Довольно большую долю массы сухого мицелия грибов составляет их клеточная стенка, а именно ох 5 до 15%. Состав ее сильно варьирует и часто весьма специфичен для определенных таксономических групп грибов. Это можно видеть из примеров ее состава у представителей дрожжей, хитридиевых и мукоровых грибов - зигомицетов, приведенного в табл. 1.3 (Aronson, 1965).
* (Количество золы без фосфатов. )
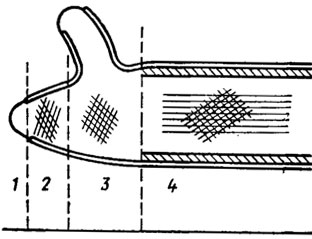
Структурно оболочки грибов построены на основе двухфазной системы, в которой имеются микрофибриллы, включенные в аморфную массу матрикса. По данным электронной микроскопии, она состоит не менее чем из двух слоев с различно направленной ориентацией фибрилл. Внутренний слой обычно ориентирован по главной оси клетки, наружные - под углом к ней (рис. 1.1). У дрожжей оболочка обычно многослойная, причем в наружном толстом слое локализован маннан, во внутреннем тонком - глюкан. У водных грибов, например у Allomyces, оболочка образует ложные перегородки - псевдосепты, напоминающие спицы колеса. У сумчатых и базидиальных грибов наблюдаются настоящие перегородки - септы. У сумчатых септы между клетками обычно имеют одну простую пору, по обе стороны которой в процессе ее формирования можно видеть по паре осмофильных телец Воронина. У базидиальных грибов эти поры нередко бывают очень сложными, снабженными колпачками - парентосомами (рис. 1.2). Такие поры были найдены в плодовых телах и ризоморфах ряда высших базидиомицетов (Мооrе, 1965; Burnett, 1968). Однако до настоящего времени остается не вполне ясным, относить ли это различие в структуре септальных пор аско- и базидиомицетов к их таксономической принадлежности или к гаплоидной и гомокариотической структуре генома сумчатых и к ди- и гетерокариотической природе базидиальных грибов. Таких исследований пока еще нет, но их значение для эволюционных и таксономических построений в царстве грибов весьма принципиальное.

Много нового добавили к изучению септальных пор грибов исследования на электронно-микроскопическом уровне (Flegler et al., 1976; Камалетдинова, Васильев, 1982). Во-первых, стало известно, что долипоры базидиальных грибов являются структурами, видимо, герметически изолирующими клетки мицелия друг от друга вплоть до начала образования ими плодовых тел (Flegler et al., 1976). Эта изоляция осуществляется растворяемыми протеолитическими ферментами и одновременно осмофильными (содержащими белки и липиды) двусторонними пробками, которые исчезают к моменту формирования плодовых тел. Их исчезновение сопровождается перфорациями парентосом и сообщение между клетками гиф восстанавливается.
Последние наблюдения над септами в плодовых телах дискомицетов (например, Peziza badia - Камалетдинова, Васильев, 1982) показали, что сходные разобщающие клетки гиф структуры существуют и у класса сумчатых грибов. В их формировании принимают участие тельца Воронина, образующиеся в колбообразных инвагинациях клеточной мембраны, осмофильное содержимое которых (тельце Воронина) высвобождается близ септы и располагается у отверстия септы, постепенно проникая в нее, создавая при этом замыкающее ее устройство. Дальнейшему проникновению тельца Воронина в вышележащий гимениальный слой, видимо, препятствует специальная перфорированная структура, лежащая над порой септы в материнской клетке будущей сумки, которая оказывается полностью изолированной от проникновения субмикроскопических органелл из субгимениального слоя. Подобные же изолирующие структуры наблюдаются и в формирующихся в плодовых телах парафизах.
Сходные структуры имеются и в спорулирующем мицелии несовершенных грибов, содержащих тельца Воронина, например у Arthrobotrys conoides. У продуцента пенициллина дейтеромицета Penicillium chrysogenum была обнаружена в септе структура, в точности соответствующая найденной Камалетдиновой и Васильевым у дискомицета Peziza badia (Kurilowich et al., 1980).
Скелетные основы оболочек грибов составлены кристаллически организованными полисахаридами: целлюлозой, хитином, хитозаном, маннаном, глюканами и другими. Все они имеют линейную структуру с β-1,4-связями исходных компонентов - мономеров гексоз, амино- и ацетаминогексоз. По результатам микрохимического тестирования (окраска рутениум красным) ранее считалось, что в составе клеточных оболочек грибов имеется пектин. Однако результаты химического анализа не подтвердили в ее составе входящего в структуру пектина мономера галактуроновой кислоты (Aronson, 1965).
Хитин и хитозан для большей части грибов очень характерны в составе их оболочек как азотсодержащие полимеры. При этом хитин грибов очень сходен с хитином насекомых и ракообразных, что было подтверждено картиной их дифракции в рентгеновских лучах. Однако азота в хитине грибов меньше, чем у животных, и среди его составных частей была обнаружена метилпентоза, называемая мицетозой. Хитин у грибов удается обнаружить микрохимически по методу Ван Висселинга, с помощью частичного щелочного деацетилирования и последующей реакцией на хитозан и по дифракции Х-лучей. Он не был найден только у оомицетов, таких, как сапролегниевые и пероноспоровые грибы. Ранее считалось, что хитин отсутствует у дрожжей, однако он содержится в клеточных перегородках - септах сахаромицетов (Кулаев, 1975).
В настоящее время установлено, что хитин можно обнаружить у Chitrydiales, Monoblepharidales, Protomycetales, Hyphochyiridiales, у всех Endomycetales, Blastocladiales, Mucorales, Entomophthorales, у всех сумчатых и базидиальных грибов и происходящих от них Fungi imperfecti (дейтеромицетов). Исключение составляют Oomycetes, у которых из полисахаридов в оболочке присутствует целлюлоза, полностью отсутствующая у представителей дрожжевых грибов.
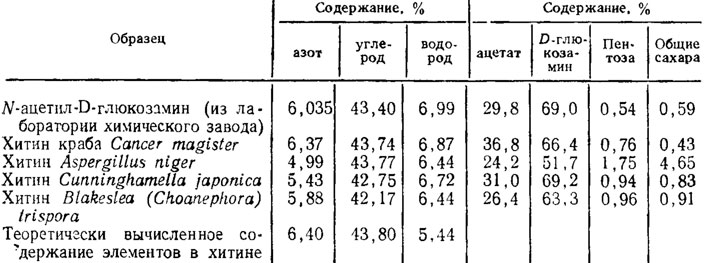
В течение последнего времени в связи с выявлением возможности практического использования хитина грибов для синтеза полимеров появилось довольно много исследований с данными по более тонкому изучению его в сопоставлении с хитином ракообразных (табл. 1.4). Ацетата и D-глюкозамина меньше всего у Aspergillus niger, что объясняется более высоким содержанием пентоз и глюкозы в составе β-глюкана и двух α-глюканов оболочки гриба. Анализ дифрактограмм показал идентичную кристаллическую структуру хитина грибов и крабов с несколько большей упорядоченностью у последних. Кроме того, хитин грибов в противоположность пластинчатой структуре хитина крабов имел волокнистое строение (Феофилова и др., 1980).
