Индивидуално развитие(онтогенеза), периодизация на онтогенезата.
Всички организми имат специфичен жизнен цикъл. За организмите, развиващи се полово, то започва от момента на появата на зиготата и завършва с естествената смърт на организма.
Съвкупността от процеси, протичащи по време на жизнения цикъл на организмите, се определя като индивидуално развитие или онтогенеза.
Онтогенезата включва 3 периода:
1 период. Преембрионална или гаметогенеза.
2 период. Ембрионален.
3 период. Постембрионален.
1. Жизнени цикли на организмите. Развитието е ларвно и директно.
Развитието на организмите е пряко и непряко с трансформация.
непряко развитиепротича през стадия на ларвата. В ларвата се образуват определени зародишни или временни органи, които осигуряват жизнената дейност на организма на този етап от развитието.
При висшите гръбначни животните развитието е директно, но временните органи се образуват и по време на ембрионалното развитие. При бозайниците това са ембрионалните обвивки (амнион, хорион, алантоис, плацента) и жълтъчната торбичка.
2. Предембрионален период (предембрионален, прогенеза) на развитие. Етапи на гаметогенезата. Промени в оогенезата, свързани с ранното развитие на ембриона (генна амплификация, ооплазмена сегрегация, натрупване на хранителни вещества).
Предембрионалният период или гаметогенезата включва няколко етапа: изолация, размножаване, растеж, съзряване, образуване (последното само в сперматозоидите).
По време на оогенезата се случват най-важните събития, необходими за развитието на бъдещия организъм.
1 събитие.По време на оогенезата настъпва усилване на рРНК гени или увеличаване на броя на копията на гените, отговорни за рРНК. Този процес се случва в профазата на мейоза 1. Копията на rRNA гените могат да бъдат до един милион.
След това тези копия се отделят от хромозомите, плават свободно в кариоплазмата, около тях се образуват нуклеоли и в нуклеолите се синтезират рибозомни субединици, които навлизат в цитоплазмата. Така броят на рибозомите се увеличава рязко предварително в яйцето.
2 събитие.По време на оогенезата различни видове иРНК се синтезират по време на профазата на мейоза 1. Процесите на транскрипция протичат върху деспирализирани участъци от хромозоми. Хромозомите в профазния стадий на мейоза 1 се наричат хромозоми на ламповата четка.
3 събитие.Хранителните вещества се съхраняват в яйцето под формата на жълтък.
4 събитие.Яйцето се характеризира с ооплазмена сегрегация, т.е. разпределението на веществата в цитоплазмата на яйцето, което води до химическа хетерогенност на цитоплазмата. Смята се, че това е необходимо за ранна клетъчна диференциация.
5 събитие. Половите клетки са специални клетки на тялото, тъй като имат тотипотентност, тоест еднаква наследственост. Само зародишните клетки, както и бластомерите при хората, на етап 2 бластомера пораждат всички видове клетки. Например, експерименти за разделяне, снаждане или смесване на бластомери на етапа на разцепване показват, че при видове с радиален тип разцепване, бластомерите от няколко поколения, ако са изолирани и поставени в подходящи условия, проявяват тотипотентност, т.е. да се развие в цялостен организъм. За еднаква наследственост и тотипотентност на клетките на човешките ембриони до етапа на 2-4 бластомера говорят случаи на раждане на двама, три, четири еднояйчни близнаци.
3. Ембрионален период на развитие, периодизация.
Ембрионалният период на онтогенезата включва няколко етапа:
1 етап на оплождане.
2-ри етап на зиготата.
3 Етап на смачкване (образуване на еднослоен ембрион).
4 Етап на гаструлация (образуване на дву- и трислоен ембрион).
5 Етап хисто- и ОТНОСНОорганогенеза (образуване на тъкани и органи).
4. Оплождане и образуване на зигота. Характеристики на оплождането при бозайниците и човека.
етап на оплождане, това е процесът на сливане на яйцеклетката и спермата, в резултат на което се образува диплоидна зигота, от която се развива диплоиден организъм. Има 3 етапа в този процес:
1 етап- конвергенция на гамети. В това важна роля играят веществата, които се секретират от яйцеклетката и спермата. Те се наричат гамони (хормони на гамети, съответно гиногамони и андрогамони). В допълнение, има редица неспецифични фактори, които увеличават вероятността от среща и взаимодействие на спермата с яйцеклетката. Те включват
координиране на началото на готовността за оплождане при мъже и жени,
поведението на мъжките и женските, което осигурява копулация и осеменяване,
излишно производство на сперма
голяма яйцеклетка,
наличието на гамони, които допринасят за конвергенцията и взаимодействието на гаметите,
наличието на копулационни органи, които осигуряват вътрешно осеменяване.
При бозайниците голямо значениеима престой на сперматозоиди в гениталния тракт на женската, в резултат на което мъжките полови клетки придобиват оплождаща способност, т.е. способност за акрозомна реакция.
Етап 2 -активирането на гамети става след техния контакт. Активирането на сперматозоида се нарича акрозомна реакция. Активирането на яйцеклетката е кортикална реакция.
Същността на акрозомалната реакция : Пропускливостта на спермата в акрозомата се променямоста на плазмената мембрана и ензими, сперматолизини, се освобождават от акрозомата. Тези ензими отпускат връзките между фоликулните клетки, които заобикалят яйцето. Сперматозоидът преминава през слоя фоликуларни клетки, след което зоната на пелуцида се разрушава и сперматозоидът преминава през тази зона.
Същността на кортикалната реакция: Състои се в сложни структурни и физико-химични промени. Поради факта, че мембраната на сперматозоидите е пропусклива за натриеви йони, последните започват да навлизат в яйцето, променяйки се мембранен потенциалклетки. След това, под формата на вълна, разпространяваща се от точката на контакт на гаметите, настъпва повишаване на съдържанието на Ca 2+ йони (в хиалоплазмата те напускат депото - EPS, ретикулум) и в яйцето се задействат биохимични процеси, след което кортикалните гранули също се разтварят на вълна. Отделените при това специфични ензими допринасят за отлепването на жълтъчната мембрана; тя го втвърдява обвивка за оплождане.
Едно от значенията на кортикалната реакция е предотвратяването на полиспермията, т.е. проникване на повече от един сперматозоид в яйцеклетката. При бозайниците коровата реакция не предизвиква образуването на оплодителната мембрана, но нейната същност е същата.
Активирането на яйцеклетката завършва с началото на протеиновия синтез на транслационно ниво, тъй като тРНК, тРНК, рибозомите и енергията са били съхранявани дори в оогенезата.
3 етап- сливане на гамети или сингамия. В този случай се образува обща плазмена мембрана в спермата и яйцето. Женският и мъжкият пронуклеус се сближават и сливат (синкарион), образувайки обща метафазна плоча. Това е моментът на окончателното сливане на гаметите - сингамията.
Характеристики на оплождането при различни видове организми.
1 пример. При бозайниците и хората сперматозоидите се свързват с яйцеклетката в зоната, където има рецептор върху зоната пелуцида. След това взаимодействие останалите рецептори се блокират.
2 пример. При морски таралежслед оплождането в яйцето се променя драстично електрически потенциалплазмената мембрана и след това се образува мембрана за оплождане, която предотвратява полиспермията.
стадий на зигота.След пенетрацията мъжкото ядро се нарича мъжки пронуклеус. Хроматинът се разхлабва в него, възниква репликация на ДНК. Женското ядро се нарича женски пронуклеус. В него се случват същите събития. При бозайниците и хората не се случва ядрен синтез, но веднага се образува метафазна плоча.
5. Изкуствено осеменяване на яйцеклетка на животни и хора, биологични и медицински аспекти.
Изкуственото оплождане на животински яйца е от голямо научно значение за медицината, тъй като в процеса на неговото изследване се разработват начини и механизми за лечение на безплодие при хората.
Изкуственото осеменяване се използва за различни формибезплодие, както мъжко, така и женско, което е трудно за лечение. Например, когато мъжът има твърде малко сперматозоиди или те са практически неподвижни, когато жената има нарушена проходимост фалопиевите тръбиили има други увреждания на вътрешните полови органи, с имунологична несъвместимост на партньорите.
6. основни характеристикисмачкване. Видове смачкване, характерни за различни животински видове. Разцепване и образуване на бластула при плацентарни бозайници.
етап на смачкване.Това е етапът на образуване на еднослоен ембрион - бластула. Вътре в бластулата има кухина, наречена бластоцел.
Характеристики на смачкване:
Клетките се делят чрез митоза.
В навечерието на всяко делене се извършва репликация на ДНК.
Делящите се клетки не растат.
Видът на смачкване зависи от вида на яйцето.
Пълно равномерно смачкване в ланцета:
Първата бразда на разцепване върви вертикално, образуват се два бластомера. Втората бразда също върви вертикално и се образуват четири бластомера. Третата бразда върви хоризонтално, образуват се осем бластомера, след което се редуват вертикални и хоризонтални бразди. След 12 цикъла раздробяването става асинхронно. На определен етап от развитието си ембрионът представлява бучка от клетки или морула. Тогава между клетките се появяват празнини и се образува кухина - бластоцел. В ланцета по време на смачкване се образува бластула, която се нарича целобластула, тоест еднослойна топка.
Пълна неравномерна фрагментация при земноводните:
Клетките на земноводните са умерено телолецитални. На анималния полюс на клетката фрагментацията протича по-бързо, отколкото на вегетативния полюс. В резултат на това клетките на анималния полюс са по-малки – микромери. На вегетативния полюс клетките са по-големи – макромери. Бластулата на земноводните се нарича amphiblastula. Бластоцелът се намира на анималния полюс.
Характеристики на смачкване при бозайници и хора:
Разцепването е напълно неравномерно, асинхронно от първите етапи; на определен етап от развитието ембрионът е морула (бучка от клетки). След това по-големите клетки се отделят към периферията, образувайки трофобласт, а по-малките клетки към центъра, образувайки ембриобласт. Бластулата се нарича бластоциста. Бластоцелът е много малък. Трофобластът насърчава въвеждането на ембриона в маточната лигавица. Този процес се нарича имплантиране. Ембриобластът дава началото на самия ембрион и някои временни органи.
7. Обща характеристика на гаструлацията. Характеристики на гаструлацията при земноводни и птици. Гаструлация при висши (плацентарни) бозайници.
Етап на гаструлация, или етапът на образуване на двуслоен ембрион, а след това на трислоен. Ембрионът на този етап се нарича гаструла.
Методи за образуване на двуслоен ембрион:
- Инвагинация (инвагинация).
– Деламинация (разслояване).
– Имиграция (заселване).
- Epiboly (замърсяване).
Инвагинация или инвагинация. Този метод е характерен за ланцетника. В определена област клетките на бластулата се инвагинират в бластоцела, което води до образуването на двуслоен ембрион. Външният слой клетки се нарича ектодерма, вътрешният слой се нарича ендодерма. Ендодермата ограничава кухината на първичното черво или гастроцела. Входът на тази кухина се нарича първична уста или бластопор. Бластопорът е заобиколен от устни.
Деламинация или стратификация. Този метод е типичен за чревни животни, при които бластулата прилича на морула, а бластоцелът практически не се експресира.
имиграция или заселване. Някои бластулни клетки се въвеждат в бластоцела, след което тези клетки се делят интензивно. В резултат на това се образува ендодерма благодарение на тези клетки.
Епиболия (замърсяване). Микромерите са разделени и, така да се каже, наслоени върху макромери. Микромерите образуват ектодермата, докато макромерите образуват ендодермата. В чистата си форма тези методи практически не се срещат, като правило те се комбинират. Амфибиите съчетават инвагинация и епиболия. Птиците и бозайниците съчетават разслояване и имиграция.
Започвайки с плоските червеи, в еволюцията се появява третият зародишен лист - мезодермата.
Методи за образуване на мезодерма:
Телобластният метод е характерен за протостоми. В областта на устните на бластопора се отделят 2 клетки, които се делят и образуват мезодермата.
Методът ентерокоел е характерен за дейтеростомите (хордовите). 2 части от клетки под формата на джобове са симетрично отделени от ендодермата. Това са мезодермални джобове. Клетките на мезодермалните джобове се делят и пораждат мезодермата. Мезодермата е зародишният слой.
8. Обща характеристика на хисто- и органогенезата (образуване на тъкани и органи).
Етап на хисто и органогенеза(етап на образуване на тъкани и органи). Условно се разделя на два периода.
1 период.Период на образование аксиални органив ембриона това е образуването на невралната тръба и хордата. Следователно този период се нарича период на неврулация, а ембрионът на този етап се нарича неврула.
2 период. Характеризира се с образуването на други тъкани и органи. От дорзалната страна на ембриона (дорзално) по цялата му дължина от ектодермата се отделя участък от клетки, които дават началото на невралната пластинка. След това краищата на невралната пластина се издигат, удебеляват и се образува неврална бразда, която постепенно потъва под ектодермата. След това ръбовете на невралната бразда се затварят, образувайки неврална тръба с кухина вътре, кухината се нарича невроцел. При гръбначните животни предната част на невралната тръба се разширява и дава начало на мозъка, а останалата част на гръбначния мозък. В същото време под невралната тръба се полага нотохорда, образувана от ендодермата и прилежащата мезодерма. Първоначално мезодермата е хомогенна клетъчна маса, но с развитието настъпва нейната сегментация. Образуват се структури, които се наричат сомити. Впоследствие те дават началото на опорно-двигателния апарат.
Производни на зародишните слоеве:
Ектодерма - зъбен емайл, нервна система и сетивни органи, епидермис на кожата и нейните придатъци, епител на предното и задното черво.
Ендодерма - епител на средното черво, храносмилателни жлези и дихателна система.
Мезодерма - мускулно-скелетна система, пикочно-полова система, кръвоносна и лимфна система, цялата съединителна тъкан.
9. Характеристики на временни органи на ембриони на гръбначни животни. Временни органи на висшите бозайници.
Временните органи функционират в ембриона и липсват в зряла възраст. Те включват жълтъчната торбичка и така наречените ембрионални мембрани - амнион, хорион и алантоис.
Жълтъчна торбичка.Жълтъчната торбичка изпълнява редица важни функции: хранене, дишане, екскреция, хемопоеза. Но поради ниското съдържание на жълтък в яйцето, той не играе съществена роля в храненето на ембриона.
Амнион. Амниотичната мембрана участва в образуването на амниона, който ограничава амнионната кухина, пълна с амниотична течност, която сега обгражда ембриона от всички страни. Поради това ембрионът се развива във водна среда, което го предпазва от механични травматични ефекти и адхезия към черупките.
алантоисобразува се като израстък на задното черво. Основната функция на алантоиса е, че той е зародишен орган на отделяне. В него се натрупват разпадни продукти, които се образуват по време на метаболизма в тялото на ембриона.
Ембриогенезата е сложен холистичен процес, който е свързан с определени явления и механизми. Много от тези явления не са напълно разбрани, въпреки че за някои от тях са получени определени данни.
1. Молекулярно-генетични промени в ранното развитие.
2. Клетъчна пролиферация (клетъчно делене).
3. Клетъчна диференциация.
4. Образуване или морфогенеза.
1. Молекулярно-генетични промени в ранното развитие (период на зигота и разцепване), ролята на цитоплазмените фактори в яйцето.
Ранното развитие включва етапите на зигота и разцепване.
Изучавайки тези етапи, учените се опитаха да отговорят на въпросите:
Първо, когато собствените гени на ембриона започнат да работят.
Второ, има ли качествени и количествени разлики в иРНК и протеиновите молекули в различни частиембрион в ранните етапи на развитие.
В зиготата генната активност е ниска, тъй като ДНК е силно свързана с хистонови протеини. Първите протеини, които се синтезират в зиготата, са от майчин произход, тъй като рибозомите и иРНК молекулите са се натрупали в яйцето предварително. Установено е, че собствените гени на ембриона при бозайниците започват да работят на етап 2-4 бластомера. При земноводните е на етап бластула. Първи започват да работят гените, отговорни за пролиферацията и общия метаболизъм, по-късно започват да работят гените, отговорни за диференциацията на клетките и тъканите. Например, когато ядрото се отстрани от зиготата, настъпва разцепване и ембрионът в своето развитие достига почти стадия на бластула, след което по-нататъшното развитие спира.
Установено е, че няма качествени разлики в иРНК и протеиновите молекули в различните части на ембриона в ранните етапи на развитие. Има само количествени разлики.
Важна роля при раздробяването играе разделянето на цитоплазмата - цитотомия. Има особено морфогенетично значение, тъй като определя вида на смачкване. По границите между отделните участъци на ооплазмата минават бразди на разцепване, което отразява феномена на ооплазмената сегрегация. Следователно цитоплазмата на различните бластомери се различава по химичен състав.
2. Клетъчна пролиферация, растеж.
Клетъчната пролиферация или клетъчното делене се извършва по време на ембриогенезата. С това е свързан растежът на тъканите и органите. Растежът на ембриона като цяло.
3. Диференциация, молекулярно-генетични механизми на диференциация.
Клетъчната диференциация е набор от процеси, в резултат на които клетките от общ произход придобиват устойчиви морфологични, физиологични, биохимични различия, което води до клетъчна специализация. Специфичността на клетките се определя от протеините, които се синтезират в тях, а за протеините отговарят съответните гени. Следователно можем да заключим, че някои гени работят в едни клетки, а други в други. Това е същността на хипотезата за диференциалната активност на гените.
В ранните етапи клетъчната диференциация е свързана с влиянието на цитоплазмените вещества върху работата на съответните гени - това е епигенетичното ниво на регулиране на работата на гените. Феноменът на ооплазмената сегрегация се извършва в яйцеклетката, в резултат на което различни части от цитоплазмата на яйцето съдържат различни вещества. По време на раздробяването се появяват бластомери, наборът от гени в тях е един и същ, а съставът на цитоплазмата е различен. Впоследствие тези вещества на цитоплазмата, очевидно, водят до диференциална активност на гените.
При характеризиране на клетъчната диференциация се използват две понятия - детерминация и компетентност.
решителностозначава, че клетъчната диференциация е генетично предопределена и необратима.
