ВКУС И ОБОНЯНИЕ
X. Альтнер, И. Бекх
13.1. Характеристика химических ощущений
Ощущения вкуса и запаха обусловлены избирательной и высокочувствительной реакцией специализированных сенсорных клеток на присутствие молекул определенных соединений. В более широком смысле специфические реакции на химические вещества, например гормоныили нейромедиаторы, характерны для многих клеток и тканей. Однако вкусовые и обонятельные сенсорные клетки действуют как экстероцепторы; их реакции дают важную информацию о внешних стимулах, обрабатываемую особыми участками мозга, которые и отвечают за соответствующие ощущения. Другие хеморецепторные клетки служат интероцепторами, определяющими, например, уровень СО 2 (разд. 21.6).
Вкус и запах можно охарактеризовать и различить на основании морфологических и физиологических критериев. Различия между двумя этими видами ощущений наиболее очевидны при сравнении типов (качеств) адекватных для них стимулов (табл. 13.1). Другие характеристики, например чувствительность к стимулам или физическое состояние последних, хотя в целом и неодинаковы, но могут и взаимоперекрываться.
По сравнению с другими ощущениями у вкуса и обоняния значительно выше адаптируемость (ср. рис. 8.5). При длительном воздействии стимула возбуждение в афферентных путях заметно ослабляется, соответственно ослабляется и восприятие; например, уже очень быстро в среде даже с сильным запахом мы перестаем его ощущать. В равной мере характерна для химических ощущений и высокая чувствительность к определенным стимулам. Вместе с тем диапазон различаемых интенсивностей стимуляции сравнительно невелик (1:500), а порог различения высок. Показатель в степенной функции Стивенса ψ = k (φ - φо) a равен 0,4–0,6 для запаха и около 1–для вкусовых стимулов (ср. рис. 8.14).
Первичные процессы и химическая специфичность .
Первое событие при стимуляции хеморецепторов это, согласно современным представлениям, химическое взаимодействие, основанное на слабом связывании адекватной молекулы с рецепторным белком. Из вкусовых органов выделены белки с ферментативными свойствами, субстратная
специфичность и кинетические особенности которых такие же, как у самих рецепторов. Последующие события, приводящие к электрической реакции клеточной мембраны, неизвестны. Каждая рецепторная клетка высокоизбирательно реагирует на специфическую группу веществ. Малейшие изменения в структуре молекулы могут изменить характер ее восприятия или сделать ее неадекватным стимулом. Стимулирующая эффективность соединения, вероятно, наиболее существенно зависит от его размера (например, длины цепи) и внутреннего распределения электрических зарядов (т. е. расположения функциональных групп). Однако тот факт, что во многих случаях молекулы веществ, сильно различающихся по химическому строению, вызывают одинаковые обонятельные ощущения, пока не получил объяснения. Например, три приведенных ниже вещества, несмотря на их структурные различия, обладают одинаковым мускусным запахом (см. Beets в ).
Высказано предположение, что хеморецепторы содержат рецепторные центры, специфичные в отношении определенных групп веществ. Такая точка зрения подтверждается случаями частичной аносмии, т. е. нечувствительности к запаху некоторых, очень близких химических соединений. Сходным образом можно интерпретировать избирательное действие некоторых лекарственных препаратов на орган вкуса. Нанесение на язык гимнемата калия (вещества, выделенного из индийского растения Gymnema silvestre ) приводит к утрате только восприятия сладкого–сахар вызывает во рту ощущение песка. Белок, содержащийся в плодах западно–африканского растения Synsepalium dulcificum , изменяет кислый вкус на сладкий, так что лимон воспринимается как апельсин (см. Kurihara в ). Нанесение на язык кокаина вызывает последовательную утрату всех четырех типов вкусовых ощущений: горького, сладкого, соленого и, наконец, кислого.
Таблица 13.1. Классификация и характеристика химических чувств
|
Вкус |
Запах |
|
|
Рецепторы |
Вторичные сенсорные клетки |
Первичные сенсорные клетки; окончания |
|
Локализация рецепторов |
Язык |
V, IX и Х черепно–мозговых нервов, Нос и глотка |
|
Афферентные черепно–мозговые нервы |
VII, IX |
I, V, IX, Х |
|
Уровни синаптических переключении в ЦНС |
1.Продолговатый мозг 2.Вентральный таламус 3. Кора (постцентральная извилина) C вязи с гипоталамусом |
1. Обонятельная луковица 2.Конечный мозг (препириформная кора) Связи с лимбической системой и гипоталамусом |
|
Адекватные стимулы |
Молекулы органических и неорганических веществ, главным образом нелетучих. Источник стимулов – вблизи рецепторов или в непосредственном контакте с ними |
Почти исключительно молекулы органических летучих веществ в газовой фазе, растворяющиеся только вблизи рецептора. Источник стимулов, как правило, удален |
|
Число качественно различимых стимулов |
Мало (4 основных) |
Очень большое (тысячи) множество нечетко очерченных качеств |
|
Абсолютная чувствительность |
Сравнительно низкая (Не менее 10 16 молекул в 1 мл раствора) |
Очень высокая для некоторых веществ (10 7 молекул, у животных–до 10 2 –10 3 молекул в 1 мл воздуха) |
|
Контактное чувство. Используется для оценки качества пищи, регуляции ее потребления и переваривания (слюноотделительные рефлексы) |
Дистантное чувство. Используется для гигиенической оценки окружающей среды и пищи; у животных–для поисков пищи, коммуникации и в половом поведении. Включает сильный эмоциональный компонент |
У взрослых сенсорные вкусовые клетки расположены на поверхности языка. Вместе с опорными клетками они образуют в эпителии его сосочков группы из 40–60 элементов–вкусовые почки (рис. 13.1). Крупные, окруженные валиком сосочки в основании языка содержат до 200 вкусовых почек каждый; в более мелких грибовидных и листовидных сосочках на его передней и боковых поверхностях их лишь по нескольку. Всего у взрослого человека несколько тысяч вкусовых почек. Железы, расположенные между сосочками, выделяют омывающую вкусовые почки жидкость. Чувствительные к стимуляции дистальные части рецепторных (сенсорных) клеток образуют микроворсинки, выходящие в общую камеру, которая через пору на поверхности сосочка сообщается с внешней средой (рис. 13.1). Стимулирующие молекулы, диффундируя через эту пору, достигают вкусовых клеток (рецепторов).
Подобно прочим вторичным сенсорным клеткам, вкусовые генерируют при стимуляции рецепторный потенциал. Это возбуждение синаптически передается афферентным волокнам
Рис. 13.1. Схема вкусовой почки, погруженной в сосочек языка; показаны базальная, сенсорные, опорные клетки и афферентные волокна соответствующего черепномозгового нерва
черепно–мозговых нервов, которые проводят его в мозг в виде импульсов. В этом процессе участвуют: барабанная струна–ветвь лицевого нерва (VII), иннервирующая переднюю и боковые части языка, и языкоглоточный нерв (IX), иннервирующий его заднюю часть (рис. 13.2). Разветвляясь, каждое афферентное волокно получает сигналы от рецепторов разных вкусовых почек.

Рис. 13.2. Схема языка человека. Окраской выделена его иннервация различными черепно–мозговыми нервами; контурами обведены области распределения разных типов сосочков (1–грибовидные, 2–окруженные валиком, 3–листовидные). Локализация зон восприятия определенных вкусовых качеств показана значками
Замещаются вкусовые клетки очень быстро; продолжительность жизни каждой из них всего около 10 дней, после чего из базальной клетки формируется новый рецептор. Он устанавливает связь с афферентными волокнами таким образом, что их специфичность не меняется. Механизм, обеспечивающий такое взаимодействие, до сих пор не ясен (см. Oakley в ).
Реакции клеток в волокон . Одиночная вкусовая клетка в большинстве случаев реагирует на вещества, представляющие различные вкусовые качества, деполяризуясь или гиперполяризуясь ими (рис. 13.3). Амплитуда рецепторного потенциала повышается вместе с концентрацией стимулирующего вещества. На тип и амплитуду ответа влияет также окружающая среда (рис. 13.4).
