarà completamente disturbata o sarà molto tardiva.
Conchiglie terziarie ben sviluppati nei pesci cartilaginei e negli anfibi, ma acquisiscono particolare complessità nei vertebrati terrestri: rettili, uccelli e mammiferi inferiori. Formare dalle secrezioni delle ghiandole dell'ovidotto, questi gusci non hanno una struttura cellulare. In tutti i vertebrati svolgono le funzioni di protezione dell'embrione dai danni meccanici e dall'azione di fattori biotici dannosi, quali fattori batterici, fungini e protozoici. Inoltre, nei vertebrati terrestri compaiono funzioni fondamentalmente nuove di immagazzinamento di acqua e sostanze nutritive per soddisfare i bisogni dell'embrione. Nei rettili, la membrana del guscio funge da pompa, prelevando l'acqua dal suolo e dall'aria. Gli uccelli hanno una riserva d'acqua guscio proteico. L'assorbimento e l'evaporazione dell'acqua sono regolati dai pori della membrana del guscio. Il guscio contiene molti sali minerali necessari per lo sviluppo dello scheletro dell'embrione.
7.3. FERTILIZZAZIONE E PARTENOGENESI
Fecondazione- questo è il processo di fusione delle cellule germinali, che culmina nell'unificazione del loro materiale genetico. La cellula diploide derivante dalla fecondazione zigote- rappresenta la fase iniziale di sviluppo di un nuovo organismo.
Il processo di fecondazione si compone di tre fasi successive:
Interazione a distanza e convergenza dei gameti;
Interazione di contatto dei gameti e attivazione dell'uovo;
Fusioni di gameti o singamia.
Approssimazione dello spermatozoo con l'uovo forniti da una combinazione di fattori non specifici che aumentano la probabilità del loro incontro e interazione. Questi includono il coordinamento dell'inizio della prontezza alla fecondazione nel maschio e nella femmina, il comportamento di maschi e femmine, che assicura la copulazione e l'inseminazione, l'eccessiva produzione di spermatozoi, le grandi dimensioni dell'uovo, nonché quelli prodotti da uova e spermatozoi sostanze chimiche, contribuendo al riavvicinamento e all'interazione delle cellule germinali. Queste sostanze, chiamate gamon(ormoni dei gameti), da un lato, attivano il movimento degli spermatozoi e, dall'altro, il loro legame. Nel movimento degli spermatozoi dei mammiferi lungo le parti superiori dell'ovidotto, il fenomeno reotassi: la loro capacità di muoversi contro il flusso in arrivo di fluido nelle tube di Falloppio.
Nei mammiferi, la permanenza degli spermatozoi nel tratto genitale femminile è di grande importanza, per cui capacitazione degli spermatozoi- la loro acquisizione della capacità fecondante. Immediatamente dopo essere entrato nel tratto genitale, lo sperma femminile non è in grado di penetrare nelle uova. Le glicoproteine e le proteine del plasma seminale vengono rimosse dal plasmolemma spermatico nella zona acrosomiale sotto l'azione delle sostanze del segreto del tratto genitale femminile, che bloccano i centri attivi delle molecole recettrici del plasmolemma spermatico, riconoscendo la superficie del plasmolemma spermatico cellula riproduttiva. Inoltre, le molecole di albumina presenti nel tratto genitale femminile si legano al colesterolo della membrana delle cellule spermatiche. Ciò porta alla destabilizzazione del plasmalemma degli spermatozoi e della sua membrana acrosomiale, che facilita il successivo rilascio di enzimi acrosomiali. È stato anche scoperto che nel processo di capacitazione, le proprietà della superficie degli spermatozoi, come la sua carica, cambiano. Inoltre, c'è un cambiamento nella mobilità (iperattivazione) degli spermatozoi e l'attivazione degli enzimi acrosomali. Negli esseri umani, la capacitazione dura circa 7 ore.
In fase interazione di contatto lo sperma distrugge le membrane dell'uovo (nei mammiferi - la corona radiante, la membrana trasparente) e la membrana citoplasmatica dell'ovocita a causa di reazione acrosomiale. Al contatto con il guscio della cellula germinale femminile sotto l'azione delle sue sostanze attivanti (una delle quali è fertilizin), viene avviato un flusso attivo di cationi di calcio nella testa dello spermatozoo. Di conseguenza, si verifica la fusione focale della cellula e delle membrane acrosomali degli spermatozoi e la loro parziale distruzione. Attraverso i microfori formati vengono rilasciati gli enzimi dello sperma - ialuronidasi, penetrasi e altri, che disconnettono i contatti tra le cellule della corona radiante, nonché tra loro e l'ovocita. L'enzima acrosomiale acrosina distrugge una parte della zona pellucida della cellula germinale femminile e lo sperma penetra nello spazio perilocale. Nel punto di contatto della testa dello spermatozoo con la membrana plasmatica dell'ovocita, avviene la fusione e la successiva distruzione delle membrane dei gameti femminili e maschili. Attraverso il ponte citoplasmatico risultante, il citoplasma di entrambi i gameti viene combinato. Quindi il nucleo e il centriolo dello spermatozoo passano nel citoplasma dell'uovo e la membrana dello spermatozoo è incorporata nella membrana della cellula uovo. La parte della coda dello spermatozoo rimane all'esterno o entra anche nell'uovo, ma poi si separa e si risolve senza svolgere alcun ruolo nell'ulteriore sviluppo.
Gli ioni di sodio iniziano a entrare attivamente nel citoplasma dell'uovo attraverso la membrana della cellula spermatica, a seguito della quale il potenziale di membrana dell'ovocita cambia drasticamente e la cellula germinale femminile diventa immune al contatto con altri spermatozoi - blocco rapido della polispermia. L'afflusso di ioni sodio provoca il rilascio di ioni calcio dai depositi intracellulari e un aumento del suo contenuto nel citoplasma dell'uovo, che si diffonde sotto forma di un'onda dal punto di contatto dei gameti. In seguito inizia questo reazione corticale: le membrane dei granuli corticali si fondono con la membrana dell'uovo e gli enzimi proteolitici da essi liberati entrano nello spazio periolino. Sotto l'influenza degli enzimi, il guscio d'uovo si ispessisce, si ispessisce, perde le proteine del recettore per gli spermatozoi e si trasforma in guscio di fecondazione. Inoltre, la glicoproteina secreta dai granuli corticali contribuisce al distacco della membrana del tuorlo dal plasmolemma dell'uovo. Come risultato di tutto quanto sopra, la penetrazione di altri spermatozoi diventa impossibile - blocco lento della polispermia. Nei mammiferi
la reazione corticale non provoca la formazione della membrana di fecondazione, ma la sua essenza è la stessa.
Come risultato del contatto dello spermatozoo con l'uovo, esso Attivazione. Consiste in complessi cambiamenti strutturali e fisico-chimici. Lo stadio iniziale dell'attivazione è la reazione corticale sopra descritta. In animali come echinodermi, pesci ossei e anfibi, i cambiamenti nel citoplasma dell'uovo sono accompagnati da visibili riarrangiamenti morfologici. Questi fenomeni sono chiamati stratificazione o segregazione plasmatica. Il suo significato per l'ulteriore sviluppo embrionale sarà discusso di seguito. In molte specie di anuri, la penetrazione di uno spermatozoo in un uovo porta a movimento dei granuli di pigmento polo animale, e un'area a forma di mezzaluna leggermente colorata appare contro il sito di penetrazione dello sperma, chiamata "falce grigia"(figura 7.3).
L'attivazione dell'uovo termina con l'inizio della sintesi proteica a livello traduzionale, poiché mRNA, tRNA, ribosomi ed energia sono stati immagazzinati nell'ovo(oo)genesi. L'attivazione dell'uovo può iniziare e procedere fino alla fine senza il nucleo dello spermatozoo e senza il nucleo dell'uovo, come dimostrato da esperimenti sull'enucleazione dello zigote.
L'uovo al momento dell'incontro con lo sperma si trova solitamente in uno degli stadi della meiosi, bloccato da un fattore specifico. Nella maggior parte dei vertebrati, questo blocco viene eseguito
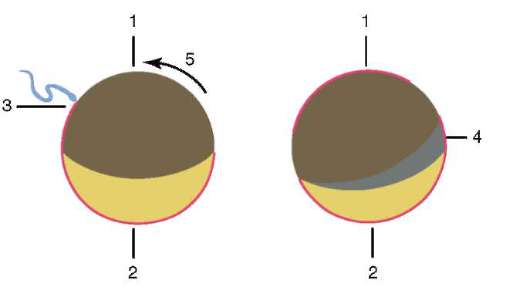
Riso. 7.3. Segregazione del citoplasma dell'uovo anfibio dopo la penetrazione dello sperma: 1 - polo animale; 2 - polo vegetativo; 3 - il luogo di introduzione dello spermatozoo; 4 - falce grigia; 5 - la direzione del movimento del citoplasma con granuli di pigmento
allo stadio della metafase II; in molti invertebrati, così come in tre specie di mammiferi (cavalli, cani e volpi), il blocco avviene allo stadio della diacinesi (profase I). Nella maggior parte dei casi, il blocco meiotico viene rimosso dopo l'attivazione dell'uovo a causa della fecondazione. Mentre la meiosi è completata nell'uovo, il nucleo dello sperma che è penetrato in esso viene modificato. Prende la forma di un'interfase e quindi di un nucleo di profase. Durante questo periodo, il DNA raddoppia e la quantità materiale ereditario v pronucleo maschile diventa p2s, quelli. contiene un insieme aploide di cromosomi raddoppiati.
Il nucleo dell'ovulo che ha completato la meiosi diventa pronucleo femminile, anche acquisendo p2s. Entrambi i pronuclei compiono movimenti complessi, poi si avvicinano e si fondono. (sincarion), formando una piastra metafasica comune. Questo, infatti, è il momento della fusione finale dei gameti - sinagoga. La prima divisione mitotica dello zigote porta alla formazione di due cellule embrionali (blastomeri) con una serie di cromosomi 2p2 in tutti.
In alcuni casi, lo sviluppo avviene senza fecondazione - partenogenesi(dal greco. partenoe- vergine). Quando partenogenesi naturale lo sviluppo procede sulla base del citoplasma e del pronucleo dell'uovo. Gli individui che si formano da un uovo hanno un set di cromosomi aploide o diploide, poiché viene attivato uno dei meccanismi per raddoppiare il numero di cromosomi. In alcuni casi, durante la meiosi della cellula germinale femminile, cade lo stadio di riduzione del numero di cromosomi e si ottiene l'uovo con un pronucleo diploide. In altri casi, la diploidizzazione si verifica durante la prima divisione di scissione, in cui non si verifica alcuna citotomia.
partenogenesi naturale, di norma, non è l'unico modo di riproduzione della specie. Si alterna alla normale riproduzione sessuale o si verifica nell'individuo gare. La partenogenesi naturale è stata trovata nelle generazioni estive di alcuni crostacei e rotiferi, nelle api, nelle vespe e in un certo numero di lepidotteri. Tra i vertebrati, la riproduzione partenogenetica è stata descritta in tre razze di lucertole rocciose dell'Armenia, costituite da sole femmine. Il 40% delle uova di tacchino deposte in assenza di un maschio può iniziare a svilupparsi, ma questo sviluppo raramente arriva alla fine, più spesso si interrompe per anomalie. In altre specie di vertebrati, la riproduzione partenogenetica naturale è sconosciuta.
partenogenesi artificiale sembra essere possibile in tutti gli animali. Lo sviluppo di metodi per lo sviluppo partenogenetico è importante
problema nelle relazioni scientifiche e applicate. Un grande contributo a questo problema è stato dato dai ricercatori domestici A.A. Tikhomirov, B.L. Astaurov, V.A. Strunnkov. È stato riscontrato che l'attivazione dell'uovo da parte dello spermatozoo non è specifica. Molti fattori fisici e chimici possono agire da attivatori. Sul baco da seta è stato dimostrato che con l'ausilio della partenogenesi artificiale è possibile regolare il rapporto tra sesso maschile e femminile nella popolazione, ottenendo un grande effetto economico.
La partenogenesi naturale si verifica più spesso con fecondazione incompleta, ad es. in quei casi in cui è avvenuta l'attivazione dell'uovo, ma il nucleo dello spermatozoo non ha partecipato alla fecondazione. Nelle uova attivate vengono utilizzate solo le informazioni dal pronucleo femminile. Questo tipo di partenogenesi è chiamato ginogenesi. Con la partenogenesi artificiale, è possibile rimuovere il pronucleo femminile, quindi lo sviluppo sarà effettuato solo a spese dei pronuclei maschili. Questo an-drogenesi. In speciali esperimenti sui ricci di mare, si è scoperto che la prole eredita solo i tratti della madre durante la ginogenesi o solo i tratti del padre durante l'androgenesi. Ciò indica che le proprietà ereditarie di un individuo sono determinate principalmente dal nucleo e non dal citoplasma.
7.4. sviluppo embrionale
7.4.1. FRAZIONAMENTO
7.4.1.1. L'essenza della fase di frantumazione
Frazionamento- questa è una serie di successive divisioni mitotiche dello zigote e dei risultanti blastomeri, che terminano con la formazione embrione multicellulare a strato singolo- blastula. La prima divisione di scissione inizia dopo l'unione del materiale ereditario dei pronuclei e la formazione di una placca metafasica comune. Vengono chiamate le cellule formate durante la scissione blastomeri(dal greco. esplodere- germoglio, germe). Una caratteristica dei cicli mitotici di frantumazione è che ad ogni divisione le cellule diventano sempre più piccole fino a raggiungere il rapporto tra i volumi del nucleo e del citoplasma che è normale per le cellule somatiche. In un riccio di mare, ad esempio, ciò richiede 6 divisioni e l'embrione è composto da 64 cellule. Non c'è crescita cellulare tra divisioni successive (periodo G 1 assente), ma il DNA è necessariamente sintetizzato. Tutti i precursori del DNA e necessari
Gli enzimi dimmerabili vengono accumulati nel processo di oogenesi. Di conseguenza, i cicli mitotici si accorciano e le divisioni si susseguono molto più velocemente che nelle normali cellule somatiche. Dopo diversi cicli di divisione, i blastomeri formati si uniscono l'uno all'altro, formando un ammasso di cellule chiamato morula. Quindi si forma una cavità tra le cellule - blastocele, riempito di liquido. Le cellule vengono spinte verso la periferia, formando il muro della blastula - blastoderma. A causa dell'assenza di crescita del blastomero durante il periodo di scissione, la dimensione totale dell'embrione allo stadio di blastula non supera la dimensione dello zigote.
7.4.1.2. Morfologia dello schiacciamento
Di norma, i blastomeri sono disposti in un ordine rigoroso l'uno rispetto all'altro e rispetto all'asse polare dell'uovo. L'ordine, o metodo di frantumazione, dipende dalla quantità, dalla densità e dalla distribuzione del tuorlo nell'uovo. Secondo le regole di Sachs-Hertwig, il nucleo cellulare tende a trovarsi al centro del citoplasma libero dal tuorlo e il fuso della divisione cellulare - nella direzione della massima estensione di questa zona.
Nelle uova oligo- e mesolecitali, scissione completare, O oloblastico. Questo tipo di schiacciamento si riscontra nelle lancette, nelle lamprede, in alcuni pesci, in tutti gli anfibi, oltre che nei marsupiali e nei mammiferi placentari. Con lo schiacciamento completo, il piano della prima divisione corrisponde al piano di simmetria bilaterale. Il piano della seconda divisione corre perpendicolare al piano della prima. Entrambi i solchi delle prime due divisioni sono meridiani, cioè iniziare dal polo animale e diffondersi al polo vegetativo. La cellula uovo è divisa in quattro blastomeri di dimensioni più o meno uguali. Il piano della terza divisione corre perpendicolare alle prime due in direzione latitudinale. Negli animali con uova isolecitali, tutti gli otto blastomeri formati sono approssimativamente uguali - frantumazione uniforme. Nelle uova mesolecitali, appare irregolarità schiacciamento. Al polo animale ci sono quattro blastomeri più piccoli: micrometri, sul vegetativo - quattro più grandi - macromeri. Quindi la divisione va di nuovo nei piani meridiani, e poi di nuovo in quelli latitudinali.
Negli ovociti polilecitali di pesci ossei, rettili, uccelli e anche mammiferi monotremi, la scissione parziale, O meroblastico, quelli. copre solo il citoplasma libero dal tuorlo. Si trova sotto forma di un disco sottile al polo animale, quindi viene chiamato questo tipo di schiacciamento discoidale.
Quando si caratterizza il tipo di frantumazione, vengono presi in considerazione anche la posizione relativa e il tasso di divisione dei blastomeri. Se i blastomeri sono disposti in file uno sopra l'altro lungo i raggi, si parla di schiacciamento radiale.È tipico dei cordati e degli echinodermi. In natura esistono altre varianti della disposizione spaziale dei blastomeri durante la frantumazione, che ne determinano le tipologie, come spirale nei crostacei bilaterale a nematodi, anarchico alla medusa.
È stata notata una relazione tra la distribuzione del tuorlo e il grado di sincronismo nella divisione dei blastomeri animali e vegetativi. Nelle uova oligolecitali di echinodermi e cefalocordi, la scissione è quasi sincrona; nelle uova mesolecitali, ad esempio, negli anfibi, la sincronia è disturbata dopo la terza divisione, poiché i blastomeri vegetativi si dividono più lentamente a causa della grande quantità di tuorlo. Negli organismi con scissione parziale (rettili, uccelli), le divisioni sono asincrone fin dall'inizio e i blastomeri che occupano una posizione centrale si dividono più velocemente.
Alla fine della frantumazione, blastula. Il tipo di blastula dipende dal tipo di frantumazione, e quindi dal tipo di uovo. Alcuni tipi di schiacciamento e blastula sono mostrati in Fig. 7.4 e schema (in tabella) 7.1. Per una descrizione più dettagliata della scissione nei mammiferi, compreso l'uomo, vedere la sezione 7.5.1.