В процеса на диференциация клетъчният материал на ембрионалните зародиши се трансформира в определен елемент от възрастния организъм. Нека разгледаме диференциацията, използвайки примера на мезодермалния сомит, който се подразделя на дерматом, склеротом и миотом. Дерматомни клетки на дермата, вторият - хрущялни клетки, третият - набраздени мускулни влакна. Следователно крайният резултат от развитието на отделните ембрионални зародиши е предварително определен или определен.
КомпетентностТова е способността на клетките да се диференцират в различни посоки, под въздействието на факторите на околната среда. Например, нотохордата и прилежащата мезодерма действат върху ектодермата, което води до образуването на невралната тръба от ектодермата. Ако няма такъв ефект, тогава ектодермата поражда епидермиса на кожата.
4. Морфогенеза (оформяне), нейните основни процеси:
Образуване или морфогенеза. Морфогенезата е съвкупност от процеси, в резултат на които ембрионът придобива характерен външен и вътрешна структура. От своя страна морфогенезата е свързана с:
а) морфогенетично движение на клетките
По време на ембриогенезата отделни клетки или групи от клетки се движат. Клетките се движат по повърхността на други клетки (поради механизма на амебоидното движение), където има специални молекули, които показват посоката на движение. Някои видове клетки се движат по градиент на химична концентрация (хемотаксис), но този механизъм е много по-рядко срещан.
Нарушаването на клетъчната миграция по време на ембриогенезата води до недоразвитие на органите или до промяна в нормалната им локализация. И двете са вродени малформации. Например, при нарушаване на миграцията на клетките - невробласти, в бялото вещество се появяват острови от сиво вещество и в същото време клетките губят способността си да се диференцират.
По този начин клетъчната миграция е под генетичен контрол, от една страна, и влиянието на околните клетки и тъкани, от друга.
б) ембрионална индукция
Това е въздействието на една тъкан (индуктор) върху друга тъкан, в резултат на което развитието на индуцираната тъкан става качествено ново. Първата и най-значима индукция е действието на нотохордата и мезодермата върху ектодермата, което води до образуването на невралната тръба. Без невралната тръба цялата ектодерма ще се трансформира в епидермиса. Това е първична ембрионална индукция, първата стъпка във верига от последователни (вторични, третични) процеси на индукция в по-нататъшното развитие.
Установено е, че има "специфични индуктори", т.е. вещества, които имат индуциращ ефект в незначителни концентрации и се различават по крайния резултат от действието си. Така екстракт от черен дроб на бозайници предизвиква предимно развитието на мозъчни структури, а екстракт от костен мозък - мезодермални.
Способността на ембрионалния зародиш да възприема индукционния стимул се нарича компетентност.
в) междуклетъчни взаимодействия
Това е взаимодействието на клетки или слоеве в контакт или на разстояние. Взаимодействието от разстояние протича с участието на биологично активни вещества (БАВ).
Това могат да бъдат протеини, хормони и др. В ранните етапи на ембрионалното развитие това са майчините хормони, тъй като ембрионът няма собствени жлези с вътрешна секреция. Хормоните не предизвикват нова диференциация, но я засилват.
Благодарение на междуклетъчните взаимодействия се извършват такива явления като морфогенетично движение на клетките, ембрионална индукция и клетъчна адхезия.
г) адхезия- способността на клетките да се слепват. В експеримента клетките на ектодермата, мезодермата и ендодермата бяха разделени и смесени една с друга. След това отново се събират в отделни групи, всяка от които представлява клетъчен агрегат от еднородни клетки. Отново се образуват три зародишни листа, разположени нормално един спрямо друг.
В процеса на адхезия участват специални протеинови молекули. Те се наричат молекули на клетъчната адхезия (MACs) и има около 100 вида.
Друга хипотеза гласи, че контактите между подобни клетки са по-силни, отколкото между чужди клетки.
Селективната адхезия на клетки от определен зародишен слой една с друга е необходимо условие за нормално развитие.
д) клетъчната смърт е необходим процес, тъй като образуването на отделни структури (канали, канали, дупки и др.) изисква разрушаване на част от клетките.
Има два фундаментално различни вида клетъчна смърт: апоптоза(в превод от гръцки "отпадане") и некроза.
Апоптозата е физиологична, генетично предопределена клетъчна смърт. Наред с други механизми на морфогенезата, той допринася за постигането на характеристиките на неговата морфофункционална организация, характерни за определен биологичен вид. Следователно апоптозата е естествен, еволюционно детерминиран и генетично контролиран механизъм на морфогенеза.
Некроза - нефизиологична клетъчна смърт, дължаща се на въздействието на неблагоприятни фактори (механични, химични, физични и др.). Некрозата обикновено е придружена от възпаление и е патологичен процес.
5. Интеграция в развитието, цялост на онтогенезата. Ролята на хормоните в координацията на процесите на развитие.
Понастоящем са известни редица вещества, които индуцират клетките да се делят, например фитохемаглутинин, някои хормони, както и комплекс от вещества, освободени по време на увреждане на тъканите. Открити са и тъканно-специфични инхибитори на клетъчното делене - кейлони. Тяхното действие е да потискат или забавят скоростта на делене на клетките в тъканите, които ги произвеждат. Например епидермалните калони действат само върху епидермиса. Тъй като са тъканно-специфични, калоните нямат видова специфичност. Така епидермалния калон от треска действа и върху епидермиса на бозайник.
Хормоните са органични съединения, произвеждани от определени клетки и предназначени да контролират функциите на тялото, тяхната регулация и координация.
Физиологичното действие на хормоните е насочено към:
1) осигуряване на хуморален, т.е. осъществява се чрез кръвта, регулирането на биологичните процеси;
2) поддържане на целостта и постоянството на вътрешната среда, хармонично взаимодействие между клетъчните компоненти на тялото;
3) регулиране на процесите на растеж, съзряване и размножаване. Хормоните регулират дейността на всички клетки на тялото. Те засягат остротата на мисленето и физическата подвижност, телосложението и растежа, определят развитието на признаци на полов диморфизъм и поведение.
6. Ролята на наследствеността и средата в ембрионалното развитие. Критични периоди на развитие. тератогенни фактори. Аномалии и малформации.
На всеки етап от онтогенезата организмът съществува в единство с заобикаляща среда. Ембриогенезата не е изключение в това отношение. Обхватът на условията, необходими за живота на даден вид, може да бъде широк. Въпреки това, за организми от всеки вид има минимум, оптимум и максимум необходими условияразвитие. Развитието на ембриона се влияе от колебания на естествени фактори (температура, влажност, атмосферно налягане, радиация, газов състав на околната среда).
Така че, в зависимост от температурата, процесите на развитие се забавят или засилват. Например жабешки яйца от същия съединител се развиват по-бързо при по-високи температури.
При аскаридите, когато достъпът на кислород до ембриона се прекрати, развитието спира.
Общо правилое, че под действието на светлината от синьо-виолетовата част на спектъра ембрионалното развитие на много животински видове се ускорява, а от червената се забавя.
По време на вътрематочното развитие факторите на околната среда са от голямо значение. Ако тези фактори водят до образуване на аномалии или дефекти в развитието, тогава те се наричат тератогенни. Тератогенните фактори могат да бъдат физически (висока температура, йонизиращо лъчение, рентген и др.), химически (лекарства, соли на тежки метали и др.) и биологични (вируси, бактерии). Тератогенните фактори водят до развитие на аномалии в определени периоди от ембрионалното развитие, които се наричат критични. Те включват:
Периодът на образуване на зародишни клетки (гаметогенеза),
етап на оплождане,
стадий на зигота,
Имплантиране на ембриона в стената на матката
плацентация,
Период на хистогенеза и органогенеза,
Малформации.
Аплазия - липса на орган или част от него
Хипоплазия - недоразвитие на даден орган
Хипотрофия - намаляване на теглото на тялото или органа
Хипертрофия - непропорционално увеличаване на масата на органа
Гигантизъм - увеличаване на дължината на тялото
Хетеротопията е нетипична локализация на група клетки или орган в тялото.
Хетероплазия - нарушение на тъканната диференциация
Стеноза - стесняване на канал или отвор
Атрезия - липса на канал или отвор
Персистенция – запазване на ембрионалните структури
В зависимост от причината вродените малформации се разделят на:
наследственапричинени от промяна в гените или хромозомите в гаметите на родителите, в резултат на което зиготата от самото начало носи генна, хромозомна или геномна мутация.
екзогененвъзникващи под въздействието на тератогенни фактори: лекарства (талидомид), хранителни добавки, вируси, промишлени отрови и др. Това са всички фактори на околната среда, които, действайки по време на ембриогенезата, нарушават развитието на тъканите и органите.
Многофакторнидефекти, които се развиват под въздействието както на екзогенни, така и на генетични фактори.
1. Постнатална онтогенеза, нейната периодизация.
Постембрионалното развитие (постнатално за човек) започва от момента на раждането и завършва с естествена смърт или смърт.
Постембрионалното развитие включва няколко периода:
1. Предрепродуктивен (ювенилен).
2. Репродуктивен (период на зрялост).
3. Пострепродуктивна (напреднала възраст).
2. Предрепродуктивен период, неговата характеристика. Растежът на тялото като важна характеристика на предрепродуктивния период.
Предрепродуктивният период започва веднага след раждането. По това време процесите на морфогенеза приключват, започват да функционират онези системи, които не са функционирали в ембриогенезата (дихателна, екскреторна и редица други).
Важна характеристика на предрепродуктивния период е растежът на организма. В този случай се получава увеличаване на размера на тялото като цяло, увеличават се надлъжните му размери; увеличаване на размера на тъканите и органите.
3. Характерът на растежа на организма и отделните му части.
Три основни процеса са в основата на растежа на организма:
1. увеличаване на броя на клетките.
2. увеличаване на размера на клетките (хипертрофия).
3. натрупване на междуклетъчно вещество.
Има два варианта за растеж: ограниченИ неограничен. Неограниченият растеж продължава през целия онтогенезис, до смъртта.
Има няколко вида растеж:
Помощни- растеж, който протича чрез увеличаване на размера на клетките.
пролиферативен- растеж, протичащ чрез размножаване на клетки: мултипликативенИ акреционен.
Мултипликативенрастежът се характеризира с факта, че и двете клетки, възникнали от деленето на родителската клетка, отново влизат в делене. Мултипликативният растеж е много ефективен и следователно почти никога не се среща в чиста форма или завършва много бързо (например в ембрионалния период).
акреционенрастеж се състои в това, че след всяко следващо делене само една от дъщерните клетки се дели отново, докато другата спира да се дели. В този случай броят на клетките нараства линейно. Такъв растеж е характерен за органи, в които се извършва обновяване на клетъчния състав.
Трябва да се отбележи, че увеличаването на надлъжните размери на тялото, което се дължи главно на растежа на дългите тръбести кости, е от особено значение за характеризиране на растежа. IN тръбести костина границата на диафизата и епифизата се изолира зона на растеж. Тук се намират хрущялни клетки, при чието делене костта нараства на дължина.
Окончателното вкостяване на всяка кост настъпва в определено време. При мъжете растежът обикновено завършва до 18-20 години, при жените - до 16-18 години. По това време последните зони на растеж изчезват. Тогава растежът на костите по дължина спира.
Трябва да се отбележи, че до 30 години човек може да нарасне с 3 см поради увеличаване на размера на прешлените.
Увеличаването на линейните размери на човек се описва с S-образна крива. Веднага след раждането се наблюдава засилен растеж на тялото, след което той намалява и рязко се ускорява към 13-14-15-годишна възраст. Това е така нареченият пубертетен скок на растеж (по време на пубертета). Освен това темпът на растеж се забавя донякъде и на възраст 30-40-45 години растежът на човек остава постоянен. По тази схема костите, мускулите и мн вътрешни органи(черен дроб, бъбреци, далак).
С напредване на възрастта се наблюдава леко намаляване на растежа.
Някои органи имат съвсем различен характер:
Тези органи включват мозъка и гръбначния мозък, лимфоидните органи, репродуктивните органи.
Теглото на мозъка на новороденото е 25% от окончателното тегло на мозъка (в зряла възраст), до 5 години - 90%, до 10 години - 95%.
Растежът на тимуса (тимус) е централният орган на имунната система. Относителното тегло на тимуса (спрямо телесното тегло) достига максимум до 12-годишна възраст. Абсолютното тегло достига максимум до 30-годишна възраст и след това има рязко намаляване на теглото на тимуса.
4. Генетичен контрол на растежа. Ролята на нервната и ендокринната система в регулацията на растежните процеси.
Височината се отнася до генетични черти, които се наследяват като цвят на косата и кожата, форма на очите и т.н. Ето защо високите родители обикновено имат високи деца и обратното. Растежът е полигенен признак; няколко гена са отговорни за проявата му във фенотипа. Гените упражняват своя контрол върху растежа чрез подходящи хормони. Най-важният хормон е растежният хормон или соматотропин, произвеждан от хипофизната жлеза.
Соматотропинът стимулира образуването на нови хрущялни клетки и отчасти тяхната осификация, насърчава протеиновия синтез в клетъчни структуриах и образуването на нови капиляри. През нощта се произвежда голямо количество от този хормон. Собственият соматотропин на детето се произвежда от 3-4 години.
Растежът също се влияе от хормоните на щитовидната жлеза и половите хормони.
5. Взаимодействие на биологичното и социалното през детството и младостта.
Ролята на наследствеността за растежа е голяма, но това не е единственият фактор. Наследствеността трябва да се разглежда като приблизителна програма, според която височината на човек може да бъде например в диапазона от 160 до 180 см. Какво всъщност ще бъде то зависи до голяма степен от външни условия, които могат да забавят наследствената програма или да допринесат за нейното изпълнение. Условия на околната среда, които влияят на човешкия растеж: хранене, физическа активност, психологически ефекти от тютюнопушене, алкохол.
Тоест населението расте (това е ускорение).
Една от вероятните причини е подобряването на условията на живот (храненето). Забелязва се, че в годините на война и природни бедствия растежът на децата намалява. Растежът се влияе леко от климата и географската среда.
6. Формиране на конституционни типове, типове тяло.
Формирането на конституционни типове хора е свързано с израстването на човек. Това трябва да се разбира като характеристиките на външните форми на тялото, характеристиките на функциите на тялото, характеристиките на поведението този човек. В зависимост от структурата на тялото, в зависимост от външните форми на тялото се разграничават определени видове физика. В момента има доста класификации. Една от тях е класификацията на М.В. Черноруцки. Според това към
Всеки организъм, независимо дали е едноклетъчен или многоклетъчен и към кое царство на живота принадлежи, преминава през индивидуално развитие, или онтогенезата(от гръцки. ontos- създание и генезис- раждане). Областта на изучаване на биологията онтогенезата, е наречен биология на развитието.
Онтогенеза - това е периодът на живот на организма от зиготата (първичната клетка) до смъртта.
При многоклетъчните организми онтогенезата обикновено започва с образуването на зигота и завършва със смъртта.
В същото време тялото не само расте, увеличавайки се по размер, но и преминава през редица различни жизнени фази, всяка от които има специална структура, функционира по различен начин, а в някои случаи и коренно различен начин на живот. .
При едноклетъчните организми началото на онтогенезата се счита за момента на отделяне от майчината или сестринската клетка. Продължава до следващото разделение или смърт. В същото време външната онтогенеза на едноклетъчните организми обикновено се проявява само като леко увеличение на размера на клетката, въпреки че всъщност това крие напълно различни периоди от нейния живот. .
Всеки вид има своя програма за онтогенеза. И това не е само набор и последователност от етапи на развитие, през които преминава, но и продължителността на всеки един от тях. В същото време всеки индивид има индивидуални характеристики на онтогенезата, които обаче не надхвърлят вида. , а те от своя страна се подчиняват на законите на онтогенезата на племенно, семейно, отрядно и дори класово ниво .
програма за онтогенезане е нищо повече от реализиране на наследствена информация, записана в гените. Следователно спецификата на онтогенезата на ниво отделни индивиди се определя от индивидуални комбинации от гени, а на ниво видове, родове, семейства - от специални гени, които са характерни само за всяка систематична група организми.
Механизмът за внедряване на наследствена информация е преди всичко в диференциал(от английски. различават- варират) генна активност. Това означава, че в различни периоди на развитие и в различни тъкани многоклетъчен организъмактивни са гени, върху чиито нуклеотидни последователности се синтезират определени тРНК. В резултат на това се синтезират специфични структурни протеини и ензими, които в крайна сметка определят характеристиките на функционирането и поведението на клетките на определени етапи от развитието на тялото, особеностите на техния метаболизъм.
При животните и цъфтящите растения онтогенезата се разделя на два периода. Ембрионално развитие, или ембриогенеза(от гръцки. ембрион- плода и генезис)продължава от образуването на зигота до раждането или излизането от яйцето, и постембрионален(от гръцки. бърз- след и ембрион) развитието продължава от раждането или излизането от яйцето и завършва със смъртта на организма.
Ходът на ембриогенезата.Общата схема на ембриогенезата включва следните фази: материал от сайта
- последователни деления на зиготата, завършващи с образуването на многоклетъчен ембрион, състоящ се от стотици и дори хиляди идентични клетки;
- диференциация (от англ. различават- разграничават) клетки, водещи до образуването на тъкани;
- полагане на органи и растеж на ембриона.
По време на онтогенезаорганизмът преминава през редица фази – състояния, в които се различава по устройство, функциониране и начин на живот. Има два етапа на онтогенезата: ембриогенеза - ембрионално развитие и постембрионално развитие- периодът на живот на организма от раждането (излизане от яйцето) до смъртта.
На тази страница материал по темите:
Обобщение на онтогенезата
Резюме на онтогенезата на индивидуалното развитие
Накратко понятието онтогенеза
-
Глава 7. ПЕРИОДИЗАЦИЯ НА ОНТОГЕНЕЗАТА
Глава 7. ПЕРИОДИЗАЦИЯ НА ОНТОГЕНЕЗАТА
7.1. етапи, периоди и етапи на онтогенезата
Индивидуалното развитие е холистичен непрекъснат процес, в който отделните събития са взаимосвързани в пространството и времето. Има няколко варианта на периодизация на онтогенезата, всеки от които е най-подходящ за решаване на конкретни научни или практически проблеми.