Генераторный потенциал вызывает соответствующий уровень возбуждения афферентных волокон, формируя реакцию, называемую «вкусовым профилем» (рис. 13.5). Их импульсация зависит от реакции рецепторов следующим образом: деполяризация последних оказывает возбуждающее, гиперполяризация – тормозное действие.
Многие волокна IX пары черепно–мозговых нервов особенно сильно реагируют на вещества с горьким вкусом. Волокна VII пары сильнее возбуждаются при действии соленого, сладкого и кислого: некоторые из них сильнее реагируют на сахар, чем на соль, другие–на соль, чем на сахар, и т.д. Эти вкусоспецифичные особенности

Рис. 13.3. Внутриклеточные записи рецепторных потенциалов двух вкусовых клеток (а, б) языка крысы. Стимулы: 0,5 моль/л NaCI; 0,02 моль/л солянокислого хинина; 0,01 моль/л HCI и 0,5 моль/л сахарозы. Длительность действия каждого стимула показана горизонтальным отрезком (по Sato , Beidler в с изменениями)

Рис. 13.4. Влияние окружающей среды на форму и амплитуду внутриклеточных записей рецепторного потенциала одиночной вкусовой клетки языка крысы, стимулируемого 0,02 моль/л солянокислого хинина. Окружающая среда: а– 41,4 ммоль/л NaCI; б–дистиллированная вода (по Sato , Beidler в с изменениями)

Рис. 13.5. Ответы двух одиночных волокон барабанной струны крысы на различные вещества: 0,1 моль/л NaCI;
0,5 моль/л сахарозы; 0,01 н. HCI; 0,02 моль/л солянокислого хинина (по с изменениями)
разных групп афферентов обеспечивают информацию о вкусовом качестве, т.е. виде стимулирующей молекулы, а общий уровень возбуждения определенной их популяции–об интенсивности стимула, т. е. о концентрации данного вещества.
Центральные нейроны. Вкусовые волокна VII и IX пар черепно–мозговых нервов оканчиваются в пределах или в непосредственной близости от ядра одиночного пути (nucleus solitarius ) продолговатого мозга. Это ядро через медиальную петлю (медиальный лемниск) связано с таламусом в области его вентрального постеромедиального ядра. Аксоны нейронов третьего порядка проходят через внутреннюю капсулу и оканчиваются в постцентральной извилине коры головного мозга. В результате обработки информации на перечисленных уровнях число нейронов с высокоспецифичной вкусовой чувствительностью возрастает. Ряд корковых клеток реагируют только на вещества с одним вкусовым качеством. Расположение таких нейронов указывает на высокую степень пространственной организации вкусового чувства. Другие клетки в этих центрах реагируют не только на вкус, но и на температурную и механическую стимуляцию языка.
Вкусовая чувствительность человека
Вкусовые качества . Человек различает четыре основных вкусовых качества: сладкое, кислое, горькое и соленое, которые достаточно хорошо характеризуются типичными для них веществами (табл. 13.2). Вкус сладкого ассоциируется главным образом с природными углеводами типа сахарозы и глюкозы; хлорид натрия–соленый; другие соли, например KCI , воспринимаются как соленые и горькие одновременно. Такие смешанные ощущения характерны для многих естественных вкусовых стимулов и соответствуют природе их компонентов. Например, апельсин–кисло–сладкий, а грейпфрут
Таблица 13.2. Вещества с характерным вкусом и эффективность их воздействия на человека (Pfaffmann в )
|
Качество |
Вещество |
Порог восприятия, моль/л |
|
Горькое |
Сульфат хинина |
0,000008 |
|
Никотин |
0,000016 |
|
|
Кислое |
Соляная кислота |
0,0009 |
|
Лимонная кислота |
0,0023 |
|
|
Сладкое |
Сахароза |
0,01 |
|
Глюкоза |
0,08 |
|
|
Сахарин |
0,000023 |
|
|
Соленое |
NaCI |
0,01 |
|
CaCI 2 |
0,01 |
кисло–сладко–горький. У кислот вкус кислый; многие растительные алкалоиды горькие.
На поверхности языка можно выделить зоны специфической чувствительности. Горький вкус воспринимается главным образом основанием языка; другие вкусовые качества воздействуют на его боковые поверхности и кончик, причем эти зоны взаимоперекрываются (рис. 13.2).
Между химическими свойствами вещества и его вкусом не существует однозначной корреляции. Например, не только сахара, но и соли свинца сладкие, а самый сладкий вкус у искусственных заменителей сахара типа сахарина. Более того, воспринимаемое качество вещества зависит от его концентрации. Поваренная соль в низкой концентрации кажется сладкой и становится чисто соленой только при ее повышении. Чувствительность к горьким веществам существенно выше. Поскольку они часто ядовиты, эта их особенность предостерегает нас от опасности, даже если их концентрация в воде или пище очень низка. Сильные горькие раздражители легко вызывают рвоту или позывы на нее. Эмоциональные компоненты вкусовых ощущений широко варьируют в зависимости от состояния организма. Например, человек, испытывающий дефицит соли, считает вкус приемлемым, даже если ее концентрация в пище так высока, что нормальный человек от еды откажется.
Вкусовые ощущения, очевидно, весьма сходны у всех млекопитающих. Поведенческие эксперименты показали, что различные животные различают те же вкусовые качества, что и человек. Однако регистрация активности отдельных нервных волокон выявила и некоторые отсутствующие у человека способности. Например, у кошек обнаружены «водяные волокна», либо реагирующие только на раздражение водой, либо демонстрирующие вкусовой профиль, включающий воду в числе эффективных стимулов (см. Sato в ).
Биологическое значение . Биологическая роль вкусовых ощущений заключается не только в проверке съедобности пищи (см. выше); они также влияют на процесс пищеварения. Связи с вегетативными эфферентами позволяют вкусовым ощущениям влиять на секрецию пищеварительных желез, причем не только на ее интенсивность, но и на состав, в зависимости, например, от того, сладкие или соленые вещества преобладают в пище.
С возрастом способность к различению вкуса снижается. К этому же ведут потребление биологически активных веществ типа кофеина и интенсивное курение.
Поверхность слизистой носа увеличена за счет носовых раковин–гребней, выступающих с боков в просвет носовой полости. Обонятельная область, содержащая большинство сенсорных клеток,

Рис. 13.6. Схема полостей носоглотки человека (сагиттальный разрез). Обонятельная область ограничена верхней и средней раковинами. Показаны зоны, иннервируемые тройничным (V), языкоглоточным (IX) и блуждающим (X) нервами
ограничена здесь верхней носовой раковиной, хотя в средней также есть небольшие островки обонятельного эпителия (рис. 13.6).
Обонятельный рецептор–это первичная биполярная сенсорная клетка, от которой отходят два отростка: сверху – дендрит, несущий реснички, от основания аксон. Реснички, внутренняя структура которых иная, чем у обычных киноцилий, погружены в слой покрывающей обонятельный эпителий слизи и не способны активно двигаться. Пахучие вещества, приносимые вдыхаемым воздухом, вступают в контакт с их мембраной–наиболее вероятным местом взаимодействия между стимулирующей молекулой и рецептором. Аксоны, направляющиеся в обонятельную луковицу, объединены в пучки (fila olfactoria ). Во всей слизистой носа находятся, кроме того, свободные окончания тройничного нерва, и некоторые из них также реагируют на запахи. В глотке обонятельные стимулы способны возбуждать волокна языкоглоточного и блуждающего нервов (рис. 13.6). Слой слизи, покрывающий обонятельный эпителий, предохраняет его от высыхания и постоянно возобновляется за счет секреции и перераспределения киноцилиями.
Молекулы пахучих веществ поступают к рецепторам (сенсорным клеткам) периодически: во время вдоха ноздрями и в меньшей степени–из полости рта, диффундируя через хоаны. Таким образом, во время еды у нас возникают смешанные ощущения, в которых сочетаются вкус и запах пищи.