Заменяющий хитин у некоторых мукоровых грибов хитозан, обнаруживаемый при слабом кислотном гидролизе реакцией на хитозансульфат, найден, кроме Mucor rouxii, также у Phycomyces blakesleeanus. Количество ацетильных групп в нем оказалось различным и варьировало у М. rouxii до нуля. Из других аминополисахаридов полимер галактозамина со свободной аминогруппой, способной связывать фосфаты, со структурой типа хитозана, был выделен из клеточных оболочек Neurospora crassa, N. sitophitla, А. niger и Botrytls drierea (Aronson, 1965). У ряда грибов встречены полимеры из аминосахаров, ассоциированных с маннаном, глюканом и протеинами.
Целлюлоза в клеточных оболочках грибов обычно не встречается одновременно с хитином. Исключение составляет один Rhizidiomyces из порядка гифохитридиевых грибов, у которого они были обнаружены одновременно. Целлюлоза была обнаружена у грибов из оомицетов порядков Acrasiales, Lagenidiales, Saprolegniales, Leptomitales, Peronosporales (Aronson, 1965).
Среди грибов, обитающих в водной среде, целлюлоза встречается обычно только в группах, имеющих двужгутиковые зооспоры. У Blastocladiales и Monoblepharidales, имеющих одножгутиковые зооспоры, ее нет. Исключение, которое составляет Rhizidiomyces из порядка Hyphochytridiales, имеющий одновременно и хитин, и целлюлозу и считающийся по Нейбл переходной формой между имеющими хитин Chytridiales и Blastocladiales и содержащими целлюлозу оомицетами, объяснимо. Эта одножгутиковая форма имеет жгутик зооспор перистый, а не бичеподобный, как у Blastocladiales и Monoblepharidales. Любопытно, что строение ворсинок перистых жгутиков оомицетов напоминает строение жгутиков бактерий, тогда как бичеподобные полностью схожи со жгутиками флагеллат.
Целлюлоза у грибов легко обнаруживается микрохимически реактивом Шпейцера или реактивом, состоящим из раствора йода в йодистом калии с добавлением 70%-го раствора серной кислоты.
Глюканы у грибов очень обильны и отличаются от целлюлозы по их структуре. Мономеры их также представляют собой глюкозу. Однако наиболее изученные глюканы дрожжей имеют менее кристаллические структуры, чем целлюлоза. Кристалличность глюка-нов увеличивается вследствие образования гидроглюканов при их обработке серной кислотой. Подобные глюканы, находящиеся в ассоциации с протеинами, были обнаружены у дрожжей и у Penicilliunt notatum (Aronson, 1965). Другой глюкан, входящий в состав оболочек грибов, - каллоза, сходная с обнаруживаемой в ситовидных трубках высших растений и сильно красящаяся основными красителями, т. е. имеющая кислую природу, в противоположность целлюлозе имеет β-1,3-глюкозидные связи. Подобный глюкан, красящийся основными красителями, был обнаружен у Sclerofinia. Глюканы имеются также у Aspergillus fischeri, Allomyces macrogynus, Neurospora crassa. У грибов встречаются также гетерополисахариды, составленные из мономеров различных сахаров, особенно частые у рода Penicillium. У дрожжеподобных форм, патогенных для животных, как у Coccidioides и Cryptococcus, подобные же, но кислые полисахариды входят в состав окружающих их клетки капсул. Примеры глюканов и других продуктов полимеризации моносахаров и сахарных кислот, входящих в состав оболочек клеток и запасных веществ грибов, приведены в табл. 1.5.

Маннаны, полисахариды, составленные из мономеров маннозы. встречаются особенно обильно у дрожжей, причем они часты у видов дрожжей, обитающих на поверхности камбиального стоя под корой деревьев. К числу таких форм относится обитающая под корой хвойных деревьев Hansenula; в окружающей клетки этих дрожжей капсуле полисахарид присутствует в форме фосфоманнана. Этот гидрофильный и слизистый полисахарид прилипает вместе с дрожжами к щетинкам, покрывающим тело жуков короедов, и таким путем дрожжи переносятся с их помощью с одного дерева на другое (Wickerham, Barton, 1961). У гифообразующих грибов маннаны пока обнаружены не были, но в гидролизатaх их клеточных стенок встречается манноза.
У грибов встречаются также полисахариды, содержащие галактозу, 6-дезоксипентозу, метилпентозы, наиболее часто фруктозу, особенно у мукоровых. В оболочках клеток Penicillium chrysogenum были найдены из числа пентоз 6-дезоксипентоза, рамноза и ксилоза, которую обнаружили также у трутовика Polysiictus sanguineus.
Полисахаридно-протеиновые комплексы были обнаружены у дрожжей, например у Candida albicans - возбудителя молочницы грудных детей. Маннаново-протеиновый комплекс был найден у Saccharomyces.
Липиды у грибов количественно сильно варьируют в зависимости от условий окружающей среды и возраста культуры. Иногда их количество достигает 35-36% от массы сухого вещества клеток. У дрожжей более 3% липидов найдено в их оболочках. Обнаружены они и в оболочках мукоровых грибов, например у Мuсоr rouxii и Phycomyces, в спорангиеносцах которого установлено около 25% липидов от их сухой массы. Вероятно, они содержатся там в образованиях типа кутикулы (Aronson, 1965).
Пигменты также входят в заметных количествах в состав клеточных оболочек грибов. Пигменты, в особенности часто локализованный в виде особого слоя черный пигмент, меланин, очень часто встречаются в клеточных стенках мицелия или в оболочках спор многих грибов. Такой меланиновый слой имеется в оболочках аскоспор Neurospora tetrasperma (Aronson, 1965).
Меланин отсутствует у грибов с низкой активностью полифенолоксидазы, принимающей участие в его биосинтезе, и с преобладанием в обмене активных дегидрогеназ. Такие грибы, к которым относятся представители родов Fusarium, Trichothecium, Arthrobotris, Cephalosporium и многие другие, характеризуются чаще всего розовой или оранжевой окраской спорулирующих структур, зависящей от обилия каротиноидов, принимающих на себя роль светозащитного экрана и антиоксиданта, принадлежащую у темноокрашенных грибов меланину. Связь наличия каротиноидов с высоким уровнем активности дегидрогеназ объясняется тем, что оптимальные условия окислительно-восстановительного режима для действия дегидрогеназ совпадают с условиями, оптимальными для биосинтеза каротиноидов и других продуктов шунта терпеноидов. Каротиноидные пигменты уже не входят обычно в состав клеточных оболочек грибов, а локализуются либо в клеточной мембране, либо в каплях липидов, рассредоточенных в цитоплазме. В отдельных случаях у грибов встречаются совершенно особые пигменты, как оммохромы, глазные пигменты насекомых, у трутовика Pycnoporus (Polyporus) cinnabarinus (Шиврина, 1965) или фикобилины у видов сыроежек Russula emetica и R. paludosa (Ефименко, 1972), входящие в систему фотосинтеза у сине-зеленых и красных водорослей.