СЪС общобиологичнигледни точки голямо събитиеонтогенеза - полово размножаване. Ако съпоставим различните времеви периоди на онтогенезата със способността на индивида да изпълнява функцията на възпроизвеждане, тогава той може да бъде разделен на три периода: предрепродуктивен, активен репродуктивен и пострепродуктивен.
IN предрепродуктивен периодиндивидът е неспособен да се размножава. Основното му съдържание е в развитието на полово зрял фенотип. През този период настъпват най-изразените структурни и функционални трансформации, реализира се основната част от наследствената информация, тялото е силно чувствително към всякакви влияния.
В активно репродуктивен периодиндивидът изпълнява функцията на сексуално размножаване, отличава се с най-стабилното функциониране на органи и системи, както и с относителна устойчивост на влияния.
Следрепродуктивен периодсвързано със стареенето на организма и се характеризира с отслабване или пълно спиране на участието в репродукцията. Намален адаптивен капацитет и устойчивост на различни влияния. По отношение на онтогенезата на човека тези периоди се характеризират допълнително със специфични социални фактори (образование, работоспособност, творчество). Всеки период има свои собствени характеристики на заболеваемост.
Предрепродуктивният период е разделен на още четири: ембрионален, ларвен, метаморфозен и ювенилен.
ембрионален,или зародишен, периодОнтогенезата започва от момента на оплождането и продължава до освобождаването на ембриона от яйцеклетката. Този период се отличава с тежестта на процесите на трансформация на зиготата в организъм, способен на повече или по-малко независимо съществуване. При повечето гръбначни животни той включва етапите (фазите) на зигота, разцепване, гаструлация, както и хисто- и органогенеза. Продължителността му варира. При плацентарните бозайници той е особено съкратен. Черупката на едно яйце се разтваря преди имплантирането на бластоциста в лигавицата на матката. До този момент ембрионът има само време да премине през етапите на зигота и смачкване. Всички по-нататъшни процеси протичат под защитата и с участието на майчиния организъм. Еволюционното значение на тези характеристики е разгледано в раздел 13.2.
Разцепването се предшества от процесите на гаметогенеза и оплождане, които са пряко свързани с индивидуалното развитие и може дори да не доведат до него, но които до голяма степен определят по-нататъшното развитие на ембриона, ако има зачеване. Тези процеси се наричат прогенеза, която предхожда същинската онтогенеза. Цитологично, процесите на гаметогенеза и оплождане представляват междинна връзка, свързваща онтогенезата на родителите с онтогенезата на тяхното потомство.
Ларвен периодв типичен вариант се наблюдава в развитието на онези гръбначни животни, чиито ембриони излизат от яйчните мембрани и започват да водят самостоятелен начин на живот, без да достигнат дефинитивните (зрели) черти на организацията. И така, той се среща в някои представители на нисшите гръбначни - миноги, повечето костни риби и земноводни. Най-характерните особености на ларвата са: ембрионалният характер на нейната организация, наличието на временни (временни) органи, ранното начало на функционирането на редица органи, което дава възможност за самостоятелно съществуване. Благодарение на активното хранене ларвата получава възможност да завърши развитието си и благодарение на активното движение има възможност да избере условия на околната среда, които са оптимални за развитие, и по този начин да избегне конкуренцията със собствените си възрастни роднини. При гръбначните животни продължителността на ларвния период е много по-голяма от тази на ембрионалния период.
Метаморфозасе състои в превръщането на ларвата в млада форма. В процеса на метаморфоза се случват такива важни морфогенетични трансформации като частично унищожаване, преструктуриране и ново образуване на органи. Степента на трансформация е толкова по-голяма, колкото повече
разликите между местообитанието на ларвата и възрастния организъм, което е добре илюстрирано от примера за развитие на безопашати земноводни (сравнете: попова лъжица и жаба).
Юношескипериодът започва от момента на завършване на метаморфозата (при плацентарните бозайници и човека - от раждането) и завършва с пубертета и началото на размножаването. Характеристиките на юношеския период се проявяват в оригиналността на храненето на младия организъм, неговото поведение и степента на зависимост от родителите. От морфологична гледна точка този период се характеризира с интензивен растеж, установяване на окончателни пропорции между различни частитяло, завършване на развитието на скелета, кожата, смяна на зъбите, завършване на развитието на половите жлези и хормонална регулация. Продължителността на ювенилния период при гръбначните животни варира от минимум 13–18 дни при мишките полевки до максимум 18–20 години при белугата, крокодила, албатроса и слона. При много представители на гръбначните животни, особено при хората, постигането на пубертета и началото на възпроизводството може да бъде разделено от значителен период от време.
Приложение екологични и ембриологичниподходът ни позволява да разделим онтогенезата на етапи, които се случват преди раждането, по време наИ след ражданетолица. Самото раждане, т.е. освобождаването на развиващ се индивид от черупките на яйцето или от тялото на майката, в различни видовевъзниква на различни етапи на зрялост. В същото време при всички видове преди раждането тялото е защитено от мембраните на яйцата или тялото на майката и не е в състояние да се храни и да изпълнява други важни функции самостоятелно. Опазването на ранните, морфофункционално незрели стадии осигурява оцеляването на вида. След раждането индивидът установява връзки с новата среда, започва да се храни сам, да се движи и да изпълнява всички други функции.
По-подробно представяне на периодизацията на онтогенезата на човека, която има практическа стойноств акушерската и педиатричната практика, вижте точка 7.6.1 и таблица. 7.3.
7.2. Морфофизиологични и еволюционни особености на яйцата на хордовите
7.2.1. ПРОИЗХОД НА ЯЙЦАТА
Яйцата на хордовите се състоят от яйцеклетка, понякога наричана още яйцеклетка, и яйчни мембрани. Ооцитиобразувани в женските полови органи
жлеза - яйчник. Те преминават през дълъг път на развитие, който започва в ембрионалния и продължава в активния репродуктивен период от онтогенезата на женските индивиди.
Първични полови клеткимного рано в ембриогенезата се изолират от соматичните клетки. И така, при безопашатите земноводни гоноцитите се изолират на етапа на бластулата, при птиците те се определят по време на образуването на първичния хипобласт, при бозайниците в епибласта на етапа на гаструлата, при опашните земноводни този процес се случва по-късно, на етапа на гаструла или дори неврула (в мезодермата). След това тези клетки се движат, като в крайна сметка достигат до рудиментите на половите жлези. Гоноцитите на бозайниците първо се озовават в екстраембрионалната област в устието на жълтъчната торбичка и след това мигрират към зачатъка на половите жлези (фиг. 7.1). Механизмите за миграция са изложени в точка 6.5.3.
Веднъж попаднали в половите жлези, първичните зародишни клетки започват да се размножават. Те се делят чрез митоза и се наричат ово(оо)гония.При повечето нисши гръбначни животни ovo(oo)gonia запазват способността си да се делят през целия репродуктивен период, например рибите отделят хиляди яйца при едно хвърляне на хайвера, земноводните - стотици. При висшите гръбначни броят на яйцата, които узряват едновременно, рядко достига 15, обикновено са по-малко, понякога едно, което също може да обясни особеностите на ово(оо)генезата.
При човешките женски ембриони ово(оо)гонията се размножава най-интензивно между 2-ия и 5-ия месец на вътрематочното развитие.
Ориз. 7.1.Локализация на първични зародишни клетки в човешки ембрион на етап 16 сомита (а) и тяхната миграция към залата на половите жлези (б): 1 - предстомашие; 2 - задно черво; 3 - алантоис; 4 - първични зародишни клетки; 5 - жълтъчна торбичка; 6 - сърце; 7 - развиваща се гонада (генитален туберкул); 8 - първичен бъбрек; 9 - клоака
период на развитие, когато броят им достига приблизително 7 млн. До 7-ия месец много ово(оо)гонии умират, а останалите навлизат в профазата на първото разделение на мейозата и спират на етапа на диакинезата. С настъпването на пубертета един овоцит овулира всеки месец, достигайки етапа на метафаза на второто разделение на мейозата. При някои яйцеклетки това се случва по време на пубертета, а при други точно преди менопаузата. Ооцитът завършва мейозата само ако настъпи оплождане.
7.2.2. СПЕЦИФИЧНОСТ И ЗНАЧЕНИЕ НА ХИМИЧНИЯ СЪСТАВ НА ЦИТОПЛАЗМАТА НА ЯЙЦЕТО
Зрялото яйце обикновено е по-голямо от ово(оо)гония и други соматични клетки. По време на ово(оо)генезата в цитоплазмата на яйцето, голям бройвещества, необходими за неговото съзряване и ранна ембриогенеза. Така в периода на растеж обемът на мишия овоцит се увеличава около 40 пъти, а на жабешкия над 400 пъти.Скоростта на синтеза на веществата в овоцитите се увеличава значително, поради редица особености. В зреещите яйцеклетки настъпва амплификация (вижте раздел 2.4.3.4) на отделни гени (вижте раздел 4), например гени на рРНК. Земноводни (Xenopus laevis)около 450 rRNA гени се намират в диплоидна клетка и повече от милион в ово(оо)цит. Освен това се повишава транскрипционната активност на генома на тези клетки. По този начин скоростта на транскрипция в растящите миши овоцити е 10 пъти по-висока, отколкото в соматичните клетки.
Функционалната роля на съхраняваните вещества е различна.
първо,това са компонентите, необходими за процесите на репликация, транскрипция и транслация, като съответните ензими, рибозоми, иРНК, тРНК и техните прекурсори.
второ,това е набор от специфични регулаторни вещества, които осигуряват координираното функциониране на всички съхранявани компоненти. Тези вещества включват фактора на разпадане на ядрената мембрана (прометафазата на 1-вото разделение на мейозата започва с разрушаването на ядрената мембрана); фактор, който причинява кондензация на хромозоми; фактор, който превръща ядрото на спермата в пронуклеус и активира синтеза на ДНК в него преди разцепването; цитостатичен фактор csf,отговорен за блока на мейозата на етапа на метафаза II (при много гръбначни, отстраняването на този блок става само в резултат на оплождане); цитоскелетна микрофиламентна система, участваща в поддържането на блока
мейоза; циклични нуклеотиди (напр. сАМР), протеин кинази и други вещества, необходими за сигнализиране от околните ovo(oo)cyt клетки и извънклетъчната среда; компоненти на така наречената инозитол фосфатна система, които осигуряват активирането на яйцеклетката след оплождане; фактор, отговорен за цитотомията по време на разцепването. Някои от тях, намирайки се в овоцита още по време на оплождането, започват да действат само във фазата на гаструлация.
Трето,това е жълтъкът, който включва протеини, фосфолипиди, неутрални мазнини, въглехидрати, минерални соли. Жълтъкът е запас от хранителни вещества и енергийни ресурси, необходими за осигуряване на ембрионалния период. Много вещества, произведени от черния дроб, навлизат в ово(оо)гонията по време на техния растеж през фоликуларните клетки на яйчника. Това изисква голям разход на енергия от женската.
четвърто,Това са специфични вещества, наречени морфогенетични детерминанти,доста твърдо определящи съдбата на клетките (бластомерите), в които попадат по време на разцепването. Подобни вещества не се намират в цитоплазмата на яйцата при всички животни.
Поради описаните особености на химичния състав на цитоплазмата на яйцето, ембрионът по време на периода на смачкване в някои случаи не използва генетичната информация на ядрата на зиготата и бластомерите за развитие. Специфични химичен състави правилното разпределение на веществата в цитоплазмата на яйцето са от голямо значение за началните фази на ембриогенезата. Съхранените хранителни и енергийни вещества осигуряват ембрионално развитие без допълнителен прием отвън.
7.2.3. РАЗМЕР НА ЯЙЦЕТО И ТЯХНАТА РОЛЯ В ЕВОЛЮЦИЯТА. ВИДОВЕ ЯЙЦА
В процеса на развитие се разкрива закономерност, която се състои в това, че колкото по-дълъг е ембрионалният период, толкова повече жълтък трябва да се натрупа в яйцето. Продължителността на ембрионалния период зависи от етапа, на който ембрионът преминава към самостоятелно съществуване във външната среда. Ако постембрионалното развитие е директно, т.е. без ларва и метаморфоза, тогава в яйцето трябва да има повече жълтък. Според броя на жълтъка, яйцата на хордата (Таблица 7.1) се разделят на алецитален, олиго-, мезо-И полилецитален,тези. с незначително малко, малко, средно и голямо количество жълтък (от гръцки. лецитос- жълтък) (Таблица 7.2).
Таблица 7.1.Систематика на типа хордови
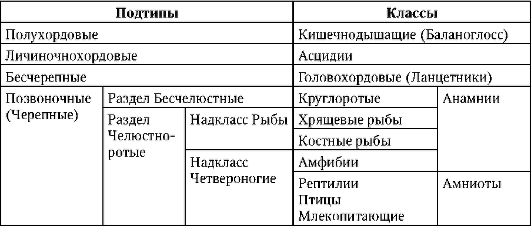
Таблица 7.2.Типове овоцити, открити в хордовите

При копие,представител на долните хордови, яйц олиголецитален.При повечето гръбначни животни яйцата съдържат значително количество жълтък. Сред нисшите гръбначни (Анамния)най-големите яйца се намират при рибите (клас Cyclostomes), при акулите и химерите (класът Хрущялни риби) и при някои земноводни. Остатъка земноводно,както и яйца от есетра мезолецитален,тези. имат средно количество жълтък. При висшите гръбначни (амниота)като влечуги, птици и яйценосни бозайници, полилецитал,тези. в яйцето има много жълтък. Ембрионалното им развитие отнема особено дълго време.
Този модел е нарушен при торбестите и плацентаренбозайници, които имат олиго-И алециталяйца, съответно. При торбестите животни ембрионът излиза от яйчните мембрани и матката с незавършена органогенеза, прехвърля се в торбичката, където продължава развитието си. В плацентата, включително при хората, ембрионът излиза
от мембраните на яйцеклетката още по-рано, на етапа на бластоциста, но след това преминава към вътрематочно съществуване, където завършва всички основни периоди на развитие, които го подготвят за раждането. Намаляванеможе да се нарече количеството жълтък в яйцата на бозайниците втори,тъй като техните предци, които овладяха земна среда, имаше, както всички амниоти, полилецитни яйца.
7.2.4. ПОЛЯРИТЕТ НА OVA
Полярност на ооцититепланира се дори на етапа на натрупване на жълтък в ово(оо)цитите по време на техния бърз (голям) растеж и се фиксира по време на изолирането на полярни (редукционни) тела. След изолирането на второто редукционно тяло полярността става стабилна и необратима, което се доказва от експериментите на Guerrier за центрофугиране на яйца на различни етапи от тяхното узряване. Полюсът, на който се открояват редукционните тела, се нарича животно,и обратното е вегетативен.Полюсите на яйцеклетката се различават по много начини: концентрация различни вещества, броят на органелите, активността на вътреклетъчните процеси и редица други. Да, експерименти последните годиниизползване на вибриращи електроди разкри електрически полетаоколо овоцитите и яйцата на редица животни и растения и протичането на електрически ток през тяхната цитоплазма. Смята се, че това се дължи на различни концентрации на йонни канали и помпи на противоположните полюси на овоцитите. В яйцето на жаба с нокти концентрацията на йонни канали е по-висока на анималния полюс, а концентрацията на помпи е по-висока на вегетативния полюс.
Натрупване на жълтък от яйцето- първата проява на неговата поляризация. С малко количество жълтък в яйцето обикновено се разпределя равномерно в цитоплазмата, а ядрото се намира приблизително в центъра. Тези яйца се наричат изолецитален(от гръцки. isos- равен). Повечето гръбначни животни имат много жълтък и той е неравномерно разпределен в цитоплазмата на яйцето. Това анизолециталклетки. По-голямата част от жълтъка се натрупва в вегетативен полюс.Тези яйца се наричат телолецитален(от гръцки. telos- край). Към обратното животносвободната от жълтъка активна цитоплазма се избутва настрани към полюса. Ако жълтъкът все още е потопен в цитоплазмата и не е изолиран от нея като отделна фракция, както при есетрите и земноводните, яйцата се т.нар. умерено телолецитален.Ако жълтъкът е напълно отделен от цитоплазмата, както при амниотите, тогава това рязко телолециталеняйца.
Характеристиките на размера и полярността на яйцата на хордовите са групирани схематично в табл. 7.2.
В процесите на поляризация на яйцето, очевидно, участва и кора- Това е повърхностният слой на цитоплазмата на яйцето, разположен непосредствено под плазмената мембрана. Съдържа микрофиламенти и кортикални гранули.Последните съдържат цял набор от вещества, участващи в образуването на мембраната за оплождане и предотвратяване на полиспермията. Сглобяването и преразпределението на елементите на цитоскелета на кортикалния слой, наблюдавани след оплождането, осигуряват придобиването на контрактилност от него, което е необходимо за деленията на разцепването на зиготата. При повечето животни първите две бразди на разцепване преминават по взаимно перпендикулярни животински-вегетативни равнини (меридиани, свързващи животинския и вегетативния полюс).
Като цяло, на анималния полюс на яйцеклетката, като правило, има повече свободна цитоплазма, органели, съхранена РНК, обикновено ядрото също се намира на анималния полюс или по-близо до него. Вегетативният полюс се характеризира с преобладаване на жълтъчни гранули.
Поляризацията на яйцеклетката е придружена от появата ovo(oo)-плазмена сегрегацияяйца, т.е. създаване на вътрешна хетерогенност на участъците от цитоплазмата на яйцето.
Животинско-вегетативната поляризация на яйцето е от решаващо значение за всички последващи процеси на ембриогенезата, тъй като определя бъдещата пространствена организация на ембриона. При възрастни животни предно-задната ос на тялото съвпада с животинско-вегетативната ос на яйцето (например при гръбначни) или е перпендикулярна на нея (например при олигохети и някои членестоноги).
7.2.5. ЧЕРЕПИ НА ЯЙЦА
Яйцата са покрити отвън с една или повече мембрани, които по-късно изпълняват, наред с други неща, функцията за защита на развиващия се ембрион.