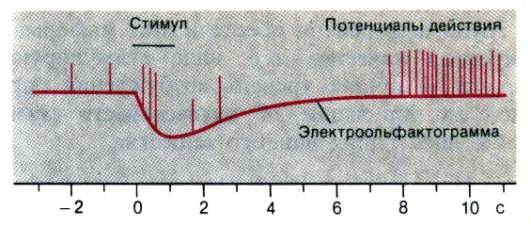
Рис. 13.7. Одновременная запись электроольфактограммы (красная линия) и потенциалов действия одиночного рецептора обонятельного эпителия лягушки при стимуляции нитробензолом. Длительность стимула (черный отрезок) –1 с (Gesteland в )
Обнюхивание–характерное поведение многих млекопитающих – значительно увеличивает приток воздуха к обонятельной слизистой и, следовательно, концентрацию в ней стимулирующих молекул.
Всего у человека в обонятельной области площадью примерно 10 см 2 около 10 7 рецепторов. У других позвоночных их бывает намного больше (у немецкой овчарки, например, 2,2–10 8). Обонятельные клетки, как и вкусовые, постоянно замещаются и из–за этого, по–видимому, не все функционируют одновременно.
Электроды, помещенные на обонятельный эпителий позвоночных, при действии запаха регистрируют медленные потенциалы сложной формы с амплитудой несколько милливольт. Эти электроольфактограммы (ЭОГ, рис. 13.7, см. Ottoson в ), как и электроретинограммы (ЭРГ), отражают суммарную активность многих клеток, поэтому не дают информации о свойствах индивидуальных рецепторов. Записать активность одиночного рецептора в обонятельной слизистой позвоночных удавалось только случайно (рис. 13.7). Показано, что спонтанная активность этих клеток очень низка (несколько импульсов в секунду), и каждая из них реагирует на множество веществ. Как и в случае вкусового профиля, можно построить спектр ответов одиночного обонятельного рецептора (см. Gesfeland в ).
Человек способен различать запах тысяч различных веществ. Обонятельные ощущения можно классифицировать на основе определенного их сходства, выявив определенные типы, или качества, запаха. Однако сделать это гораздо труднее, чем в случае вкусовых ощущений. Неопределенность категорий очевидна и в том, что классификации, предложенные разными авторами, неодинаковы. Корреляция между химической структурой и качеством запаха еще меньше, чем в случае вкусовых раздражителей. Табл. 13.3 показывает, что классы запахов, как правило, называют по их природным
Таблица 13.3. Отличительные характеристики классов запахов (Amoor , Skramlik )
|
Класс запахов |
Известные типичные вещества |
Сходство с запахом |
“Стандартный” источник |
|
Цветочный |
Гераниол |
Розы |
d –1–β–фенилэтилметилкарбинол |
|
Эфирный |
Бензилацетат |
Груши |
1,2–дихлорэтан |
|
Мускусный |
Мускон |
Мускуса |
3–метилциклопентадекан–1 –он |
|
Камфарный |
Цинеол, камфара |
Эвкалипта |
1,8–цинеол |
|
Гнилостный |
Сероводород |
Тухлых яиц |
Диметилсульфид |
|
Едкий |
Муравьиная кислота, уксусная кислота |
Уксуса |
Муравьиная кислота |
источникам или типичным веществам; каждую категорию можно также охарактеризовать «стандартным» источником.
Нейрофизиологические основы отнесения запахов к тому или иному классу до сих пор не обнаружены. Точка зрения, согласно которой группы, объединяющие близкие по запаху вещества, чем–то отличаются друг от друга, подтверждается случаями частичного нарушения обоняния (частичной аносмии). При таких дефектах (по крайней мере некоторые из них генетической природы) порог восприятия определенных обонятельных стимулов повышается. При этом часто он изменяется для нескольких веществ, относящихся, как правило, к одному и тому же классу запахов. Экспериментальные данные, применяемые для классификации запахов, можно получить при анализе перекрестной адаптации. Она заключается в том, что когда длительное действие какого–либо запаха вызывает повышение порога его восприятия, чувствительность к запаху некоторых других веществ также понижается (рис. 13.8). Изучая количественно такие взаимные изменения порогов, можно построить схему перекрестных адаптационных взаимоотношений (рис. 13.9). Однако и это не достаточно для однозначной и детальной классификации множества пахучих веществ по вызываемым ими ощущениям .
При интерпретации особенностей человеческого обоняния следует учитывать, что к пахучим веществам чувствительны и окончания тройничного нерва в слизистой носа, а также языкоглоточного и блуждающего нервов в глотке. Все они участвуют в формировании обонятельного ощущения (рис. 13.6). Их роль, никак не связанная с обонятельным нервом, сохраняется и при нарушениях функции обонятельного эпителия вследствие, например, инфекции (гриппа), опухолей (и связанных с ними операций на мозге) или черепно–мозговых травм. В подобных случаях, объединяемых термином гипосмия, порог восприятия существенно выше нормальною, однако способность различать запахи снижается лишь незначительно. При гипофизарном гипогонадизме (синдроме Кальмана) обоняние обеспечивается исключительно этими черепно–мозговыми нервами, поскольку при этом врожденном заболевании происходит аплазия обонятельных луковиц. Вредные температурные и химические воздействия могут вызвать обратимую или необратимую острую или хроническую гипосмию или аносмию в зависимости от природы и способа воздействия. Наконец, чувствительность к запахам снижается с возрастом.
Обоняние у человека очень чувствительно, хотя известно, что у некоторых животных этот аппарат еще более совершенен. В табл. 13.4 приведены концентрации двух пахучих веществ, достаточные для того, чтобы вызвать у человека соответствующие ощущения. При действии очень малых их количеств возникающее ощущение неспецифично; лишь после превышения некоторого порогового уровня запах не только выявляется, но и распознается. Например, скатол в низких концентрациях пахнет вполне приемлемо; при более высоких–отталкивающе. Таким образом, необходимо различать порог выявления и порог распознавания запаха.
Такие пороги, определяемые по ответам испытуемых или поведенческим реакциям животных, не позволяют установить чувствительность одиночной сенсорной клетки (рецептора). Однако, зная пространственную протяженность обонятельного органа человека и число рецепторов в его составе, можно вычислить и их чувствительность. Такие расчеты показывают, что одиночная сенсорная клетка деполяризуется и генерирует потенциал действия в ответ на одну, самое большее на несколько молекул пахучего вещества. Конечно, поведенческая реакция требует активации значительного числа рецепторов т.е. превышения определенного критического уровня отношения сигнал/шум в афферентных волокнах.
Кодирование. Кодирование обонятельных стимулов рецепторами пока можно описать только в первом приближении. Во–первых, индивидуальная сенсорная клетка способна реагировать на множество различных пахучих веществ. Во–вторых,

Рис. 13.8. Повышение интенсивности ощущения при увеличении интенсивности стимуляции (пропанолом) без адаптации (черная прямая) и после адаптации к пентанолу (красная кривая с черными треугольниками) (Cain , Engen в с изменениями)

Рис. 13.9. Перекрестные адаптационные взаимоотношения семи пахучих веществ: 1–цитраль, 2–циклопентанон, 3–бензилацетат, 4–сафрол, 5–м–ксилен, 6–метилсалицилат,7–бутилацетат.Реципрокные взаимодействия, как правило, неравносильны. Степень повышения порога восприятия обозначена следующим образом: черные линии–очень большая; красные сплошные линии большая; красные прерывистые линии–умеренная; красный пунктир–слабая (по с изменениями)
Таблица 13.4. Порог выявления для запахов масляной кислоты и бутилмеркаптана (Neuhaus , Stuiver )
|
Вещество г |
Число молекул в 1 мл воздуха |
Концентрация вещества вблизи источника стимула, моль/л |
|
Масляная кислота |
2,4–10 9 |
10 –10 |
|
Бутилмеркаптан |
10 7 |
2,7– 10 –12 |
у различных обонятельных рецепторов (как и у вкусовых) перекрывающиеся профили реакции. Таким образом, каждое пахучее вещество специфически возбуждает целую популяцию сенсорных клеток; при этом концентрация вещества отражается в общем уровне возбуждения.