Клеточная стенка. Представляет собой многослойную оболочку из 9...10 слоев различной электронной плотности. Система микрофибрилл, встроенных в аморфный матрикс, формирует скелет клетки. Фибриллы в зависимости от видовой принадлежности могут состоять из целлюлозы, глюкона и хитина. Другие полисахариды, белки, пигменты, липиды служат цементирующими веществами, образующими химические связи с микрофибриллярной частью клеточной стенки. Наличие таких комплексов обеспечивает избирательную проницаемость для одних веществ и блокаду других.
Опорные микрофибриллы клеточной стенки и ее матрикс отличаются по механизму образования и биосинтезу. Образование фибрилл и матрикса происходит несинхронно, в первую очередь регенерируется фибриллярный остов стенки. Биосинтез этих двух частей клеточной стенки осуществляется с участием ферментов.
Процесс образования клеточной стенки происходит двумя способами: новый материал может либо внедряться в стенку поляри- зованно, либо равномерно накладываться по всей ее поверхности. В первом случае происходит образование цилиндрических клеток, во втором - сферических.
Клеточная стенка служит защитным приспособлением и предохраняет грибную клетку от воздействия различных факторов окружающей среды, например осмотическим барьером, обусловливающим избирательную проницаемость для различных веществ. Она придает форму вегетативным клеткам гиф и органов размножения. На поверхности клеточной стенки и цитоплазматической мембраны локализованы ферменты, осуществляющие превращение не усвояемых клеткой (не растворимых в воде) полимеров.
В результате лизиса клеточная стенка грибов может разрушиться под воздействием ферментов, выделяемых другими клетками и образующихся в клетке самого гриба.
Основные компоненты клеточной стенки грибов - хитин, глюканы, белок и жиры. Азотистые и безазотистые полисахариды с жировыми веществами образуют растворимые и нерастворимые комплексы. Основу клеточной стенки составляют 4...6 моносахаров, соотношение которых у различных грибов незначительно варьирует. В состав полисахаридных фракций входят глюкозамин, манноза, глюкоза, ксилоза и др. Следует подчеркнуть, что состав клеточной оболочки различных клеток одного и того же гриба неодинаковый.
Протопласт - содержимое клетки, заключенное в клеточную стенку: Имеет цитоплазматическую мембрану, эндоплазмати- ческий ретикулум, одно или несколько ядер с ядрышками, а также митохондрии, рибосомы с РНК, лизосомы, аппарат Гольджи, вакуоли, пластинчатый комплекс, секреторные гранулы, а также другие структуры и различные включения.
Цитоплазматическая мембрана. Тонкая трехслойная оболочка, располагается непосредственно под клеточной стенкой и отделяет ее от цитоплазмы. Цитоплазматическая мембрана обладает избирательной проницаемостью для веществ, входящих в клетку и выходящих из нее. Цитоплазматическая мембрана содержит до 40 % липидов и до 38 % белков. Различной формы инвагинации и ущемления цитоплазматической мембраны называются мезосомами.
Основное функциональное назначение цитоплазматической мембраны заключается в следующем: осуществление поступления в клетку различных веществ, ферментативная переработка и выделение продуктов метаболизма. Переработанные в цитоплазматической мембране вещества поступают в протопласт клетки и участвуют в обмене веществ.
Эндоплазматический ретикулум. Состоит из пузырьков, канальцев и вакуолей, служащих своеобразным депо питательных веществ.
Митохондрии. Многочисленные подвижные замкнутые образования эллипсовидной формы, с перегородками, покрытые одно- или двухслойной оболочкой. Предполагают, что митохондрии, благодаря собственной ДНК кольцевой структуры, способны к репродукции. Митохондрии окружены мембраной, на которой происходит локализация ферментов: пируватоксидазы, сукциндегидрогеназы, щелочной и кислой фосфатаз, пероксидазы и др. Митохондрии служат генераторами энергии в клетке. В зависимости от условий культивирования и физиологического состояния клетки форма митохондрий и их количество в клетке варьируют.
Рибосомы. Округлые зерна рибонуклеопротеидной природы размером до 200\ принимают участие в синтезе клеточных белков. Количество рибосом значительно отличается у различных видов грибов и зависит от внешних факторов, возраста культуры и др.
Аппарат Гольджи. Представлен группой пузырьков очень мелкого диаметра (0,000 002...О,ООО 01 мкм) или параллельно лежащими дисковидными пластинками. Этот органоид располагается в клетке на участке, свободном от рибосом.
Лизосомы. Производные аппарата Гольджи, размешаются между клеточной оболочкой и цитоплазматической мембраной. Представляют собой зернистые образования, окруженные однослойной липопротеидной мембраной. Содержат фермент, гидролизирующий белок, и выполняют функцию зашиты клеток от неблагоприятного воздействия токсичных веществ экзо- и эндогенного происхождения.
Липосомы. Капельки жировых веществ, окруженные однослойными мембранами.
Ядро. Находится в центре или на полюсах клетки. В грибных клетках могут быть одиночные и множественные ядра. Они отвечают за наследственные функции. Форма ядер округлая или удлиненная. Каждое ядро окружено двухслойной пористой нуклеомембраной с ядрышком из плотных зерен и тонких фибрилл. Ядрышки содержат в составе хромосом ДНК. Через анастомозы ядра могут мигрировать из одной клетки в другую.
Включения. В грибных клетках многочисленные включения: волютин, гликоген, липиды, пигменты, миелоидные образования, соли органических кислот, аминокислоты и др. Считается, что гликоген ответствен за эндогенное дыхание, а волютин служит запасным питательным веществом, участвующим в энергетических процессах.
Следует отметить, что в процессе жизнедеятельности в клетках грибов накапливаются различные продукты метаболизма - антибиотики, ферменты, токсины, витамины и др.
Все многочисленные морфологические элементы микроскопических грибов подразделяют на две группы: мицелий и споры. Они бывают различной формы и размеров. Морфологическое различие спор и мицелия служит важным дифференциальным признаком при определении вида гриба.
Мицелий. Представляет собой узкую круглую трубку, диаметр которой варьирует у микромицетов от одного до нескольких микрон.
При обильном ветвлении гифы мицелия, соприкасаясь друг с другом, могут образовывать слияния между клетками, - анастомозы. При наличии большого их количества мицелий приобретает характерный сетчатый вид. Развитие анастомозов наблюдается у различных грибов с многоклеточным мицелием. Благодаря им возможно перемещение клеточного ядра из одной клетки в другую и переход от гаплоидного к диплоидному мицелию. Однако в большинстве случаев они осуществляют вегетативные функции и развиваются у многих форм при недостатке питания. Длина клеток мицелия колеблется от нескольких микрон до десятков и реже сотен микрон.