Разграничете първична обвивка,образувани от самата яйцеклетка вторична обвивка- продукт от дейността на яйчниковите фоликуларни клетки и третични черупки,с които яйцето е заобиколено по време на преминаване през яйцепровода.
първична обвивка,понякога се нарича жълтък,открити в яйцата на всички животни. При гръбначните, включително бозайниците,
първичната обвивка е част от плътната обвивка, образувайки нейната вътрешна част. Външната част на плътната черупкапроизвеждат фоликуларни клетки вторична обвивка.Плътната обвивка е пронизана отвътре от микровласинките на яйцето, а отвън - от микровласинките на фоликуларните клетки. За техните оптични свойствапри бозайниците се нарича лъскава черупка(зона пелуцида).Така че тази черупка съчетава основно и второстепенно.На върха на zona pellucida е сияйна корона(корона радиата),образувани от фоликуларни клетки, които се придържат към яйцеклетката, докато е в яйчниковия фоликул (фиг. 7.2).
Zona pellucida е сложен комплекс от извънклетъчни гликопротеини, които при бозайниците се наричат ЗП.При мишки се състои от три различни сулфатирани гликопротеина - ZP1, ZP2, ZP3. Като се използва ZP3сперматозоидите се свързват със зоната пелуцида и след проникването на един сперматозоид този гликопротеин се модифицира и проникването на друг
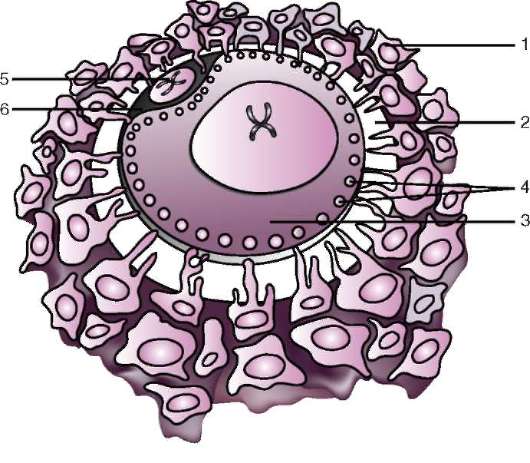
Ориз. 7.2.Яйцеклетка (овоцит от първи ред) на бозайници: 1 - лъчиста корона; 2 - лъскава черупка; 3 - цитоплазма; 4 - кортикални гранули; 5 - полярно тяло; 6 - периолково пространство
gih сперматозоиди става невъзможно. Освен това гликопротеините са специфични за вида, което предотвратява междувидовото оплождане. При други хордови видовата специфика на оплождането се определя от взаимодействието на протеините на спермата с рецепторите. жълтъчна мембрана. Zona pellucida не приляга директно към мембраната на яйцето, а се отделя перивителиново (периотелиново) пространство.След проникването на първия сперматозоид в яйцеклетката, овопероксидазата на кортикалните гранули навлиза в това пространство. Смята се, че действието на този ензим се променя ZP3И Zp2,което води до втвърдяване на zona pellucida. Той персистира около ембриона през целия предимплантационен период или в значителна част от този период. Блестящата обвивка предотвратява слепването на съседните ембриони и залепването на ембрионите по стените на яйцепровода и матката. Известно е, че в началните етапи на раздробяване до бластоциста бластомерите имат висока адхезивност. Ако ембрион, лишен от zona pellucida, бъде трансплантиран в яйцепровода, бластомерите неизбежно ще се прилепят към стената на яйцепровода и ембрионът ще умре. Освен това, благодарение на zona pellucida, бластомерите са компактни и подредени, което спомага за образуването на контакти и взаимодействие между тях и осигурява нормалното развитие на ембриона на този етап. Ако обвивката се отстрани, тогава фрагментацията ще продължи, но бластомерите ще бъдат подредени във верига и тяхното уплътняване ще бъде напълно нарушено или ще бъде много късно.
Третични черупкидобре развити при хрущялните риби и земноводните, но придобиват особена сложност при сухоземните гръбначни животни - влечуги, птици и низши бозайници. Формиране от секретите на жлезите на яйцепровода,тези черупки нямат клетъчна структура. При всички гръбначни животни те изпълняват функциите за защита на ембриона от механични повреди и действието на вредни биотични фактори, като бактериални, гъбични и протозойни фактори. В допълнение, фундаментално нови функции за съхранение на вода и хранителни вещества се появяват при сухоземните гръбначни животни, за да задоволят нуждите на ембриона. При влечугите мембраната на черупката действа като помпа, извличайки вода от почвата и въздуха. Птиците имат водоснабдяване протеинова обвивка.Абсорбцията и изпарението на водата се регулира от пори в черупковата мембрана. Черупката съдържа много минерални соли, необходими за развитието на скелета на ембриона.
7.3. ОПЛОЖДАНЕ И ПАРТЕНОГЕНЕЗА
Оплождане- това е процес на сливане на зародишни клетки, завършващ с обединяването на техния генетичен материал. Диплоидната клетка, получена в резултат на оплождане зигота- представлява началния етап от развитието на нов организъм.
Процесът на оплождане се състои от три последователни фази:
Дистанционно взаимодействие и конвергенция на гамети;
Контактно взаимодействие на гамети и активиране на яйцеклетката;
Сливане на гамети или сингамия.
Сближаване на сперматозоида с яйцеклеткатаосигурени от комбинация от неспецифични фактори, които увеличават вероятността от тяхната среща и взаимодействие. Те включват координацията на началото на готовността за оплождане при мъжките и женските, поведението на мъжките и женските, което осигурява копулация и осеменяване, прекомерното производство на сперматозоиди, големия размер на яйцето, както и тези, произведени от яйца и сперматозоиди химически вещества, допринасяйки за сближаването и взаимодействието на зародишните клетки. Тези вещества, т.нар хамони(хормоните на гаметите), от една страна, активират движението на сперматозоидите, а от друга страна, тяхното свързване. При движението на сперматозоидите на бозайници по горните части на яйцепровода, явлението реотаксис:способността им да се движат срещу идващия поток от течност във фалопиевите тръби.
При бозайниците престоят на сперматозоидите в женските полови пътища е от голямо значение, в резултат на което капацитет на спермата- придобиването им на оплождаща способност. Непосредствено след навлизане в гениталния тракт женските сперматозоиди не са в състояние да проникнат в яйцеклетките. Гликопротеините и семенните плазмени протеини се отстраняват от плазмолемата на сперматозоидите в областта на акрозомата под действието на веществата от секрета на женския генитален тракт, които блокират активните центрове на рецепторните молекули на плазмолемата на спермата, разпознавайки повърхността на женската репродуктивна клетка. В допълнение, молекулите на албумина, намиращи се в женския генитален тракт, се свързват с холестерола на мембраната на сперматозоидите. Това води до дестабилизиране на плазмалемата на спермата и нейната акрозомна мембрана, което улеснява последващото освобождаване на акрозомни ензими. Установено е също, че в процеса на капацитация се променят свойствата на повърхността на сперматозоидите, като например нейния заряд. Освен това има промяна в подвижността (хиперактивация) на сперматозоидите и активиране на акрозомните ензими. При хората капацитетът продължава около 7 часа.
Във фаза контактно взаимодействиеспермата разрушава мембраните на яйцеклетката (при бозайниците - лъчистата корона, прозрачната мембрана) и цитоплазмената мембрана на яйцеклетката поради акрозомна реакция.При контакт с обвивката на женската зародишна клетка под действието на нейните активиращи вещества (едно от които е фертилизин) се инициира активен поток от калциеви катиони в главата на спермата. В резултат на това се получава фокално сливане на клетъчните и акрозомалните мембрани на сперматозоидите и тяхното частично разрушаване. През образуваните микродупчици се отделят сперматозоиди - хиалуронидаза, пенетраза и други, които прекъсват контактите между клетките на лъчистия венец, както и между тях и овоцита. Акрозомният ензим акрозин разрушава част от зоната пелуцида на женската зародишна клетка и сперматозоидите проникват в околоплодното пространство. В точката на контакт на главата на спермата с плазмената мембрана на яйцеклетката се получава сливане и последващо разрушаване на мембраните на женските и мъжките гамети. Чрез получения цитоплазмен мост цитоплазмата на двете гамети се комбинира. След това ядрото и центриола на сперматозоида преминават в цитоплазмата на яйцеклетката, а мембраната на сперматозоида е вградена в мембраната на яйцеклетката. Опашната част на сперматозоида или остава отвън, или също навлиза в яйцеклетката, но след това се отделя и разтваря, без да играе никаква роля в по-нататъшното развитие.
Натриевите йони започват активно да навлизат в цитоплазмата на яйцето през мембраната на сперматозоидите, в резултат на което мембранният потенциал на ооцита се променя драстично и женската зародишна клетка става имунизирана срещу контакт с други сперматозоиди - бърз блок на полиспермия.Притокът на натриеви йони предизвиква освобождаване на калциеви йони от вътреклетъчните депа и увеличаване на съдържанието му в цитоплазмата на яйцето, което се разпространява под формата на вълна от мястото на контакт на гаметите. След това започва кортикална реакция:мембраните на кортикалните гранули се сливат с мембраната на яйцеклетката и освободените от тях протеолитични ензими навлизат в периолковото пространство. Под въздействието на ензими черупката на яйцето се удебелява, удебелява, губи рецепторни протеини за сперматозоидите и се превръща в обвивка за оплождане.В допълнение, гликопротеинът, секретиран от кортикалните гранули, допринася за отделянето на жълтъчната мембрана от плазмолемата на яйцето. В резултат на всичко по-горе, проникването на други сперматозоиди става невъзможно - бавен блок на полиспермия.При бозайниците
кортикалната реакция не предизвиква образуването на оплодителната мембрана, но нейната същност е същата.
В резултат на контакта на сперматозоида с яйцето, то активиране.Състои се в сложни структурни и физико-химични промени. Началният етап на активиране е кортикалната реакция, описана по-горе. При животни като бодлокожи, костни риби и земноводни, промените в цитоплазмата на яйцето са придружени от видими морфологични пренареждания. Тези явления се наричат стратификация или плазмена сегрегация.Неговото значение за по-нататъшното ембрионално развитие ще бъде обсъдено по-долу. При много видове анурани проникването на сперматозоид в яйцеклетката води до движение на пигментни гранулиживотински полюс и бледо оцветена зона с форма на полумесец се появява срещу мястото на проникване на спермата, т.нар. "сив сърп"(фиг. 7.3).
Активирането на яйцеклетката завършва с началото на протеиновия синтез на ниво транслация, тъй като тРНК, тРНК, рибозомите и енергията са били съхранени обратно в ово(оо)генезата. Активирането на яйцеклетката може да започне и да продължи до края без ядро на сперматозоида и без ядро на яйцеклетка, което е доказано чрез експерименти за енуклеация на зигота.
Яйцеклетката по време на срещата със спермата обикновено е на един от етапите на мейозата, блокирана от специфичен фактор. При повечето гръбначни този блок се осъществява
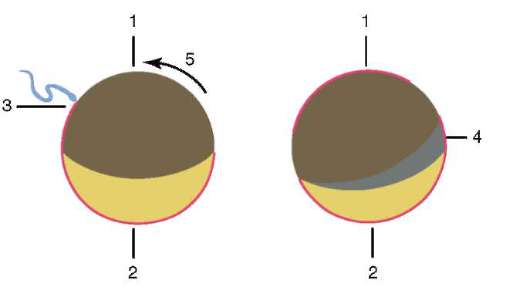
Ориз. 7.3.Сегрегация на цитоплазмата на яйцеклетката на земноводните след проникване на сперма: 1 - животински полюс; 2 - вегетативен полюс; 3 - мястото на въвеждане на сперматозоида; 4 - сив сърп; 5 - посоката на движение на цитоплазмата с пигментни гранули
на етапа на метафаза II; при много безгръбначни, както и при три вида бозайници (коне, кучета и лисици), блокът възниква на етапа на диакинеза (профаза I). В повечето случаи мейотичният блок се отстранява след активирането на яйцеклетката поради оплождането. Докато мейозата е завършена в яйцеклетката, ядрото на спермата, която е проникнала в нея, се модифицира. Приема формата на интерфазно и след това профазно ядро. През това време ДНК се удвоява, а количеството наследствен материал V мъжки пронуклеусстава p2s, тези. той съдържа хаплоиден набор от редупликирани хромозоми.
Ядрото на завършилата мейоза яйцеклетка става женски пронуклеус,също придобиване p2s. И двата пронуклеуса извършват сложни движения, след което се приближават един към друг и се сливат. (синкарион),образуващи обща метафазна плоча. Това всъщност е моментът на окончателното сливане на гаметите - сингамия.Първото митотично делене на зиготата води до образуването на две ембрионални клетки (бластомери) с набор от хромозоми 2p2s във всеки.
В някои случаи развитието протича без оплождане - партеногенеза(от гръцки. партеное- девствена). Кога естествена партеногенезаразвитието протича на базата на цитоплазмата и пронуклеуса на яйцето. Индивидите, които се образуват от яйцеклетка, имат хаплоиден или диплоиден набор от хромозоми, тъй като се задейства един от механизмите за удвояване на броя на хромозомите. В някои случаи, по време на мейозата на женската зародишна клетка, етапът на намаляване на броя на хромозомите отпада и яйцето се получава с диплоиден пронуклеус. В други случаи диплоидизацията настъпва по време на първото делене на разцепване, при което не настъпва цитотомия.
естествена партеногенеза,като правило не е единственият начин за възпроизвеждане на вида. То или се редува с нормално сексуално размножаване, или се среща при индивида състезания.Естествена партеногенеза е установена в летните поколения на някои ракообразни и ротифери, при пчели, оси и редица Lepidoptera. Сред гръбначните животни партеногенетичното размножаване е описано в три раси на скалния гущер от Армения, състоящи се само от женски. 40% от пуешките яйца, снесени в отсъствието на мъж, могат да започнат да се развиват, но това развитие рядко достига до края, по-често спира поради аномалии. При други видове гръбначни естественото партеногенетично размножаване не е известно.
изкуствена партеногенезаизглежда възможно при всички животни. Важно е разработването на методи за партеногенетично развитие
проблем в научно-приложните отношения. Голям принос към този проблем направиха местните изследователи A.A. Тихомиров, Б.Л. Астауров, В.А. Струников. Установено е, че активирането на яйцеклетката от сперматозоида не е специфично. Много физични и химични фактори могат да действат като активатори. На копринената буба е показано, че с помощта на изкуствена партеногенеза е възможно да се регулира съотношението на мъжкия и женския пол в популацията, като се получава голям икономически ефект.
Естествената партеногенеза възниква най-често при непълно оплождане, т.е. в случаите, когато се е случило активиране на яйцеклетката, но ядрото на сперматозоида не е участвало в оплождането. В активираните яйцеклетки се използва само информация от женския пронуклеус. Този тип партеногенеза се нарича гиногенеза.При изкуствена партеногенеза е възможно да се отстрани женският пронуклеус и след това развитието ще се извърши само за сметка на мъжките пронуклеуси. Това андрогенеза.При специални експерименти с морски таралежи беше установено, че потомството наследява или само чертите на майката по време на гиногенезата, или само чертите на бащата по време на андрогенезата. Това показва, че наследствените свойства на индивида се определят главно от ядрото, а не от цитоплазмата.
7.4. ембрионално развитие
7.4.1. РАЗДЕЛЯНЕ
7.4.1.1. Същността на етапа на смачкване
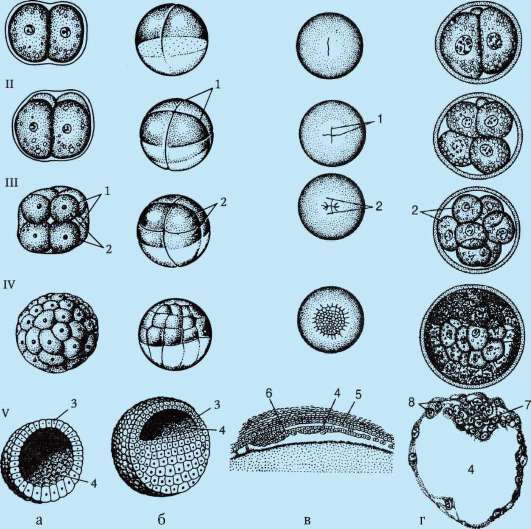
Разделяне- това е поредица от последователни митотични деления на зиготата и получените бластомери, завършващи с образуването многоклетъчен еднослоен ембрион- бластула.Първото делене на разцепване започва след обединяването на наследствения материал на пронуклеусите и образуването на обща метафазна пластина. Клетките, образувани по време на разцепването, се наричат бластомери(от гръцки. взривявам- кълнове, зародиш). Характеристика на митотичните цикли на раздробяване е, че с всяко делене клетките стават все по-малки и по-малки, докато достигнат съотношението на обемите на ядрото и цитоплазмата, което е обичайно за соматичните клетки. При морския таралеж например това изисква 6 деления, а ембрионът се състои от 64 клетки. Няма клетъчен растеж между последователните деления (период G 1 отсъства), но ДНК задължително се синтезира. Всички прекурсори на ДНК и необходими
Димируемите ензими се натрупват в процеса на оогенезата. В резултат митотичните цикли се съкращават и деленията следват много по-бързо, отколкото в обикновените соматични клетки. След няколко цикъла на делене, образуваните бластомери се прилепват един към друг, образувайки клъстер от клетки, т.нар. морула.Тогава между клетките се образува кухина - бластоцел,пълни с течност. Клетките се изтласкват към периферията, образувайки стената на бластулата - бластодермия.Поради липсата на растеж на бластомера по време на периода на разцепване, общият размер на ембриона в стадия на бластула не надвишава размера на зиготата.
7.4.1.2. Морфология на смачкване
По правило бластомерите са подредени в строг ред един спрямо друг и полярната ос на яйцето. Редът, или начинът на разбиване зависи от количеството, плътността и разпределението на жълтъка в яйцето. Според правилата на Sachs-Hertwig клетъчното ядро има тенденция да бъде разположено в центъра на цитоплазмата, свободна от жълтъка, а вретеното на клетъчното делене - в посока на най-голямото разпространение на тази зона.