Центральная обработка обонятельной информации
Обонятельная луковица . Гистологически обонятельная луковица подразделяется на несколько слоев, характеризующихся клетками специфической формы, от которых отходят отростки определенного типа с характерными связями между ними. Основные черты обработки информации здесь следующие: значительная конвергенция сенсорных клеток на митральных; выраженные тормозные механизмы и эфферентный контроль входной импульсации. В клубочковом (гломерулярном) слое аксоны приблизительно 1000 рецепторов оканчиваются на первичных дендритах одной митральной клетки (рис. 13.10). Эти дендриты образуют также реципрокные дендродендритные синапсы с перигломерулярными клетками. Митрально–перигломерулярные контакты–возбуждающие, противоположные по направлению–тормозные. Аксоны перигломерулярных клеток оканчиваются на дендритах митральных клеток соседнего клубочка. Такая организация позволяет модулировать локальный дендритный ответ, обеспечивая аутоторможение и торможение окружающих клеток. Клетки–зерна также образуют дендродендритные синапсы с митральными клетками (в данном случае–с их вторичными деидритами) и тем самым влияют на генерирование ими импульсов. Входы на митральных клетках тоже тормозные, т.е. реципрокные контакты участвуют в аутоторможении. Наконец, клетки–зерна образуют синапсы с коллатералями митральных клеток, а также с эфферентными (бульбопетальными) аксонами различного происхождения. Некоторые из этих центробежных волокон приходят из контралатеральной луковицы через переднюю комиссуру (спайку).
Особенность торможения, вызываемого лишенными аксонов клетками–зернами,–то, что в отличие от типичного торможения по Реншоу они могут активироваться частично, т. е. с пространственным градиентом. Эта

Рис. 13.10. Схема нейронных связей в обонятельной луковице. В клубочках (гломерулах) аксоны обонятельных рецепторов оканчиваются на первичных дендритах (D 1) митральных клеток. Перигломерулярные клетки и клетки–зерна образуют реципрокные синапсы с первичными и вторичными (D 2) дендритами митральных клеток. К–коллатерали. Направление синаптической передачи показано стрелками: возбуждающие влияния –черными, тормозные –красными (по с обобщениями и изменениями)
картина очень сложных взаимодействий вполне сравнима с известной в сетчатке, хотя обработка информации основана на другом принципе клеточной организации . Все описанное выше–лишь грубая схема происходящих в обонятельной луковице событий. Помимо митральных, к вторичным нейронам относятся и разнообразные пучковатые клетки, отличающиеся своими проекциями и медиаторами.
Центральные связи . Аксоны митральных клеток образуют латеральный обонятельный тракт, направляющийся в препириформную кору и пириформную долю. Синапсы с нейронами высших порядков обеспечивают связь с гиппокампом и, через миндалину, с вегетативными ядрами гипоталамуса. Нейроны, отвечающие на обонятельные стимулы, обнаружены также в ретикулярной формации среднего мозга и в орбитофронтальной коре.
Влияние обоняния на другие функциональные системы . Прямая связь с лимбической системой (см. разд. 16.6) объясняет выраженный эмоциональный компонент обонятельных ощущений. Запахи могут вызывать удовольствие или отвращение (гедонические компоненты), влияя соответствующим образом на аффективное состояние организма. Кроме того, нельзя недооценивать значение обонятельных стимулов в регуляции полового поведения, хотя результаты экспериментов на животных, особенно опытов по блокаде обоняния у грызунов, нельзя прямо переносить на человека. На животных также показано, что ответы нейронов обонятельного тракта можно изменить инъекцией тестостерона. Таким образом, на их возбуждение влияют и половые гормоны.
Функциональные нарушения . Помимо гипосмии и аносмии встречаются неправильное восприятие запаха (иаросмия) и обонятельные ощущения в отсутствие пахучих веществ (обонятельные галлюцинации). Причины таких расстройств различны. Например, они могут возникать при аллергических ринитах и травмах головы. Обонятельные галлюцинации неприятного характера (какосмия) наблюдаются главным образом при шизофрении.
Учебники и руководства
1. Beidler LM. (Ed.). Chemical Senses. Part 1. Olfaction. Part 2. Taste. Handbook of Sensory Physiology, Vol. IV, Berlin–Heidelberg–New York, Springer, 1971.
2. PfaffD. (Ed.). Taste. Olfaction and Central Nervous System. New York, Rockfeller University Press, 1985.
Оригинальные статьи и обзоры
3. Breipohl W. (Ed.). Olfaction and Endocrine Regulation, London, IRL Press, 1982.
4. Denton D. A., Coghlan J. P. (Eds.). Olfaction and Taste, Vol. V, New York, Academic Press, 1975.
5. Hayushi Т . (Ed.). Olfaction and Taste, Vol. II, Oxford–London New York–Paris, Pergamon Press, 1967.
6. Kare M. R., Mailer 0. (Eds.). The Chemical Senses and Nutrition, New York–San Francisco London, Academic Press, 1977.
7. Koster E. Adaptation and Cross Adaptation in Olfaction, Rotterdam, Bronder, 1971.
8. Le Magnen J., Mac Lead P. (Eds.). Olfaction and Taste., Vol. VI, London–Washington DC, IRL Press, 1977.
9. Norris D. M. (Ed.). Perception of Behavioral Chemicals, Amsterdam–New York–Oxford, Elsevier/North Holland, 1981.
10. Pfaffman С . (Ed.). Olfaction and Taste, Vol. Ill, New York, Rockfeller University Press, 1969.
11. Sato Т . Receptor potential in rat taste cells. In: Autrum H., Ottoson D., PerlE.R., Schmidt R.F., Shimazu H., Willis W.D. (Eds.). Progress in Sensory Physiology, Vol. 6, p. 1–37, Berlin–Heidelberg–New York–Tokyo, Springer, 1986.
12. Schneider D. (Ed.). Olfaction and Taste, Vol. IV, Stuttgart, Wiss, Verlagsges, 1972.
13. Shepherd G. M. Synaptic organization of the mammalian olfactory bulb. Physiol. Rev. 52, 864, (1972).
14. Van der Starre H. (Ed.). Olfaction and Taste. Vol. VII, London–Washington DC, IRL Press, 1980.
15. Zotterman Y. (Ed.). Olfaction and Taste. Vol. I. Oxford–London–New York Paris, Pergamon Press, 1963.
16. Chemical Senses. London. IRL Press (Published in regular installments).
Почему одно и то же блюдо разными людьми воспринимается по-разному? Например, суп вам кажется замечательным в первозданном виде, а вашей второй половинке хочется его поперчить или посолить. В первом случае речь идет о категории людей «супертейстеров», на языке которых находится много сосочков и вкус кажется полным. Как правило, «супертейстеры» предпочитают мягкую еду, вместо острой, а в кофе любят добавлять сливки. Категория людей с низкой плотностью сосочков – «субтейстеры» - любят острую пищу, «обжигающую» полость рта. Хотя на вкусовую чувствительность оказывают влияние не только сосочки.
Известно, что человеческий мозг различает пять вкусов – кислый, соленый, горький, сладкий и умами (насыщенный экзотический вкус восточной пищи). Однако набор химических веществ, вызывающих эти сигналы, имеет отличия у разных людей. Эксперты отмечают, что люди обладают от 20 до 40 генов, несущих ответственность за детекторы горького вкуса.
Разное восприятие горького, скорее всего, последствие эволюционного давления в разных уголках планеты. Самые ядовитые растения наделены горьким вкусом и кочевые племена, которые контактировали с разными видами растений, со временем получили различные рецепторы.