Мицелий окружен двухконтурной оболочкой, которая у молодых культур более нежная. В перегородках, делящих мицелий на отдельные клетки, имеются поры, через которые в процессе роста переливается цитоплазма, а с ней и питательные вещества. В клетках много различных включений: в старых цитоплазма становится зернистой из-за множества вакуолей. Молодой мицелий состоит из удлиненных прямоугольных клеток, старый - из коротких округлых или многогранных. Мицелий, имеющий перегородки, называется септированным. Однако у некоторых низших грибов мицелий состоит из гиф, лишенных поперечных перегородок, и представляет собой как бы одну, сильно разветвленную гигантскую клетку с многочисленными ядрами и называется несептированным мицелием.
Как же происходит развитие мицелия? Из споры выпячивается ростковая трубочка, которая удлиняется и затем отчленяется перегородкой от средней части, включающей спору. Ростовые трубочки затем еще удлиняются и получают новую перегородку, разделяясь на дистальную, или верхушечную, клетку и проксимальную, или внутреннюю. В дальнейшем верхушечная клетка удлиняется и вновь делится, отделяя вторую, более молодую по сравнению с первой, внутреннюю клетку. Так повторяется и дальше. В этом процессе внутренние клетки только вытягиваются, поперечное деление их происходит редко, но зато из них развиваются боковые ветви. На дистальном конце внутренней клетки образуется боковое выпячивание, принимающее цилиндрическую форму и отделяющееся затем перегородкой от производящей ее клетки. Новая клетка вырастает затем в боковую ветвь, растушую и ветвящуюся таким же образом, как и главная. Благодаря развитию ветвей на протяжении главной гифы они тем старше и сильнее развиты, чем ближе к основанию лежит то их отхождения - акропетальное ветвление.
Развитие несептированного мицелия происходит в общем таким же образом, но без образования поперечных перегородок. Рост происходит на кончиках гиф, где накапливается обильная протоплазма, заполняющая весь просвет, а в более задних частях происходит значительное развитие центральных вакуолей. В однородной среде, например на поверхности питательной желатины, гифы мицелия (как неклеточного, так и многоклеточного) разрастаются равномерно и радиально, так что мицелий имеет форму круга, нарастающего с краев. Центральная часть в нем самая старая, даже иногда отмершая, а периферическая - наиболее молодая.
При общем однообразии развития мицелия, который можно назвать типичным, в отдельных случаях наблюдается ряд специфических черт как макроскопического вида и общего характера роста, так и микроскопического строения. Макроскопический вид мицелия определяют прежде всего воздушные гифы. В одних случаях они формируются на самой поверхности субстрата и отчасти внутри его и тогда мицелий имеет вид плоского, прижатого к субстрату кружка; в других случаях, кроме того, развиваются более или менее обильные гифы, поднимающиеся в воздух и придающие мицелию некоторое сходство, например с куском ваты, возвышающимся над субстратом. Характер роста может быть различным у одного и того же гриба в зависимости от влажности, питания и др. Однако ряд форм грибов имеет специфические особенности, например образование пышного воздушного мицелия - разрушителя древесины.
Цвет мицелия чаше всего бывает снежно-белый, но с возрастом приобретает бурую окраску разных оттенков. Это связано с отложением пигмента в клеточных стенках и реже внутри самой клетки.
Различают мицелий истинный и псевдомицелий. Последний характеризуется тем, что отдельные клетки не связаны друг с другом и не имеют общей оболочки. Вместо истинного ветвления здесь наблюдается древовидное расположение клеток.
Для прикрепления к субстрату и извлечения из него питательных веществ в ходе эволюции у некоторых грибов сформировались специально предназначенные для этого органы: ризоиды и аппрессории, которые учитывают при идентификации грибов. Ризоиды - это корешкообразные, а аппрессории - короткие расширенные, иногда лопастеобразные выросты мицелия.
Склероции , тяжи, ризоморфы и хламидоспоры также являются видоизменениями мицелиального роста.
Склероции представляют собой септированные гифы грибов, образующие особые тела. При формировании склероциев оболочки гиф утолщаются и приобретают темную окраску. Сильно утолщена стенка гиф наружного слоя склероция, внутри же гифы более тонкостенные и обычно не окрашены. Склероции - это защитные приспособительные тела, которые позволяют грибу длительное время сохраняться в окружающей среде и обеспечивают его устойчивость к воздействию различных внешних факторов: температуры, солнечных лучей и др. Зрелые склероции содержат меньше влаги по сравнению с мицелием и много запасных веществ - липидов, гликогена.
Размеры склероциев колеблются от нескольких миллиметров до нескольких десятков сантиметров, а форма бывает самая разнообразная: сферическая, неправильная, в виде прямых или изогнутых рожков и др.
Структура клеток склероциев и механизм их образования различны, однако их формирование происходит путем увеличения ветвления мицелия и септирования гиф. Известны два способа образования склероциев: терминальный - на концах гиф; интеркалярный - в отдельных фрагментах главных гиф.
У многих грибов при развитии плодовых тел и некоторых вегетативных структур образуется ложная ткань - плектенхима (псевдопаренхима). В отличие от настоящей ткани паренхимы, возникающей в результате деления клеток в трех направлениях, плектенхима образуется путем сплетения и срастания. Если она состоит из клеток более или менее изодиаметрических, то ее называют параплектенхимой; если в ней заметно явное гифообразное строение (клетки удлиненной формы), то ее называют прозоплектенхимой.
Мицелиальные тяжи - вегетативная структура линейно агрегированных гиф. Диаметр мицелиальных тяжей зависит от количества гиф, которые концентрируются вокруг центральной основы.
В простейшем случае небольшое количество параллельно идущих гиф склеиваются друг с другом ослизненными наружными оболочками или вступают в более прочное соединение путем фор. мирования многочисленных коротких анастомозов. В других слу. чаях, когда тяжи массивны, их гифы получают определенную дифференцировку. Наружные элементы бывают более тонкими, образуя как бы кору вокруг центрального толстого ствола.
Ризоморфы - более сложные по агрегации гифы, которые отличаются у различных грибов интенсивностью роста центральной гифы, протяженностью боковых ветвлений, а также степенью дифференциации клеток гиф.
Наружные части у ризоморфы обычно темноокрашены и имеют определенное сходство с корнями высших растений. Они широко распространены у грибов с крупными плодовыми телами: у базидиальных, сумчатых и др.
Основное назначение мицелиальных тяжей и ризоморф состоит в обеспечении распространения грибов в субстрате и передвижении по гифам питательных веществ.