Tabella 7.3.Tipi di schiacciamento e tipi di blastole cordate
7.4.1.3. Caratteristiche dei processi genetico-molecolari e biochimici durante la frantumazione
Come notato sopra, i cicli mitotici durante il periodo di scissione sono notevolmente accorciati, specialmente all'inizio. Ad esempio, l'intero ciclo di fissione nelle uova di riccio di mare dura 30-40 minuti con una durata di Fasi S solo 15 min.
GJ-, e in molti animali G 2- periodi sono praticamente assenti, poiché l'apporto necessario di tutte le sostanze è stato creato nel citoplasma dell'uovo, e maggiore è, più grande è. È stato dimostrato che dopo la rimozione dallo zigote
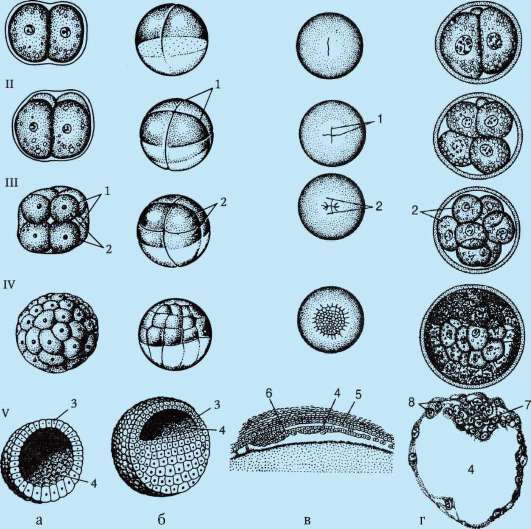
Riso. 7.4. Scissione in cordati con diversi tipi di uova: a - lancetta; b - rana; c - uccello; d - mammifero: I - 2 blastomeri; II - 4 blastomeri; III - 8 blastomeri; IV - morula; V - blastula; 1 - solchi schiaccianti; 2 - blastomeri; 3 - blastoderma; 4 - blastocele; 5 - epiblasto; 6 - ipoblasto; 7 - embrioblasto; 8 - trofoblasto; la dimensione degli embrioni nella figura non riflette la dimensione reale degli embrioni
si verifica la scissione del nucleo e l'embrione raggiunge nel suo sviluppo quasi lo stadio di blastula. L'ulteriore sviluppo si interrompe.
Prima di ogni divisione, avviene però la sintesi del DNA e degli istoni, la durata Periodo S accorciato. Durante la frammentazione durante la replicazione del DNA, la velocità di avanzamento della forcella di replicazione è normale. Allo stesso tempo, ci sono più punti di inizio nel DNA del blastomero che in
cellule somatiche, quindi i repliconi sono più corti. La sintesi del DNA avviene in tutti i repliconi contemporaneamente (in modo sincrono). Pertanto, il tempo di replicazione del DNA nel nucleo coincide con il tempo di raddoppio di un replicone, per di più accorciato.
All'inizio della scissione, altri tipi di attività nucleare, come la trascrizione, sono praticamente assenti. In diversi tipi di uova, la trascrizione genica e la sintesi dell'RNA iniziano in fasi diverse. Nei casi in cui nel citoplasma sono presenti molte sostanze diverse, come ad esempio negli anfibi, la trascrizione non si attiva immediatamente. La sintesi dell'RNA in essi inizia nella fase della prima blastula. Al contrario, nei mammiferi, la sintesi dell'RNA inizia già allo stadio di due blastomeri.
Durante il periodo di scissione si formano RNA e proteine, simili a quelli sintetizzati durante l'oogenesi. Principalmente istoni, proteine membrane cellulari ed enzimi necessari per la divisione cellulare. Queste proteine vengono utilizzate immediatamente insieme a quelle precedentemente immagazzinate nel citoplasma delle uova. Insieme a questo, durante il periodo di scissione, è possibile la sintesi di proteine che prima non erano possibili, come evidenziato dai dati sulla presenza di differenze regionali nella sintesi di RNA e proteine tra i blastomeri. A volte queste sostanze iniziano ad agire nelle fasi successive.
Un ruolo importante nella frantumazione è svolto dalla divisione del citoplasma - citotomia. Ha un particolare significato morfogenetico, poiché determina il tipo di frantumazione. Nel processo di citotomia, si forma prima una costrizione con l'aiuto di un anello contrattile di microfilamenti. L'assemblaggio di questo anello avviene sotto l'influenza diretta dei poli del fuso mitotico. Dopo la citotomia, i blastomeri delle uova oligolecitiche rimangono collegati tra loro solo da sottili ponti. È in questo momento che sono più facili da separare. Questo perché la citotomia porta ad una riduzione dell'area di contatto tra le cellule a causa della superficie limitata delle membrane.
Immediatamente dopo la citotomia, inizia la sintesi di nuove sezioni della superficie cellulare, la zona di contatto aumenta ei blastomeri iniziano a toccarsi strettamente, il che offre la possibilità di interazione intercellulare. I solchi di scissione corrono lungo i confini tra le singole sezioni dell'ovoplasma, quindi il citoplasma di diversi blastomeri differisce nella composizione chimica. Ciò riflette il fenomeno segregazione ovoplasmatica.
Il risultato principale del periodo di schiacciamento è la trasformazione di un embrione unicellulare - uno zigote - in multicellulare E singolo strato embrione - blastula. Inoltre, durante la frantumazione, la formazione di
ing contatti intercellulari e un aumento della loro area, la distribuzione di varie sezioni del citoplasma zigote tra diversi blastomeri dell'embrione, l'attivazione graduale del genoma dell'embrione, il ripristino del rapporto nucleo-citoplasmatico caratteristico delle cellule somatiche. Nella fase di frantumazione, tutte le cellule dell'embrione sono omogenee rispetto allo stato funzionale dell'apparato genetico. La differenza nel citoplasma dei blastomeri e le interazioni intercellulari determinano (predeterminano) la direzione dell'ulteriore sviluppo cellulare.
7.4.2. GASTRULAZIONE
7.4.2.1. L'essenza dello stadio della gastrulazione
L'essenza della fase di gastrulazione è che un embrione a strato singolo - la blastula - si trasforma in multistrato- due- O a tre strati, chiamato gastrula(dal greco. gaster- stomaco e suffisso diminutivo "st").
Nei cordati primitivi, ad esempio, nella lancetta, un blastoderma monostrato omogeneo durante la gastrulazione si trasforma in uno strato germinale esterno - ectoderma- e lo strato germinale interno - endoderma. L'endoderma forma l'intestino primario con una cavità all'interno - gastrocele. Viene chiamato il foro che porta al gastrocele blastopore o bocca primaria. Due strati germinali stanno definendo caratteristica morfologica gastrulazione. La loro esistenza ad un certo stadio di sviluppo in tutti gli animali pluricellulari, dai celenterati ai vertebrati superiori, permette di pensare all'omologia degli strati germinali e all'unità dell'origine di tutti questi animali.
Nei vertebrati, oltre ai due strati germinali menzionati, un terzo si forma durante la gastrulazione - mesoderma, situato tra l'ecto- e l'endoderma. Lo sviluppo dello strato germinale medio, che è cordomesoderma,- complicazione evolutiva della fase di gastrulazione nei vertebrati, associata all'accelerazione del loro sviluppo nelle prime fasi dell'embriogenesi. Nei cordati più primitivi, come la lancetta, il cordomesoderma di solito si forma all'inizio della fase che segue la gastrulazione - organogenesi. Lo spostamento nel tempo di sviluppo di alcuni organi rispetto ad altri nei discendenti rispetto ai gruppi ancestrali è una manifestazione di eterocronia. I cambiamenti nei tempi della formazione degli organi più importanti nel corso dell'evoluzione sono abbastanza comuni.
Il processo di gastrulazione è caratterizzato importanti trasformazioni cellulari, come i movimenti diretti di gruppi e singole cellule, la propagazione selettiva e l'ordinamento delle cellule, l'inizio della citodifferenziazione e le interazioni di induzione. I meccanismi cellulari elencati di ontogenesi sono discussi in dettaglio nella sezione 8.2.
I metodi di gastrulazione sono diversi. Si distinguono quattro tipi di movimenti cellulari diretti spazialmente, che portano alla trasformazione dell'embrione da un singolo strato a un multistrato (due o tre strati).
Intussuscezione- invaginazione di una delle sezioni del blastoderma verso l'interno come un intero strato. Nella lancetta, le cellule del polo vegetativo si invaginano; negli anfibi, l'intussuscezione si verifica al confine tra il polo animale e quello vegetativo nella regione della mezzaluna grigia. Il processo di invaginazione è possibile solo nelle uova con una piccola o media quantità di tuorlo.
epibolia- fouling con piccole cellule in rapida divisione del polo animale di cellule più grandi, in ritardo nel tasso di divisione e meno mobili del polo vegetativo. Questo processo è chiaramente espresso negli anfibi.
Delaminazione- stratificazione delle cellule di blastoderma in due strati sovrapposti. La delaminazione può essere osservata nella discoblastula degli embrioni con schiacciamento incompleto di animali come rettili, uccelli e mammiferi ovipari. La delaminazione si manifesta nell'embrioblasto dei mammiferi placentari, portando alla formazione di ipoblasto ed epiblasto.
Immigrazione- movimento di gruppi o singole celle che non sono unite in un unico strato. L'immigrazione si verifica in tutti gli embrioni, ma è più caratteristica della seconda fase della gastrulazione nei vertebrati superiori (rettili, uccelli e mammiferi).
In ogni caso specifico di embriogenesi, di norma, vengono combinati diversi metodi di gastrulazione.
7.4.2.2. Morfologia della gastrulazione
Un esame più dettagliato della gastrulazione nella lancetta, nella rana, nel pollo e nei mammiferi aiuterà a comprendere meglio le relazioni evolutive e comprendere i modelli di sviluppo individuale.
7.4.2.2-a. Gaetrulazione della lancetta
gastrulazione lancetta mostrato in fig. 7.5. Diversi marcatori allo stadio di blastula (Fig. 7.5, a) contrassegnati presuntivo(presunto) rudimenti. Queste sono aree della blastula, dal materiale cellulare
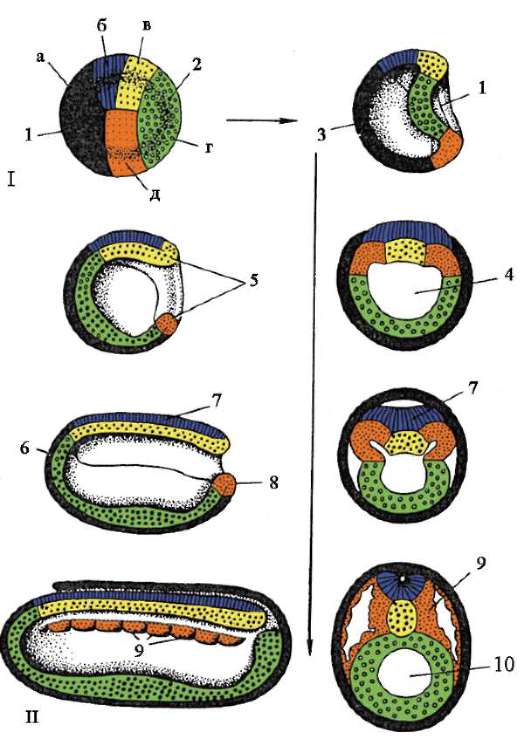
Riso. 7.5.Presunti rudimenti, gastrulazione e neurulazione in una lancetta. I - presunti rudimenti allo stadio di blastula (vista esterna) e gastrula precoce (vista in sezione); II - gastrula tardiva e neurulazione sulle sezioni sagittale (fila sinistra) e trasversale (fila destra); III - modello plastico dell'embrione alla fine del periodo di neurulazione:
1 - palo animale; 2 - polo vegetativo; 3 - blastocele; 4 - gastrogol; 5 - labbra dorsali e ventrali del blastoporo; 6 - l'estremità della testa dell'embrione; 7 - placca midollare; 8 - coda dell'embrione; 9 - parte dorsale del mesoderma; 10 - cavità dell'intestino secondario; 11 - somiti segmentati; 12 - parte addominale del mesoderma; a, b, c, d, e - designazioni di organi presunti e in via di sviluppo: a - ectoderma cutaneo; b - tubo neurale; in - accordo; d - endoderma, epitelio intestinale; d - mesoderma
di cui, durante la gastrulazione e l'organogenesi precoce (neurulazione), si formano solitamente strati germinali e organi completamente definiti (Fig. 7.5, bec).
L'invaginazione inizia al polo vegetativo. A causa della divisione più rapida, le cellule del polo animale crescono e spingono le cellule del polo vegetativo nella blastula. Ciò è facilitato da un cambiamento nello stato del citoplasma nelle cellule che formano le labbra del blastopore e ad esse adiacenti. A causa dell'invaginazione, il blastocele diminuisce e il gastrocele aumenta. Contemporaneamente alla scomparsa del blastocele, l'ectoderma e l'endoderma entrano in stretto contatto. La lancetta, come tutti gli altri deuterostomi(questi includono il phylum Echinodermata, il phylum Chordata e alcuni altri tipi di animali relativamente piccoli), la regione del blastopore si trasforma nella parte della coda dell'organismo, a differenza dei protostomi, in cui il blastopore corrisponde alla parte della testa. Apertura della bocca nei deuterostomi formata all'estremità dell'embrione di fronte al blastopore.
7.4.2.2-6. Gaetrulazione negli anfibi
La gastrulazione negli anfibi ha molto in comune con la gastrulazione della lancetta, ma poiché il tuorlo nelle loro uova è molto più grande e si trova principalmente al polo vegetativo, i grandi blastomeri dell'anfiblastula non sono in grado di gonfiarsi verso l'interno. Intussuscezione va un po' diversamente. Al confine tra i poli animale e vegetativo nella regione della falce grigia, le cellule prima si allungano fortemente verso l'interno, assumendo la forma di "a forma di fiasco" (Fig. 7.6, a), quindi tirano le cellule dello strato superficiale della blastula insieme a loro. Compaiono un solco a forma di mezzaluna e un labbro dorsale (dorsale) del blastopore (Fig. 7.6, b).
Allo stesso tempo, le cellule più piccole del polo animale, dividendosi più velocemente, iniziano a muoversi verso il polo vegetativo. Nella regione del labbro dorsale, si alzano e si invaginano, e cellule più grandi crescono sui lati e sul lato opposto al solco a forma di falce. Poi il processo epibolia porta alla formazione delle labbra laterali (laterali) e ventrali (ventrali) del blastopore. Il blastoporo si chiude ad anello, all'interno del quale sono visibili da tempo grandi cellule luminose del polo vegetativo sotto forma delle cosiddette tappo di tuorlo. Successivamente, sono completamente immersi verso l'interno e il blastopore si restringe.
Utilizzando il metodo di marcatura con coloranti vitali (vitali) negli anfibi, i movimenti cellulari sono stati studiati in dettaglio
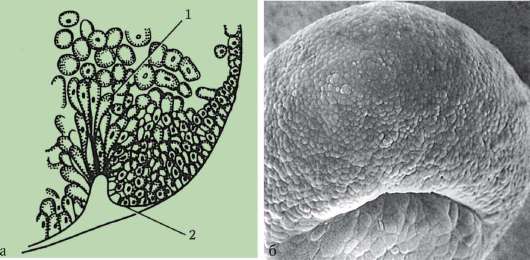
Riso. 7.6.Formazione del blastopore della prima gastrula degli anfibi: a - cellule a forma di fiasco nella regione del labbro dorsale del blastoporo; b - vista generale del solco falciforme e del labbro dorsale; 1 - celle a forma di pallone; 2 - labbro dorsale del blastoporo
blastula durante la gastrulazione, è stato riscontrato che aree specifiche del blastoderma, chiamate presuntivo(dal lat. praesumptio- assunzione), durante il normale sviluppo, compaiono prima nella composizione di alcuni rudimenti di organi, e poi nella composizione degli organi stessi (Fig. 7.7). È noto che negli anfibi senza coda, il materiale della presunta notocorda e mesoderma allo stadio di blastula non si trova sulla sua superficie, ma negli strati interni della parete dell'anfiblastula, tuttavia, approssimativamente agli stessi livelli mostrati nella figura.
Una certa somiglianza tra i processi di gastrulazione e le aree degli organi presunti negli anfibi e nella lancetta, ad es. l'omologia degli organi principali, come il tubo neurale, la notocorda, l'intestino secondario, indica la loro relazione filogenetica.
7.4.2.2- c. Gastrulazione negli uccelli
La gastrulazione negli embrioni con un tipo meroblastico di scissione e sviluppo ha le sue caratteristiche. A uccelli inizia dopo la scissione e la formazione della blastula durante il passaggio dell'embrione attraverso l'ovidotto. Quando l'uovo viene deposto, l'embrione è già costituito da diversi strati: viene chiamato lo strato superiore epiblasto, inferiore - ipoblastoma primitivo(vedi Fig. 7.4, c). Tra di loro c'è uno stretto spazio: il blastocele. Quindi formato ipoblasto secondario, la cui modalità di formazione non è del tutto chiara. Ci sono prove che nelle primarie
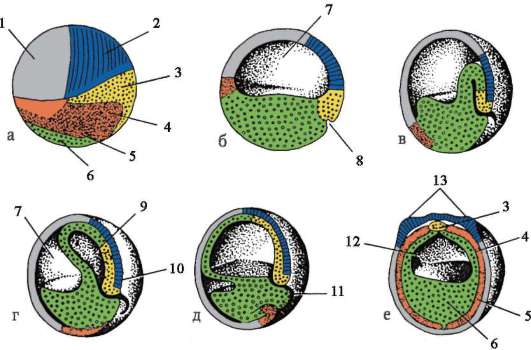
Riso. 7.7. Mappa delle aree di presunti rudimenti di organi nelle prime fasi dello sviluppo embrionale dell'anfibio: a - stadio di blastula (vista a sinistra); b-e - fasi successive della gastrulazione (sezioni sagittali); e - l'inizio della neurulazione (sezione trasversale); 1 - ectoderma cutaneo; 2 - tubo neurale; 3 - accordo; 4 - mesoderma somitico; 5 - mesoderma di splancnotomi; 6 - endoderma; 7 - blastocele; 8 - solco a mezzaluna; 9 - gastrocele; 10 - blastopore labiale dorsale; 11 - tappo di tuorlo; 12 - cavità dell'intestino secondario; 13 - rulli nervosi
Nell'area degli uccelli hanno origine le cellule germinali primarie e il secondario forma un endoderma extra-embrionale. La formazione dell'ipoblasto primario e secondario è considerata un fenomeno che precede la gastrulazione.