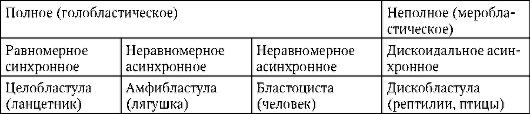
В олиго- и мезолециталните яйца, разцепване пълен,или холобластичен.Този тип смачкване се среща при ланцетници, миноги, някои риби, всички земноводни, както и торбести и плацентарни бозайници. При пълно смачкване равнината на първото деление съответства на равнината на двустранна симетрия. Равнината на второто деление е перпендикулярна на равнината на първото. И двете бразди на първите две деления са меридианни, т.е. започва от анималния полюс и се разпространява към вегетативния полюс. Яйцеклетката е разделена на четири повече или по-малко еднакви по размер бластомери. Равнината на третото деление е перпендикулярна на първите две в посока на ширината. При животни с изолецитни яйца, всичките осем образувани бластомера са приблизително еднакви - равномерно раздробяване.В мезолециталните яйца се появява нередностсмачкване. На анималния полюс има четири по-малки бластомера - микрометри,върху вегетативното - четири по-големи - макромери.След това разделянето отново преминава в меридианните равнини, а след това отново в ширините.
В полилецитни ооцити на костни риби, влечуги, птици, а също и еднопроходни бозайници, разцепване частично,или меробластичен,тези. покрива само цитоплазмата, свободна от жълтък. Разположен е под формата на тънък диск на анималния полюс, поради което този вид смачкване се нарича дискоидален.
При характеризиране на вида на смачкване се вземат предвид и относителното положение и скоростта на разделяне на бластомерите. Ако бластомерите са подредени в редове един над друг по радиусите, раздробяването се нарича радиална.Характерен е за хордови и бодлокожи. В природата съществуват и други варианти на пространствено разположение на бластомерите при раздробяване, което определя видовете му, като напр. спиралав миди двустраннопри кръгли червеи, анархиченпри медуза.
Отбелязана е връзка между разпределението на жълтъка и степента на синхронност в разделянето на животински и вегетативни бластомери. В олиголециталните яйца на бодлокожите и главохордите разцепването е почти синхронно; в мезолециталните яйца, например при земноводните, синхронът се нарушава след третото делене, тъй като вегетативните бластомери се делят по-бавно поради голямото количество жълтък. При организми с частично разцепване (влечуги, птици) деленията са асинхронни от самото начало и бластомерите, заемащи централно положение, се делят по-бързо.
До края на смачкване, бластула.Видът на бластулата зависи от вида на раздробяването и следователно от вида на яйцето. Някои видове смачкване и бластула са показани на фиг. 7.4 и схема (в таблицата) 7.1. За по-подробно описание на разцепването при бозайници, включително хора, вижте раздел 7.5.1.
Таблица 7.3.Видове раздробяване и видове хордови бластули
7.4.1.3. Характеристики на молекулярно-генетичните и биохимични процеси при раздробяване
Както беше отбелязано по-горе, митотичните цикли по време на периода на разцепване са значително съкратени, особено в самото начало. Например, целият цикъл на делене в яйца на морски таралеж продължава 30-40 минути с продължителност от S-фазисамо 15 мин.
Жj-,и при много животни G 2- периодипрактически липсват, тъй като необходимият запас от всички вещества е създаден в цитоплазмата на яйцеклетката и колкото по-голям, толкова по-голям е той. Доказано е, че при отстраняване от зиготата
Ориз. 7.4.Разцепване при хордови с различни видове яйца: а - ланцет; б - жаба; c - птица; d - бозайник: I - 2 бластомера; II - 4 бластомера; III - 8 бластомера; IV - морула; V - бластула; 1 - смачкващи бразди; 2 - бластомери; 3 - бластодерма; 4 - бластоцел; 5 - епибласт; 6 - хипобласт; 7 - ембриобласт; 8 - трофобласт; размерът на ембрионите на фигурата не отразява истинския размер на ембрионите
настъпва разцепване на ядрото и ембрионът достига в развитието си почти до стадий бластула. По-нататъшното развитие спира.
Преди всяко делене се случва синтеза на ДНК и хистони, но продължителността S-периодсъкратен. По време на фрагментация по време на репликация на ДНК скоростта на напредване на вилицата за репликация е нормална. В същото време има повече начални точки в бластомерната ДНК, отколкото в
соматични клетки, така че репликоните са по-къси. Синтезът на ДНК се извършва във всички репликони едновременно (синхронно). Следователно времето на репликация на ДНК в ядрото съвпада с времето на удвояване на един, освен това съкратен, репликон.
В началото на разцепването други видове ядрена активност, като транскрипция, практически отсъстват. В различните видове яйцеклетки генната транскрипция и синтезът на РНК започват на различни етапи. В случаите, когато в цитоплазмата има много различни вещества, като например при земноводните, транскрипцията не се активира веднага. Синтезът на РНК в тях започва на етапа на ранната бластула. Напротив, при бозайниците синтезът на РНК започва още на етапа на два бластомера.
По време на периода на разцепване се образуват РНК и протеини, подобни на тези, синтезирани по време на оогенезата. Предимно хистони, протеини клетъчни мембрании ензими, необходими за клетъчното делене. Тези протеини се използват незабавно заедно с тези, съхранявани преди това в цитоплазмата на яйцата. Заедно с това по време на периода на разцепване е възможен синтезът на протеини, които преди това не са били възможни, което се доказва от данните за наличието на регионални различия в синтеза на РНК и протеини между бластомерите. Понякога тези вещества започват да действат на по-късни етапи.
Важна роля при раздробяването играе разделянето на цитоплазмата - цитотомия.Има особено морфогенетично значение, тъй като определя вида на смачкване. В процеса на цитотомия първо се образува стеснение с помощта на контрактилен пръстен от микрофиламенти. Сглобяването на този пръстен става под прякото влияние на полюсите на митотичното вретено. След цитотомия бластомерите на олиголецитните яйца остават свързани помежду си само чрез тънки мостове. По това време те се разделят най-лесно. Това е така, защото цитотомията води до намаляване на площта на контакт между клетките поради ограничената повърхност на мембраните.
Веднага след цитотомията започва синтезът на нови участъци от клетъчната повърхност, контактната зона се увеличава и бластомерите започват да се докосват плътно, което осигурява възможност за междуклетъчно взаимодействие. По границите между отделните участъци на овоплазмата минават бразди за разцепване, така че цитоплазмата на различните бластомери се различава по химичен състав. Това отразява явлението овоплазмена сегрегация.
Основният резултат от периода на смачкване е трансформацията на едноклетъчен ембрион - зигота - в многоклетъченИ един слойембрион – бластула. Освен това, по време на раздробяването, образуването на
инж междуклетъчни контактии увеличаване на тяхната площ, разпределението на различни участъци от цитоплазмата на зиготата между различни бластомери на ембриона, постепенното активиране на собствения геном на ембриона, възстановяването на ядрено-цитоплазменото съотношение, характерно за соматичните клетки. На етапа на раздробяване всички клетки на ембриона са хомогенни по отношение на функционалното състояние на генетичния апарат. Разликата в цитоплазмата на бластомерите и междуклетъчните взаимодействия определят (предопределят) посоката на по-нататъшното развитие на клетката.
7.4.2. ГАСТРУЛАЦИЯ
7.4.2.1. Същността на етапа на гаструлация
Същността на етапа на гаструлация е, че еднослоен ембрион - бластула - се превръща в многопластов- две-или трислоен,Наречен гаструла(от гръцки. гастър- стомах и умалителна наставка "st").
При примитивните хордови, например в ланцета, хомогенна еднослойна бластодерма по време на гаструлацията се трансформира във външен зародишен слой - ектодерма- и вътрешния зародишен слой - ендодерма.Ендодермата образува първичното черво с кухина вътре - гастроцел.Дупката, водеща до гастроцела, се нарича бластопорили първична уста. Два зародишни слояса определящи морфологичен признакгаструлация. Тяхното съществуване на определен етап от развитието при всички многоклетъчни животни, от кишечнополовите до висшите гръбначни, ни позволява да мислим за хомологията на зародишните слоеве и единството на произхода на всички тези животни.
При гръбначните освен двата споменати зародишни листа по време на гаструлацията се образува и трети – мезодерма,разположени между екто- и ендодермата. Развитието на средния зародишен лист, който е хордомезодерма,- еволюционно усложнение на фазата на гаструлация при гръбначните животни, свързано с ускоряването на тяхното развитие в ранните етапи на ембриогенезата. При по-примитивните хордови, като ланцетника, хордомезодермата обикновено се образува в началото на фазата след гаструлацията - органогенеза.Изместването във времето на развитие на някои органи спрямо други в потомците в сравнение с предшествениците е проява на хетерохронност.Промените във времето на формиране на най-важните органи в хода на еволюцията са доста чести.
Характеризира се процесът на гаструлация важни клетъчни трансформации,като насочени движения на групи и отделни клетки, селективно размножаване и сортиране на клетки, начало на цитодиференциация и индукционни взаимодействия. Изброените клетъчни механизми на онтогенезата са разгледани подробно в раздел 8.2.
Методите за гаструлация са различни. Разграничават се четири вида пространствено насочени клетъчни движения, водещи до превръщането на ембриона от еднослоен в многослоен (дву- или трислоен).
Инвагинация- инвагинация на един от участъците на бластодермата навътре като цял слой. В ланцетата клетките на вегетативния полюс се инвагинират; при земноводните инвагинацията настъпва на границата между животинския и вегетативния полюс в областта на сивия полумесец. Процесът на инвагинация е възможен само при яйца с малко или средно количество жълтък.
епиболия- замърсяване с малки бързо делящи се клетки на анималния полюс на по-големи, изоставащи в скоростта на делене и по-малко подвижни клетки на вегетативния полюс. Този процес е ясно изразен при земноводните.
Разслояване- стратификация на клетките на бластодермата в два слоя, разположени един над друг. Деламинация може да се наблюдава в дискобластулата на ембриони с непълно раздробяване на животни като влечуги, птици и яйценосни бозайници. Деламинацията се проявява в ембриобласта на плацентните бозайници, което води до образуването на хипобласт и епибласт.
Имиграция- движение на групи или отделни клетки, които не са обединени в един слой. Имиграцията се среща при всички ембриони, но е най-характерна за втората фаза на гаструлацията при висшите гръбначни (влечуги, птици и бозайници).
Във всеки конкретен случай на ембриогенеза, като правило, се комбинират няколко метода на гаструлация.
7.4.2.2. Морфология на гаструлацията
По-подробно изследване на гаструлацията при ланцетника, жабата, пилетата и бозайниците ще помогне да се разберат по-добре еволюционните връзки и да се разберат моделите на индивидуалното развитие.
7.4.2.2-а. Гетрулация на ланцетника
гаструлация ланцетникпоказано на фиг. 7.5. Маркирани са различни маркери на етапа на бластула (фиг. 7.5, а). предполагаем(предполага се) рудименти.Това са участъци от бластулата, от клетъчния материал
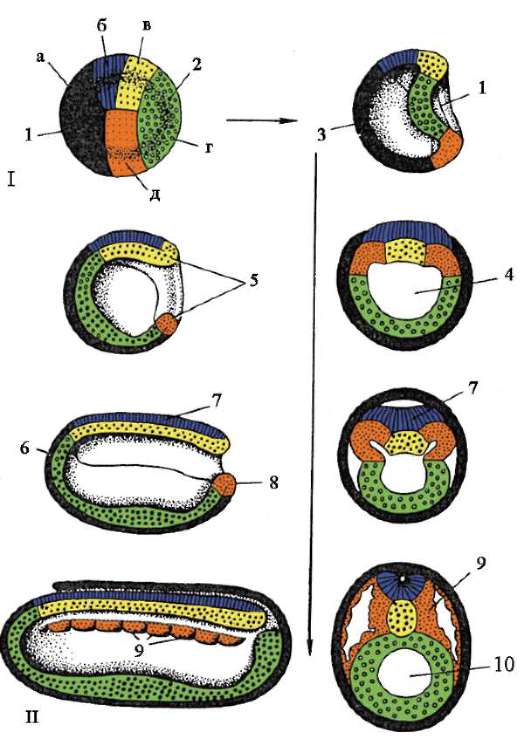
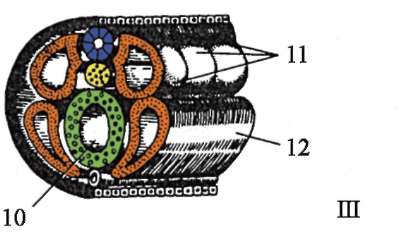
Ориз. 7.5.Предполагаеми зачатъци, гаструлация и неврулация в ланцетник. I - предполагаеми зачатъци на етапа на бластула (изглед отвън) и ранна гаструла (изглед в разрез); II - късна гаструла и неврулация на сагиталните (ляв ред) и напречните (десен ред) секции; III - пластмасов модел на ембриона в края на периода на неврулация:
1 - животински стълб; 2 - вегетативен полюс; 3 - бластоцел; 4 - гастро-гол; 5 - дорзални и вентрални устни на бластопора; 6 - главата на ембриона; 7 - медуларна плоча; 8 - опашката на ембриона; 9 - дорзалната част на мезодермата; 10 - кухина на вторичното черво; 11 - сегментирани сомити; 12 - коремна част на мезодермата; a, b, c, d, e - обозначения на предполагаеми и развиващи се органи: a - кожна ектодерма; b - неврална тръба; в - акорд; d - ендодерма, чревен епител; d - мезодерма
от които по време на гаструлацията и ранната органогенеза (неврулация) обикновено се образуват напълно дефинирани зародишни слоеве и органи (фиг. 7.5, b и c).
Инвагинацията започва от вегетативния полюс. Благодарение на по-бързото делене клетките на анималния полюс растат и избутват клетките на вегетативния полюс в бластулата. Това се улеснява от промяна в състоянието на цитоплазмата в клетките, които образуват устните на бластопора и съседните на тях. Поради инвагинацията бластоцелът намалява, а гастроцелът се увеличава. Едновременно с изчезването на бластоцела, ектодермата и ендодермата влизат в близък контакт. Ланцетът, като всички останали дейтеростоми(те включват тип Echinodermata, тип Chordata и някои други сравнително малки видове животни), областта на бластопора се превръща в опашната част на организма, за разлика от протостомите, в които бластопорът съответства на частта на главата. Отваряне на устата при дейтеростомитеобразуван в края на ембриона срещу бластопора.
7.4.2.2-6. Гаетрулация при земноводните
Гаструлацията при земноводните има много общо с гаструлацията на ланцетника, но тъй като жълтъкът в техните яйца е много по-голям и е разположен главно на вегетативния полюс, големите бластомери на амфибластулата не могат да изпъкнат навътре. Инвагинациявърви малко по-различно. На границата между животинския и вегетативния полюс в областта на сивия сърп клетките първо силно се разтягат навътре, приемайки формата на "колба" (фиг. 7.6, а), а след това издърпват клетките на повърхностния слой на бластулата заедно с тях. Появяват се жлеб във формата на полумесец и гръбна (дорзална) устна на бластопора (фиг. 7.6, b).
В същото време по-малките клетки на анималния полюс, като се делят по-бързо, започват да се придвижват към вегетативния полюс. В областта на дорзалната устна те се обръщат нагоре и инвагинират, а по-големи клетки растат отстрани и от страната, противоположна на сърповидната бразда. След това процесът епиболияводи до образуването на странични (странични) и вентрални (вентрални) устни на бластопора. Бластопорът се затваря в пръстен, вътре в който известно време се виждат големи светли клетки на вегетативния полюс под формата на т.нар. жълтък щепсел.По-късно те са напълно потопени навътре и бластопорът се стеснява.
С помощта на метода за маркиране с витални (жизненоважни) багрила при земноводните са изследвани подробно клетъчните движения
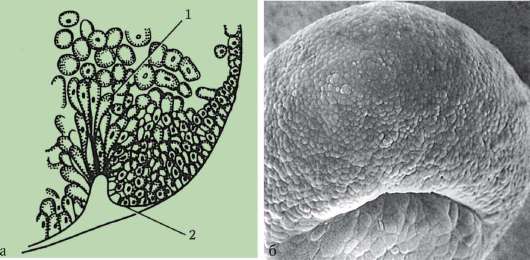
Ориз. 7.6.Образуване на бластопора на ранната гаструла на земноводни: а - клетки с форма на колба в областта на дорзалната устна на бластопора; б - общ изглед на фалциформен жлеб и гръбначна устна; 1 - клетки с форма на колба; 2 - дорзална устна на бластопора
бластула по време на гаструлацията е установено, че специфични участъци от бластодермата, т.нар предполагаем(от лат. praesumptio- предположение), по време на нормалното развитие те първо се появяват в състава на определени рудименти на органи, а след това в състава на самите органи (фиг. 7.7). Известно е, че при безопашатите земноводни материалът на презумптивната нотохорда и мезодермата на етапа на бластула не лежи на повърхността му, а във вътрешните слоеве на стената на амфибластулата, но приблизително на същите нива, както е показано на фигурата.
Известно сходство между процесите на гаструлация и областите на презумптивните органи при земноводните и ланцетните, т.е. хомологията на основните органи, като невралната тръба, хордата, вторичното черво, показва тяхната филогенетична връзка.