Хламидоспоры - это изменения мицелия в зрелых и старых культурах на концах или по его ходу. Основная функция хламидоспор не размножение, а сохранение вида. Форма их обычно круглая, овальная или слегка удлиненная, диаметр превышает диаметр мицелия. У некоторых грибов стенка двухконтурная, поверхность гладкая или шероховатая. Хламидоспоры могут возникать на концах мицелия, тогда они называются терминальными, по ходу мицелия - интерполярными (промежуточными).
В старых культурах часто наблюдают большие скопления хламидоспор причудливой формы, напоминающей четки или ожерелье. Молодые и зрелые хламидоспоры способны прорастать. Старые клетки дегенерируют.
Споры . С помощью спор грибы не только размножаются, но также и распространяются в окружающей среде. Этому способствует высокая устойчивость оболочек спор к воздействию агрессивных факторов. Споры подразделяют на эндоспоры, образующиеся внутри особых вместилищ - спорангиев (сумок), и экзоспоры, располагающиеся на мицелии.
У совершенных грибов споры подразделяют на ооспоры, зигоспоры, аскоспоры, базидиоспоры, эндоспоры, фиалоспоры, хламидоспоры. Споры несовершенных грибов в соответствии с размерами и происхождением также делят на несколько групп. К эндоспорам, образующимся внутри мицелия путем сегментации последнего, относят таллоспоры, включающие в себя артроспоры, хламидоспоры и бластоспоры. Кроме того, для несовершенных грибов характерно образование конидий, макроконидий, алейрий (микроконидий) и гемиспор, считающихся несовершенными конидиями.
Гемиспоры более прочно связаны с мицелием и представляют, собой один или два сегмента, отшнуровывающихся после поперечного деления мицелиальной нити. Форма их цилиндрическая, иногда округлая или многогранная, оболочка двухконтурная.
МИКРОБИОЛОГИЯ, 2010, том 79, № 6, с. 723-733
УДК 582.281(047)
КЛЕТОЧНАЯ СТЕНКА ГРИБОВ: СОВРЕМЕННЫЕ ПРЕДСТАВЛЕНИЯ О СОСТАВЕ И БИОЛОГИЧЕСКОЙ ФУНКЦИИ
© 2010 г. Е. П. Феофилова1
Учреждение Российской академии наук Институт микробиологии им. С.Н. Виноградского РАН, Москва
Поступила в редакцию 05.11.2009 г.
Обзор посвящен мало изученной поверхностной структуре клетки мицелиальных грибов - клеточной стенке (КС). Приводятся данные о методах выделения и проверке чистоты на отсутствие цитоплазмати-ческого содержимого во фракции КС и о ее химическом составе. Подробно рассматриваются структурные (каркасные) и внутриструктурные компоненты КС - аминополисахариды, а- и в-глюканы, белки, липиды, уроновые кислоты, гидрофобины, спорополленин и меланины. Особое внимание уделяется хитину, его новой функции в антистрессовой защите клеток, а также отличиям этого аминополисахарида грибов от хитина водорослей и Arthropoda. Обсуждается феномен апикального роста гифы и участие специальных микровезикул в морфогенезе грибной клетки. Приводятся данные о ферментах, участвующих в синтезе и лизисе КС. В заключение обсуждается функциональная значимость КС у грибов по сравнению с поверхностными структурами высших эукариот.
Ключевые слова: мицелиальные грибы, клеточная стенка, методы выделения, химический состав, апикальный рост, физиологические функции, морфогенез.
Одной из ключевых проблем современной биологии является вопрос - каким образом в процессе развития происходит образование морфологически одинаковых организмов, какие биохимические механизмы и какие клеточные структуры участвуют в этом процессе, который уже миллиарды лет протекает на нашей планете. Не менее интересными представляются данные о том, как влияет стресс на морфогенез и какие биополимеры контролируют внешнюю форму клеток. Как показывают результаты исследований последних лет, растущая гифа ми-целиальных грибов является уникальной моделью, позволяющей понять процесс формообразования клеток и то, как сохраняется в онтогенезе постоянство клеточной морфологии. Очень большой вклад в исследование этого феномена внесли данные по изучению состава и биологической функции КС мицелиальных грибов.
КС растений была открыта в 1665 году, а КС грибов только в начале 18 столетия. Однако эта поверхностная клеточная структура длительное время практически не изучалась, так как считалось, что КС выполняет только "каркасную" функцию, и ее сравнивали со стенами дома, несущими опорную нагрузку. Но в начале XX века отношение к КС резко изменилось, ее начали интенсивно исследовать, но в основном у растений и бактерий. Однако интенсивное развитие грибоводства и биотехнологических производств, в которых продуцентами БАВ были мицелиальные грибы, интенсифицировали разви-
1 Адрес для корреспонденции (e-mail: [email protected]).
тие научных исследований по изучению КС грибов. Данные по этой поверхностной структуре за период до 80-х годов прошлого столетия были обобщены в первой в мире книге по КС грибов . В последующие годы основной массив работ пришелся на конец XX и начало XXI веков. Были проведены основополагающие исследования, посвященные систематике, химическому составу КС, апикальному росту гифы, метаболизму хитина, антигрибным препаратам, гидрофобинам, ковалентно связанным белкам, ферментам, участвующим в образовании КС, ветвлении гиф и лизисе КС . Эти данные не были обобщены, но именно обсуждение их в совокупности позволяет показать, что эта поверхностная структура клетки несет полифункциональную нагрузку и выполняет такие важнейшие функции, как, например, защита клетки от действия неблагоприятных факторов, контроль морфогенеза, участие в процессах репродукции, определение антигенных и адгезивных свойств, контроль процессов диморфизма и образования покоящихся клеток грибов, восприятие внешнего сигнала и передача его на мембрану и внутриклеточные мессенджеры. Учитывая сказанное, мы считали целесообразным остановиться на обсуждении следующих данных: о химическом составе КС, биологической функции ее основных биополимеров, строении КС и межклеточных взаимодействиях в гифах грибов, феномене апикального роста гифы, основных ферментах, участвующих в образовании и лизисе КС.