Жители стран, где распространена малярия, чаще всего обладают геном, делающим их менее чувствительными к определенным горьким веществам, особенно тем, которые содержат цианид. Исследователи считают, что цианид в небольших количествах способен нейтрализовать малярийных насекомых, при этом, не отравляя человека. Кстати, у людей есть естественное отвращение к горечи и некоторым запахам, именно поэтому любимое многими пиво редко кому нравится при первой пробе.
Если хотите понять, кто вы – «супертейстер» или «субтейстер», - положите на язык пищу с синим красителем. Эта синяя краска не способна прилипать к вкусовым сосочкам на языке. Если синевы останется на языке мало – вы «супертейстер», если весь язык посинел, вы – «субтейстер».
Оглавление темы "Вестибулярная сенсорная система. Вкус. Вкусовая чувствительность. Обонятельная сенсорная система. Запах (запахи). Классификация запахов.":1. Вестибулярная сенсорная система. Функция вестибулярной системы. Вестибулярный аппарат. Костный лабиринт. Перепончатый лабиринт. Отолиты.
2. Волосковые клетки. Свойства рецепторных клеток вестибулярного аппарата. Стереоцилии. Киноцилий.
3. Отолитовый аппарат. Отолитовый орган. Адекватные раздражители рецепторов отолитовых органов.
4. Полукружные каналы. Адекватные раздражители рецепторов полукружных каналов.
5. Центральная часть вестибулярной системы. Вестибулярные ядра. Кинетозы.
6. Вкус. Вкусовая чувствительность. Вкусовая сенсорная система. Вкусовая рецепция. Вкусовая пора.
8. Центральный отдел вкусовой системы. Проводящие пути вкусовой чувствительности. Ядра вкуса.
9. Вкусовое восприятие. Обонятельная сенсорная система. Макросматики. Микросматики.
10. Запах (запахи). Классификация запахов. Стереохимическая теория запахов.
Мембрана микроворсинок вкусовых клеток содержит специфические участки (рецепторы), предназначенные для связывания растворенных в жидкой среде полости рта химических молекул. Существует четыре разновидности вкусовых ощущений, или четыре вкусовые модальности: сладкое, кислое, соленое и горькое. Строгой зависимости между химической природой вещества и вкусовым ощущением нет : например, сладким вкусом обладают не только сахара, но и некоторые неорганические соединения (соли свинца, бериллия), а самым сладким веществом является не усваиваемый организмом сахарин. Большинство вкусовых клеток полимодальны, т. е. могут реагировать на стимулы всех четырех вкусовых модальностей.
Присоединение к специфическим рецепторам молекул, обладающих сладким вкусом, активирует систему вторичных посредников аденилатциклазы - циклического аденозинмонофосфата, которые закрывают мембранные каналы ионов калия, и поэтому мембрана рецепторной клетки деполяризуется. Вещества, обладающие горьким вкусом, активируют одну из двух систем вторичных посредников: 1) фосфолипазу С - инозитол-3-фос-фат, что приводит к выходу из внутриклеточного депо ионов кальция с последующим выделением медиатора из рецепторной клетки; 2) специфический G-белок гастдуцин, регулирующий внутриклеточную концентрацию цАМФ, которая управляет катионными каналами мембраны и этим определяет возникновение рецепторного потенциала. Действие на рецепторы молекул, имеющих соленый вкус, сопровождается открытием управляемых натриевых каналов и деполяризацией вкусовой клетки. Вещества, обладающие кислым вкусом, закрывают мембранные каналы для ионов калия, что ведет к деполяризации рецепторной клетки.
Величина рецепторного потенциала зависит от вкусового качества и концентрации химического вещества , действующего на клетку. Возникновение рецепторного потенциала приводит к выделению вкусовой клеткой медиатора, действующего через синапс на афферентное волокно первичного сенсорного нейрона, в котором через 40-50 мс от начала действия стимула повышается частота потенциалов действия. Возникшие в афферентных волокнах нервные импульсы проводятся к ядрам одиночных пучков продолговатого мозга. При повышении концентрации действующего вещества общее количество реагирующих чувствительных волокон возрастает за счет вовлечения высокопороговых афферентов в передачу информации от рецепторов.
Вкусовая чувствительность
Пороги вкусовой чувствительности выявляются поочередным нанесением на поверхность языка растворов веществ, обладающих разными вкусовыми качествами (табл. 17.4). Абсолютным порогом чувствительности считают появление определенного вкусового ощущения, отличающегося от вкуса дистиллированной воды. Вкус одного и того же вещества может восприниматься по-разному в зависимости от его концентрации в растворе; например, при малой концентрации хлорида натрия он ощущается сладким, а при большей концентрации - соленым. Максимальная способность различать концентрацию растворов одного и того же вещества и, соответственно, самый низкий дифференциальный порог вкусовой чувствительности характерны для среднего диапазона концентраций, а при высоких концентрациях вещества дифференциальный порог повышается.
Таблица 17.4. Абсолютные пороги восприятия веществ с характерным вкусом
Абсолютные пороги вкусовой чувствительности индивидуально различаются, но у подавляющего большинства людей самым низким оказывается порог определения веществ с горьким вкусом. Эта особенность восприятия возникла в эволюции, она способствует отказу от употребления в пищу веществ горького вкуса, к которым принадлежат алкалоиды многих ядовитых растений. Вкусовые пороги различаются у одного и того же человека в зависимости от его потребности в тех или иных веществах, они повышаются вследствие длительного употребления веществ с характерным вкусом (например, сладостей или соленостей) или курения, потребления алкоголя, обжигающих напитков. Разные области языка различаются вкусовой чувствительностью к различным веществам, что обусловлено особенностями распределения вкусовых рецепторов. Кончик языка более других областей чувствителен к сладкому, боковые стороны языка - к кислому и соленому, а корень языка - к горькому. Вкусовые ощущения в большинстве случаев мультимодальны и основаны не только на избирательной химической чувствительности вкусовых рецепторных клеток, но и на раздражении пищей терморецепторов и механорецепторов ротовой полости , а также действии летучих компонентов пищи на обонятельные рецепторы .
Определяет формирование вкусовых ощущений, является рефлексогенной зоной. С помощью вкусового анализатора оцениваются различные качества вкусовых ощущений, сила ощущений, которая зависит не только от силы раздражения, но и от функционального состояния организма.
Структурно-функциональная характеристика вкусового анализатора .
Периферический отдел . Рецепторы вкуса (вкусовые клетки с микроворсинками) - это вторичные рецепторы, они являются элементом вкусовых почек, в состав которых входят также опорные и базальные клетки. Во вкусовых почках обнаружены клетки, содержащие серотонин, и клетки, образующие гистамин. Эти и другие вещества играют определенную роль в формировании чувства вкуса. Отдельные вкусовые почки являются полимодальными образованиями, так как могут воспринимать различные виды вкусовых раздражителей. Вкусовые почки в виде отдельных включений находятся на задней стенке глотки, мягком нёбе, миндалинах, гортани, надгортаннике и входят также в состав вкусовых сосочков языка как органа вкуса.
Периферический отдел вкусового анализатора представлен вкусовыми луковицами, которые расположены главным образом в сосочках языка. Вкусовые клетки усеяны на своем конце микроворсинками, которые называют еще вкусовыми волосками. Они выходят на поверхность языка через вкусовые поры.
На вкусовой клетке имеется большое число синапсов, которые образуют волокна барабанной струны и языкоглоточного нерва. Волокна барабанной струны (ветвь язычного нерва) подходят ко всем грибовидным сосочкам, а волокна языкоглоточного нерва - к желобоватым и листовидным. Корковый конец вкусового анализатора находится в гиппокампе , парагиппокамповой извилине и в нижней части заднецентральной извилины .