7.4.2.2-c. Гаструлация при птици
Гаструлацията при ембриони с меробластичен тип разцепване и развитие има свои собствени характеристики. При птицизапочва след разцепването и образуването на бластулата при преминаването на ембриона през яйцепровода. Към момента на снасяне на яйцето ембрионът вече се състои от няколко слоя: горният слой се нарича епибласт,нисък - първичен хипобластом(виж Фиг. 7.4, c). Между тях има тясна междина - бластоцел. След това се образува вторичен хипобласт,чийто начин на образуване не е напълно ясен. Има данни, че в първич
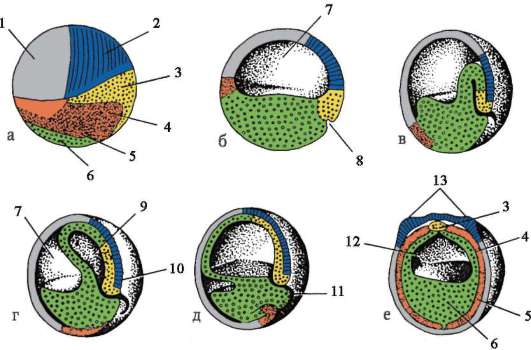
Ориз. 7.7.Карта на областите на предполагаеми рудименти на органи в ранните етапи на ембрионалното развитие на земноводните: а - етап на бластула (изглед отляво); b-e - последователни етапи на гаструлация (сагитални сечения); д - началото на неврулация (напречно сечение); 1 - кожна ектодерма; 2 - неврална тръба; 3 - акорд; 4 - сомитна мезодерма; 5 - мезодерма на спланхнотоми; 6 - ендодерма; 7 - бластоцел; 8 - полумесец жлеб; 9 - гастроцел; 10 - гръбначна устна бластопор; 11 - жълтък щепсел; 12 - кухина на вторичното черво; 13 - нервни ролки
В областта на птиците възникват първичните зародишни клетки, а вторичните образуват екстраембрионална ендодерма. Образуването на първичен и вторичен хипобласт се счита за явление, предхождащо гаструлацията.
Основните събития на гаструлацията и окончателното формиране на трите зародишни слоя започват след яйцеполагането с началото на инкубацията. В задната част на епибласта има натрупване на клетки в резултат на неравномерната скорост на делене на клетките и движението им от страничните части на епибласта към центъра, една към друга. Така нареченият първична линия,който се простира към края на главата. В центъра на първичната ивица се образува първична бразда,и по ръбовете първични ролки.Появява се удебеляване в края на главата на първичната лента - Хенсен възел,и в него - първична ямка(фиг. 7.8).
Когато епибластните клетки навлязат в първичната бразда, тяхната форма се променя. По форма наподобяват "колбовидните" клетки на гаструлата на земноводните. След това тези клетки стават звездовидни и потъват под епибласта, за да образуват мезодермата. Част от мигриращите клетки, вградени в хипобласта, по-късно дават началото на зародишната ендодерма (фиг. 7.9). По този начин ендодермата се формира на базата на първичен и вторичен хипобласт с добавяне на ново поколение ендодермални клетки, мигриращи от горните слоеве на бластодермата. Наличието на няколко поколения ендодермални клетки показва удължаване на периода на гаструлация във времето.
Част от клетките, мигриращи от епибласта през възела на Хенсен, образуват бъдещата нотохорда. Едновременно с анлагирането и удължаването на хордата възелът на Хенсен и първичната ивица постепенно се изместват в посока от предния към опашния край и изчезват. Това съответства на стесняването и затварянето на бластопора. Тъй като първичната ивица се свива, тя оставя след себе си образуваните участъци от аксиалните органи на ембриона в посока от главата към опашката. Изглежда разумно да се разглеждат клетъчните движения в пилешкия ембрион като хомоложни на тези при земноводните, а първичната ивица и възелът на Хенсен като хомоложни на бластопора и дорзалната (гръбна) устна на гаструлата на тези животни.
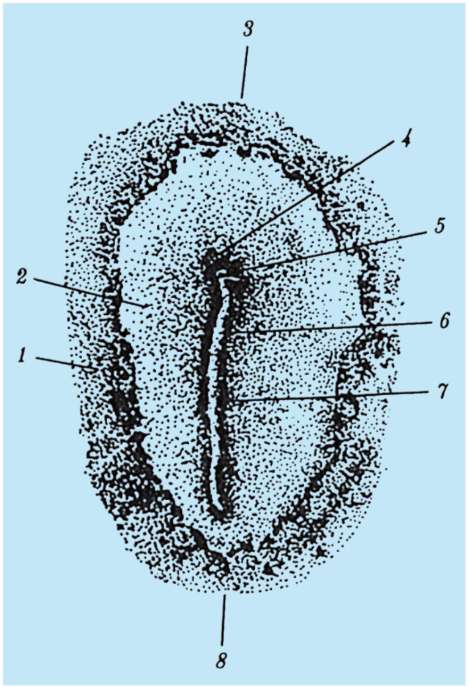
Ориз. 7.8.Пилешки ембрион на етапа на първичната ивица (изглед от гръб): 1 - тъмна зона; 2 - полупрозрачна област на зародишния диск; 3 - край на главата; 4 - възел на Хенсен; 5 - първична ямка; 6 - първичен жлеб; 7 - първична ролка; 8 - край на опашката
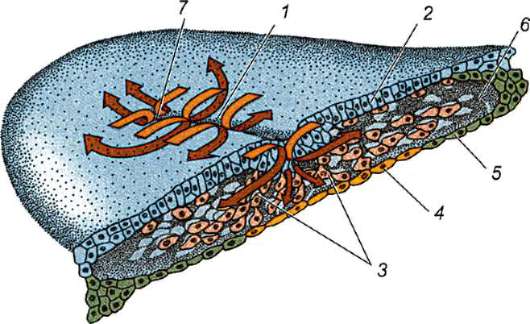
Ориз. 7.9.Пилешки ембрион на етапа на първичната лента (напречно сечение): 1 - първичен жлеб; 2 - епибласт (ектодерма); 3 - мигриращи клетки (мезодерма); 4 - зародишна ендодерма; 5 - хипобласт (ендодерма); 6 - бластоцел; 7 - възел на Хенсен
7.4.2.2-г. Гаструлация при бозайници
Интересно е да се отбележи, че клетките на ембриони на бозайници (виж параграф 7.5.1), въпреки факта, че при тези животни яйцата имат малко количество жълтък и фрагментацията е пълна, във фазата на гаструлация те запазват движенията, характерни за ембриони на влечуги и птици. Това потвърждава идеята за произхода на бозайниците от група предци, чиито яйца са били богати на жълтък.
7.4.2.3. Характеристики на етапа на гаструлация
Гаструлацията се характеризира с различни клетъчни процеси. Митотик продължава клетъчно възпроизвеждане,освен това има различна интензивност в различните части на ембриона. Интерфазите на митотичните цикли включват всички периоди (G 1 , S, G 2), следователно, започвайки от етапа на гаструлация, се наблюдава растеж на развиващия се организъм. Въпреки това, най Характеристикагаструлацията се състои от движение на клетъчни маси.Това води до промяна в структурата на ембриона и превръщането му от бластула в гаструла. продължава сортиранеклетки по принадлежността им към различни зародишни слоеве, вътре в които се "разпознават".
Започва фазата на гаструлация диференциацияклетки, което означава преход към активно използване на биологичната информация на собствения геном. Един от регулаторите на генетичната активност е различният химичен състав на цитоплазмата на ембрионалните клетки, който се установява благодарение на ово(оо)плазмената сегрегация.и двете произхождат от вегетативния полюс на яйцето.
По време на гаструлацията ролята е много голяма ембрионална индукция- взаимодействия между клетъчни комплекси (части) на развиващия се ембрион. Доказано е, че появата на първичната ивица при птиците е резултат от индуктивно взаимодействие между хипобласта и епибласта. Хипобластът има полярност. Промяна в позицията на хипобласта спрямо епибласта причинява промяна в ориентацията на примитивната ивица. Всички тези процеси са описани подробно в глава 8.
7.4.3. ОБРАЗУВАНЕ НА ОРГАНИ И ТЪКАНИ 7.4.3.1. Същността на етапа на органогенезата
органогенеза,състоящи се в образуването на отделни органи, съставляват основното съдържание на ембрионалния период. Те продължават в ларвата и завършват в ювенилния период. Органогенезата се отличава с най-сложните и разнообразни морфогенетични (оформящи) трансформации. Необходима предпоставка за прехода към органогенезата е постигането от ембриона на стадия на гаструла, а именно образуването на зародишни слоеве. Заемайки определено положение един спрямо друг, зародишните листове чрез контакт и взаимодействие осигуряват такива взаимоотношения между различните клетъчни групи, които стимулират тяхното развитие в определена посока. Този т.нар ембрионална индукция- най-важната последица от взаимодействието между зародишните слоеве.
В хода на органогенезата формата, структурата и химичният състав на клетките се променят, изолират се клетъчни групи, които са зачатъци на бъдещи органи. Постепенно се развива определена форма на органи, между тях се установяват пространствени и функционални връзки. Процесите на морфогенеза са придружени от диференциация
клетъчна диференциация и образуване на тъкани, диференциация на тъкани и структури, както и техния избирателен и неравномерен растеж. Предпоставка за органогенезата, заедно с възпроизвеждането, миграцията и сортирането на клетките, е тяхната селективна смърт (вижте глава 8).
7.4.3.2. Неврулация
Самото начало на органогенезата се нарича неврулация. Неврулацияобхваща процесите от появата на първите признаци на образуване на невралната пластинка до нейното затваряне в невралната тръба (фиг. 7.10). По същото време, акордИ вторично черво,и мезодермата, разположена отстрани на хордата, се разделя в краниокаудална посока на сегментирани сдвоени структури - сомити(формира се комплекс от аксиални органи - първична органогенеза).
Нервната система на гръбначните животни, включително хората, се характеризира със стабилност на основния структурен план през цялото време. еволюционна историяподтип. Във формирането на невралната тръба всички хордови имат много общо. Първоначално неспециализирано дорзална ектодерма,в отговор на индукционния ефект от хордомезодермата, той се превръща в неврална пластина,подадено невроепителенцилиндрични клетки.
Невралната пластина не остава сплескана за дълго. Скоро страничните му ръбове се издигат, образувайки нервни гънки,които лежат от двете страни на плитка надлъжна невронна бразда.След това ръбовете на нервните гънки се затварят, образувайки затворена нервна тръба с канал вътре - невроцеле.На първо място, затварянето на нервните гънки се случва на нивото на началото (границата между гръбначния мозък и мозъка) на гръбначния мозък и след това се разпространява в мозъка и опашката.
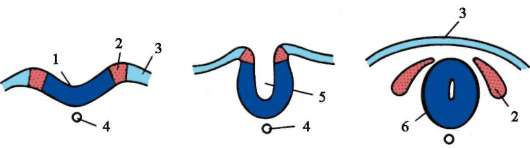
Ориз. 7.10.Последователни етапи в образуването на невралната тръба и невралния гребен (напречно сечение на ембриона): 1 - неврална пластина; 2 - неврален гребен; 3 - ектодерма; 4 - акорд; 5 - нервна бразда; 6 - neurocoel; 7 - нервни гънки
посоки. Доказано е, че микротубулите и микрофиламентите на невроепителните клетки играят важна роля в морфогенезата на невралната тръба. Разрушаване на тези клетъчни структури от колхицин и цитохалазин INкара невралната пластина да остане отворена. Незатварянето на невралните гънки води до вродени малформации на невралната тръба.
След затварянето на невралните гънки се образуват клетките, които първоначално са били разположени между невралната пластина и бъдещата кожна ектодерма нервен гребен.Клетките на нервния гребен се отличават със способността си за екстензивни, но строго регулирани миграции в тялото (виж раздел 8.2.2, фиг. 8.9) и образуват два основни потока. Клетките на един от тях - повърхностен- включват се в епидермиса или дермата на кожата, където се диференцират в пигментни клетки. Друг поток мигрира във вентралната посокаи образува чувствителни спинални ганглии, симпатикови ганглии, надбъбречна медула, парасимпатикови ганглии. Клетките от черепния неврален гребен дават началото както на нервните клетки, така и на редица други структури, като хрилни хрущяли, някои покривни кости на черепа.
Ендодермата във всички ембриони в крайна сметка образува епитела на вторичното черво и много от неговите производни. Самото вторично черво винаги се намира под хордата.
Така в процеса на неврулация се образува комплекс аксиални органи:неврална тръба, хорда, черво, които са най-характерната черта на организацията на тялото на всички хордови. Еднаквият произход, развитие и взаимно разположение на аксиалните органи разкриват тяхната пълна хомология и еволюционна приемственост.
Задълбоченото изследване и сравнение на процесите на неврулация при специфични представители на хордовия тип разкрива някои разлики, които са свързани главно с характеристики, които зависят от структурата на яйцата, метода на смачкване и гаструлация (фиг. 7.11). Обръща се внимание на различната форма на ембрионите и изместването на времето на полагане на аксиалните органи един спрямо друг, т.е. хетерохронност, описана по-горе.
7.4.3.3. диференциация на мезодермата
Мезодермата, която заема място отстрани на хордата и се простира по-нататък между кожната ектодерма и ендодермата на вторичното черво, е разделена на дорзална и вентрална област. Дорсален
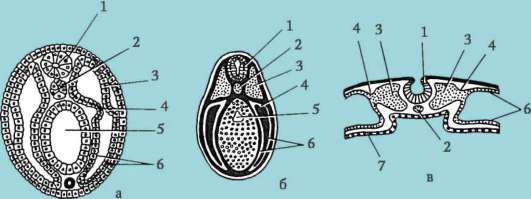
Ориз. 7.11.Неврули на различни хордови: а - ланцет; б - жаба; в - пиле; 1 - неврална тръба; 2 - акорд; 3 - сомит; 4 - сомитен крак; 5 - вторично черво; 6 - странична плоча; 7 - ендодерма
част е сегментирана и представена по двойки сомити.Полагането на сомитите върви от главата към края на опашката. Вентралната несегментирана част на мезодермата, която изглежда като тънък слой клетки, се нарича странична плоча.Сомитите са свързани към страничната пластина чрез междинната мезодерма под формата на сегментирани сомити крака.
Всички области на мезодермата постепенно се диференцират. В началото на образуването сомитите имат конфигурация, характерна за епител с кухина вътре. Под индукционния ефект, идващ от хордата и невралната тръба, вентромедиалните части на сомитите - склеротоми- се превръщат във вторичен мезенхим, изхвърлят се от сомита и обграждат нотохордата и вентралната част на невралната тръба. В крайна сметка от тях се образуват прешлени, ребра и лопатки.
Дорзолатералната част на сомитите се диференцира в две посоки. Образуват се участъци от сомити, обърнати към покривната ектодерма дерматоми,които дават началото на вътрешния слой на кожата – дермата. Образуват се клетки, разположени от вътрешната страна на дорзолатералната част на сомитите миотоми,от които ще се развият напречнонабраздените скелетни мускули на тялото и крайниците. От областта на краката на сомитите с рудименти нефротомИ гонотомобразуват се отделителни органи и полови жлези.
Несегментираните странични плочи се разделят на два листа, ограничаващи вторичната телесна кухина - общо взето. вътрешен лист,съседен на ендодермата се нарича висцерален. Той обгражда червата от всички страни и образува мезентериума, покриващ белодробната
паренхим и сърдечен мускул. Външен листстраничната плоча е в непосредствена близост до ектодермата и се нарича париетален.В бъдеще той образува външните листове на перитонеума, плеврата и перикарда.
7.4.3.4. Производни на зародишните слоеве
Ектодерма, мезодерма и ендодерма в хода на по-нататъшното развитие, взаимодействайки помежду си, участват в образуването на специфични органи (частна или локална органогенеза). Появата на рудимента на орган е свързана с локални промени в определена област на съответния зародишен слой. Така, от ектодермасе развива епидермиса на кожата и нейните производни (пера, коса, нокти, кожа и млечни жлези), компоненти на органите на зрението, слуха, обонянието, епитела на устната кухина и зъбния емайл. Най-важните ектодермални производни са невралната тръба, невралния гребен и всички неврални структури, образувани от тях.
Производни на ендодерматаса епител на стомаха и червата, чернодробни клетки, секретиращи клетки на панкреаса, чревни и стомашни жлези. Ендодермата на предната част на ембрионалните черва образува епитела на белите дробове и дихателните пътища, както и секретиращите клетки на предния и средния лоб на хипофизата, щитовидната и паращитовидните жлези.
мезодерма,в допълнение към вече описаните по-горе скелетни структури, скелетните мускули, дермата на кожата, органите на отделителната и репродуктивната системи, той образува сърдечно-съдовата система, лимфната система, плеврата, перитонеума и перикарда. от мезенхим,със смесен произход поради клетките на трите зародишни слоя се развиват всички видове съединителна тъкан, гладки мускули, кръв и лимфа (за повече подробности вижте параграф 8.2.5).
Зачатъкът на определен орган първоначално се формира от специфичен зародишен слой, но след това органът става по-сложен и в резултат на това два или три зародишни слоя участват в неговото формиране.
7.4.4. ПРЕДПАЗНИ ОРГАНИ НА ЕМБРИОНА НА ГРЪБНАЧНИ
временно,или временно,органи се образуват в ембриогенезата на редица представители на гръбначните за осигуряване на жизнените функции на ембриона като дишане, хранене, отделяне, движение и др. Недоразвитите органи на най-развиващото се животно все още не са в състояние да функционират по предназначение, въпреки че трябва
играят някаква роля в системата на развиващия се цял организъм. Веднага след като ембрионът достигне необходимата степен на зрялост, когато повечето органи са способни да изпълняват жизненоважни функции, временните органи се резорбират или изхвърлят.
Времето за образуване на временни органи зависи от това какви запаси от хранителни вещества са натрупани в яйцето и в какви условия на околната среда се развива ембрионът. При безопашатите земноводни, например, поради достатъчното количество жълтък в яйцето и факта, че развитието протича във вода, ембрионът осъществява газообмен и отделя продукти на дисимилация директно през мембраните на яйцето и достига стадия на попова лъжица. На този етап се формират временни органи на дишане (хриле), храносмилане и движение, адаптирани към водния начин на живот. Изброените ларвни органи позволяват на поповата лъжица да продължи развитието си. При достигане на състояние на морфологична и функционална зрялост на органите от възрастен тип, временните органи изчезват в процеса на метаморфоза.
Влечугите и птиците имат повече резерви от жълтък в яйцето, но ембрионът се развива не във вода, а на сушата. В тази връзка много рано възниква необходимостта от осигуряване на дишане и отделяне, както и предпазване от изсъхване. При тях още в ранната ембриогенеза, почти успоредно с неврулацията, започва образуването на провизорни органи, като напр. амнион, хорионИ жълтъчна торбичка.Малко по-късно се образува алантоис. При плацентарните бозайници същите тези временни органи се образуват дори по-рано, тъй като в яйцето има много малко жълтък. Развитието на такива животни се случва вътреутробно, образуването на временни органи в тях съвпада във времето с периода на гаструлация.