ХИМИЧЕСКИЙ СОСТАВ КЛЕТОЧНОЙ
СТЕНОКИ ГРИБОВ И БИОЛОГИЧЕСКАЯ ФУНКЦИЯ ЕЕ ОСНОВНЫХ КОМПОНЕНТОВ
Изучение химического состава КС грибов начинается с получения чистой фракции этой структуры, т.е. очистки КС от цитоплазматического загрязнения. Первоначальная процедура - разрушение клеток грибов и отмывка от клеточного содержимого - осуществляется при низкой температуре (около 4- 5°С) для того, чтобы предотвратить разрушительную деятельность деградирующих ферментов. Перед разрушением клеток их замораживают при температуре жидкого азота. Разрушение клеток осуществляется либо в специальных гомогенизаторах, либо на прессах методом "твердого давления". Хорошие результаты дает дальнейшее использование ультразвукового дезинтегратора. Отмывка клеточного содержимого осуществляется холодной водой и повторяется 4-5 раз. Для более тщательного удаления цитоплазматического содержимого используют отмывку с использованием №С1, 8 М мочевины, 1 М аммония или 0.5 М уксусной кислоты. В настоящее время разработаны методы выделения КС грибов в процессе их онтогенеза, причем эти методы существенно различаются для мицелия и покоящихся клеток . В зависимости от направления исследований, в частности, при определении полисахарид-ного состава, из КС извлекают липиды, используя органические растворители, например, хлороформ и метанол в соотношении 2: 1, иногда хорошие результаты дает дополнительная обработка КС серным эфиром. Этот растворитель хорошо подсушивает КС и иногда может заменить лиофильную сушку. Следующий этап - определение чистоты полученной фракции КС. С этой целью используют световую и электронную микроскопию и специализированные методы окраски. Наиболее часто применяется реакция, основанная на взаимодействии I3- с хитозаном. Изолированные чистые КС с раствором Люголя окрашиваются в розовый или фиолетовый цвет, в то время как интактные КС имеют красную окраску . Используется также окраска на наличие ядер с DAPI (4,6-ёкт1ёто-2-ркепуЦп-ёо1). Следует особо подчеркнуть, что выделение чистой фракции КС является очень ответственной процедурой, от которой зависят последующие результаты анализа ее химического состава. В этом отношении интересна работа , в которой у представителя мукоровых грибов обнаружен в мицелии глюкан, точнее, хитозан-глюкановый комплекс, хотя все предыдущие исследования свидетельствовали об отсутствии в мицелии Mucorales глюкана .
В настоящее время компоненты КС грибов разделяют на структурные компоненты (хитин, р-(1- 3)-р(1-6)-глюканы, р-(1-4)-глюкан (целлюлоза), и внутриструктурные (их называют матрикс), к которым стали относить маннопротеины, галакто-ман-нопротеины, ксило-маннопротеины, глюкуроно-
маннопротеины и а-(1-3)-глюкан . Наименее изучены а-глюканы, представляющие собой линейный полимер глюкозы (у Schizosaccharomyces pombe такой глюкан содержит приблизительно около 260 остатков глюкозы). Этот полимер состоит из двух связанных линейных цепей, содержащих около 120 (1-3)-а^-глюкозных остатков и (1-4)-a-D-глюкозных остатков на концах молекулы полимера. Предполагают, что этот глюкан необходим для протекания процесса морфогенеза . Из КС мицелия Penicillium roqueforti выделен нерастворимый в воде глюкан, который после высушивания теряет способность растворяться в щелочи, однако его удалось растворить в 10% растворе хлорида лития в диме-тилсульфоксиде. При изучении глюкана методом метилирования были получены два, три-О-метил-производных глюкозы, отвечающие связям 1 - 3 и 1 -»- 4 между моносахаридами в соотношении около 5: 2, и следы тетра-О-метил-производного, отвечающего концевым невосстанавливающим мо-носахаридным остаткам. Такой результат свидетельствует о линейной структуре молекул, содержащих только 1 -»- 3 и 1 -»- 4 связи между остатками глюкозы. Эти данные были подтверждены анализом спектра 13С-ЯМР полисахарида, из которого также следует а-конфигурация всех остатков глюкозы . Такие полисахариды, особенно после химической модификации (сульфатирования или карбосимети-лирования), способной придать им растворимость в воде, могут быть использованы как биологически активные полимеры и представляют интерес для изучения их биологической функции в клеточной стенке грибов и растений.
Нерастворимые в воде (1 -»- 3)-а^-глюканы были выделены ранее из нескольких видов высших и низших грибов, в том числе из Penicillium chrysoge-num , а родственные а^-глюканы с двумя типами связей (1 -3) и (1 -»- 4), различающиеся их соотношением, находили в высших растениях (Aconi-tum kusnezoffii Reichb ).
Структурный компонент - целлюлоза - характерен для оомицетных грибов, которых по современной систематике относят к псевдогрибам . Мо-носахаридный состав КС включает глюкозу, манно-зу, ксилозу, причем, преобладающим сахаром является глюкоза, составляющая до 68%. В состав КС грибов входят также аминокислоты, липиды (не более 3%) и ^ацетил^-глюкозамин. Глюканы, например, 1,3-глюкан, образуют прочный комплекс с хитином, называемый хитин-глюкановым комплексом (ХГК), который составляет скелет грибной клетки. Этот комплекс присутствует в КС практически у всех грибов, за исключением зигомицетов, что служит надежным систематическим признаком. У некоторых грибов на поверхности КС обнаружен слизистый материал, состоящий из полисахаридов, представленных ß-1,3-глюканами, содержащими только глюкозные единицы, соединенные ß-1,6-связями с каждой третьей единицей глюкозы по
Прокариотическая клетка
Прокариоты - организмы, не обладающие, в отличие от эукариот, оформленным клеточным ядром и другими внутренними мембранными органоидами (за исключением плоских цистерн у фотосинтезирующих видов, например, у цианобактерий). Единственная крупная кольцевая (у некоторых видов - линейная) двухцепочечная молекула ДНК, в которой содержится основная часть генетического материала клетки (так называемый нуклеоид) не образует комплекса с белками-гистонами (так называемого хроматина). К прокариотам относятся бактерии, в том числе цианобактерии (сине-зелёные водоросли), и археи. Потомками прокариотических клеток являются органеллы эукариотических клеток - митохондрии и пластиды. Основное содержимое клетки, заполняющее весь её объём, - вязкая зернистая цитоплазма.
Эукариоты - организмы, обладающие, в отличие от прокариот, оформленным клеточным ядром, отграниченным от цитоплазмы ядерной оболочкой. Генетический материал заключён в нескольких линейных двухцепочных молекулах ДНК (в зависимости от вида организмов их число на ядро может колебаться от двух до нескольких сотен), прикреплённых изнутри к мембране клеточного ядра и образующих у подавляющего большинства (кроме динофлагеллят) комплекс с белками-гистонами, называемый хроматином. В клетках эукариот имеется система внутренних мембран, образующих, помимо ядра, ряд других органоидов(эндоплазматическая сеть, аппарат Гольджи и др.). Кроме того, у подавляющего большинства имеются постоянные внутриклеточные симбионты-прокариоты - митохондрии, а у водорослей и растений - также и пластиды.
2) цитоплазма
Цитопла́зма - внутренняя среда живой или умершей клетки, кроме ядра и вакуоли, ограниченная плазматической мембраной. Включает в себя гиалоплазму - основное прозрачное вещество цитоплазмы, находящиеся в ней обязательные клеточные компоненты - органеллы, а также различные непостоянные структуры - включения.
В состав цитоплазмы входят все виды органических и неорганических веществ. В ней присутствуют также нерастворимые отходы обменных процессов и запасные питательные вещества. Основное вещество цитоплазмы - вода.