I principali eventi di gastrulazione e la formazione finale dei tre strati germinali iniziano dopo l'ovodeposizione con l'inizio dell'incubazione. C'è un accumulo di cellule nella parte posteriore dell'epiblasto a causa della velocità irregolare della divisione cellulare e del loro movimento dalle parti laterali dell'epiblasto al centro, l'una verso l'altra. Il cosidetto linea primaria, che si estende verso l'estremità della testa. Al centro della striscia primaria si forma solco primario, e lungo i bordi rotoli primari. Appare un ispessimento all'estremità di testa della striscia primaria - Nodo di Hensen, e in esso - fossa primaria(figura 7.8).
Quando le cellule epiblastiche entrano nel solco primario, la loro forma cambia. Assomigliano nella forma alle cellule "a forma di fiasco" della gastrula degli anfibi. Queste cellule diventano quindi stellate e affondano sotto l'epiblasto per formare il mesoderma. Una parte delle cellule in migrazione, incorporandosi nell'ipoblasto, dà successivamente origine all'endoderma germinale (Fig. 7.9). Pertanto, l'endoderma si forma sulla base dell'ipoblasto primario e secondario con l'aggiunta di una nuova generazione di cellule endodermiche che migrano dagli strati superiori del blastoderma. La presenza di diverse generazioni di cellule endodermiche indica il prolungamento nel tempo del periodo di gastrulazione.
Parte delle cellule che migrano dall'epiblasto attraverso il nodo di Hensen formano la futura notocorda. Contemporaneamente all'anlage e all'allungamento della corda, il nodo di Hensen e la striscia primaria vengono gradualmente spostati nella direzione dall'estremità anteriore a quella caudale e scompaiono. Ciò corrisponde al restringimento e alla chiusura del blastopore. Quando la striscia primaria si contrae, lascia dietro di sé le sezioni formate degli organi assiali dell'embrione nella direzione dalla testa alle sezioni della coda. Sembra ragionevole considerare i movimenti cellulari nell'embrione di pollo come omologhi a quelli negli anfibi, e la striscia primaria e il nodo di Hensen come omologhi al blastopore e al labbro dorsale (dorsale) della gastrula di questi animali.
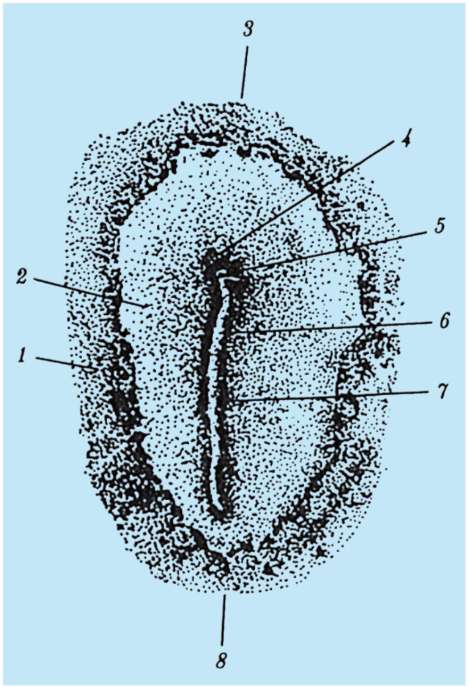
Riso. 7.8.Embrione di pollo allo stadio della striscia primaria (vista dorsale): 1 - area scura; 2 - area traslucida del disco germinale; 3 - testata; 4 - Nodo di Hensen; 5 - fossa primaria; 6 - scanalatura primaria; 7 - rullo primario; 8 - coda
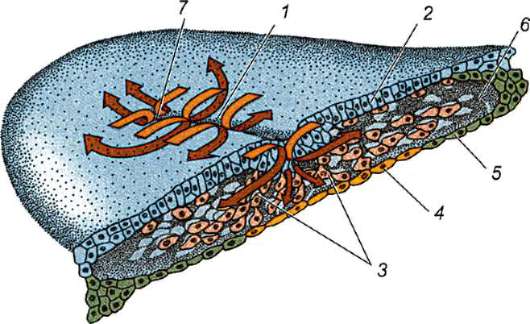
Riso. 7.9.Embrione di pollo nella fase della striscia primaria (sezione trasversale): 1 - solco primario; 2 - epiblasto (ectoderma); 3 - cellule migratorie (mesoderma); 4 - endoderma germinale; 5 - ipoblasto (endoderma); 6 - blastocele; 7 - Nodo di Hensen
7.4.2.2-d. Gastrulazione nei mammiferi
È interessante notare che le cellule degli embrioni di mammifero (vedi paragrafo 7.5.1), nonostante in questi animali le uova abbiano una piccola quantità di tuorlo, e la frammentazione sia completa, nella fase di gastrulazione conservano i movimenti caratteristici di gli embrioni di rettili e uccelli. Ciò conferma l'idea dell'origine dei mammiferi da un gruppo ancestrale le cui uova erano ricche di tuorlo.
7.4.2.3. Caratteristiche dello stadio della gastrulazione
La gastrulazione è caratterizzata da una varietà di processi cellulari. Il mitotico continua riproduzione cellulare, inoltre, ha intensità diversa nelle diverse parti dell'embrione. Le interfasi dei cicli mitotici includono tutti i periodi (SOL 1 , S, G 2), quindi, a partire dallo stadio di gastrulazione, si osserva la crescita dell'organismo in via di sviluppo. Tuttavia, il più caratteristica la gastrulazione consiste in movimento delle masse cellulari. Ciò porta a un cambiamento nella struttura dell'embrione e alla sua trasformazione da blastula a gastrula. in corso ordinamento cellule per la loro appartenenza a diversi strati germinali, all'interno dei quali si "riconoscono" a vicenda.
Inizia la fase di gastrulazione differenziazione cellule, il che significa una transizione all'uso attivo delle informazioni biologiche del proprio genoma. Uno dei regolatori dell'attività genetica è la diversa composizione chimica del citoplasma delle cellule embrionali, che si instaura per segregazione ovo(oo)plasmatica, entrambi originati dal polo vegetativo dell'uovo.
Durante la gastrulazione, il ruolo è molto ampio induzione embrionale- interazioni tra complessi cellulari (parti) dell'embrione in via di sviluppo. È stato dimostrato che la comparsa della striscia primaria negli uccelli è il risultato di un'interazione induttiva tra l'ipoblasto e l'epiblasto. L'ipoblasto ha polarità. Un cambiamento nella posizione dell'ipoblasto rispetto all'epiblasto provoca un cambiamento nell'orientamento della striscia primitiva. Tutti questi processi sono descritti in dettaglio nel capitolo 8.
7.4.3. FORMAZIONE DI ORGANI E TESSUTI 7.4.3.1. L'essenza dello stadio dell'organogenesi
organogenesi, consistenti nella formazione di singoli organi, costituiscono il contenuto principale del periodo embrionale. Continuano nel periodo larvale e terminano nel periodo giovanile. L'organogenesi si distingue per le trasformazioni morfogenetiche (formanti) più complesse e diverse. Un prerequisito necessario per il passaggio all'organogenesi è il raggiungimento da parte dell'embrione dello stadio della gastrula, ovvero la formazione degli strati germinali. Occupando una certa posizione l'uno rispetto all'altro, gli strati germinali, entrando in contatto e interagendo, forniscono tali relazioni tra diversi gruppi cellulari che stimolano il loro sviluppo in una certa direzione. Questo cosiddetto induzione embrionale- la conseguenza più importante dell'interazione tra gli strati germinali.
Nel corso dell'organogenesi, la forma, la struttura e la composizione chimica delle cellule cambiano, i gruppi cellulari vengono isolati, che sono i rudimenti degli organi futuri. Una certa forma di organi si sviluppa gradualmente, vengono stabilite connessioni spaziali e funzionali tra di loro. I processi di morfogenesi sono accompagnati da differenziazione
differenziazione cellulare e formazione dei tessuti, differenziazione di tessuti e strutture, nonché la loro crescita selettiva e irregolare. Un prerequisito per l'organogenesi, insieme alla riproduzione cellulare, alla migrazione e allo smistamento, è la loro morte selettiva (vedi Capitolo 8).
7.4.3.2. Neurulazione
L'inizio dell'organogenesi è chiamato neurulazione. Neurulazione copre i processi dalla comparsa dei primi segni della formazione della placca neurale alla sua chiusura nel tubo neurale (Fig. 7.10). Allo stesso tempo, accordo E intestino secondario, e il mesoderma che giace ai lati della corda si divide nella direzione craniocaudale in strutture accoppiate segmentate - somiti(si forma un complesso di organi assiali - organogenesi primaria).
Il sistema nervoso dei vertebrati, compreso l'uomo, è caratterizzato dalla stabilità del piano strutturale principale in tutto storia evolutiva sottotipo. Nella formazione del tubo neurale, tutti i cordati hanno molto in comune. Inizialmente non specializzato ectoderma dorsale, rispondendo all'effetto di induzione del cordomesoderma, si trasforma in piastra neurale, presentato neuroepiteliale cellule cilindriche.
La placca neurale non rimane appiattita a lungo. Presto, i suoi bordi laterali si alzano, formandosi pieghe nervose, che giacciono su entrambi i lati di un longitudinale poco profondo solco neurale. I bordi delle pieghe neurali poi si chiudono, formando un tubo neurale chiuso con un canale all'interno - neurocele. Innanzitutto, la chiusura delle pieghe neurali avviene a livello dell'inizio (il confine tra midollo spinale e cervello) del midollo spinale, per poi diffondersi al cervello e alla coda.
Riso. 7.10. Fasi successive nella formazione del tubo neurale e della cresta neurale (sezione trasversale dell'embrione): 1 - placca neurale; 2 - cresta neurale; 3 - ectoderma; 4 - accordo; 5 - solco nervoso; 6 - neurocele; 7 - pieghe nervose
indicazioni. È stato dimostrato che i microtubuli ei microfilamenti delle cellule neuroepiteliali svolgono un ruolo importante nella morfogenesi del tubo neurale. Distruzione di queste strutture cellulari da parte di colchicina e citocalasina IN fa sì che la placca neurale rimanga aperta. La mancata chiusura delle pieghe neurali porta a malformazioni congenite del tubo neurale.
Dopo la chiusura delle pieghe neurali, si formano le cellule che originariamente si trovavano tra la placca neurale e il futuro ectoderma cutaneo cresta neurale. Le cellule della cresta neurale si distinguono per la loro capacità di migrazioni estese, ma strettamente regolate in tutto il corpo (vedi sezione 8.2.2, Fig. 8.9) e formano due flussi principali. Le cellule di uno di loro - superficiale- sono inclusi nell'epidermide o nel derma della pelle, dove si differenziano in cellule pigmentate. Un altro flusso migra nella direzione ventrale e forma gangli spinali sensibili, gangli simpatici, midollo surrenale, gangli parasimpatici. Le cellule della cresta neurale cranica danno origine sia alle cellule nervose che a una serie di altre strutture, come la cartilagine branchiale, alcune ossa tegumentarie del cranio.
L'endoderma in tutti gli embrioni forma infine l'epitelio dell'intestino secondario e molti dei suoi derivati. L'intestino secondario stesso si trova sempre sotto l'accordo.
Quindi, nel processo di neurulazione, un complesso organi assiali: tubo neurale, corda, intestino, che sono la caratteristica più caratteristica dell'organizzazione del corpo di tutti i cordati. La stessa origine, lo sviluppo e la disposizione reciproca degli organi assiali rivelano la loro completa omologia e continuità evolutiva.
Un esame approfondito e un confronto dei processi di neurulazione in specifici rappresentanti del tipo cordato rivela alcune differenze che sono principalmente associate a caratteristiche che dipendono dalla struttura delle uova, dal metodo di schiacciamento e gastrulazione (Fig. 7.11). Si richiama l'attenzione sulla diversa forma degli embrioni e sullo spostamento nel tempo di deposizione degli organi assiali l'uno rispetto all'altro, ad es. eterocronia sopra descritta.
7.4.3.3. differenziazione mesodermica
Il mesoderma, che occupa un posto ai lati della notocorda e si estende ulteriormente tra l'ectoderma cutaneo e l'endoderma dell'intestino secondario, è suddiviso in regioni dorsale e ventrale. Dorsale
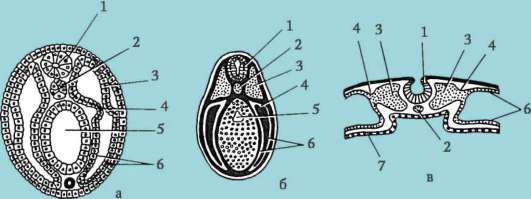
Riso. 7.11. Neurule di vari cordati: a - lancetta; b - rana; c - pollo; 1 - tubo neurale; 2 - accordo; 3 - somitico; 4 - gamba somitica; 5 - intestino secondario; 6 - piastra laterale; 7 - endoderma
la parte è segmentata e presentata in coppia somiti. La deposizione dei somiti va dalla testa alla coda. La parte ventrale non segmentata del mesoderma, che assomiglia a un sottile strato di cellule, è chiamata piatto di contorno. I somiti sono collegati alla piastra laterale dal mesoderma intermedio sotto forma di segmentato gambe somite.
Tutte le aree del mesoderma si differenziano gradualmente. All'inizio della formazione, i somiti hanno una configurazione caratteristica di un epitelio con una cavità all'interno. Sotto l'effetto di induzione proveniente dalla corda e dal tubo neurale, le parti ventromediali dei somiti... sclerotomi- si trasformano in mesenchima secondario, vengono sfrattati dal somite e circondano la notocorda e la parte ventrale del tubo neurale. Alla fine, da loro si formano vertebre, costole e scapole.
La parte dorsolaterale dei somiti si differenzia in due direzioni. Aree di somiti rivolte verso la forma ectodermica tegumentaria dermatomi, che danno origine allo strato interno della pelle - il derma. Le cellule situate sul lato interno della parte dorsolaterale dei somiti si formano miotomi, da cui si svilupperanno i muscoli scheletrici striati del corpo e degli arti. Dalla regione delle gambe dei somiti con rudimenti nefrotomo E gonotomo si formano organi escretori e ghiandole sessuali.
Placche laterali non segmentate divise in due foglie, che limitano la cavità del corpo secondario - generalmente. foglio interno, adiacente all'endoderma è chiamato viscerale. Circonda l'intestino da tutti i lati e forma il mesentere, copre il polmone
parenchima e muscolo cardiaco. Foglia esterna la placca laterale è adiacente all'ectoderma ed è chiamata parietale. In futuro, forma i fogli esterni del peritoneo, della pleura e del pericardio.
7.4.3.4. Derivati degli strati germinali
Ectoderma, mesoderma ed endoderma nel corso dell'ulteriore sviluppo, interagendo tra loro, partecipano alla formazione di organi specifici (organogenesi privata o locale). L'emergere del rudimento di un organo è associato a cambiamenti locali in una certa area del corrispondente strato germinale. COSÌ, dall'ectoderma si sviluppano l'epidermide della pelle e dei suoi derivati (piuma, capelli, unghie, pelle e ghiandole mammarie), componenti degli organi della vista, dell'udito, dell'olfatto, dell'epitelio del cavo orale e dello smalto dei denti. I derivati ectodermici più importanti sono il tubo neurale, la cresta neurale e tutte le strutture neurali da essi formate.
Derivati endodermici sono l'epitelio dello stomaco e dell'intestino, le cellule del fegato, le cellule secernenti del pancreas, le ghiandole intestinali e gastriche. L'endoderma della parte anteriore dell'intestino embrionale forma l'epitelio dei polmoni e delle vie aeree, così come le cellule secernenti dei lobi anteriore e medio dell'ipofisi, della tiroide e delle ghiandole paratiroidi.
mesoderma, oltre alle strutture scheletriche già descritte sopra, i muscoli scheletrici, il derma della pelle, gli organi dell'apparato escretore e riproduttivo, forma il sistema cardiovascolare, il sistema linfatico, la pleura, il peritoneo e il pericardio. Da mesenchima, avendo un'origine mista dovuta alle cellule dei tre foglietti germinativi, si sviluppano tutti i tipi di tessuto connettivo, muscoli lisci, sangue e linfa (per maggiori dettagli si veda il paragrafo 8.2.5).
Il rudimento di un particolare organo è inizialmente formato da uno specifico strato germinale, ma poi l'organo diventa più complesso e, di conseguenza, due o tre strati germinali prendono parte alla sua formazione.
7.4.4. ORGANI PROVVISORI DELL'EMBRIONE DEI VERTEBRATI
provvisorio, O temporaneo, gli organi si formano nell'embriogenesi di un numero di rappresentanti dei vertebrati per garantire le funzioni vitali dell'embrione, come la respirazione, la nutrizione, l'escrezione, il movimento e altri. Gli organi sottosviluppati dell'animale più in via di sviluppo non sono ancora in grado di funzionare come previsto, anche se devono
svolgere un ruolo nel sistema di un intero organismo in via di sviluppo. Non appena l'embrione raggiunge il necessario grado di maturità, quando la maggior parte degli organi è in grado di svolgere funzioni vitali, gli organi temporanei vengono riassorbiti o scartati.
Il tempo di formazione degli organi provvisori dipende da quali riserve di nutrienti sono state accumulate nell'uovo e in quali condizioni ambientali si sviluppa l'embrione. Negli anfibi senza coda, ad esempio, a causa della sufficiente quantità di tuorlo nell'uovo e del fatto che lo sviluppo avviene in acqua, l'embrione effettua scambi gassosi e rilascia prodotti di dissimilazione direttamente attraverso le membrane dell'uovo e raggiunge lo stadio di girino. In questa fase si formano gli organi provvisori della respirazione (branchie), della digestione e del movimento adattati allo stile di vita acquatico. Gli organi larvali elencati consentono al girino di continuare il suo sviluppo. Al raggiungimento dello stato di maturità morfologica e funzionale degli organi del tipo adulto, gli organi temporanei scompaiono nel processo di metamorfosi.