Вкусовые клетки непрерывно делятся и непрерывно гибнут. Особенно быстро происходит замещение клеток, расположенных в передней части языка, где они лежат более поверхностно. Замена клеток вкусовой почки сопровождается образованием новых синаптических структур
Проводниковый отдел . Внутрь вкусовой почки входят нервные волокна, которые образуют рецепторно-афферентные синапсы. Вкусовые почки различных областей полости рта получают нервные волокна от разных нервов: вкусовые почки передних двух третей языка - от барабанной струны, входящей в состав лицевого нерва; почки задней трети языка, а также мягкого и твердого нёба, миндалин - от языкоглсточного нерва; вкусовые почки, расположенные в области глотки, надгортанника и гортани, - от верх-пегортанного нерва, являющегося частью блуждающего нерва.
Эти нервные волокна являются периферическими отростками биполярных нейронов, расположенных в соответствующих чувствительных ганглиях, представляющих первыйнейрон проводникового отдела вкусового анализатора. Центральные отростки этих клеток входят в состав одиночного пучка продолговатого мозга , ядра которого представляют второй нейрон. Отсюда нервные волокна в составе медиальной петли подходят к зрительному бугру (третий нейрон).
Центральный отдел. Отростки нейронов таламуса идут в кору больших полушарий (четвертый нейрон). Центральный, или корковый, отдел вкусового анализатора локализуется в нижней части соматосенсорной зоны коры в области представительства языка. Большая часть нейронов этой области мультимодальна, т. е. реагирует не только на вкусовые, но и на температурные, механические и ноцицептивные раздражители. Для вкусовой сенсорной системы характерно то, что каждая вкусовая почка имеет не только афферентные, но и эфферентные нервные волокна, которые подходят к вкусовым клеткам из ЦНС , благодаря чему обеспечивается включение вкусового анализатора в целостную деятельность организма.
Механизм вкусового восприятия . Чтобы возникло вкусовое ощущение, раздражающее вещество должно находиться в растворенном состоянии. Сладкое или горькое вкусовое вещество, растворяющееся в слюне до молекул, проникает в поры вкусовых луковиц, вступает во взаимодействие с гликокаликсом и адсорбируется на клеточной мембране микроворсинки, в которую встроены «сладкочувствующие» или «горькочувствующие» рецепторные белки. При воздействии соленых или кислых вкусовых веществ изменяется концентрация электролитов около вкусовой клетки. Во всех случаях повышается проницаемость клеточной мембраны микроворсинок, возникает движение ионов натрия внутрь клетки, происходят деполяризация мембраны и образование рецеп-горного потенциала, который распространяется и по мембране, и по микротубулярной системе вкусовой клетки к ее основанию. В это время во вкусовой клетке образуется медиатор (ацетилхолин, серотонин, а также, возможно, гормоноподобные вещества белковой природы), который в рецепторно-афферентном синапсе ведет к возникновению генераторного потенциала, а затем потенциала действия во внесинаптических отделах афферентного нервного волокна.
Восприятие вкусовых раздражителей. Микроворсинки вкусовых клеток являются образованиями, непосредственно воспринимающими вкусовой раздражитель. Мембранный потенциал вкусовых клеток колеблется от -30 до -50 мВ. При действии вкусовых раздражителей возникает рецепторный потенциал величиной от 15 до 40 мВ. Он представляет собой деполяризацию поверхности вкусовой клетки, которая является причиной возникновения в волокнах барабанной струны и языкоглоточного нерва генераторного потенциала, переходящего по достижении критического уровня в распространяющиеся импульсы. С рецепторной клетки возбуждение передается через синапс на нервное волокно с помощью ацетилхолина. Некоторые вещества, как например СаС1 2 , хинин, соли тяжелых металлов, вызывают не первичную деполяризацию, а первичную гиперпрляризацию. Ее возникновение связано с осуществлением отрицательных отвергаемых реакций . Распространяющихся импульсов при этом не возникает.
Чувствительность рецепторов к разным видам вкусовых раздражений .
Различные вкусовые клетки обладают разной чувствительностью к различным вкусовым веществам, которые делятся на четыре группы: кислое, соленое, сладкое, горькое. Каждая клетка отвечает всегда более чем на одно вкусовое вещество, иногда даже на все четыре, но наибольшей чувствительностью обладает к одному из них. Соответственно в зависимости от расположения клеток с особо высокой чувствительностью к тому или иному вкусовому раздражителю разные участки языка обладают также разной чувствительностью.
Установлено, что кончик языка и передняя его треть наиболее чувствительны к сладкому, где расположены грибовидные сосочки, боковые поверхности - к кислому и соленому (листовидные сосочки), а корень языка - к горькому (желобоватые сосочки, или вкусовые сосочки, окруженные валом).
Для вкусовых клеток характерны колебания порога раздражения и различный в разных условиях характер ответа на одни и те же раздражители . Их возбудимость зависит от постоянных влияний друг на друга, а также от состояния рецепторов пищеварительного тракта, обонятельных и др. В норме существует определенная «настройка» вкусовых рецепторов в соответствии с состоянием организма, в частности с состоянием сытости.
Отправить свою хорошую работу в базу знаний просто. Используйте форму, расположенную ниже
Студенты, аспиранты, молодые ученые, использующие базу знаний в своей учебе и работе, будут вам очень благодарны.
Размещено на http://www.allbest.ru/
Кафедра Физиологии
Физиология вкуса
Введение
1. Морфология органов вкуса; субъективная физиология вкуса. Ориентация и строение вкусовых почек
2. Центральные связи
3. Основные вкусовые ощущения
4. Интенсивность ощущений
5. Объективная физиология вкуса
6. Первичный процесс
7. Роль вкусовой чувствительности
Литература
Введение
Человек и животное непрерывно получают информацию о бесконечном многообразии изменений, которые происходят во внешней и внутренней среде. Это осуществляется благодаря наличию у организма специализированных структур, которые получили название анализаторы (сенсорные системы).
Под анализаторами понимают совокупность образований, обеспечивающих восприятие энергии раздражителя, трансформацию ее в специфические процессы возбуждения, проведение этого возбуждения в структуры ЦНС и к клеткам коры, анализ и синтез специфическими зонами коры этого возбуждения с последующим формированием ощущения.
Понятие об анализаторах введено в физиологию И. П. Павловым в связи с учением о высшей нервной деятельности. Каждый анализатор состоит из трех отделов:
Периферический или рецепторный отдел, который осуществляет восприятие энергии раздражителя и трансформацию ее в специфический процесс возбуждения.
Проводниковый отдел, представленный афферентными нервами и подкорковыми центрами, он осуществляет передачу возникшего возбуждения в кору головного мозга.
Центральный или корковый отдел анализатора, представленный соответствующими зонами коры головного мозга, где осуществляется высший анализ и синтез возбуждений и формирование соответствующего ощущения.
Роль анализаторов при формировании приспособительных реакций чрезвычайно велика и многообразна. Согласно концепции функциональной системы П. К. Анохина формирование любой приспособительной реакции осуществляется в несколько этапов. Анализаторы принимают непосредственное участие в формировании всех этапов функциональной системы. Они являются поставщиками афферентных посылок определенной модальности и различного функционального назначения, причем, одна и та же афферентация может быть обстановочной, пусковой, обратной и ориентировочной в зависимости от этапа формирования приспособительной деятельности.
вкус физиология анализатор орган
1. Морфология органов вкуса; субъективная физиология вкуса. Ориен тация и строение вкусовых почек
Язык у человека покрыт слизистой оболочкой, складки которой во многих местах образуют маленькие выпуклости в форме колышков, называемые сосочками
Эти три типа распределены по-разному. Только грибовидные сосочки рассеяны по всей поверхности.Желобоватые сосочки, которых у человека всего 7-12, сверху имеют вид круглых образований 1-3 мм в диаметре; они находятся в ограниченной зоне поперек спинки языка у его корня. Третий тип, листовидныесосочки, образуют тесно расположенные складки вдоль задних краев языка. Они хорошо развиты у детей, но гораздо менее выражены и менее многочисленны у взрослых.