Цитоплазма постоянно движется, перетекает внутри живой клетки, перемещая вместе с собой различные вещества, включения и органоиды. Это движение называется циклозом. В ней протекают все процессы обмена веществ.
Цитоплазма способна к росту и воспроизведению и при частичном удалении может восстановиться. Однако нормально функционирует цитоплазма только в присутствии ядра. Без него долго существовать цитоплазма не может, так же как и ядро без цитоплазмы.
Важнейшая роль цитоплазмы заключается в объединении всех клеточных структур (компонентов) и обеспечении их химического взаимодействия. Так же цитоплазма поддерживает тургор(объём) клетки, поддержание температуры.
3)Клеточная стенка
Клеточная стенка - жёсткая оболочка клетки, расположенная снаружи от цитоплазматической мембраны и выполняющая структурные, защитные и транспортные функции. Обнаруживается у большинства бактерий, архей, грибов и растений. Животные и многие простейшиене имеют клеточной стенки.
Клеточные стенки прокариот
Клеточные стенки бактерий состоят из пептидогликана (муреина) и бывают двух типов: грамположительного и грамотрицательного. Клеточная стенка грамположительного типа состоит исключительно из толстого слоя пептидогликана, плотно прилегающего к клеточной мембране и пронизанного тейхоевыми илипотейхоевыми кислотами. При грамотрицательном типе слой пептидогликана существенно тоньше, между ним и плазматической мембраной находится периплазматическое пространство, а снаружи клетка окружена ещё одной мембраной, представленной т. н. липополисахаридом и являющаяся пирогенным эндотоксином грамотрицательных бактерий.
Клеточные стенки грибов
Клеточные стенки грибов состоят из хитина и глюканов.
Актиномицеты (Actinomycetales, от греч. aktis – луч, mykes – гриб) – это ветвящиеся бактерии, принадлежащие типу Актинобактерии (Actinobacteria). Они являются частью нормальной микрофлоры пищеварительной системы наземных позвоночных и беспозвоночных животных, а также в обилии присутствуют в грунте и играют важнейшую роль в экологии и круговороте веществ в почве.
Эти микроорганизмы являются возбудителями многих оппортунистических патологий – таких, которые возникают в результате снижения функции иммунной системы организма. Актиномицеты широко используются в биотехнологии, так как являются источником целого ряда антибактериальных и противоопухолевых веществ.
Рис. 1. Стрептомицеты синтезируют огромное количество антибактериальных и противоопухолевых препаратов.
Строение актиномицетов: почему все же бактерии, а не грибы?
1. Организация генетического материала
Наследственный материал Актиномицетов заключен в одной молекуле дезоксирибонуклеиновой кислоты, имеющей кольцевую форму и свободно располагающейся в цитоплазме – такая же форма организации генетического материала, называемая нуклеоидом, характерна и для других бактерий. У грибов же генетический материал организован и входит в состав клеточного ядра.
ДНК актиномицетов содержит большое количество ГЦ-пар (65-75% от общего количества нуклеотидов). Этот признак постоянен, не зависит от мутаций и потому используется в систематике микроорганизмов. Такое содержание ГЦ-пар делает ДНК актиномицетов весьма тугоплавкой, потому на анализ ДНК актиномицетов уходит больше времени по сравнению с другими бактериями.

Рис. 2. Схематическое строение клеточной стенки Гр+ бактерий.

Рис. 3. Актиномицеты, окрашенные по Граму.
Актиномицеты имеют плотную бактериальную клеточную стенку, которая расположена снаружи от цитоплазматической мембраны и обуславливает их положительное окрашивание по Грамму. Как и у других Грамм-положительных бактерий, она состоит из нескольких десятков слоев полимера муреина (пептидогликана), который пронизан тейхоевыми и липотейхоевыми кислотами. Липотейхоевые кислоты заякорены в цитоплазматической мембране бактерии и соединяют её с клеточной стенкой. Тейхоевые кислоты придают клеточной стенке отрицательный заряд. Клеточная стенка грибов же состоит из других полимеров – хитина и глюкана.
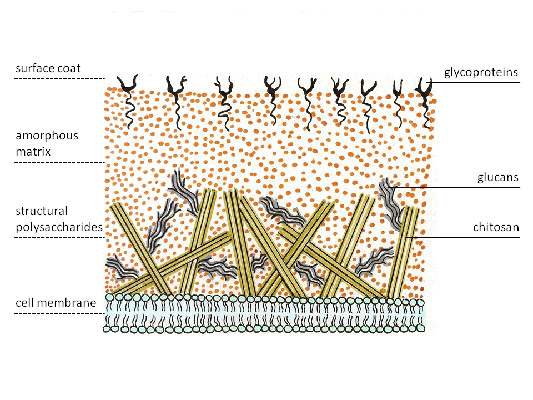
Рис. 4. Клеточная стенка грибов.
3. Клеточные органеллы
Актиномицеты, подобно другим бактериям, не имеют мембранных органелл. У актиномицетов 70S рибосомы, тогда как грибы имеют 80S рибосомы, равно как и остальные эукариотические организмы.
4. Рост колоний
Образование мицелия при росте – это то, что более всего роднит актиномицеты с грибами. Мицелий в случае актиномицетов представляет собой ветвящуюся совокупность гиф. Гифы разделены перегородками на длинные бактериальные клетки, содержащие несколько нуклеоидов. Перегородки у ряда видов могут проходить во взаимно перпендикулярном направлении. Ветвятся гифы путем почкования.
Мицелий, врастающий в субстрат (землю, ил или питательную среду), называется субстратным. Он обеспечивает колонию питательными веществами. Над субстратом возвышается воздушный мицелий, придающий колонии «пушистость» — он образует споры, а также так называемые «вторичные метаболиты» (в отличие от «первичных метаболитов» субстратного мицелия), среди которых много антибактериальных веществ.
Жизненный цикл и физиология актиномицетов
В процессе жизненного цикла большая часть актиномицетов образует споры. Некоторые актиномицеты размножаются путем фрагментации мицелия.
1. Спорообразование
Споры актиномицетов происходят из воздушного мицелия. Это экзоспоры – они развиваются снаружи материнской клетки. Гифы воздушного мицелия, из которых развиваются споры, называются спороносцами. Споры могут содержаться в утолщении на конце спороносца – спорангии (например, у стрептомицетов, актинопланов и плимелий), а могут располагаться цепочкой вдоль спорангиеносца (например, у нокардий и актиномадур).
По числу образующихся спор актиномицеты делятся на:
- Моноспоровые (к примеру, Saccaromonospora, Micromonospora, Thermomonospora) – образуют одиночные споры, чаще путем отпочковывания и последующего отделения перегородкой от материнского гифа;
- Олигоспоровые (например, Actinomadura) – образуют короткие цепочки спор вдоль спороносца;
- Полиспоровые (большинство других актиномицетов, к примеру, Streptomyces, Frankia, Geodermatophilus) – образуют множество спор, заключенных в спорангии.