Rettili e uccelli hanno più riserve di tuorlo nell'uovo, ma l'embrione non si sviluppa nell'acqua, ma sulla terra. A questo proposito, molto presto è necessario garantire la respirazione e l'escrezione, nonché la protezione dall'essiccamento. In essi già all'inizio dell'embriogenesi, quasi parallelamente alla neurulazione, inizia la formazione di organi provvisori, come amnio, corion E sacco vitellino. Poco dopo si forma l'allantois. Nei mammiferi placentari, questi stessi organi provvisori si formano anche prima, poiché nell'uovo c'è pochissimo tuorlo. Lo sviluppo di tali animali avviene in utero, la formazione di organi provvisori in essi coincide nel tempo con il periodo di gastrulazione.
La presenza o l'assenza dell'amnion e di altri organi provvisori è alla base della divisione dei vertebrati in due gruppi: Amniota E Anamnia(vedi tabella 7.1). I vertebrati evolutivamente più vecchi che si sviluppano esclusivamente nell'ambiente acquatico e sono rappresentati da classi come ciclostomi, pesci e anfibi non hanno bisogno di acqua aggiuntiva e altri gusci dell'embrione e costituiscono il gruppo anamnium. Il gruppo degli amnioti comprende i vertebrati terrestri primari, cioè quelli il cui sviluppo embrionale avviene in condizioni terrestri. Queste sono le tre classi: Rettili, Uccelli e Mammiferi. Appartengono ai vertebrati superiori, poiché hanno sistemi di organi coordinati e altamente efficienti che ne assicurano l'esistenza nella maggior parte condizioni difficili quali sono le condizioni del terreno. Questi
le classi includono un gran numero di specie che sono passate secondariamente nell'ambiente acquatico. Pertanto, i vertebrati superiori sono stati in grado di dominare tutti gli habitat. Tale perfezione sarebbe impossibile, anche senza la fecondazione interna e la formazione di speciali organi embrionali provvisori, detti anche membrane embrionali. Le membrane amniotiche comprendono l'amnion, il corion (sierosa), il sacco vitellino e l'allantoide.
C'è molto in comune nella struttura e nelle funzioni degli organi provvisori di vari amnioti. Descrivendo nei termini più generali gli organi provvisori degli embrioni dei vertebrati superiori, va notato che essi si sviluppano tutti dal materiale cellulare di strati germinali già formati. Ci sono alcune caratteristiche nello sviluppo delle membrane embrionali dei mammiferi placentari, che saranno discusse di seguito.
Amnioè un sacco contenente l'embrione e riempito di liquido amniotico. La membrana amniotica è formata da ectoderma extraembrionale e somatopleura. La sua parte ectodermica è specializzata per la secrezione e l'assorbimento del liquido amniotico che bagna l'embrione. L'amnione svolge un ruolo primario nel proteggere l'embrione dall'essiccamento e dai danni meccanici, creando per esso l'ambiente acquatico più favorevole e naturale. La parte mesodermica dell'amnion dà origine a fibre muscolari lisce. Le contrazioni di questi muscoli fanno pulsare l'amnione ei lenti movimenti oscillatori comunicati all'embrione in questo processo apparentemente aiutano a garantire che le sue parti in crescita non interferiscano l'una con l'altra.
Corion (sierosa)- la membrana germinale più esterna adiacente al guscio (sierosa) o ai tessuti materni (corion), originata, come l'amnion, dall'ectoderma e dalla somatopleura. Questo guscio serve per lo scambio tra l'embrione e l'ambiente. Nelle specie ovipare, la funzione principale della sierosa è la partecipazione alla respirazione (scambio gassoso); nei mammiferi, il corion svolge funzioni molto più estese, partecipando, oltre alla respirazione, alla nutrizione, all'escrezione, alla filtrazione e alla sintesi di sostanze, come gli ormoni.
Sacco vitellino formato dall'endoderma extraembrionale e dal mesoderma viscerale ed è direttamente collegato al tubo intestinale dell'embrione. Negli embrioni con una grande quantità di tuorlo, prende parte alla nutrizione. Negli uccelli, ad esempio, si sviluppa un sistema vascolare nello splanchnopleuron del sacco vitellino. Il tuorlo non passa attraverso il dotto vitellino, che collega il sacco all'intestino. Viene prima tradotto in
forma solubile sotto l'azione degli enzimi digestivi prodotti dalle cellule endodermiche della parete del sacco. Quindi entra nei vasi e si diffonde con il sangue in tutto il corpo dell'embrione.
I mammiferi non hanno riserve di tuorlo e la conservazione del sacco vitellino può essere associata a importanti funzioni secondarie. Il mesoderma del sacco vitellino produce le cellule del sangue dell'embrione. Inoltre, il sacco vitellino dei mammiferi è riempito con un fluido ad alta concentrazione di aminoacidi e glucosio, che indica la possibilità del metabolismo proteico nel sacco vitellino.
Il destino del sacco vitellino in diversi animali è leggermente diverso. Negli uccelli, entro la fine del periodo di incubazione, i resti del sacco vitellino sono già all'interno dell'embrione, dopodiché scompare rapidamente e si risolve completamente entro la fine dei 6 giorni dopo la schiusa. Nei mammiferi, il sacco vitellino si sviluppa in modi diversi. Nei predatori è relativamente grande, con una rete di vasi molto sviluppata, mentre nei primati si restringe rapidamente e scompare senza lasciare traccia prima del parto.
Allantois si sviluppa un po' più tardi rispetto ad altri organi provvisori. È una conseguenza simile a un sacco della parete ventrale dell'intestino posteriore. Pertanto, è formato dall'endoderma all'interno e dalla splancnopleura all'esterno. Nei rettili e negli uccelli, l'allantoide cresce rapidamente fino al corion e svolge diverse funzioni. Prima di tutto, è un serbatoio per l'urea e l'acido urico, che sono i prodotti finali del metabolismo contenente azoto. materia organica. L'allantoide ha una rete vascolare ben sviluppata, grazie alla quale, insieme al corion, partecipa allo scambio di gas. Alla schiusa la parte esterna dell'allantoide viene scartata, mentre la parte interna si conserva come vescica.
In molti mammiferi anche l'allantoide è ben sviluppato e, insieme al corion, forma la placenta corioallantoidea. Termine placenta significa stretta sovrapposizione o fusione delle membrane germinali con i tessuti dell'organismo genitore. Nei primati e in alcuni altri mammiferi, la parte endodermica dell'allantoide è rudimentale e le cellule mesodermiche formano un denso cordone che si estende dalla regione cloacale al corion. I vasi crescono lungo il mesoderma allantoide fino al corion, attraverso il quale la placenta svolge funzioni escretorie, respiratorie e nutrizionali.
È più accessibile e più facile studiare la formazione e la struttura delle membrane embrionali usando l'esempio di un embrione di pollo. Allo stadio neurula, tre strati germinali passano direttamente dall'embrione all'extra-
parte respirante, senza delimitare in alcun modo. Man mano che l'embrione prende forma, attorno ad esso si formano diverse pieghe che, per così dire, tagliano l'embrione, lo separano dal tuorlo e stabiliscono confini netti tra l'embrione e le aree extraembrionali. Si chiamano pieghe del tronco(figura 7.12).
Innanzitutto, si forma una piega della testa. Taglia la parte della testa dal basso. Le estremità posteriori di questa piega passano nelle pieghe laterali del tronco, che delimitano il corpo dell'embrione dai lati. La piega della coda delimita l'estremità posteriore dell'embrione. Restringe gradualmente la gamba che collega l'intestino medio e il sacco vitellino, formato
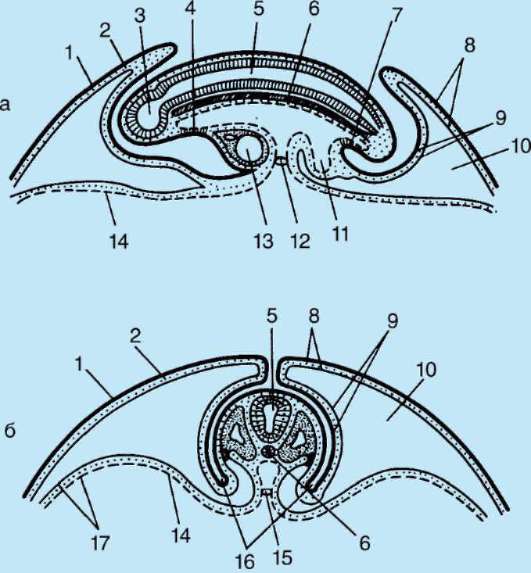
Riso. 7.12. Formazione di pieghe del tronco e membrane embrionali in un embrione di pollo: a - sezione longitudinale; b - sezione trasversale; 1 - ectoderma; 2 - mesoderma; 3 - il rudimento del cervello; 4 - membrana faringea; 5 - tubo neurale; 6 - accordo; 7 - membrana cloacale; 8 - corion; 9 - amnio; 10 - esocele; 11 - allantois; 12 - area dell'ombelico; 13 - il rudimento del cuore; 14 - endoderma; 15 - segnalibro dell'intestino; 16 - pieghe del busto; 17 - sacco vitellino
intestino anteriore e posteriore. Allo stesso tempo, dall'ectoderma e dalla somatopleura ad esso adiacente, si forma prima una piega della testa (Fig. 7.13), che, come un cappuccio, cresce dall'alto sull'embrione. Le estremità della testa si piegano sui lati pieghe amniotiche. Crescono sopra l'embrione l'uno verso l'altro e crescono insieme, formando immediatamente le pareti dell'amnion, adiacente all'embrione, e il corion, che si trova all'esterno.
Successivamente si forma l'allantoide (Fig. 7.14). La vista generale dell'embrione di pollo al sesto giorno di incubazione è mostrata in Fig. 7.15. In vari mammiferi, i processi di formazione degli organi provvisori sono più o meno simili a quelli sopra descritti negli uccelli. Le caratteristiche del loro sviluppo nei primati, compresi gli umani, sono mostrate nel prossimo capitolo.
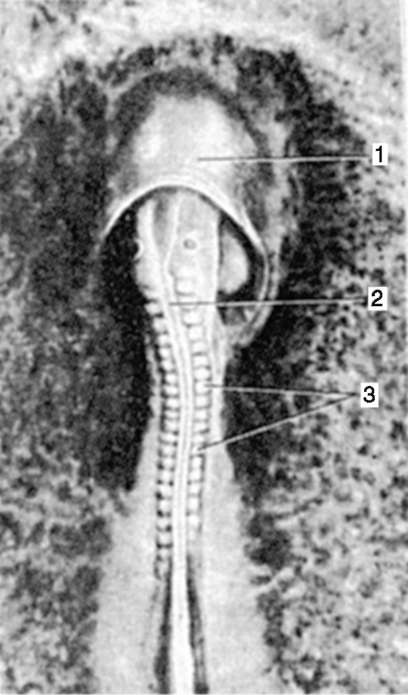
Riso. 7.13.Embrione di pollo per circa 40 ore di incubazione: 1 - piega della testa di amnion; 2 - tubo neurale; 3 - somiti
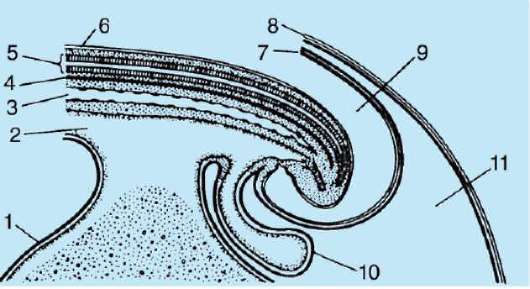
Riso. 7.14.La formazione di allantois in un embrione di pollo (sezione longitudinale della coda): 1 - sacco vitellino; 2 - intestino medio; 3 - aorta; 4 - accordo; 3 - tubo neurale; 6 - ectoderma; 7 - amnio; 8 - corion; 9 - cavità amniotica; 10 - allantois; 11 - esoceloma
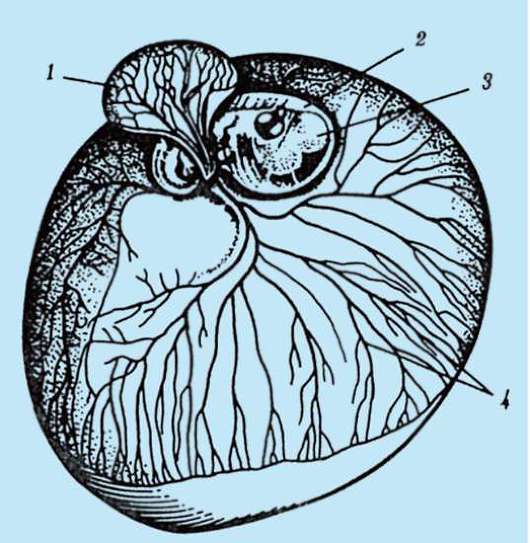
Riso. 7.15. Embrione di pollo al 6° giorno di incubazione (rimozione di proteine e corion, allantois spostato verso l'alto): 1 - allantois; 2 - amnio; 3 - embrione; 4 - vasi del sacco vitellino
7.5. sviluppo embrionale dei mammiferi
7.5.1. PERIODIZZAZIONE E SVILUPPO PRECOCE DELL'EMBRIONE
Lo studio dello sviluppo umano prenatale e, in particolare, embrionale (intrauterino) è molto importante, in quanto aiuta a comprendere meglio la relazione tra organi, nonché i meccanismi delle malformazioni congenite. Nello sviluppo embrionale di diverse specie di mammiferi, ci sono caratteristiche comuni, ma ci sono anche differenze. In tutti i placentali, ad esempio, i processi di embriogenesi precoce differiscono significativamente da quelli precedentemente descritti in altri vertebrati. Allo stesso tempo, ci sono caratteristiche specifiche tra quelle placentari.
Frazionamento Lo zigote umano è caratterizzato dalle seguenti caratteristiche. Il piano della prima divisione passa attraverso i poli dell'uovo, cioè, come in altri vertebrati, è un meridiano. In questo caso, uno dei blastomeri risultanti è più grande dell'altro, il che indica irregolarità schiacciamento. I primi due blastomeri entrano nella divisione successiva in modo asincrono. Il solco corre lungo il meridiano ed è perpendicolare al primo solco. Quindi, c'è uno stadio di tre blastomeri. Durante la divisione del blastomero minore, una coppia di blastomeri minori formati ruota di 90°, in modo che il piano del solco di fissione sia perpendicolare ai primi due
Riso. 7.16.Primi stadi di scissione dello zigote di coniglio: I - piano del primo solco di scissione; IIa - il piano del secondo solco che schiaccia uno dei primi due blastomeri; IIb - piano del secondo solco di scissione del secondo dei primi due blastomeri
solchi. Una disposizione simile dei blastomeri allo stadio di 4 cellule è stata descritta in topi, conigli, visoni e scimmie (Fig. 7.16). A causa della scissione asincrona, potrebbero esserci stadi con un numero dispari di blastomeri - 3, 5, 7, 9, che, a causa dell'assenza di uno stadio di crescita, differiscono per le loro dimensioni. Quindi lo schiacciamento dell'uomo completo asincrono non uniforme.
Come risultato della frantumazione, si forma un accumulo di cellule - morula. I blastomeri localizzati superficialmente formano uno strato cellulare, mentre i blastomeri che si trovano all'interno della morula sono raggruppati in un nodo cellulare centrale. Presto, il liquido appare all'interno della morula, si forma una cavità (blastocele) e l'embrione si trasforma in blastocisti. La formazione di una morula avviene allo stadio di 16 blastomeri e la cavitazione (formazione di una cavità) a partire dallo stadio di 32 blastomeri.
IN blastocisti distinguere lo strato esterno delle cellule (trofoblasto) e la massa cellulare interna (nodulo germinale, o embrioblasto). La massa cellulare interna viene spinta da parte dal fluido verso uno dei poli della blastocisti. Più tardi da trofoblasto si svilupperà la membrana germinale esterna: il corion e da embrioblasto- l'embrione stesso e alcuni organi extraembrionali. È stato dimostrato che l'embrione vero e proprio si sviluppa da un numero molto piccolo di cellule del nodulo germinale.
La fase di frantumazione avviene sotto il guscio lucido. La Figura 7.17 mostra le prime fasi dell'embriogenesi umana, mostrando dove si trova l'embrione nel corpo della madre. La frammentazione dello zigote umano e l'emergenza della blastocisti sono mostrate schematicamente in Fig. 7.18 e 7.19.