Нитевидные сосочки, занимающие остальную поверхность языка, не показаны на рис. 1, потому что в них нет вкусовых почек. Название «почка» говорит о форме этих органов (рис. 2). Положение их на сосочках варьирует; в случае желобоватых и листовидных сосочков много вкусовых почек заложено в боковых стенках, а на верхушке их нет. В грибовидных сосочках вкусовые почки ограничены поверхностью «шляпки гриба», которая может достигать 1 мм в диаметре.
Отдельная вкусовая почка имеет около 70 мкм в высоту и около 40 мкм в диаметре. Всего у человека около 2000 вкусовых почек, из них около половины-на желобоватых сосочках. Каждая вкусовая почка содержит 40-60 отдельных клеток.
В соединительную ткань под желобоватыми и листовидными сосочками погружены серозные железы, протоки которых открываются в углубления у основания сосочка, их секрет служит для смывания частиц пищи и микроорганизмов. Кроме того, он понижает концентрацию стимулирующего вещества вблизи вкусовых почек.
Внутри вкусовых почек различают три типа клеток: сенсорные, опорные и базальные (рис. 2). Растворимые в воде вещества, попадающие на поверхность языка, диффундируют через пору в наполненное жидкостью пространство над вкусовой почкой; здесь они соприкасаются с мембранами микровиллей, которые образуют наружные концы сенсорных клеток. Вкусовые рецепторы представляют собой вторичные сенсорные клетки без аксонов, которые проводят импульсы в центральном направлении. Их ответы передаются афферентными волокнами, которые образуют синапсы близ оснований сенсорных клеток. На рис. 2 показаны только два волокна, но в действительности в каждую вкусовую почку входят и разветвляются в ней около 50 волокон.
Продолжительность жизни сенсорных клеток во вкусовых почках невелика; происходит их непрерывная смена. В среднем одна сенсорная клетка замещается новой уже через 10 дней. За сменой клеток можно проследить, помечая их ядра 3Н-тимидином и определяя число меченых ядер, сохранившихся через некоторое время. Утраченные сенсорные клетки замещаются новыми, которые образуются из базальных клеток. При этой смене должны прерваться синапсы между афферентными волокнами и старыми клетками и возникнуть новые синапсы. В связи с такой перестройкой возникает много интересных вопросов, особенно если учесть тот факт, что сенсорные клетки различаются по своей чувствительности к разным стимулам. Так, смена сенсорных клеток может привести к изменению «вкусового профиля» - характерной формы ответов в афферентных волокнах, о чем пойдет речь в следующем разделе.
2. Центральные связи
Афферентные волокна, проводящие ответы от скоплений вкусовых луковиц, распределяются по двум черепномоз-говым нервам -лицевому (VII) и языкоглоточному (IX). Такое деление обычно соответствует областям языка, которые снабжаются этими волокнами. Так, волокна от желобоватых и листовидных сосочков идут преимущественно в составе языкоглоточного нерва, а волокна от грибовидных сосочков в передней части языка входят в барабанную струну (chorda tympani), ветвь лицевого нерва. У детей имеются добавочные вкусовые органы в эпителии мягкого нёба и задней стенки глотки до гортани; они иннервируются главным образом блуждающим нервом (X).
В головном мозгу вкусовые волокна на каждой стороне объединяются в солитарный тракт. Он оканчивается в продолговатом мозгу, в ядре солитарного тракта, где афферентные волокна образуют синапсы с нейронами второго порядка. Аксоны этих нейронов идут к вентральному таламусу в составе медиального лемниска. Третья совокупность нейронов связывает эту область с корой больших полушарий. Вкусовые зоны коры расположены в латеральной части постиентральной извилины.
3. Основные вкусовые ощущения
В обычных условиях, например при еде, слизистая ротовой полости подвергается действию сложных стимулов, включающих несколько модальностей. Благодаря тому, что ротовая полость сообщается с носовой, пахучие вещества могут достичь обонятельных рецепторов в носу и вызвать другие ощущения. Кроме того, в слизистой оболочке рта и языка имеются терморецепторы, механорецепторы и болевые волокна, которые тоже подвергаются стимуляции. То, что обычно называют «вкусом», в действительности является мультимодальным ощущением, в котором на собственно вкусовые ощущения накладываются ощущения запаха, тепла или холода, давления и, возможно, даже боли.
Существуют четыре четко различимых основных вкусовых ощущения: сладкое, кислое, соленое и горькое.
Пороги обнаружения для разных качеств приходятся на разные концентрации. Пороговая концентрация сернокислого хинина (8 мкмоль/л, или 0,006 г/л) служит хорошим примером того, что вещества с горьким вкусом обнаруживаются при очень низких концентрациях. Порог обнаружения для сахарина составляет 23 мкмоль/л (0,0055 г/л), для виноградного сахара-0,08 моль/л, а для тростникового сахара-0,01 моль/л (соответственно 14,41 и 3,42 г/л). Эти данные характерны, и они показывают, что пороги для моно- и дисахаридов значительно выше, чем для синтетических сладостей. Пороги для уксусной кислоты (0,18 моль/л, или 0,108 г/л) и столовой соли (0,01 моль/л, или 0,585 г/л) служат иллюстрацией того общего правила, что пороги для кислого и соленогоприблизительно того же порядка, что и для указанных выше сахаридов. Пороги для кислот приблизительно отражают степень их диссоциации. Сравнение порогов для виноградного и тростникового Сахаров говорит о том, что раствор виноградного сахара должен быть более концентрированным, чем раствор тростникового сахара, для того чтобы они были одинаково сладкими. Экспериментальная проверка растворов разных надпороговых концентраций соответствует этому различию.
Но польза от таких точных пороговых данных ограничена, потому что для большинства веществ пороги подвержены значительной индивидуальной вариабельности. Разумнее было бы говорить о диапазоне пороговых значений
4. Интенсивность ощущений
Простое сравнение разных растворов показывает, что интенсивность вкусового ощущения зависит отконцентрации вещества. При определении порогов обнаружено, что эффект от разбавления раствора стимулирующего вещества может быть компенсирован стимуляцией большей поверхности языка, т.е. большего числа рецепторов Вероятно, это происходит благодаря пространственной суммации. В пороговой области существует входное соотношение между концентрацией и продолжительностью действия стимула. Кроме того, следует помнить, что чувство вкуса подвержено определенной адаптации -при длительном действии стимула интенсивность ощущения снижается. Еще одним фактором является секреция серозных желез, которая разжижает действующее у вкусовых луковиц вещество и тем самым влияет на интенсивность ощущения.
Испытание ряда разведений солевых растворов в околопороговой области во многих случаях показывает, что ощущение может менять свое качество в зависимости от концентрации. Растворы столовой соли 0,02-0,03 моль/л имеют сладкий вкус, а в концентрации 0,04 моль/л или больше -- соленый. Этот сдвиг качества, вероятно, можно понять, исходя из того, что вкусовые волокна обладают широким спектром чувствительности в пределах каждого качества.
Разные области языка у человека варьируют по чувствительности к четырем основным качествам. Кончик языка особенно чувствителен к сладким веществам, средние части краев отвечают лучше всего на кислые стимулы. Соленые стимулы всего эффективнее в области края языка, которая частично перекрывает первые две. Горькие вещества сильнее всего действуют на рецепторы близ корня языка, в области желобоватых сосочков. Поэтому повреждение языкоглоточного нерва понижает способность к обнаружению горечей, а после блокады проведения в лицевом нерве обнаруживаются только они одни.
5. Объективная физиология вкуса
Способность к различению вкусовых качеств зависит от специфичности рецепторных молекул в мембранах сенсорных клеток. Для регистрации активности как отдельных сенсорных клеток, так и афферентных волокон можно воспользоваться микроэлектродами. Такие записи показывают, что ни сами рецепторы, ни волокна, идущие к ЦНС, не дают качественно специфических ответов; как правило, эффективными оказываются стимулы более чем одной категории. Очевидно, что каждое волокно реагирует на стимулы нескольких категорий, но при рассмотрении разных градаций чувствительности выявляются различия. Иными словами, стимуляция раствором вещества в определенной концентрации активирует различные волокна в разной степени. Характер возбуждения, типичный для каждого отдельного волокна при ответах на ряд веществ, называется вкусовым профилем. Волокнами, наиболее близкими к качественной специфичности, являются те, которые реагируют на растворы Сахаров увеличением частоты разрядов. Сравнительные исследования показали, что такие относительно специфичные волокна особенно характерны для обезьян.