Наличието или отсъствието на амнион и други временни органи е в основата на разделянето на гръбначните животни на две групи: АмниотаИ Анамния(виж таблица 7.1). Еволюционно по-старите гръбначни животни, които се развиват изключително във водна среда и са представени от такива класове като циклостоми, риби и земноводни, не се нуждаят от допълнителна вода и други черупки на ембриона и съставляват групата anamnium. Групата на амниотите включва първични сухоземни гръбначни, т.е. тези, чието ембрионално развитие протича в земни условия. Това са трите класа: влечуги, птици и бозайници. Те принадлежат към висшите гръбначни животни, тъй като имат координирани и високоефективни системи от органи, които осигуряват съществуването им в най-голяма степен трудни условиякакви са условията на земята. Тези
класовете включват голям брой видове, вторично преминали във водната среда. Така висшите гръбначни животни успяха да овладеят всички местообитания. Такова съвършенство би било невъзможно, включително и без вътрешно оплождане и формирането на специални временни ембрионални органи, т.нар. ембрионални мембрани.Амниотичните мембрани включват амнион, хорион (сероза), жълтъчна торбичка и алантоис.
Има много общо в структурата и функциите на временните органи на различните амниоти. Описвайки най-общо временните органи на ембрионите на висшите гръбначни животни, трябва да се отбележи, че всички те се развиват от клетъчния материал на вече формираните зародишни слоеве. Има някои особености в развитието на ембрионалните мембрани на плацентарни бозайници, които ще бъдат разгледани по-долу.
Амнионе торбичка, съдържаща ембриона и пълна с амниотична течност. Амниотичната мембрана се образува от екстраембрионална ектодерма и соматоплевра. Ектодермалната му част е специализирана за секрецията и абсорбцията на околоплодната течност, която къпе ембриона. Амнионът играе основна роля в защитата на ембриона от изсушаване и механични повреди, създавайки за него най-благоприятната и естествена водна среда. Мезодермалната част на амниона води до гладкомускулни влакна. Контракциите на тези мускули карат амниона да пулсира и бавните осцилаторни движения, съобщавани на ембриона в този процес, очевидно помагат да се гарантира, че неговите растящи части не си пречат една на друга.
хорион (сероза)- най-външната зародишна мембрана, съседна на черупката (сероза) или майчините тъкани (хорион), възникваща, подобно на амниона, от ектодермата и соматоплеврата. Тази обвивка служи за обмена между ембриона и околната среда. При яйценосните видове основната функция на серозата е участието в дишането (обмен на газ); при бозайниците хорионът изпълнява много по-обширни функции, участвайки, освен в дишането, в храненето, екскрецията, филтрацията и синтеза на вещества, като хормони.
Жълтъчна торбичкаобразува се от извънембрионалната ендодерма и висцералната мезодерма и е директно свързана с чревната тръба на ембриона. При ембриони с голямо количество жълтък той участва в храненето. При птиците, например, васкулатурата се развива в splanchnopleuron на жълтъчната торбичка. Жълтъкът не преминава през жълтъчния канал, който свързва торбичката с червата. Първо се превежда на
разтворима форма под действието на храносмилателни ензими, произведени от ендодермалните клетки на стената на сака. След това навлиза в съдовете и се разпространява с кръв в тялото на ембриона.
Бозайниците нямат резерви за жълтък и запазването на жълтъчната торбичка може да бъде свързано с важни второстепенни функции. Мезодермата на жълтъчната торбичка произвежда кръвните клетки на ембриона. В допълнение, жълтъчната торбичка на бозайниците е пълна с течност с висока концентрация на аминокиселини и глюкоза, което показва възможността за протеинов метаболизъм в жълтъчната торбичка.
Съдбата на жълтъчната торбичка при различните животни е малко по-различна. При птиците до края на инкубационния период остатъците от жълтъчната торбичка вече са вътре в ембриона, след което бързо изчезват и напълно се разтварят до края на 6 дни след излюпването. При бозайниците жълтъчната торбичка е развита по различен начин. При хищниците той е сравнително голям, със силно развита мрежа от съдове, докато при приматите бързо се свива и изчезва безследно преди раждането.
алантоиссе развива малко по-късно от другите временни органи. Представлява торбовиден израстък на вентралната стена на задното черво. Следователно тя се образува от ендодермата отвътре и спланхноплеврата отвън. При влечуги и птици алантоисът бързо расте до хориона и изпълнява няколко функции. На първо място, това е резервоар за урея и пикочна киселина, които са крайните продукти на азотсъдържащия метаболизъм. органична материя. Алантоисът има добре развита съдова мрежа, поради което заедно с хориона участва в газообмена. При излюпването външната част на алантоиса се изхвърля, докато вътрешната част се запазва като пикочен мехур.
При много бозайници алантоисът също е добре развит и заедно с хориона образува хориоалантоисната плацента. Срок плацентаозначава тясно припокриване или сливане на зародишните мембрани с тъканите на родителския организъм. При примати и някои други бозайници ендодермалната част на алантоиса е рудиментарна, а мезодермалните клетки образуват плътна връв, простираща се от клоакалната област до хориона. Съдовете растат по мезодермата на алантоиса до хориона, чрез който плацентата изпълнява екскреторни, дихателни и хранителни функции.
По-достъпно и по-лесно е да се изследва образуването и структурата на ембрионалните мембрани, използвайки примера на пилешки ембрион. На етапа на неврулата три зародишни слоя преминават директно от ембриона към екстра-
дишаща част, без да се ограничава по никакъв начин. Докато ембрионът се оформя, около него се образуват няколко гънки, които сякаш подрязват ембриона, отделят го от жълтъка и установяват ясни граници между ембриона и екстраембрионалните области. Те се наричат гънки на багажника(фиг. 7.12).
Първо се образува гънка на главата. Тя изрязва главата отдолу. Задните краища на тази гънка преминават в страничните гънки на багажника, които ограничават тялото на ембриона отстрани. Опашната гънка ограничава задния край на ембриона. Постепенно стеснява крака, свързващ средното черво и жълтъчната торбичка, образувана
Ориз. 7.12.Образуване на гънки на багажника и ембрионални мембрани в пилешки ембрион: а - надлъжен разрез; b - напречно сечение; 1 - ектодерма; 2 - мезодерма; 3 - зачатъкът на мозъка; 4 - фарингеална мембрана; 5 - неврална тръба; 6 - акорд; 7 - клоакална мембрана; 8 - хорион; 9 - амнион; 10 - екзоцел; 11 - алантоис; 12 - зона на пъпа; 13 - зачатъкът на сърцето; 14 - ендодерма; 15 - маркер на червата; 16 - гънки на торса; 17 - жълтъчен сак
предни и задни черва. В същото време от ектодермата и прилежащата към нея соматоплевра първо се образува гънка на главата (фиг. 7.13), която като качулка расте върху ембриона отгоре. Краищата на гънката на главата се оформят отстрани амниотични гънки.Те растат върху ембриона един към друг и растат заедно, като веднага образуват стените на амниона, съседен на ембриона, и хориона, който лежи отвън.
По-късно се образува алантоис (фиг. 7.14). Общият изглед на пилешкия ембрион на 6-ия ден от инкубацията е показан на фиг. 7.15. При различни бозайници процесите на образуване на временни органи са повече или по-малко подобни на описаните по-горе при птиците. Характеристиките на тяхното развитие при примати, включително хора, са показани в следващата глава.
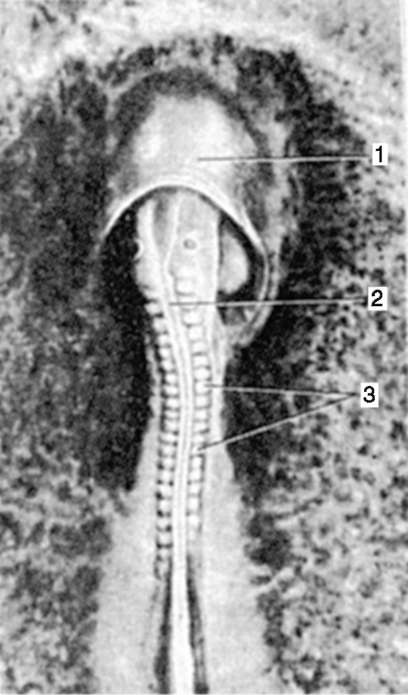
Ориз. 7.13.Пилешки ембрион за около 40 часа инкубация: 1 - гънка на главата на амниона; 2 - неврална тръба; 3 - сомити
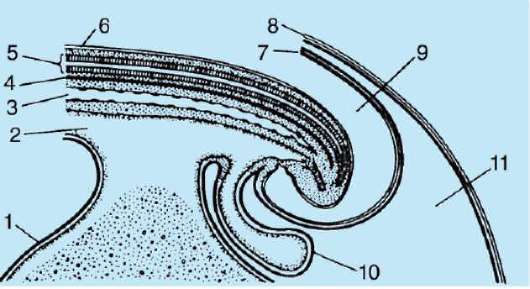
Ориз. 7.14.Образуването на алантоис в пилешки ембрион (надлъжен разрез на опашката): 1 - жълтъчен сак; 2 - средно черво; 3 - аорта; 4 - акорд; 3 - неврална тръба; 6 - ектодерма; 7 - амнион; 8 - хорион; 9 - амнионна кухина; 10 - алантоис; 11 - екзокоелом
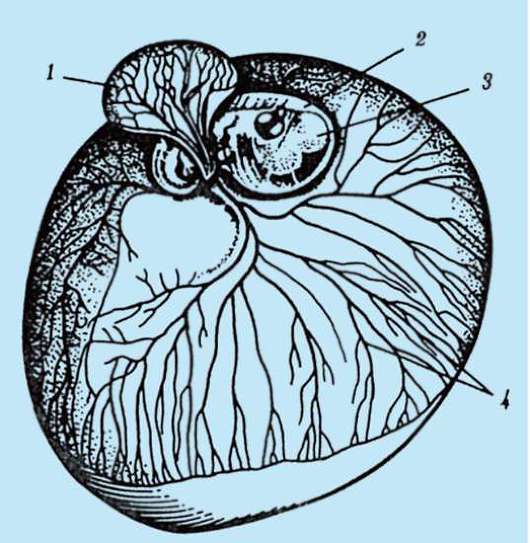
Ориз. 7.15.Пилешки ембрион на 6-ия ден от инкубацията (протеинът и хорионът са отстранени, алантоисът е преместен нагоре): 1 - алантоис; 2 - амнион; 3 - ембрион; 4 - съдове на жълтъчната торбичка
7.5. ембрионално развитие на бозайници
7.5.1. ПЕРИОДИЗАЦИЯ И РАННО ЕМБРИОННО РАЗВИТИЕ
Изследването на пренаталното и по-специално на ембрионалното (вътрематочно) развитие на човека е много важно, тъй като помага да се разбере по-добре връзката между органите, както и механизмите на вродените малформации. В ембрионалното развитие на различни видове бозайници има Общи черти, но има и разлики. Във всички плаценти, например, процесите на ранна ембриогенеза се различават значително от тези, описани по-рано при други гръбначни животни. В същото време сред плацентарните има специфични особености.
РазделянеЧовешката зигота се характеризира със следните характеристики. Равнината на първото деление минава през полюсите на яйцето, т.е., както при другите гръбначни животни, това е меридиан. В този случай един от получените бластомери е по-голям от другия, което показва нередностсмачкване. Първите два бластомера влизат в следващото деление асинхронно.Браздата минава по меридиана и е перпендикулярна на първата бразда. По този начин има етап от три бластомера. По време на разделянето на по-малкия бластомер, образуваната двойка по-малки бластомери се завърта на 90 °, така че равнината на браздата на делене е перпендикулярна на първите два
Ориз. 7.16.Ранни етапи на разцепване на заешката зигота: I - равнина на първата бразда на разцепване; IIa - равнината на втората бразда, смачкваща един от първите два бластомера; IIb - равнина на втората бразда на разцепване на втория от първите два бластомера
бразди. Подобно разположение на бластомерите в 4-клетъчния стадий е описано при мишки, зайци, норки и маймуни (фиг. 7.16). Поради асинхронното разцепване може да има етапи с нечетен брой бластомери - 3, 5, 7, 9, които поради липса на етап на растеж се различават по своя размер. Така че смачкването на човека пълен асинхронен неравномерен.
В резултат на раздробяването се образува натрупване на клетки - морула.Повърхностно разположените бластомери образуват клетъчен слой, а бластомерите, разположени вътре в морулата, са групирани в централен клетъчен възел. Скоро вътре в морулата се появява течност, образува се кухина (бластоцел) и ембрионът се превръща в бластоциста.Образуването на морула става на етап 16 бластомера, а кавитацията (образуване на кухина) започва от етапа на 32 бластомера.
IN бластоцистаразличават външния слой на клетките (трофобласт)и вътрешната клетъчна маса (зародишно възелче или ембриобласт).Вътрешната клетъчна маса се отблъсква от течност към един от полюсите на бластоциста. По-късно от трофобластще се развие външната зародишна мембрана - хорион, и от ембриобласт- самият ембрион и някои извънембрионални органи. Доказано е, че истинският ембрион се развива от много малък брой клетки на зародишния възел.
Етапът на смачкване се извършва под лъскавата черупка. Фигура 7.17 показва ранните етапи на човешката ембриогенеза, показвайки къде се намира ембрионът в тялото на майката. Фрагментацията на човешката зигота и появата на бластоциста са показани схематично на фиг. 7.18 и 7.19.
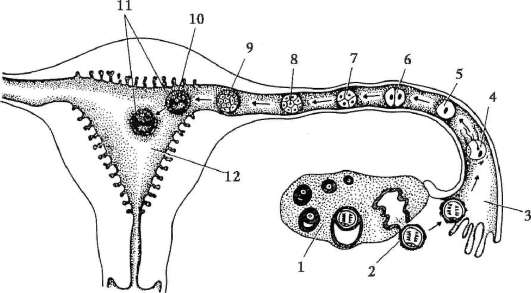
Ориз. 7.17.Овулацията, оплождането и човешкият ембрион през 1-вата седмица от развитието: 1 - яйчник; 2 - овоцит II ред (овулация); 3 - яйцепровод; 4 - оплождане; 5 - зигота; 6 - ембрион на етапа на два бластомера; 7 - ембрион на етап от четири бластомера; 8 - ембрион на етап от осем бластомера; 9 - морула; 10, 11 - бластоцист; 12 - задната стена на матката
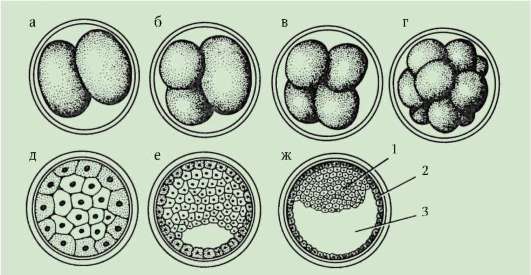
Ориз. 7.18.Разцепване на човешката зигота: а - два бластомера; b - три бластомера; c - четири бластомера; g - морула; e - секция на морулата; f, g - разрез на ранен и късен бластоцист: 1 - ембриобласт; 2 - трофобласт; 3 - бластоцел
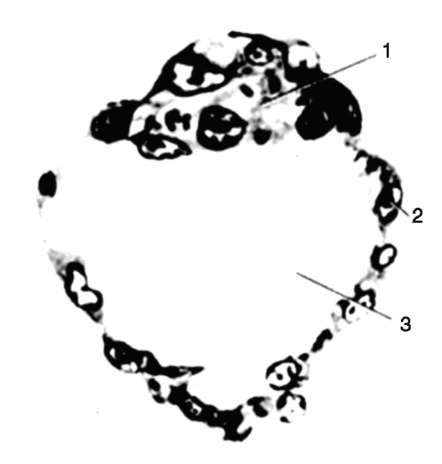
Ориз. 7.19.Бластоцист на човешки ембрион (разрез): 1 - ембриобласт; 2 - трофобласт; 3 - бластоцел
Приблизително на 6-7-ия ден след оплождането ембрионът, който плава свободно в маточната кухина в продължение на 2-3 дни, е готов за имплантиране, т.е. до потапяне в лигавицата му. Под действието на ензимите, отделяни от трофобласта, zona pellucida се разрушава частично и ембрионът излиза от нея („люкове“). С участието на интегринови протеини, синтезирани както от маточния епител, така и от клетките на трофобласта, бластулата е прикрепена към стената на матката с ембрионалния си полюс. Влизайки в контакт с тъканите на майката, клетките на трофобласта се размножават бързо и, освобождавайки протеолитични ензими, ги унищожават (тези тъкани). Това предизвиква реакция на стената на матката, придружена от активно образуване на съдове на мястото на имплантиране. Нарушаването на синтеза на интегрини и ензими, необходими за имплантиране, прави този процес невъзможен, което от своя страна води до хипоксия и смърт на ембриона. По време на имплантирането трофобластът се диференцира на два слоя: вътрешният слой, т.нар цитотрофобластом,защото спестява клетъчна структура, и външната, т.нар синцитио-трофобластом,тъй като е синциций. Фигура 7.20 показва човешки ембрион в процес на имплантиране.