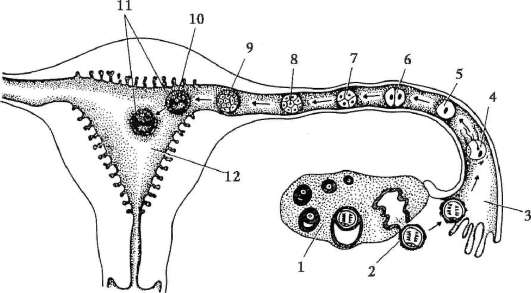
Riso. 7.17.Ovulazione, fecondazione e embrione umano alla 1a settimana di sviluppo: 1 - ovaio; 2 - ovocita II ordine (ovulazione); 3 - ovidotto; 4 - fecondazione; 5 - zigote; 6 - embrione allo stadio di due blastomeri; 7 - embrione allo stadio di quattro blastomeri; 8 - embrione allo stadio di otto blastomeri; 9 - morula; 10, 11 - blastocisti; 12 - parete posteriore dell'utero
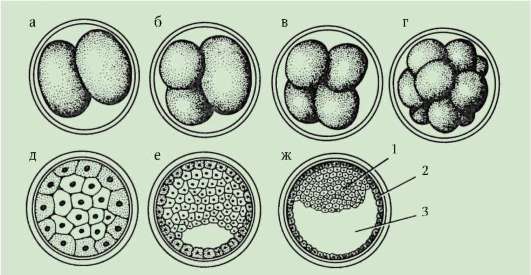
Riso. 7.18.Scissione dello zigote umano: a - due blastomeri; b - tre blastomeri; c - quattro blastomeri; g - morula; e - sezione della morula; f, g - sezione di blastocisti precoce e tardiva: 1 - embrioblasto; 2 - trofoblasto; 3 - blastocele
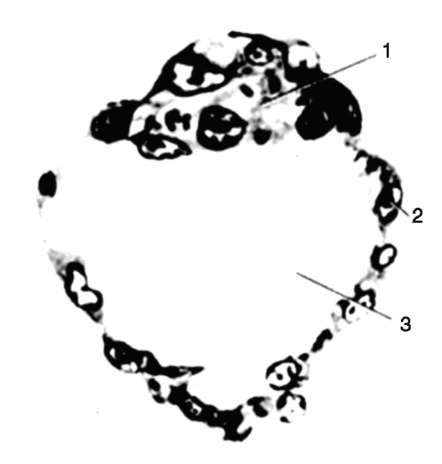
Riso. 7.19.Blastocisti dell'embrione umano (sezione): 1 - embrioblasto; 2 - trofoblasto; 3 - blastocele
Approssimativamente il 6-7° giorno dopo la fecondazione, l'embrione, che galleggia liberamente nella cavità uterina da 2-3 giorni, è pronto per l'impianto, ad es. all'immersione nella sua membrana mucosa. Sotto l'azione degli enzimi secreti dal trofoblasto, la zona pellucida viene parzialmente distrutta e da essa emerge l'embrione ("schiusa"). Con la partecipazione di proteine integrine sintetizzate sia dall'epitelio uterino che dalle cellule del trofoblasto, la blastula è attaccata alla parete uterina con il suo polo embrionale. Entrate in contatto con i tessuti materni, le cellule del trofoblasto si moltiplicano rapidamente e, rilasciando enzimi proteolitici, le distruggono (questi tessuti). Ciò provoca una reazione della parete uterina, accompagnata dalla formazione attiva di vasi nel sito di impianto. La violazione della sintesi di integrine ed enzimi necessari per l'impianto rende impossibile questo processo, che a sua volta porta all'ipossia e alla morte dell'embrione. Durante l'impianto, il trofoblasto si differenzia in due strati: lo strato interno, chiamato citotrofoblastoma, perché fa risparmiare struttura cellulare, e quello esterno, chiamato sincizio-trofoblastoma, perché è un sincizio. La Figura 7.20 mostra un embrione umano nel processo di impianto.
gastrulazione nei mammiferi, è strettamente correlato ad altre trasformazioni embrionali. Contemporaneamente alla divisione del trofoblasto in due strati, il nodulo germinale si appiattisce e si trasforma in un bistrato scudo germinale. Lo strato inferiore dello scudo - ipoblasto, o endoderma primario, secondo la maggior parte degli autori, è formato dalla delaminazione della massa cellulare interna, approssimativamente come avviene nel disco embrionale degli uccelli. Primario
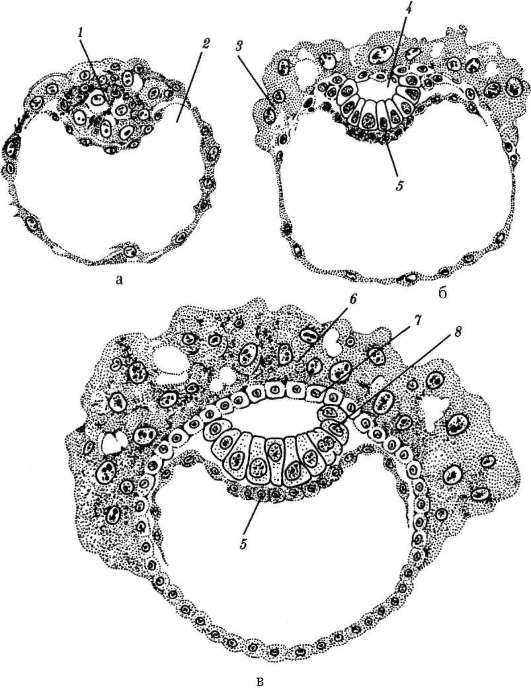
Riso. 7.20.Stadi successivi di impianto e sviluppo dell'embrione umano alla fine della 1a e 2a settimana: a - blastocisti; b - blastocisti all'inizio dell'impianto (7° giorno di sviluppo); c - blastocisti parzialmente impiantate (8° giorno di sviluppo)
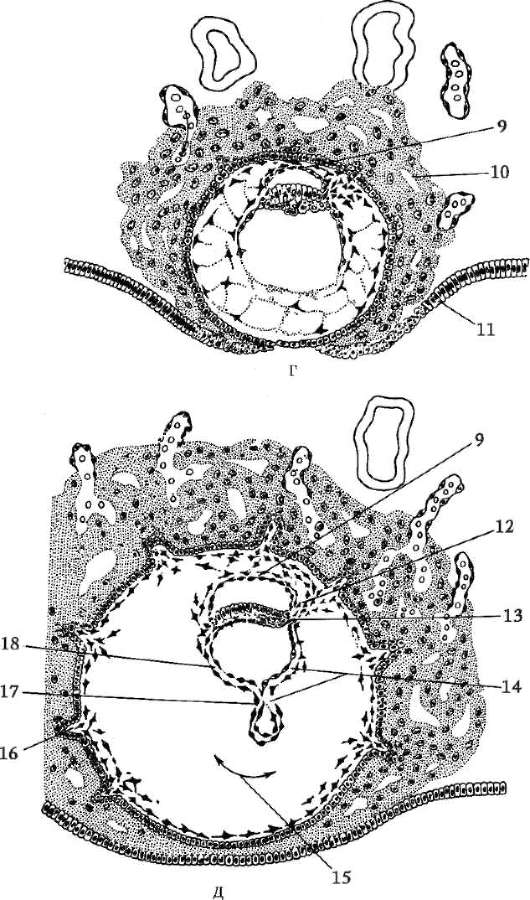
Riso. 7.20. Finale: d - embrione nel 9-10° giorno di sviluppo; e - embrione il 13 ° giorno di sviluppo: 1 - embrioblasto, 2 - blastocele, 3 - trofoblasto, 4 - cavità amniotica, 5 - ipoblasto, 6 - sinciziotrofoblasto, 7 - citotrofoblasto, 8 - epiblasto, 9 - amnion, 10 - lacuna trofoblasto, 11 - epitelio uterino, 12 - gambo corporeo, 13 - rene allantoide, 14 - segno tuorlo, 15 - celoma extra-respiratorio, 16 - villi coriali, 17 - sacco vitellino primario, 18 - sacco vitellino secondario
naya endoderma è completamente speso per la formazione di endoderma extraembrionale. Foderando la cavità del trofoblasto, forma il sacco vitellino primario dei mammiferi.
Strato cellulare superiore epiblasto- la fonte del futuro ectoderma, mesoderma ed endoderma secondario. Alla 3a settimana si forma l'epiblasto linea primaria, il cui sviluppo è accompagnato da quasi gli stessi movimenti delle masse cellulari come nella formazione della striscia primaria degli uccelli (Fig. 7.21). All'estremità della striscia primaria, Nodo di Hensen E buco primario, omologo al labbro dorsale del blastoporo di altri vertebrati. Le cellule che si muovono nella regione della fossa primaria sono dirette sotto l'epiblasto verso il piatto precordale.
piatto precordale situato all'estremità della testa dell'embrione e segna il sito della futura membrana orofaringea. Le cellule che si muovono lungo l'asse centrale formano il rudimento della notocorda e del mesoderma e si compongono processo cordomesodermico. Il nodo di Hensen si sposta gradualmente verso l'estremità caudale dell'embrione, la striscia primaria si accorcia e il rudimento della notocorda si allunga. Ai lati del processo cordomeso-dermico si formano placche mesodermiche che si espandono in entrambe le direzioni. La Figura 7.22 presenta un diagramma generalizzato di alcuni dei processi dello sviluppo embrionale precoce.
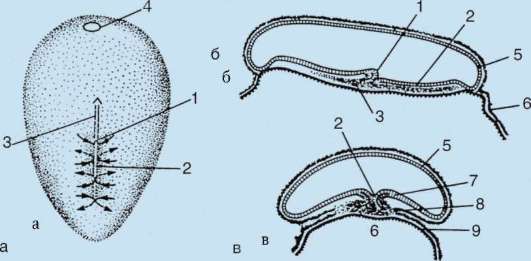
Riso. 7.21.Sviluppo dell'embrione umano nella fase della striscia primaria (15-17 giorni): a - vista dell'embrione dall'alto (l'amnion viene rimosso); b - sezione longitudinale; c - sezione trasversale attraverso la striscia primaria: 1 - nodo di Hensen; 2 - striscia primaria; 3 - accordo; 4 - piatto precordale; 5 - amnio; 6 - sacco vitellino; 7 - ectoderma; 8 - mesoderma; 9 - endoderma
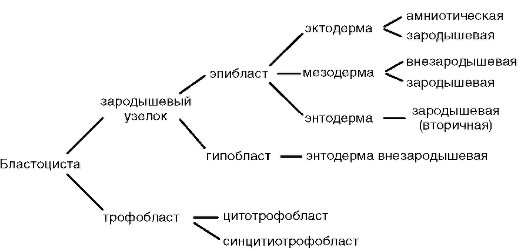
Riso. 7.22. Differenziazione degli strati germinali dei mammiferi
Entro la fine della 3a settimana, nell'ectoderma dell'embrione sopra il rudimento della notocorda, placca neurale. Consiste di celle cilindriche alte. Al centro della piastra neurale si forma una deflessione nella forma grondaia nervosa, e sui suoi lati si alzano pieghe nervose. Questo è l'inizio della neurulazione. Nella parte centrale dell'embrione si verifica la chiusura delle pieghe neurali - a tubo neurale. Quindi la chiusura si allarga nelle direzioni della testa e della coda. Il tubo neurale e le parti adiacenti dell'ectoderma, da cui successivamente si sviluppa cresta neurale, completamente immersi e separati dall'ectoderma che cresce insieme sopra di loro (vedi Fig. 7.10). La striscia di cellule che si trova sotto il tubo neurale si trasforma in una corda. Segmenti del mesoderma dorsale - somiti - compaiono ai lati della notocorda e del tubo neurale nella parte centrale dell'embrione. Entro la fine della 4a settimana, si diffondono alla testa e alla coda, il loro numero raggiunge circa 40 paia.
L'inizio della formazione dell'intestino secondario, l'anlage del cuore e la vascolarizzazione del sacco vitellino appartengono allo stesso periodo. La Figura 7.23 mostra il rapporto tra la dimensione dell'embrione e gli organi extraembrionali al 21° giorno di sviluppo. Più in dettaglio, la separazione del corpo dell'embrione dalle membrane embrionali e la deposizione degli organi possono essere viste in Fig. 7.24, che mostra non solo la vista generale dell'embrione, ma anche i piani di taglio. Si richiama l'attenzione sulla rapida (entro 7 giorni dalla 4a settimana) formazione dell'embrione sotto forma di un corpo allungato e ricurvo, sollevato e tagliato dalle pieghe del tronco dal sacco vitellino. Dietro
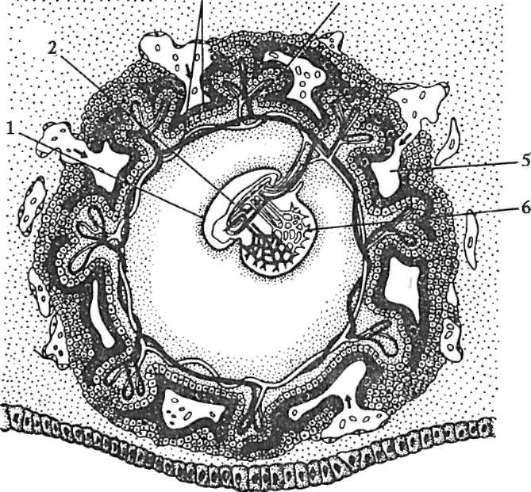
Riso. 7.23.L'embrione e gli organi umani extra-embrionali nel 21° giorno di sviluppo: 1 - amnion; 2 - embrione; 3 - corion; 4 - villo terziario; 5 - sangue materno; 6 - sacco vitellino
in questo momento vengono deposti tutti i somiti, quattro paia di archi branchiali, il tubo cardiaco, i reni degli arti, l'intestino medio, nonché le "tasche" dell'intestino anteriore e posteriore.
Nelle successive quattro settimane di sviluppo embrionale, vengono deposti tutti gli organi principali. La violazione del processo di sviluppo durante questo periodo porta alle malformazioni congenite più gravi e multiple.
Come notato sopra, lo sviluppo di organi provvisori extra-embrionali nei mammiferi e nell'uomo ha caratteristiche specifiche. Questi organi si formano molto presto, contemporaneamente alla gastrulazione, e in modo leggermente diverso rispetto ad altri amnioti. L'inizio dello sviluppo del corion e dell'amnion cade il 7-8° giorno, cioè coincide con l'inizio dell'impianto. Corion nasce dal trofoblasto, che si è già diviso in citotrofoblasto e sinciziotrofoblasto. Quest'ultimo, sotto l'influenza del contatto con la mucosa dell'utero, cresce e lo distrugge. Entro la fine della seconda settimana,
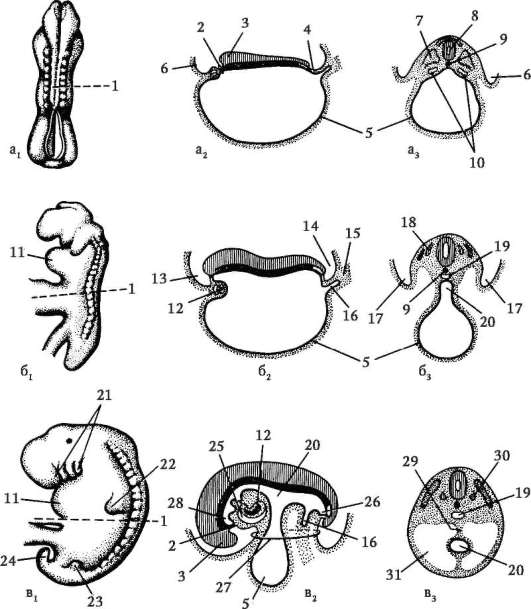
Riso. 7.24.Lo sviluppo dell'embrione umano alla 4a settimana: a 1 b 1 in 1 - visione d'insieme; a 2 b 2 c 2 - sezione longitudinale; a 3 b 3 c 3 - sezione trasversale; a 1 a 2 a 3 - 22 giorni; b 1 b 2 b 3 - 24 giorni; in 1 in 2 in 3 - 28 giorni: 1 - il livello della sezione trasversale; 2 - membrana orofaringea; 3 - cervello; 4 - membrana cloacale; 5 - sacco vitellino; 6 - amnio; 7 - somiti; 8 - tubo neurale; 9 - accordo; 10 - segnalibri accoppiati dell'aorta addominale; 11 - sporgenza cardiaca; 12 - cuore; 13 - piega del tronco della testa; 14 - piega del tronco della coda; 15 - gamba del corpo; 16 - allantois; 17 - pieghe laterali del tronco; 18 - cresta neurale; 19 - aorta dorsale; 20 - intestino medio; 21 - archi branchiali; 22 - rene della zampa anteriore; 23 - rene dell'arto posteriore; 24 - coda; 25 - pericardio; 26 - tasca dell'intestino posteriore; 27 - cordone ombelicale; 28 - tasca dell'intestino anteriore; 29 - mesentere dorsale; 30 - nodo nervoso della radice posteriore; 31 - intero intraembrionale
i villi primari del corion si formano sotto forma di un accumulo di cellule epiteliali del citotrofoblasto. All'inizio della 3a settimana, il mesenchima mesodermico cresce al loro interno e compaiono i villi secondari, e quando i vasi sanguigni compaiono all'interno del nucleo del tessuto connettivo entro la fine della 3a settimana, vengono chiamati villi terziari. Viene chiamata l'area in cui i tessuti del corion e la mucosa dell'utero sono strettamente adiacenti placenta.
Nell'uomo, come in altri primati, i vasi della parte materna della placenta perdono la loro continuità ei villi coriali sono di fatto bagnati dal sangue e dalla linfa dell'organismo materno. Questa placenta si chiama emocoriale. Con il progredire della gravidanza, i villi aumentano di dimensioni, si ramificano, ma il sangue del feto dall'inizio alla fine viene isolato dal sangue materno dalla barriera placentare.
Barriera placentare consiste di trofoblasto, tessuto connettivo ed endotelio vascolare del feto. Questa barriera è permeabile all'acqua, agli elettroliti, ai nutrienti e ai prodotti di dissimilazione, nonché agli antigeni degli eritrociti fetali e agli anticorpi materni, alle sostanze tossiche e agli ormoni. Le cellule della placenta producono quattro ormoni, inclusa la gonadotropina corionica umana, che si trova nelle urine di una donna incinta dalla 2-3a settimana di gravidanza.
Amnio si verifica per divergenza delle cellule epiblastiche della massa cellulare interna. L'amnio umano è chiamato schizamnion(vedi Fig. 7.20) in contrasto con pleurico uccelli e alcuni mammiferi. La cavità amniotica è per qualche tempo delimitata da cellule epiblastiche e in parte da un sito di trofoblasto. Quindi, le pareti laterali dell'epiblasto formano pieghe dirette verso l'alto, che successivamente si uniscono. La cavità è completamente rivestita da cellule epiblastiche (ectodermiche). All'esterno, l'ectoderma amniotico è circondato da cellule mesodermiche extraembrionali. Il liquido amniotico contiene una varietà di sostanze, come proteine, carboidrati, sali, oligoelementi, ormoni, ecc. La sua composizione cambia durante lo sviluppo, inoltre, viene costantemente aggiornata. Nel processo di sviluppo dell'embrione, svolge un ruolo importante nella formazione delle cavità nasali e orali, degli organi digestivi e respiratori.
Sacco vitellino appare quando un sottile strato dell'ipoblasto si separa dalla massa cellulare interna e le sue cellule endodermiche extra-embrionali, muovendosi, rivestono la superficie del tro-
foblasto. Il risultante sacco vitellino primario collassa il 12-13° giorno e viene convertito in un sacco vitellino secondario associato all'embrione dal dotto vitellino. Le cellule endodermiche sono ricoperte esternamente dal mesoderma extraembrionale. Alla 3a settimana di sviluppo, le cellule germinali primarie, le cellule del sangue e i vasi sanguigni iniziano a essere rilevate nella parete del sacco vitellino. Fino alla 7-8a settimana di sviluppo, questo sacco vitellino è il principale organo ematopoietico dell'embrione.