Рис. 5. Спорангий актиномицетов рода Frankia.
Споры актиномицетов могут быть подвижными – в таком случае, спора имеет жгутик и может передвигаться (споры актинопланов, геодерматофилов и дерматофилов). В большинстве случаев споры неподвижны и распространяются ветром, водой или животными.

Рис. 6. Дерматофилы, световая микроскопия.
Споруляция у актиномицетов особенно активно протекает в неблагоприятных условиях. Устойчивость спор к нагреванию невелика по сравнению со спорами других бактерий, однако высушивание они выдерживают не хуже других, а потому имеют колоссальное приспособительное значение. Актиномицеты доминируют над другими микроорганизмами в сухих пустынных почвах.
Прорастание своры требует определенной влажности внешней среды. В присутствии воды спора набухает, в ней активируются ферменты и запускаются метаболические процессы, сопровождающиеся выходом ростовых трубок (будущих бактериальных тел) и синтезом нуклеиновых кислот.
2. Тип дыхания
Большинство актиномицетов — аэробы (нуждаются в кислороде для поддержания жизнедеятельности). Факультативные анаэробы (бактерии, способные жить как при наличии, так и в отсутствие кислорода) встречаются среди видов с непродолжительной мицелиальной стадией, размножающиеся фрагментацией мицелия.
3.Кислотоустойчивость
Актиномицеты обладают ацидотолерантностью – устойчивостью в кислой среде, которая позволяет им обитать в насыщенных кислотами лесных почвах. Кислотоустойчивость в лаборатории может быть определена окраской препарата, содержащего актиномицеты, по Цилю-Нильсену (фуксином с последующей протравкой серной кислотой и окраской метиленовой синью). Большинство актиномицетов при такой окраске не обесцвечиваются после протравки кислотой и сохраняют красный фуксиновый цвет. Щелочная среда неблагоприятна для этих бактерий: при повышенном рН они склонны к спорообразованию.
4. Особенности метаболизма
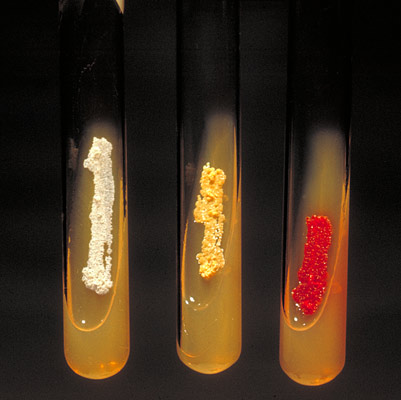
Рис. 7. Аэробные актиномицеты образуют пигмент на скошенном агаре. Слева направо: Actinomadura madurae, Nocardia asteroides, Micromonospora.
Выше упоминалось образование воздушным мицелием «вторичных метаболитов». Среди них:
- пигменты, обуславливающие различную расцветку воздушного мицелия при росте на средах;
- летучие пахучие вещества, придающие характерный запах почве после дождя, застоявшейся воде, кожным покровам некоторых животных;
- антибиотики:
a. противогрибковые – полиены;
b. противобактериальные – например, стрептомицин, эритромицин, тетрациклин, ванкомицин;
c. противоопухолевые – антрациклины, блеомицин.
Где обитают актиномицеты?
Актиномицеты в наибольшем количестве обнаруживаются в почвах, притом мицелиальных форм значительно меньше, чем спор. Они играют значительную роль в образовании гумуса, разлагая органические вещества, труднодоступные для утилизации другими бактериями. Актиномицеты в связи с этим используют в качестве санитарно-показательных микроорганизмов в санитарно-эпидемиологическом деле: обнаружение их в большим количестве в почве или воде указывает на наличие компоста в соответствующем субстрате.

Рис. 8. Актиномицеты в компосте.
Актиномицеты являются симбионтами многих растений, помогая им фиксировать азот. В то же время, многие микроорганизмы этого класса являются возбудителями заболеваний растений.

Рис. 9. Стрептомикоз картофеля.
Они также обнаруживаются в составе нормальной микрофлоры пищеварительной системы целого ряда животных, начиная от почвенных кольчатых червей (например, дождевых) и заканчивая крупным домашним скотом.
Эти микроорганизмы помогают расщеплять целлюлозу, в обилии присутствующую в растительной пище. У человека актиномицеты обнаруживаются в полости рта (десна и зубной налет), кишечнике (дистальные отделы толстого кишечника), на коже (лицо, крылья носа, за ушами, между пальцами) и в органах дыхательной системы (преимущественно в верхних дыхательных путях).

Рис. 10. Микрофлора кожи человека. Тип Актинобактерии обозначены оттенками синего, класс Актиномицеты – ярко-голубым.
Актиномицеты при условии снижения иммунной реактивности организма могут стать причиной актиномикозов – оппортунистических заболеваний, заключающихся в формировании актиномикозных гранулем – скоплений бактериальных тел, напоминающих зерна желтой серы («друз»), окруженных иммунокомпетентными клетками. Воспалительная реакция ведет к расплавлению гранулем, образованию свищей, ведущему к перфорациям органов и разносу бактерий кровью.
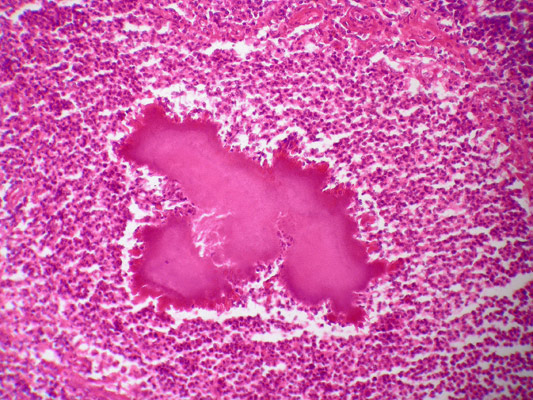
Рис. 11. Актиномикозная друза, окраска по Граму.
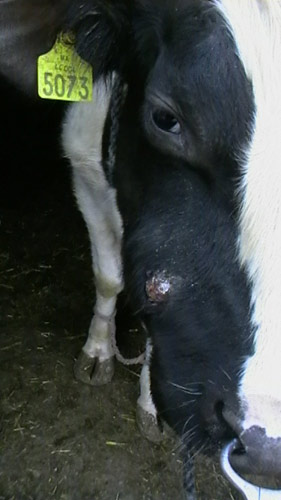
Рис. 12. Актиномикоз верхней челюсти у коровы.

Рис. 13. Максиллярный актиномикоз человека.
Актиномицеты – удивительные организмы, до сих пор вводящие в заблуждение множество ученых своей схожестью с грибами. Наряду с потенциальной опасностью в виде оппортунистических актиномикозов, эти организмы дарят человеку плодоносную почву и оружие для борьбы с инфекционными и онкологическими заболеваниями – антибиотики и цитостатики.