Регистрация активности отдельных сенсорных клеток показала, что они обладают градуальной относительной специфичностью. Ответы волокон, идущих от этих клеток, в этом отношении отражают ответы клеток. Но афферентные волокна ветвятся во вкусовых луковицах, так что каждое волокно получает возбуждение от многих сенсорных клеток, которые, надо полагать, различаются по степени специфичности. Кроме того, обнаружено, что сенсорные клетки в разных сосочках образуют синапсы с коллатералями от одного афферентного волокна. Это значит, что вкусовые волокна получают входы от сенсорных клеток, распределенных по значительным участкам языка. Эти участки называются рецептивными полями. Ситуация с рецептивными полями усложняется тем, что отдельные сенсорные клетки могут получать иннервацию от нескольких различных волокон.
Градуальная относительная специфичность вкусовых волокон создается 1) градуальной относительной специфичностью сенсорных клеток и 2) ветвлением вкусовых волокон, создающим рецептивные поля. Частота импульсации в одиночном афферентном волокне поэтому меняется как от качества стимула, так и от его концентрации. Разумеется, важным фактором является также степень, в какой стимулируемая область покрывает рецептивное поле волокна. Очевидным выводом в отношении кодирования стимула является то, что активность одного волокна не может дать однозначную информацию о качестве или концентрации. Толькосравнение уровня возбуждения в нескольких волокнах может выявить характерные распределения (паттерны) активности, которые говорят что-то о качестве стимула. При условии, что качество известно, частота импульсации в каждом отдельном волокне может служить мерой концентрации стимулирующего вещества. Отличительные черты вещества, следовательно, кодируются таким образом, что сложный, но характерный паттерн возбуждения создается одновременными, но разными ответами множества нейронов.
6. Первичный процесс
Условием возбуждения вкусового рецептора является взаимодействие между молекулами стимулирующего вещества и специально дифференцированными точками в мембране сенсорной клетки, где лежатрецепторные молекулы. Это взаимодействие называется первичным процессом; как полагают, он начинается с адсорбции молекулы вещества-стимула. Предполагают, что, когда это происходит, рецепторная-вероятно белковая-молекула изменяет свою структуру. Такое конформационное изменение рецепторной молекулы приводит в свою очередь к локальному изменению проницаемости мембраны клетки. Этот клеточный «усилительный механизм» мог бы послужить причиной генерации рецепторного потенциала.
К свидетельствам существования специфических рецепторных молекул относится наблюдение, что некоторые растительные вещества и препараты, например кокаин и гимневая кислота (получаемая из индийского растения Gymnema sylvestre), избирательно блокируют некоторые вкусовые ощущения. Очевидно, эта кислота связывается с рецепторными молекулами для сладких веществ, поскольку ее нанесение делает такие вещества безвкусными. Первичный процесс в мембранах вкусовых сенсорных клеток еще по-настоящему не объяснен, но, согласно рабочей гипотезе, он сходен с процессом в холинергических синапсах, где особые молекулы меняют проницаемость в особых точках мембраны.
7. Роль вкусовой чувствительности
Вкусовые луковицы на языке реагируют на стимулы, локализованные во рту. Иными словами, вкусовая чувствительность у всех позвоночных участвует в ориентации на близком расстоянии. В то же время у рыб чувство вкуса может служить также ориентации на далеком расстоянии. В воде вкусовые вещества перемещаются благодаря диффузии и конвекции из очень далеких источников к вкусовым луковицам, которые могут быть рассеяны по всей поверхности тела рыбы.
Помимо своей роли в ориентации на близком расстоянии чувство вкуса у человека выполняет важную функцию, запуская ряд рефлексов. Например, отмывание языка секретом из серозных желез контролируется рефлексом, который находится под действием вкусовых луковиц. Секреция слюны тоже запускается рефлекторно соответствующей стимуляцией вкусовых рецепторов. Даже состав слюны варьирует в зависимости от характера стимулов, действующих на сенсорные клетки, и вкусовые стимулы влияют также на выделение желудочного сока. Наконец, доказано, что рвота вызывается при участии вкусовой чувствительности.
Литература
1. Батуев А.С., Куликов Г.Л. Введение в физиологию сенсорных систем. -- М.: Высшая школа, 1983. -263 с.
2. Лекции по физиологии центральной нервной системы: Учебное пособие. Биолого-химический факультет УдГУ, Проничев И.В. -- Powered by swift.engine.edu, 2003. - 162 с.
3. Шульговский В. В. Основы нейрофизиологии: Учебное пособие для студентов вузов. -- М.: Аспект Пресс, 2000. с. 277.
4. Шульговский В. В. Физиология высшей нервной деятельности с основами нейробиологии: Учебник для студ.биол. специальностей вузов.- М.: Издательский центр «Академия», 2003. - 464 с.
Размещено на Allbest.ru
Подобные документы
Неоднородная структура органа вкуса. Около 2000 вкусовых луковиц находится в ткани языка, неба, надгортанника и верхней части пищевода. Большинство из них размещены в слизистой мембране вкусовой луковицы. Нервные волокна и образование вкусовой почки.
реферат , добавлен 02.03.2009
Общая физиология сенсорных систем. Соматосенсорный, вкусовой и обонятельный анализаторы. Определение точек прикосновения. Определение пространственных порогов тактильной рецепции и локализации болевых рецепторов. Определение вкусовых ощущений и порогов.
методичка , добавлен 07.02.2013
Общая характеристика организма собаки, особенности его анатомии и физиологии, функции отдельных органов. Описание основных систем организма: системы костей, мышечной, кожной и нервной. Особенности органов зрения, вкуса, слуха осязания и обоняния.
реферат , добавлен 09.11.2010
Анатомия и физиология сердечно–сосудистой системы. Вены, распределение и ток крови, регулирование кровообращения. Давление крови, кровеносные сосуды, артерии. Определение показателя состояния осанки и плоскостопия у учащихся. Орган вкуса, виды сосочков.
курсовая работа , добавлен 25.12.2014
Изучение особенностей технологии разработки, видов (сироп, инъекции, ингаляции, гранулы, мазь, гель) и состава лекарственных форм для детей. Характеристика методов определения вкуса лекарств, числовых индексов и органолептической оценки корригенов.
реферат , добавлен 27.01.2010
Преддверно-улитковый орган (орган слуха и равновесия): структура и взаимодействие элементов, функции в жизнедеятельности организма человека. Распространение звука в органе слуха. Расположение органа обоняния и вкуса, закономерности их функционирования.
презентация , добавлен 27.08.2013
Строение и физиология сердца, его основные функции. Характеристика схемы и механизма кровообращения. Фазы сердечного цикла, электрическая активность клеток миокарда и параметры центральной гемодинамики. Понятие и особенности процесса иннервации сердца.
презентация , добавлен 12.01.2014
Нормальная физиология. Патологическая физиология. Хронологическая таблица. Классификация по группам и подгруппам. Химическое строение, механизм действия. Источники происхождения и др. Механизм биологической активности препаратов данной группы.
курсовая работа , добавлен 03.07.2008
Изучение анатомии и физиологии ЛОР-органов как дистантных анализаторов. Анатомия уха, носа, глотки, гортани. Физиология носа и придаточных пазух, слухового и вестибулярного анализатора. Дыхательная, защитная и голосообразовательная функции гортани.
реферат , добавлен 29.01.2010
Строение промежуточного мозга. Роль печени и поджелудочной железы в пищеварении. Торможение центральной нервной системы. Анатомия и физиология вегетативной нервной системы, ее возрастные особенности. Состав крови и физико-химические свойства плазмы.