Allantois nasce nell'embrione umano, come in altri amnioti, sotto forma di una tasca nella parete ventrale dell'intestino posteriore, ma la sua cavità endodermica rimane una struttura rudimentale. Tuttavia, nelle sue pareti si sviluppa un'abbondante rete di vasi che si collegano con i principali vasi sanguigni dell'embrione. Il mesoderma allantoide si connette con il mesoderma corion, dandogli vasi sanguigni, che forniscono nutrimento all'embrione in via di sviluppo fino a circa il secondo mese di embriogenesi. Pertanto, si verifica la vascolarizzazione della placenta corioallantoidea.
Alcune fasi e termini di sviluppo degli organi negli embrioni umani sono presentati nella Tabella. 7.4.
Tabella 7.4. Principali periodi ed eventi dell'ontogenesi umana primitiva
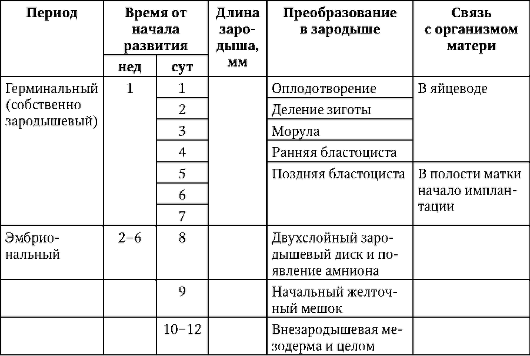
Continuazione della tavola. 7.4
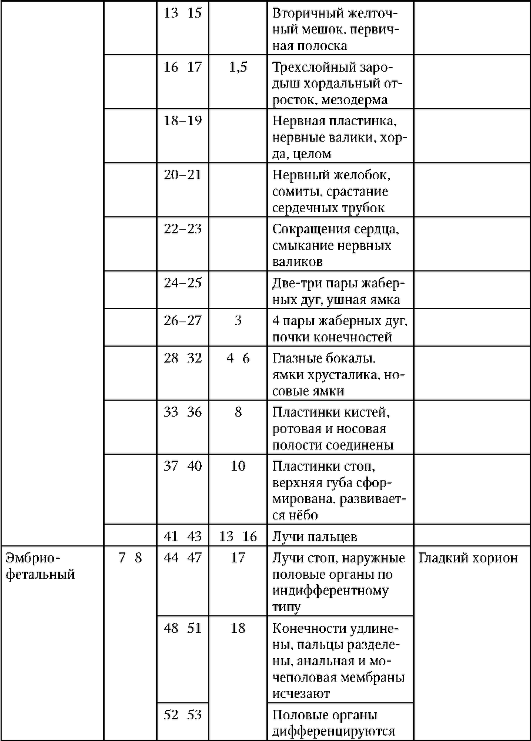
La fine del tavolo. 7.4
7.5.2. ESEMPI DI ORGANOGENESI UMANA CHE RIFLETTONO L'EVOLUZIONE DELLA SPECIE
Questa sezione non ha lo scopo di descrivere lo sviluppo di tutti gli organi umani. Saranno considerati solo alcuni processi morfogenetici, illustrando i seguenti punti biologici generali:
Il valore delle interazioni intercellulari, tissutali e interorgano nella morfogenesi;
Riflessione nella morfogenesi umana di caratteristiche evolutivamente più antiche dell'organizzazione dei vertebrati;
Caratteristiche dell'organogenesi, che consentono di valutare lo stadio e i meccanismi dei disturbi dello sviluppo in alcune malformazioni congenite.
Struttura segmentale colonna vertebrale riflette la sua origine dai somiti dell'embrione. Inoltre, la formazione delle vertebre è strettamente correlata alla notocorda e al midollo spinale. I corpi vertebrali sono formati da ammassi di cellule mesenchimali che migrano verso la notocorda e la circondano, che originano dagli sclerotomi. Le cellule mesenchimali di due coppie adiacenti di somiti sono coinvolte nella formazione dei corpi vertebrali, e quindi i corpi vertebrali non si trovano a livello dei somiti, ma tra di loro. Allo stesso tempo, i muscoli che si sviluppano dai miotomi del somi-
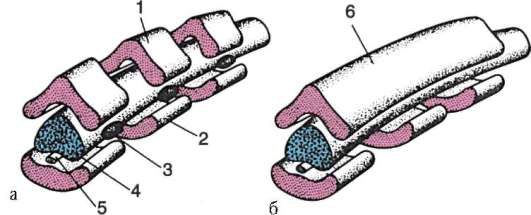
Riso. 7.25. Lo sviluppo degli archi dorsali delle vertebre in un embrione di pollo: a - sviluppo normale; b - dopo la rimozione dei gangli spinali: 1 - arco dorsale; 2 - corpo vertebrale; 3 - nodo spinale; 4 - midollo spinale; 5 - accordo; 6 - archi dorsali non segmentati delle vertebre
comov, come se gettato sulle articolazioni intervertebrali e garantire il movimento delle vertebre.
Gli archi vertebrali sono formati da accumuli di cellule sclerotomiche mesenchimali che si diffondono nella direzione dorsale (dorsale), mentre i processi trasversali e le costole sono formati da accumuli di cellule sclerotomiche che migrano lateralmente. Il midollo spinale e i gangli spinali nelle fasi appropriate dello sviluppo intrauterino sono coinvolti nella morfogenesi degli archi dorsali delle vertebre. Quindi, se i gangli spinali vengono rimossi, si forma la cartilagine degli archi dorsali, ma sembra un bastoncino non segmentato (Fig. 7.25).
La struttura istologica delle vertebre cambia gradualmente. Durante la 4a-5a settimana si formano le vertebre mesenchimali, alla 6a settimana compaiono focolai di condrogenesi e poi all'ottava settimana inizia l'ossificazione, che dura fino a quasi 25 anni. Le sezioni della corda situate all'interno dei corpi vertebrali scompaiono gradualmente. All'interno dei dischi intervertebrali, la notocorda è conservata come una struttura simile al muco nota come nucleo polposo.
Formazione orecchio medio associato alle trasformazioni ontogenetiche del primo e del secondo archi viscerali (branchie). La Figura 7.26 mostra schematicamente le aree della testa e del collo alla 4a e 24a (fine del primo-secondo trimestre di sviluppo prenatale) settimane di sviluppo umano. Alla 4a settimana, gli archi viscerali (branchie) vengono deposti nelle regioni della testa e del collo dell'embrione a destra ea sinistra. Dall'interno, dal lato della faringe, si formano tra di loro tasche faringee(figura 7.27). Queste sono sporgenze (o sporgenze) dell'endoderma,
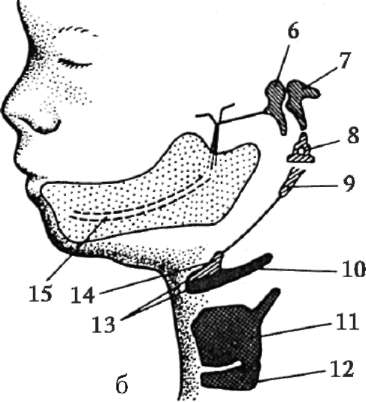
Riso. 7.26.Testa e collo di un embrione umano, vista laterale: a - alla 4a settimana; b - nella 24a settimana: 1 - il primo arco; 2 - posizione dell'orecchio interno in via di sviluppo; 3 - secondo arco; 4 - terzo arco; 5 - quarto prato; 6 - martello. 7 - incudine; 8 - staffa; 9 - processo stiloideo; 10 - grande corno dell'osso ioide; 11 - cartilagine tiroidea; 12 - cartilagine cricoidea; 13 - il corpo dell'osso ioide; 14 - piccolo corno; 15 - mascella inferiore; anlages di archi viscerali (a) e organi che si sviluppano da loro (b) sono indicati dalla stessa ombreggiatura
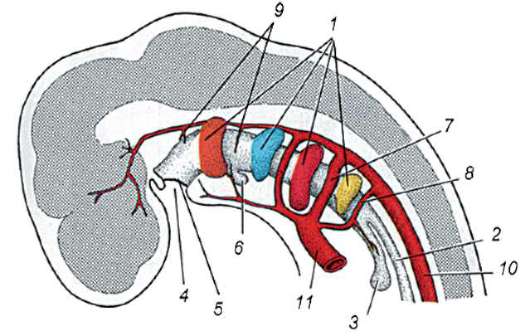
Riso. 7.27.Regione branchiale di un embrione umano di 5 settimane (ectoderma tegumentario e mesenchima rimossi). 1 - tasche faringee; 2 - esofago; 3 - trachea e rene polmonare; 4 - stomodeo; 5 - placca stomodale (orale); 6 - rudimento della ghiandola tiroidea; 7 - 4° arco aortico; 8 - 6° arco aortico; 9 - riduzione del 1o e 2o arco aortico; 10 - aorta discendente dorsale; 11 - aorta ascendente addominale (sacco arterioso)
crescendo lateralmente. Fuori, secondo le escrescenze faringee, ectodermiche tasche branchiali.
L'endoderma e l'ectoderma delle prime tasche faringee e branchiali entrano in contatto alla 4a settimana di sviluppo (Fig. 7.28, a). Il contatto non dura a lungo, l'estremità cieca della tasca faringea si allontana dalla superficie ed è circondata dal mesenchima (Fig. 7.28, b). Dalla parte distale della tasca faringea inizia a svilupparsi la cavità dell'orecchio medio (cavità timpanica) e dalla parte prossimale si forma la tuba uditiva (di Eustachio). Dal mesenchima si differenziano ulteriormente gli anlages degli ossicini uditivi, che servono a condurre il suono (Fig. 7.28, c). Il martello e l'incudine originano dal materiale cellulare del primo arco viscerale, la staffa - dal materiale cellulare della parte dorsale del secondo arco viscerale. Muscoli e nervi collegati all'apparato ossiculare che conduce il suono dagli ossicini dell'orecchio medio sono formati dal materiale cellulare della regione del primo e del secondo arco viscerale.
Alla fine della vita intrauterina ed entro pochi mesi dalla nascita, il tessuto connettivo embrionale nella cavità timpanica viene riassorbito. Solo dopo questo, gli ossicini uditivi acquisiscono mobilità e le vibrazioni della membrana timpanica possono essere trasmesse liberamente alla membrana della finestra ovale dell'orecchio interno.
Nella formazione delle strutture zona facciale E cavità orale prendono parte le cellule ecto e mesodermiche. La maggior parte struttura
Riso. 7.28. Sviluppo dell'orecchio medio, sezione frontale: a - alla 4a settimana; b - la 5a settimana; c - fasi successive; 1 - primo piatto branchia; 2 - prima tasca branchiale; 3 - il primo arco viscerale (branchiale); 4 - vescicola uditiva; 5 - prima tasca faringea; 6 - derivati del primo arco viscerale; 7 - derivato del secondo arco viscerale; 8 - cavità tubo-schiavo; 9 - tappo del canale uditivo esterno. 10 - meato uditivo esterno primario; 11 - martello; 12 - incudine; 13 - staffa; 14 - cavità timpanica; 15 - tuba uditiva (di Eustachio).
il giro della regione facciale si sviluppa da masse di cellule mesenchimali di origine ectodermica che migrano dalla cresta neurale (4° strato germinativo). Da questo mesenchima si sviluppano i tessuti connettivi, cartilaginei e ossei e il tessuto della polpa dentale. I muscoli striati dei muscoli del viso e della faringe sono formati dal mesenchima di origine mesodermica. Qui è importante capire come si è svolto il processo di "espansione e cambiamento di funzioni" nell'antropogenesi.
La cavità orale è formata da un'invaginazione ectodermica stomo-deum, adiacente all'avambraccio che termina alla cieca. Sono separati da sottile piatto stomodale, costituito da fogli ectodermici (dal lato dello stomodeo) ed endodermici (dal lato dell'intestino) (vedi Fig. 7-26). Questa piastra si rompe alla fine della 4a settimana di sviluppo e la cavità orale è collegata al tubo intestinale inizialmente chiuso alla cieca.
Le ghiandole salivari parotidi si sviluppano dall'ectoderma dello stomodeo e le ghiandole salivari sublinguali e sottomandibolari si sviluppano dall'endoderma della parte ventrale della faringe.
In un feto di 5 settimane apertura della bocca limitato dalle seguenti strutture (Fig. 7.29, a): nella linea mediana sopra la cavità orale è presente un'area sporgente arrotondata - sporgenza frontale (frontale); ai suoi lati si trovano i rilievi a ferro di cavallo che circondano le fosse olfattive. I processi medi di queste elevazioni
Riso. 7.29. Fasi successive della formazione del viso (vista frontale): a - alla 5a settimana; b - alla decima settimana. 1 - stomodeo; 2 - sporgenza frontale (frontale); 3 - processo nasale mediale; 4 - processo nasale laterale; 5 - fossa olfattiva; 6 - processo mascellare; 7 - posa dell'arco mandibolare; 8 - filtro; 9 - occhio
chiamato nasale mediale, e quelli laterali laterale
processi nasali. Dagli angoli laterali superiori della cavità orale crescono verso la linea mediana processi mascellari.
Limitare la cavità orale dal basso mascella inferiore formato da segnalibri accoppiati dell'arco mandibolare. All'inizio compaiono su ciascun lato della linea mediana, quindi aumentano l'uno verso l'altro e crescono insieme.
Durante la sesta settimana c'è un rapido sviluppo mascella superiore. I processi mascellari crescono verso la linea mediana. Allo stesso tempo, i processi nasali aumentano, spingendo verso l'alto la parte inferiore della sporgenza frontale situata tra di loro. I processi nasali mediali crescono in modo particolarmente intenso, che si spostano verso la linea mediana e si fondono tra loro, quindi si fondono con i processi mascellari, formando così un arco mascellare completo alla 10a settimana di sviluppo. La pelle ektoderm sopra i processi medi uniti è formata filtro- un solco longitudinale sopra il labbro superiore sotto il naso (Fig. 7.29, b).
Entro la fine del secondo mese di sviluppo, quando si forma la mascella superiore, inizia a svilupparsi cielo solido. Nella sua formazione, così come nella formazione dell'arco della mascella superiore, sono coinvolti processi nasali medi e mascellari(Fig. 7.30, a). Dalla regione mediana della mascella superiore, nata dalla fusione dei processi nasali medi, si forma una piccola parte triangolare - palato primario. Parte principale - palato secondario- si sviluppa da escrescenze palatine laterali (processi), che compaiono sulla mascella superiore (Fig. 7.30, b). All'inizio sono diretti verso il basso, quindi i bordi dei processi palatali si alzano e verso la linea mediana. Entrano in contatto e si fondono per formare la sutura del palato. La parte principale formata del palato separa la parte più alta della cavità stomodale originaria. Il palato secondario cresce insieme a quello primario e il setto nasale cresce fino alla loro superficie cranica (Fig. 7.30, c). Pertanto, contemporaneamente alla separazione della cavità nasale dalla cavità orale, è divisa nelle metà destra e sinistra.
Nel periodo iniziale della sua formazione (nella quinta settimana di embriogenesi) linguaè una sacca formata dalla membrana mucosa, piena di massa muscolare in crescita. Si ritiene che la mucosa della lingua derivi da ectoderma stomodeo, e i muscoli vengono da miotomi occipitali.
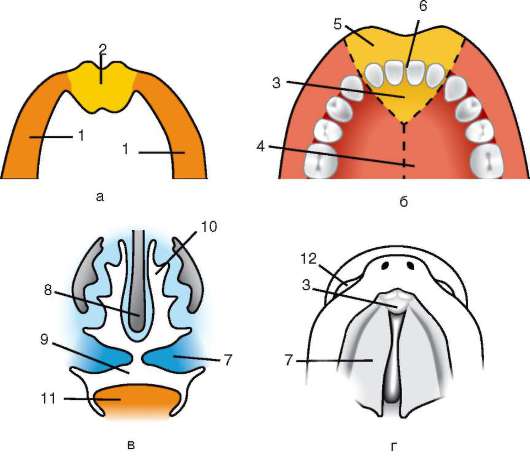
Riso. 7.30. Formazione del palato duro: a - una rappresentazione schematica della mascella superiore (vista dal lato della cavità orale); b - 10a settimana di sviluppo (vista dalla cavità orale); c - 8a settimana di sviluppo (sezione frontale); d - 8a settimana di sviluppo (vista dalla cavità orale): 1 - processo mascellare; 2 - processi nasali mediali uniti; 3 - cielo primario; 4 - palato secondario (escrescenze palatine laterali); 5 - filtro labiale; 6 - mascella superiore con incisivi; 7 - escrescenza palatina laterale; 8 - setto nasale; 9 - cavità orale; 10 - cavità nasale; 11 - lingua; 12 - occhio
Diverse strutture del pavimento della cavità orale sono coinvolte nella formazione della mucosa della lingua. Il corpo della lingua è formato dalla fusione di ispessimenti laterali accoppiati - tubercoli linguali laterali e situato tra loro una piccola eminenza mediana, chiamata tubercolo impar(figura 7.31). Si ritiene che il corpo della lingua sia un derivato del materiale cellulare della regione del primo arco viscerale. Partecipa alla formazione della radice della lingua corula(parentesi)- eminenza mediana, situata dietro tubercolo impar e accendi
Riso. 7.31. Sviluppo del linguaggio: a - embrione di 5 settimane; b - feto di 5 mesi; 1 - tubercolo linguale laterale; 2- tubercolo impar 3 - copula (parentesi); 4 - forame cieco; 5 - solco terminale; 6 - il corpo della lingua; 7 - la radice della lingua; 8 - posa della cartilagine epiglottica; 9 - glottide; 10 - tubercolo aritenoideo; 11 - epiglottide, tonsilla palatina. I-IV - numero di archi viscerali
il materiale cellulare delle aree del secondo, terzo e quarto arco viscerale. Dopo la fusione di tutti gli anlages e la formazione finale della lingua, il suo corpo viene separato dalla radice solco terminale(solco terminale), al centro di cui è buco cieco(forame cieco)- una traccia di invaginazione del fondo della faringe, da cui appare la ghiandola tiroidea (vedi Fig. 7.27). È considerato il confine tra il corpo e la radice della lingua.
Successivamente, la lingua si separa quasi completamente dal fondo della cavità orale e la connessione tra loro viene preservata sotto forma di una piega della mucosa: il frenulo della lingua.
Un interessante esempio di organogenesi è la formazione denti, a cui partecipa l'ectoderma tegumentario - l'epitelio dello stomodeo (smalto) e le cellule mesenchimali originate dalla cresta neurale (dentina, cemento e polpa) (Fig. 7.32). Nel corso dello sviluppo, si notano interazioni di induzione tra le strutture mesenchimali ed epiteliali.
Durante la vita si sviluppano due generazioni di denti. Il cambio di denti avviene durante l'infanzia, vengono chiamati i primi denti latticini. Ce ne sono 20: 10 nella mascella superiore e 10 nella mascella inferiore. I denti da latte erompono completamente a circa 2 anni di età. Questa dentatura serve il bambino per i successivi 4 anni, dopodiché i denti da latte gradualmente
Riso. 7.32. Denti permanenti umani: 1 - incisivi; 2 - zanne; 3 - piccoli molari; 4 - grandi molari
ma cadono e vengono sostituiti Denti permanenti(seconda generazione di denti) che funzionano in una persona per il resto della vita. Il periodo di sostituzione dei denti dura approssimativamente dai 6 ai 12 anni. 32 denti permanenti: 16 superiori e 16 inferiori (vengono aggiunti molari grandi) (vedi Fig. 7.32). Nella forma sono simili ai latticini, ma sono di dimensioni maggiori.
Segnalibro e istruzione germi dentali i denti da latte iniziano da 6-7 settimane di sviluppo intrauterino. L'epitelio della cavità orale è immerso nel mesenchima sottostante sotto forma di un denso filamento chiamato placca dentale(figura 7.33). Piccole sporgenze epiteliali compaiono sulla placca dentale - gemme dentali, da cui si svilupperanno i denti da latte. Man mano che la lamina dentale cresce, ogni germe del dente aumenta di dimensioni, penetra più in profondità nel mesenchima e assume la forma di una coppa rovesciata. Questa struttura si forma organo smaltato, e si forma il mesenchima sottostante, che riempie la cavità della ciotola papilla dentale. In ulteriore sviluppo, quest'ultimo forma la polpa del dente. Al 3° mese di sviluppo intrauterino, l'organo dello smalto aumenta di dimensioni, cambia forma e si separa gradualmente dalla placca dentale. La forma del dente futuro è determinata dalla componente mesenchimale. Quando combinato in vitro mesenchima della posa del dente molare (molare) con la componente epiteliale della posa dell'incisivo, si sviluppa il molare. Al contrario, come risultato della combinazione dell'ectoderma del molare e del mesenchima dell'incisivo, si sviluppa un incisivo.
Durante l'istogenesi dei tessuti dentali, che inizia alla fine del 4° mese, dentina, smalto e polpa(Fig. 7.33, d). Smalto
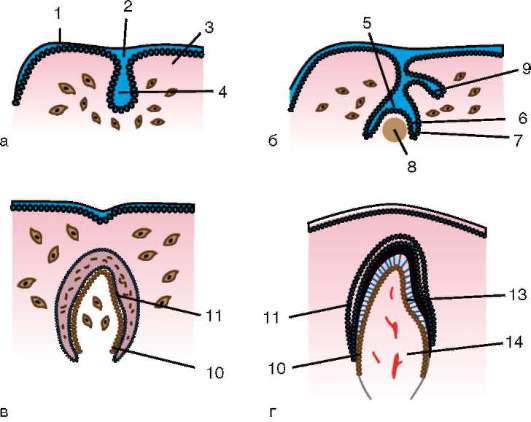
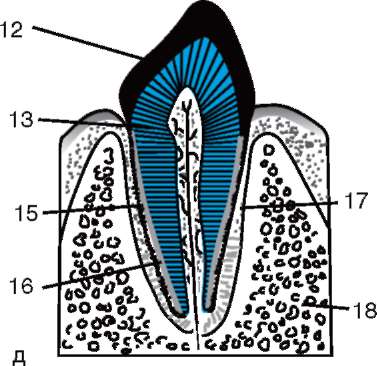
Riso. 7.33.Sviluppo dei denti: a - embrione di 8 settimane; b - feto di 10 settimane; c - feto di 3 mesi; g - feto di 6 mesi; d - dopo l'eruzione; 1 - epitelio della cavità orale; 2 - placca dentale; 3 - mesenchima; 4 - il germe di un dente da latte; 5 - organo smaltato; 6 - lo strato interno dell'organo dello smalto; 7 - strato esterno dell'organo dello smalto; 8 - papilla dentale; 9 - il germe di un dente permanente; 10 - odontoblasti; 11 - ameloblasti; 12 - smalto; 13 - dentina; 14 - polpa del dente; 15 - cementoblasti; 16 - cemento; 17 - parodontale; 18 - osso alveolare
produrre cellule dello strato interno dell'organo dello smalto - esplosioni di smalto. I loro processi secernono la base organica dello smalto - prismi di smalto, che poi calcificano. Fonte di sviluppo dentina- odontoblasti (dentinoblasti), cellule pulpari superficiali. L'apice dei dentinoblasti ha anche processi che secernono sostanze organiche di una struttura fibrillare, chiamate predentin e formando la matrice della dentina. Dalla fine del 5° mese, nella predentina si depositano i sali di calcio e fosforo e si forma la dentina finale. La formazione della dentina e dello smalto differisce dallo sviluppo dell'osso in quanto le cellule non sono murate sostanza intercellulare, ma allontanati: ameloblasti - verso l'esterno, odontoblasti - verso l'interno. Inoltre, il contenuto di sostanze organiche in questi tessuti è diverso: l'osso contiene circa il 45% di materia organica, la dentina - 28% e lo smalto - meno del 5%.
Sulla superficie esterna della dentina dal mesenchima che circonda il dente (sacca dentale) appaiono poco prima della dentizione cementoblasti. Queste gabbie secernono fibre di collagene e sostanza intercellulare, formando cemento. Si forma solo 4-5 mesi dopo la nascita durante lo sviluppo delle radici dei denti. Dallo strato esterno del sacco dentale si forma il dente. legamento (parodonto)(Fig. 7.33, e). Pertanto, nello sviluppo prenatale, si sviluppano solo le corone dei denti da latte.
L'eruzione del dente è fornita da tre fattori. La prima e più importante è la crescita e lo sviluppo della radice, che porta all'espulsione della chioma attraverso la mucosa. Il secondo fattore è la crescita della polpa, che porta ad un aumento della pressione all'interno del germe del dente. Inoltre, si verifica un'ulteriore deposizione nella parte inferiore dell'alveolo dentale. tessuto osseo strati.
La deposizione dei denti permanenti inizia molto presto, all'inizio del 5° mese di sviluppo intrauterino. Il germe del dente permanente si trova dietro il germe del dente da latte e lo sviluppo della radice di quest'ultimo nei primi due anni dopo la nascita ha un effetto di induzione sullo sviluppo del germe del dente permanente. I processi istologici che portano alla formazione del latte e dei denti permanenti sono gli stessi. Quando i denti da latte eruttano, lo smalto e la dentina si formano nei rudimenti dei denti permanenti. Per qualche tempo rimangono in uno stato latente e si attivano solo dopo che la mascella ha raggiunto dimensioni sufficientemente grandi. Nel processo di sostituzione, la crescita di un dente permanente e la pressione del suo smalto sulla radice del latte portano al riassorbimento.
osteoclasti (cellule che distruggono il tessuto osseo) di tessuto più morbido - la dentina di un dente da latte. Quest'ultimo viene espulso e sostituito da uno permanente.
Sviluppo del dipartimento digestivo i sistemi sono difficili da considerare in dettaglio, poiché il tubo intestinale è notevolmente allungato, differenziato e connesso nel suo sviluppo con molti altri sistemi: endocrino, respiratorio, escretore, circolatorio, nervoso e cavità secondaria - celoma. Consideriamo solo i primi e più comuni processi che si verificano nel tubo intestinale, nonché i cambiamenti nella sua connessione con il sacco vitellino.
Il tubo intestinale nasce dall'endoderma del tetto del sacco vitellino e dal foglio viscerale adiacente del mesoderma nel processo di separazione del corpo dell'embrione dalle parti extraembrionali usando le pieghe della testa, della coda e del tronco laterale (vedi Fig. 7.24, b2, b3). Alla 4a settimana, è rappresentato da un tubo relativamente semplice, costituito dall'intestino anteriore, la placca stomodale (orale) ciecamente chiusa anteriormente, l'intestino medio associato al sacco vitellino
attraverso il gambo del tuorlo (dotto) e l'intestino posteriore, chiuso alla cieca all'estremità della coda da una membrana cloacale (anale)
(figura 7.34).
Durante il periodo dalla 4a alla fine dell'8a settimana di sviluppo embrionale (intrauterino), l'intestino si allunga e comincia a differenziarsi. Dalla sua parte anteriore si formano la faringe, l'esofago, lo stomaco e il duodeno con i suoi derivati (fegato e pancreas) prima che vi scorra il dotto biliare comune.
L'intestino medio si forma Varie a forma di U un anello che è collegato al sacco vitellino attraverso il dotto vitellino (Fig. 7.35, a). La sezione dell'intestino tra il gambo del tuorlo e lo stomaco si trasformerà nell'intestino tenue e nella sezione
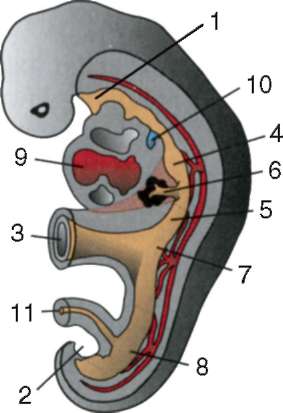
Riso. 7.34.Apparato digerente di un embrione di 4 settimane: 1 - piastra stomo-affare; 2 - membrana cloacale; 3 - dotto tuorlo; 4 - stomaco; 5 - duodeno; 6 - rudimento di fegato; 7 - intestino medio; 8 - intestino posteriore; 9 - cuore; 10 - posa degli organi respiratori; 11 - uraco
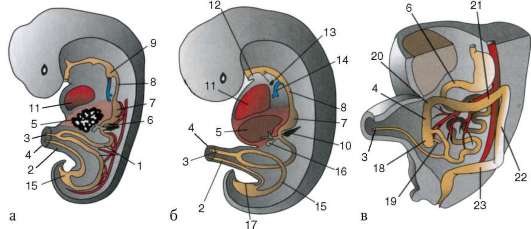
Riso. 7.35.Sviluppo dell'apparato digerente umano: a - il 32esimo giorno; b - il 36° giorno; c - fasi successive; 1 - Ansa a forma di U dell'intestino; 2 - uraco; 3 - dotto tuorlo; 4 - gambo addominale (cordone ombelicale); 5 - fegato; 6 - duodeno; 7 - stomaco; 8 - esofago; 9 - laringe; 10 - pancreas; 11 - cuore; 12 - lingua; 13 - segnalibro della ghiandola tiroidea; 14 - rudimento della trachea e dei polmoni; 15 - intestino posteriore; 16 - cistifellea; 17 - membrana cloacale; 18 - cieco; 19 - appendice; 20-23 - sezioni dell'intestino crasso
premendo caudalmente al dotto vitellino, - nell'intestino crasso. L'intestino medio si allunga, parte di esso sporge nella cavità del gambo addominale, formando una sporgenza erniaria della parete addominale dell'embrione.
Nello stesso tempo, alcune sezioni dell'intestino, ruotando, cambiano posizione (Fig. 7.35, b). Entro la decima settimana, la parte sporgente dell'ansa intestinale viene tirata indietro attraverso il bordo ombelicale, prendendo la sua posizione finale nella cavità addominale. Un po' prima, nel punto di transizione dell'intestino tenue nell'intestino crasso, si sviluppa il cieco. All'inizio aumenta di dimensioni, ma entro il 3 ° mese la sua parte distale inizia a rimanere indietro rispetto alla crescita di altre parti (crescita irregolare), per cui il diametro di questa parte è molto più piccolo e un'appendice è formato. Dall'intestino posteriore, l'intestino crasso si forma sotto l'angolo splenico, compreso il retto (Fig. 7.35, c). Lo sviluppo dell'estremità cloacale dell'intestino posteriore è strettamente correlato allo sviluppo dell'orifizio urogenitale (Fig. 7.36). Il setto urorettale, che separa la cloaca nel seno urogenitale e nel retto, si forma alla 5a-6a settimana. La rottura della membrana anale avviene nell'ottava settimana.
Al 2 ° mese di sviluppo intrauterino inizia la rapida proliferazione dell'epitelio dell'esofago e del duodeno, che
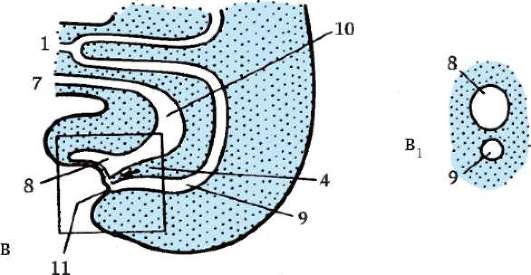
Riso. 7.36. Sviluppo della regione cloacale dell'intestino posteriore nell'embrione umano: a - alla 4a settimana; b - la sesta settimana; c - alla 7a settimana; a 1 b 1 e c 1 - sezioni trasversali attraverso la cloaca (i livelli di taglio sono indicati da quadrati in Fig. a; b; c). 1 - gambo di tuorlo; 2 - intestino medio; 3 - intestino posteriore; 4 - setto urogenitale; 5 - cloaca; 6 - coda; 7 - allantoide; 8 - seno urogenitale; 9 - retto; 10 - vescia; 11 - membrana anale; le frecce mostrano la direzione della crescita
porta alla chiusura temporanea del loro lume. Entro la fine del 2° mese si verifica la ricanalizzazione di questi organi, ad es. ripristino della permeabilità.
Cuore una persona inizia a svilupparsi dalla fine della 3a - l'inizio della 4a settimana dallo splancnomesoderma sotto forma di rudimenti accoppiati situati sotto la faringe (Fig. 7.37). Poiché il corpo dell'embrione stesso è delimitato dalle pieghe del tronco e l'intestino anteriore è chiuso dal lato ventrale, i tubi endocardici accoppiati si chiudono in uno, giacendo lungo la linea mediana (vedi Fig. 7.37). Per gli anfibi è stata dimostrata l'esistenza di un'azione induttiva dell'endoderma sul mesoderma precordiale. Ciò è evidenziato anche da un esperimento su un embrione di pollo, quando, dopo aver asportato il fondo dell'intestino anteriore, è stato ottenuto un embrione con doppio cuore. Anlage accoppiati delle cavità coelomiche si fondono sul lato ventrale, formando la cavità pericardica.
La differenziazione regionale del cuore inizia con un rapido allungamento del tubo cardiaco primario, che porta alla sua flessione e all'acquisizione di una forma a 5 (Fig. 7.38). Si noti che la formazione di una curva con cambiamenti locali nella forma delle cellule lungo il tubo cardiaco viene stimolata dal tubo cardiaco stesso. Essere espiantato (situazione "fuori dal corpo", o ex vivo), il tubo cardiaco conserva la capacità di formare una struttura a forma di 5.
Riso. 7.37. Sviluppo del cuore umano nel 21°-22° giorno: a - visione generale dell'embrione; b - sezione longitudinale del cono della testa dell'embrione; c - sezione trasversale dell'embrione: 1 - livello di taglio; 2 - rullo nervoso; 3 - somiti; 4 - striscia primaria; 5 - cervello; 6 - anteriore; 7 - tubo cardiaco; 8 - pericardio; 9 - amnio; 10 - membrana orofaringea; 11 - solco neurale; 12 - accordo; 13 - segnalibri accoppiati dell'aorta dorsale; 14 - fusione di tubi cardiaci; 15 - sacco vitellino
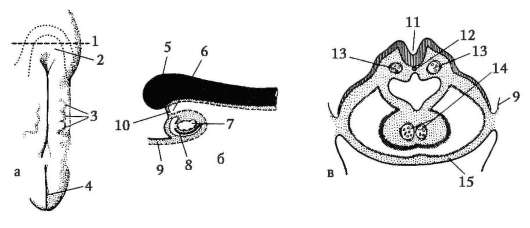
Riso. 7.38. Il cuore umano in via di sviluppo: a - il 21°-22° giorno; b - il 23° giorno; c - il 24° giorno (vista dal lato ventrale); d - il 28° giorno (sezione longitudinale): 1 - il primo paio di archi aortici; 2 - fusione di tubi cardiaci; 3 - tubi cardiaci non uniti; 4 - tronco arterioso; 5 - bulbo del cuore; 6 - ventricolo; 7 - atrio; 8 - seno venoso; 9 - Condotto di Cuvier; 10 - vena ombelicale; 11 - vena di tuorlo; 12 - canale atrioventricolare
All'estremità originariamente caudale si trova il seno venoso, in cui confluiscono grandi vene: i dotti di Cuvier, ombelicale e vitellino. Il seno venoso diventerà successivamente parte dell'atrio destro. Cranialmente al seno venoso, l'atrio è formato dalla parte espansa del tubo cardiaco e il ventricolo è formato dalla parte centrale curva. L'area di transizione in cui il ventricolo si restringe è chiamata cono o bulbo. Successivamente sarà incorporato nella parete del ventricolo destro. Il cono passa nel tronco arterioso, da cui partono le radici dell'aorta addominale.
Dopo la formazione dell'atrio e del ventricolo, compaiono segni esterni dell'imminente divisione del cuore nelle metà destra e sinistra, appare il solco mediano. In questo momento, nei tessuti del cuore in via di sviluppo, si nota la morte cellulare focale e l'interazione di elementi extracellulari e cellulari di fattori. Qualche tempo dopo, sulla superficie interna del ventricolo, corrispondente al livello del solco esterno, si forma un setto di corde muscolari che crescono dall'apice del cuore verso l'atrio. Sulle pareti dorsali e ventrali del canale atrioventricolare ristretto, si formano cuscinetti endocardici dal mesenchima sciolto. Successivamente si trasformano in densi tessuto connettivo, crescono insieme e dividono il canale nei condotti destro e sinistro. Allo stesso tempo, appare un setto mediano primario, che viene successivamente sostituito da uno secondario. Ha un foro, chiamato ovale, attraverso il quale
il sangue dall'atrio destro scorre nel sinistro. È necessario per la circolazione sanguigna del feto.
Contemporaneamente ai cambiamenti nella parte principale del cuore come organo indipendente, il tronco arterioso è diviso in due canali. Questo processo inizia alla radice dell'aorta addominale tra il quarto e il sesto arco viscerale. La separazione avviene a causa della formazione di pieghe longitudinali. Le pieghe sono disposte a spirale, crescono all'interno del tronco arterioso e, incontrandosi, lo dividono nell'aorta e nel tronco polmonare. Inoltre, le pieghe si diffondono verso il cono, dove le valvole semilunari dell'aorta e del tronco polmonare si formano da aree specializzate, e più avanti nei ventricoli, dove si incontrano con il tessuto endocardico in movimento del canale atrioventricolare e del setto atriale. Ciò porta alla completa crescita eccessiva dell'orifizio interventricolare.
Fino alla fine della vita intrauterina rimangono aperte solo la valvola nel forame ovale del setto interatriale e il dotto arterioso (botall) che collega il tronco polmonare all'aorta. Sarà discusso più dettagliatamente di seguito. Il dotto arterioso serve a deviare parte del sangue dal ventricolo destro all'aorta, bypassando i polmoni, fino a quando non hanno raggiunto il necessario sviluppo e non hanno ricevuto uno stimolo funzionale in connessione con l'inizio della funzione respiratoria (Fig. 7.39).
Grandi arterie si sviluppano in combinazione con il cuore, a partire dalla 4a settimana. Il sistema circolatorio primario dell'embrione funziona dalla fine della 5a settimana, entro la fine dell'8a settimana viene implementato il piano di base della struttura sistema arterioso. Gli archi viscerali accoppiati formati alla 4a settimana ricevono un adeguato apporto arterioso sotto forma di archi aortici. Questi archi aortici sorgono dal tronco arterioso, o meglio, dalla sua parte espansa - il sacco arterioso (chiamato anche aorta addominale o ascendente) - e terminano nelle radici dell'aorta dorsale, o discendente (vedi Fig. 7.27).
Nonostante siano posate sei paia di archi aortici (viscerali), non coesistono contemporaneamente. Durante il periodo in cui si forma la sesta coppia di archi, le prime due coppie stanno già degenerando. Il quinto arco appare solo su poco tempo sotto forma di un vaso rudimentale.
Nella formazione dei vasi principali giocano un ruolo essenziale le radici dell'aorta addominale e dorsale, nonché la terza, la quarta e la sesta coppia di archi aortici (Fig. 7.40).
Le parti prossimali della terza coppia formano le arterie carotidi comuni (Fig. 7.41). Il quarto arco sinistro fa parte dell'arco aortico.
Riso. 7.39. Schema di circolazione fetale: 1 - finestra ovale; 2 - setto interatriale; 3 - vena polmonare; 4 - vena cava superiore; 5 - dotto arterioso (botall); 6 - arteria polmonare; 7 - aorta discendente; 8 - arterie ombelicali; 9 - vena ombelicale; 10 - vena cava inferiore; undici - dotto venoso; 12 - vena porta del fegato. io - fegato; II - vena cava inferiore; III - atrio destro; IV - atrio sinistro; V - aorta discendente
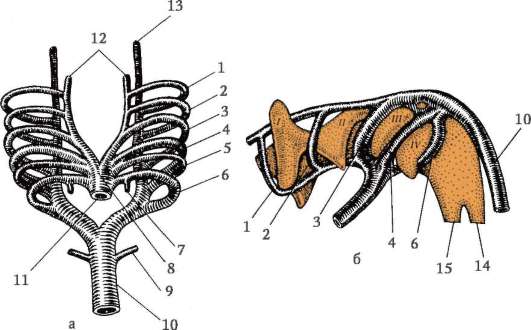
Riso. 7.40. Sviluppo degli archi aortici: a - vista dal lato ventrale: 1-6 - archi aortici; 7 - arteria polmonare; 8 - sacco arterioso, aorta addominale; 9 - settima arteria intersegmentale, succlavia; 10 - aorta discendente dorsale; 11 - tronco arterioso; 12 - arteria carotide esterna, radice dell'aorta ascendente addominale; 13 - arteria carotide interna, radice aortica dorsale
Il quarto arco destro diventa la parte prossimale dell'arteria succlavia destra. La parte distale di questa arteria origina dall'aorta dorsale destra. La parte prossimale del sesto arco sinistro si trasforma nella parte prossimale dell'arteria polmonare sinistra e la parte distale nel dotto arterioso (botall). La parte prossimale del sesto arco destro forma la parte prossimale dell'arteria polmonare destra e la parte distale è ridotta. Le radici dell'aorta dorsale si fondono molto presto nell'aorta dorsale spaiata, ma alla 7a settimana di sviluppo, la parte distale della radice dell'aorta dorsale destra di solito si unisce nell'arteria succlavia destra, come già menzionato sopra. Ciò diventa possibile grazie allo sviluppo inverso del sito che collega la radice destra a sinistra. La presentazione di materiale relativo allo sviluppo del cuore e delle grandi arterie offre motivi di confronto
Riso. 7.41. Formazione delle arterie maggiori, derivati del tronco arterioso, del sacco aortico, degli archi aortici e dell'aorta dorsale: a - embrione di 6 settimane; b - feto di 7 settimane; c - feto di 6 mesi: 1-6 - archi aortici; 7 - tronco arterioso; 8 - arteria polmonare; 9 - arteria succlavia; 10 - aorta discendente dorsale; 11 - sacco aortico; 12 - arteria carotide interna; 13 - dotto arterioso (botall); 14 - aorta ascendente; 15 - tronco polmonare; 1b - arteria carotide esterna; 17 - arteria carotide comune
dell'ontogenesi umana con lo sviluppo filogenetico dei vertebrati (vedi capitolo 14), e ci permette anche di avvicinarci alla comprensione dei meccanismi di insorgenza di malformazioni congenite del cuore e dei vasi sanguigni.
Domande per l'autocontrollo
1. Quali tipi di ontogenesi esistono? Elencare e descrivere le fasi principali dell'ontogenesi.
2. Quali sono le caratteristiche della struttura e delle funzioni delle cellule germinali?
3. Come avviene la fecondazione e quali fasi comprende questo processo?
4. Descrivere l'essenza e le forme morfologiche delle fasi dello sviluppo embrionale in rappresentanti di classi diverse
cordati (frantumazione, gastrulazione, organogenesi e formazione di organi profilattici).
5. Quali sono le caratteristiche dello sviluppo embrionale dei mammiferi?
6. Fornisci esempi dello sviluppo embrionale degli organi umani, che riflettono l'evoluzione dei gruppi ancestrali di cordati.
L'ontogenesi è lo sviluppo individuale di un organismo, un insieme di successive trasformazioni morfologiche, fisiologiche e biochimiche subite da un organismo, dalla fecondazione (durante la riproduzione sessuata) o dal momento della separazione dall'individuo materno (durante la riproduzione asessuata) fino alla fine della vita.
Assegna periodi pre-embrionali, embrionali e post-embrionali. Negli animali placentari e nell'uomo si distinguono i periodi prenatale (antenatale) e postnatale (postnatale).
La totalità degli eventi che si verificano nell'ovo(oo)genesi, ma nell'interesse del processo di sviluppo individuale di un nuovo organismo, costituisce il contenuto del periodo di progenesi - pre-embrionale periodo di sviluppo individuale. Inizia prima del momento della fecondazione e della formazione di uno zigote ed è associato principalmente alla cellula germinale femminile. Quest'ultimo nel corso della gametogenesi (ovogenesi) acquisisce alcune caratteristiche che verranno utilizzate non da sé, ma dall'individuo della nuova generazione che ha iniziato lo sviluppo individuale. Una di queste caratteristiche, forse la più nota, è la formazione nel citoplasma dell'uovo di una quantità maggiore o minore di tuorlo, a seconda del tipo di animale, che viene utilizzato come materiale nutritivo nello sviluppo della prole . L'attività genetica funzionale di un certo numero di geni, che si manifesta nella loro trascrizione e nei cambiamenti post (post) trascrizionali nella trascrizione primaria e (m) dell'RNA, è anche correlata nel tempo al periodo precedente la fecondazione. L'i(m)RNA formatosi come risultato di questa attività organizza la sintesi di proteine importanti per le prime fasi dell'embriogenesi.
Tipi di uova.
Sono classificati in base alla quantità e alla distribuzione del tuorlo. Può essere:
1. Uova alecitali - tuorlo assente o molto piccolo
Esempio: nei mammiferi
2. Isolecithal - c'è poco tuorlo, è uniformemente distribuito nel citoplasma
Esempio: lancetta
3. Polylecital: a) telolecital - il tuorlo è spostato verso il polo vegetativo
b) centrolecithal - tuorlo al centro.
Fecondazione
Fusione di uno spermatozoo con un uovo. Pietre miliari processo di fecondazione: 1) penetrazione dello sperma nell'ovulo; 2) attivazione di vari processi sintetici nell'uovo; 3) fusione dei nuclei dell'uovo e dello spermatozoo con il ripristino dell'insieme diploide dei cromosomi.
La durata della vita e la capacità di fecondare ovuli e spermatozoi sono limitate. Nella maggior parte dei mammiferi, l'uovo conserva la capacità di fecondare per 24 ore e negli esseri umani 12-24 ore dopo l'ovulazione. Gli spermatozoi conservano la loro capacità fecondante nel tratto genitale femminile anche per diverse ore.
Il contatto diretto dello spermatozoo e dell'ovulo provoca cambiamenti regolari in entrambe le cellule, che provocano la penetrazione del nucleo e del centriolo del gamete maschile nell'ovulo e l'attivazione del metabolismo di quest'ultimo. Questi cambiamenti da parte dello spermatozoo sono chiamati reazione acrosomiale e da parte dell'uovo reazione corticale. L'essenza della reazione acrosomiale è la seguente. Al momento del contatto con l'uovo nella parte superiore della testa dello spermatozoo, la membrana plasmatica e la parte della membrana della vescicola acrosomiale adiacente ad essa si dissolvono. A causa del rilascio di enzimi dal granulo acrosomiale, la porzione adiacente della membrana dell'uovo si dissolve. La membrana acrosomiale sporge verso l'esterno e forma una crescita sotto forma di un tubo cavo. Quest'ultimo si allunga, passa attraverso le membrane dell'uovo ed entra in contatto con la membrana plasmatica dell'uovo. Nell'area di tale contatto si verifica una sporgenza o tubercolo di fecondazione, dopodiché le membrane plasmatiche di entrambi i gameti si fondono e inizia l'unione dei loro contenuti. Da questo momento in poi, lo sperma e l'uovo sono un'unica cellula: lo zigote. La reazione acrosomiale è estremamente rapida.
Attivazione dell'uovo o reazione corticale che si sviluppa a seguito del contatto con uno spermatozoo. Cambiamenti nello strato corticale superficiale dell'ooplasma e formazione della membrana di fertilizzazione. Questa membrana, chiamata anche membrana del tuorlo, si forma staccando la superficie dell'uovo. Sotto di esso si forma uno spazio periorlo, nel quale viene versato il contenuto dei granuli dello strato corticale del citoplasma dell'uovo. La membrana di fecondazione protegge l'ovulo dalla penetrazione di spermatozoi soprannumerari, cioè fornisce un "blocco di polispermia". Il fluido che si accumula nello spazio del periuovo funge da ambiente specifico in cui procede lo sviluppo dell'embrione fino al momento in cui lascia le membrane dell'uovo.
IN embrionale periodo, ci sono tre fasi principali: schiacciamento, gastrulazione e organogenesi primaria. Il periodo embrionale, o germinale, dell'ontogenesi inizia dal momento della fecondazione e continua fino a quando l'embrione emerge dalle membrane dell'uovo. Nella maggior parte dei vertebrati, include gli stadi di zigote, scissione, gastrulazione, isto e organogenesi.
Uno zigote è un embrione unicellulare. Nello zigote si verifica il processo di differenziazione del citoplasma. Si osserva il movimento dell'ooplasma. In esso compaiono aree da cui successivamente si formano presunti (supposti) rudimenti di futuri organi.
Scissione - una serie di successive divisioni mitotiche di un uovo fecondato o iniziato per lo sviluppo. La scissione è il primo periodo dello sviluppo embrionale, che è presente nell'ontogenesi di tutti gli animali pluricellulari e porta alla formazione di un embrione chiamato blastula (embrione a uno strato). Allo stesso tempo, la massa dell'embrione e il suo volume non cambiano, cioè rimangono gli stessi dello zigote e l'uovo si divide in cellule sempre più piccole: i blastomeri. Dopo ogni divisione di scissione, le cellule dell'embrione diventano sempre più piccole, cioè le relazioni nucleare-plasma cambiano: il nucleo rimane lo stesso e il volume del citoplasma diminuisce. Il processo procede fino a quando questi indicatori raggiungono i valori caratteristici delle cellule somatiche. Il tipo di schiacciamento dipende dalla quantità di tuorlo e dalla sua posizione nell'uovo. Se c'è poco tuorlo ed è distribuito uniformemente nel citoplasma (uova isolecitali: echinodermi, platelminti, mammiferi), la frantumazione procede in base al tipo di uniforme completa: i blastomeri hanno le stesse dimensioni, l'uovo intero viene schiacciato. Se il tuorlo è distribuito in modo non uniforme (uova telolecitali: anfibi), allora la frantumazione procede secondo il tipo di disomogeneità completa: i blastomeri sono di dimensioni diverse, quelli che contengono il tuorlo sono più grandi, l'uovo è schiacciato nel suo insieme. Con la frantumazione incompleta, c'è così tanto tuorlo nelle uova che i solchi di frantumazione non possono separarlo completamente. La scissione di un uovo, in cui è schiacciato solo il "cappello" del citoplasma concentrato sul polo animale, dove si trova il nucleo dello zigote, è chiamata discoidale incompleta (uova telolecitali: rettili, uccelli). Con uno schiacciamento superficiale incompleto nelle profondità del tuorlo, si verificano le prime divisioni nucleari sincrone, che non sono accompagnate dalla formazione di confini intercellulari. I nuclei, circondati da una piccola quantità di citoplasma, sono uniformemente distribuiti nel tuorlo. Quando ce ne sono abbastanza, migrano nel citoplasma, dove poi, dopo la formazione dei confini intercellulari, appare il blastoderma (uova centrolecitali: insetti).
· Gastrulazione - il processo di movimento cellulare, accompagnato da crescita, riproduzione, differenziazione. L'embrione in questa fase è chiamato gastrula.
Inizialmente, si formano i fogli esterno (ectoderma) e interno (endoderma). Successivamente, appare un terzo strato germinale medio (mesoderma), situato nel corpo dell'embrione tra l'ectoderma e l'endoderma.
Ci sono 4 modi principali per formare i fogli esterni e interni. Spesso, tuttavia, esiste una combinazione di diversi modi di gastrulazione.
Invaginazione a 1 via, che sta nel fatto che una certa sezione del blastoderma, pur mantenendo la struttura dello strato, è avvitata nel blastocele. Quindi il blastocele scompare e appare il gastrocele. L'apertura attraverso la quale questa cavità comunica con l'ambiente esterno è detta bocca primaria o blastoporo.
Epibolia a 2 vie– fouling di macromeri con micromeri in rapida divisione del polo animale. Negli embrioni di tali animali inizialmente non si forma il blastopore ed è assente il gastrocele.
Immigrazione a 3 vie– calcolo di una parte delle cellule della parete della blastula all'interno del blastocele.
Delaminazione a 4 vie - le cellule all'esterno vengono trasformate in uno strato
a-ectoderma; b-endoderma; v-blastocele.
La deposizione del terzo strato germinale avviene in due modi: teloblastico ed enterocelo.
Teloblastico - un metodo di formazione del mesoderma nei protostomi separando due strisce mesodermiche da due cellule embrionali primarie (teloblasti).
Enterocelo - consiste nel fatto che si formano sporgenze dall'endoderma dell'intestino primario su entrambi i lati - tasche (sacche celomiche), che successivamente si allacciano e crescono tra l'ecto - e l'endoderma, formando il mesoderma; le loro cavità si fondono l'una con l'altra e si forma una cavità corporea secondaria, o celoma.
Isto- e organogenesi
Si possono distinguere due fasi.
1 neurulazione, consiste nella formazione di un complesso di organi assiali: il tubo neurale, l'accordo. Un embrione allo stadio di neurulazione è chiamato neurula. In primo luogo, si verifica l'appiattimento dello strato cellulare, che porta alla formazione della placca neurale, i cui bordi, sollevandosi, formano le pieghe neurali. A causa del movimento delle cellule lungo la linea mediana della placca neurale, si verifica una depressione. In questa fase della neurulazione, il segnalibro sistema nervoso detto solco neurale. Allo stesso tempo, la placca neurale sembra piegarsi lungo la linea mediana e poco dopo i suoi bordi si chiudono. Come risultato di questi processi, appare un tubo neurale con una cavità: un neurocele. la chiusura delle creste avviene prima al centro e poi nella parte posteriore del solco neurale. Questo accade per ultimo nella parte della testa, che è più ampia rispetto ad altre aree. La sezione anteriore espansa nell'ulteriore sviluppo forma il cervello, il resto del tubo neurale - il midollo spinale.
2 isto- e organogenesi.
Dall'ectoderma si formano l'epidermide della pelle e i suoi derivati, le ghiandole, l'epitelio della cavità orale, l'epitelio vaginale, lo smalto dei denti e le cellule nervose del recettore.
Dall'endoderma si formano l'epitelio delle vie aeree e dei polmoni, parte delle cellule del pancreas, le cellule secernenti, l'epitelio dello stomaco, l'intestino.
Dal mesoderma: la cartilagine e lo scheletro osseo, i muscoli, i reni, i vasi sanguigni.