гаструлацияпри бозайниците тя е тясно свързана с други ембрионални трансформации. Едновременно с разделянето на трофобласта на два слоя, зародишният възел се сплесква и се превръща в двуслоен зародишно щитче. Долният слой на щита - хипобласт,или първична ендодерма, според повечето автори, се образува чрез разслояване на вътрешната клетъчна маса, приблизително както се случва в ембрионалния диск на птиците. Първичен
Ориз. 7.20.Последователни етапи на имплантиране и развитие на човешкия ембрион в края на 1-ва и 2-ра седмица: а - бластоциста; b - бластоциста в самото начало на имплантирането (7-ми ден от развитието); c - частично имплантиран бластоцист (8-ми ден от развитието)
Ориз. 7.20. Край: d - ембрион на 9-10-ия ден от развитието; e - ембрион на 13-ия ден от развитието: 1 - ембриобласт, 2 - бластоцел, 3 - трофобласт, 4 - амнионна кухина, 5 - хипобласт, 6 - синцитиотрофобласт, 7 - цитотрофобласт, 8 - епибласт, 9 - амнион, 10 - лакуна трофобласт, 11 - маточен епител, 12 - стебло на тялото, 13 - алантоисен бъбрек, 14 - жълтъчен белег, 15 - екстрадишащ целом, 16 - хорионни въси, 17 - първичен жълтъчен сак, 18 - вторичен жълтъчен сак
naya ендодерма се изразходва напълно за образуването на екстраембрионална ендодерма. Покривайки кухината на трофобласта, той образува първичната жълтъчна торбичка на бозайниците.
Горен клетъчен слой епибласт- източникът на бъдещата ектодерма, мезодерма и вторична ендодерма. На 3-та седмица в епибласта се образува първична линия,чието развитие е придружено от почти същите движения на клетъчните маси, както при образуването на първичната ивица на птиците (фиг. 7.21). В горния край на първичната ивица, Възелът на ХенсънИ първичен отвор,хомоложни на дорзалната бластопорна устна на други гръбначни. Клетките, които се движат в областта на първичната ямка, се насочват под епибласта към прехордалната плоча.
прехордална плочаразположен в края на главата на ембриона и маркира мястото на бъдещата орофарингеална мембрана. Клетките, движещи се по централната ос, образуват зачатъка на нотохордата и мезодермата и се изграждат хордомезодермен процес.Възелът на Хенсен постепенно се измества към каудалния край на ембриона, първичната ивица се скъсява и зачатъкът на хордата се удължава. Отстрани на хордомезо-дермалния процес се образуват мезодермални пластинки, които се разширяват в двете посоки. Фигура 7.22 представя обобщена диаграма на някои от процесите на ранното ембрионално развитие.
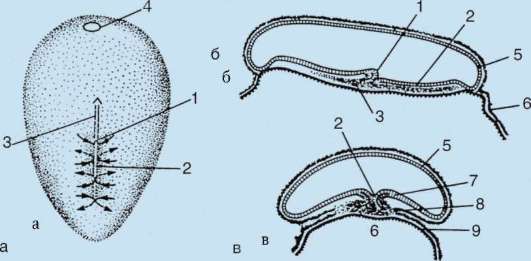
Ориз. 7.21.Развитие на човешкия ембрион на първичния етап (15-17 дни): а - изглед на ембриона отгоре (амнионът е отстранен); b - надлъжен разрез; c - напречен разрез през първичната лента: 1 - възел на Хенсен; 2 - първична лента; 3 - акорд; 4 - прехордална плоча; 5 - амнион; 6 - жълтъчна торбичка; 7 - ектодерма; 8 - мезодерма; 9 - ендодерма
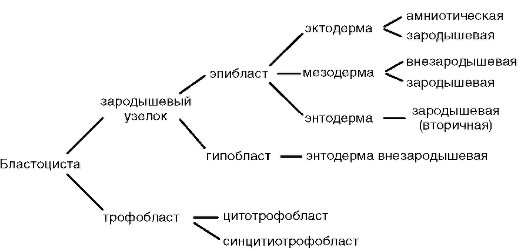
Ориз. 7.22.Диференциация на зародишните слоеве на бозайниците
До края на 3-та седмица, в ектодермата на ембриона над зачатъка на хордата, неврална пластина.Състои се от високи цилиндрични клетки. В центъра на невралната пластина се образува деформация във формата нервен канал,а отстрани се издигат нервни гънки.Това е началото на неврулацията. В средната част на ембриона настъпва затваряне на невралните гънки - а неврална тръба.След това затварянето се разпространява в посоките на главата и опашката. Невралната тръба и съседните части на ектодермата, от които впоследствие се развива нервен гребен,напълно потопени и отделени от ектодермата, растяща заедно над тях (виж фиг. 7.10). Ивицата от клетки, разположена под невралната тръба, се превръща в хорда. Сегменти от дорзалната мезодерма - сомити - се появяват отстрани на хордата и невралната тръба в средната част на ембриона. До края на 4-та седмица те се разпространяват в края на главата и опашката, броят им достига около 40 двойки.
Началото на образуването на вторичното черво, залата на сърцето и васкулатурата на жълтъчната торбичка принадлежи към същото време. Фигура 7.23 показва съотношението на размера на ембриона и екстраембрионалните органи на 21-ия ден от развитието. По-подробно отделянето на тялото на ембриона от ембрионалните мембрани и полагането на органи може да се види на фиг. 7.24, който показва не само общия изглед на ембриона, но и плановете на разреза. Обръща се внимание на бързото (в рамките на 7 дни от 4-та седмица) образуване на ембриона под формата на удължено и извито тяло, повдигнато и отрязано от гънките на багажника от жълтъчната торбичка. Отзад
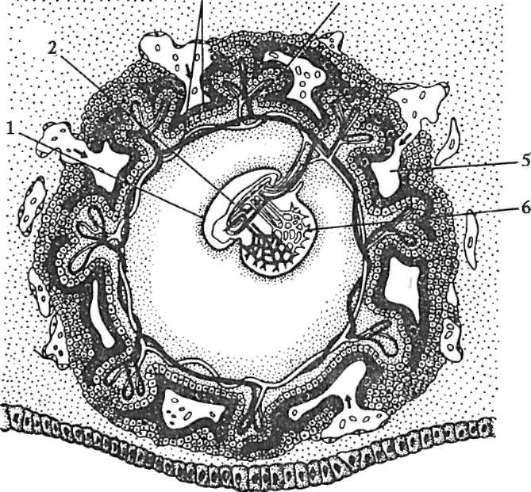
Ориз. 7.23.Ембрионът и екстраембрионалните човешки органи на 21-ия ден от развитието: 1 - амнион; 2 - ембрион; 3 - хорион; 4 - третичен вил; 5 - майчина кръв; 6 - жълтъчна торбичка
по това време се полагат всички сомити, четири чифта хрилни дъги, сърдечната тръба, бъбреците на крайниците, средното черво, както и "джобовете" на предното и задното черво.
През следващите четири седмици от ембрионалното развитие се залагат всички основни органи. Нарушаването на процеса на развитие през този период води до най-тежките и множество вродени малформации.
Както беше отбелязано по-горе, развитието на екстраембрионални временни органи при бозайници и хора има специфични характеристики. Тези органи се образуват много рано, едновременно с гаструлацията и малко по-различно от другите амниоти. Началото на развитието на хориона и амниона пада на 7-8-ия ден, т.е. съвпада с началото на имплантирането. хорионвъзниква от трофобласта, който вече е разделен на цитотрофобласт и синцитиотрофобласт. Последният, под въздействието на контакт с лигавицата на матката, расте и я унищожава. До края на 2-та седмица,
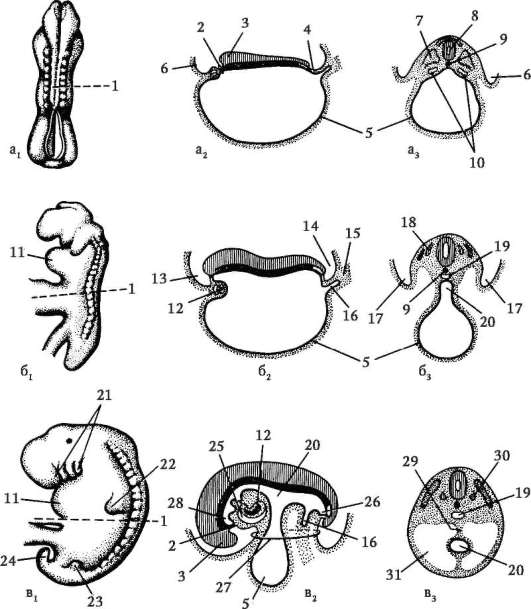
Ориз. 7.24.Развитието на човешкия ембрион на 4-та седмица: a 1 b 1 в 1 - общ изглед; a 2 b 2 c 2 - надлъжен разрез; a 3 b 3 c 3 - напречно сечение; a 1 a 2 a 3 - 22 дни; b 1 b 2 b 3 - 24 дни; в 1 в 2 в 3 - 28 дни: 1 - нивото на напречното сечение; 2 - орофарингеална мембрана; 3 - мозък; 4 - клоакална мембрана; 5 - жълтъчна торбичка; 6 - амнион; 7 - сомити; 8 - неврална тръба; 9 - акорд; 10 - сдвоени маркери на коремната аорта; 11 - сърдечен перваз; 12 - сърце; 13 - гънка на багажника на главата; 14 - гънка на опашката на багажника; 15 - крак на тялото; 16 - алантоис; 17 - странични гънки на багажника; 18 - неврален гребен; 19 - гръбначна аорта; 20 - средно черво; 21 - хрилни дъги; 22 - бъбрек на предния крайник; 23 - бъбрек на задния крайник; 24 - опашка; 25 - перикард; 26 - джоб на задното черво; 27 - пъпна връв; 28 - джоб на предното черво; 29 - дорзален мезентериум; 30 - нервен възел на задния корен; 31 - интраембрионално цяло
първичните власинки на хориона се образуват под формата на натрупване на цитотрофобластни епителни клетки. В началото на 3-та седмица мезодермалния мезенхим прораства в тях и се появяват вторични вили, а когато до края на 3-та седмица се появят кръвоносни съдове вътре в сърцевината на съединителната тъкан, те се наричат третични вили. Областта, където тъканите на хориона и лигавицата на матката са тясно съседни, се нарича плацента.
При хората, както и при другите примати, съдовете на майчината част на плацентата губят своята непрекъснатост и хорионните въси всъщност се къпят в кръвта и лимфата на майчиния организъм. Тази плацента се нарича хемохориален.С напредването на бременността вилите се увеличават по размер, разклоняват се, но кръвта на плода от самото начало до края е изолирана от кръвта на майката чрез плацентарната бариера.
Плацентарна бариерасе състои от трофобласт, съединителна тъкан и съдов ендотел на плода. Тази бариера е пропусклива за вода, електролити, хранителни вещества и продукти на дисимилация, както и за фетални еритроцитни антигени и майчини антитела, токсични вещества и хормони. Клетките на плацентата произвеждат четири хормона, включително човешки хорионгонадотропин, който се намира в урината на бременна жена от 2-3-та седмица на бременността.
Амнионвъзниква чрез дивергенция на епибластни клетки от вътрешната клетъчна маса. Човешкият амнион се нарича шизамнион(виж фиг. 7.20) за разлика от плеврамнионптици и някои бозайници. Амниотичната кухина за известно време е ограничена от епибластни клетки и отчасти от място на трофобласт. След това страничните стени на епибласта образуват гънки, насочени нагоре, които впоследствие се сливат. Кухината е изцяло облицована с епибластни (ектодермални) клетки. Отвън амниотичната ектодерма е заобиколена от екстраембрионални мезодермални клетки. Амниотичната течност съдържа различни вещества, като протеини, въглехидрати, соли, микроелементи, хормони и др. Съставът й се променя по време на развитието, освен това се актуализира постоянно. В процеса на развитие на ембриона той играе важна роля при формирането на носната и устната кухина, храносмилателните и дихателните органи.
Жълтъчна торбичкасе появява, когато тънък слой от хипобласта се отдели от вътрешната клетъчна маса и неговите екстраембрионални ендодермални клетки, движейки се, покриват повърхността на тро-
фобласт. Полученият първичен жълтъчен сак се свива на 12-13-ия ден и се превръща във вторичен жълтъчен сак, свързан с ембриона чрез жълтъчния канал. Ендодермалните клетки са покрити отвън от екстраембрионалната мезодерма. На 3-та седмица от развитието първичните зародишни клетки, кръвните клетки и кръвоносните съдове започват да се откриват в стената на жълтъчната торбичка. До 7-8-та седмица от развитието този жълтъчен сак е основният хематопоетичен орган на ембриона.
алантоисвъзниква в човешкия ембрион, както и при други амниоти, под формата на джоб във вентралната стена на задното черво, но неговата ендодермална кухина остава рудиментарна структура. Въпреки това в стените му се развива изобилна мрежа от съдове, свързващи се с главните кръвоносни съдове на ембриона. Мезодермата на алантоиса се свързва с мезодермата на хориона, вкарвайки в нея кръвоносни съдове, които осигуряват храненето на развиващия се ембрион до около втория месец от ембриогенезата. По този начин се получава васкуларизация на хориоалантоичната плацента.
Някои етапи и срокове на развитие на органите в човешките ембриони са представени в таблица. 7.4.
Таблица 7.4.Основни периоди и събития в ранния онтогенезис на човека
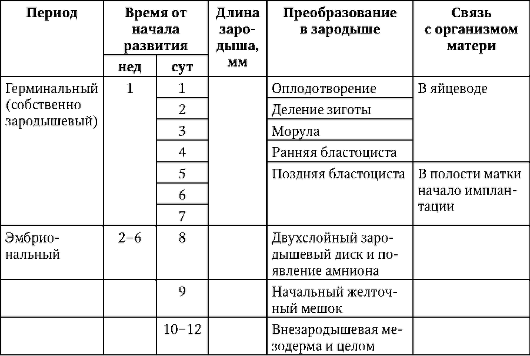
Продължение на таблицата. 7.4
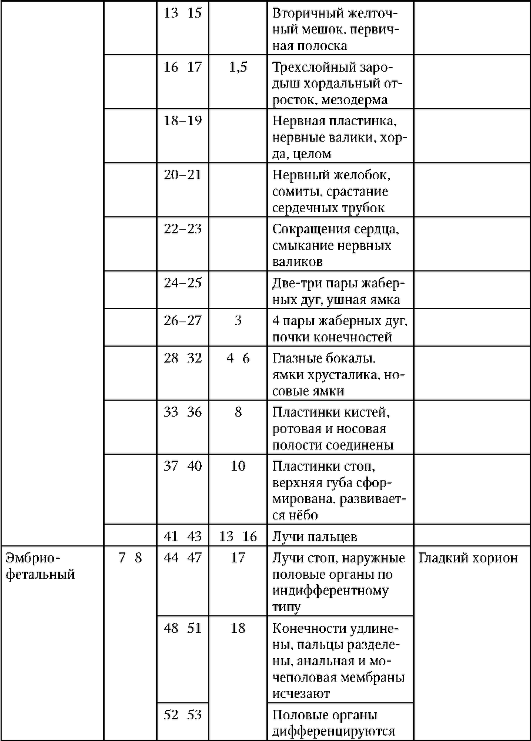
Краят на масата. 7.4
7.5.2. ПРИМЕРИ ЗА ЧОВЕШКА ОРГАНОГЕНЕЗА, ОТРАЗЯВАЩА ЕВОЛЮЦИЯТА НА ВИДА
Този раздел няма за цел да опише развитието на всички човешки органи. Ще бъдат разгледани само някои морфогенетични процеси, илюстриращи следните общи биологични точки:
Значението на междуклетъчните, тъканните и междуорганните взаимодействия в морфогенезата;
Отражение в човешката морфогенеза на еволюционно по-древни особености на организацията на гръбначните животни;
Характеристики на органогенезата, позволяващи да се оцени етапът и механизмите на нарушения в развитието при някои вродени малформации.
Сегментна структура гръбначен стълботразява неговия произход от сомитите на ембриона. В допълнение, формирането на прешлените е тясно свързано с хордата и гръбначния мозък. Телата на прешлените се образуват от клъстери от мезенхимни клетки, мигриращи към нотохордата и около нея, които произхождат от склеротоми. Мезенхимните клетки на две съседни двойки сомити участват в образуването на телата на прешлените и следователно телата на прешлените са разположени не на нивото на сомитите, а между тях. В същото време мускулите, развиващи се от миотомите на соми-
Ориз. 7.25.Развитието на дорзалните дъги на прешлените при пилешки ембрион: а - нормално развитие; b - след отстраняване на гръбначните ганглии: 1 - гръбначна дъга; 2 - гръбначно тяло; 3 - гръбначен възел; 4 - гръбначен мозък; 5 - акорд; 6 - несегментирани гръбни дъги на прешлените
comov, сякаш хвърлен върху междупрешленните стави и осигурява движението на прешлените.
Гръбначните дъги се образуват от натрупвания на мезенхимни склеротомични клетки, разпространяващи се в дорзална (дорзална) посока, докато напречните процеси и ребра се образуват от натрупвания на склеротомни клетки, мигриращи настрани. Гръбначният мозък и гръбначните ганглии на съответните етапи на вътрематочно развитие участват в морфогенезата на дорзалните дъги на прешлените. Така че, ако се отстранят гръбначните ганглии, тогава се образува хрущялът на дорзалните дъги, но изглежда като несегментиран прът (фиг. 7.25).
Хистологичната структура на прешлените постепенно се променя. През 4-5-та седмица се образуват мезенхимни прешлени, на 6-та седмица се появяват огнища на хондрогенеза, а след това на 8-та седмица започва осификация, която продължава до почти 25 години. Участъци от хордата, разположени вътре в телата на прешлените, постепенно изчезват. В междупрешленните дискове нотохордата е запазена като подобна на слуз структура, известна като пулпозно ядро.
Формиране средно ухосвързани с онтогенетични трансформации на първото и второто висцерални (хрилни) дъги.Фигура 7.26 схематично показва областите на главата и шията на 4-та и 24-та (края на първия-втори триместър на пренаталното развитие) седмици от човешкото развитие. На 4-та седмица в областта на главата и шията на ембриона отдясно и отляво се полагат висцерални (хрилни) дъги. Отвътре, от страната на фаринкса, между тях се образуват фарингеални джобове(фиг. 7.27). Това са издатини (или издатини) на ендодермата,
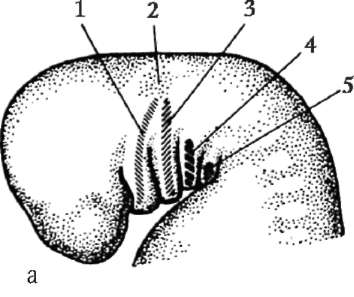
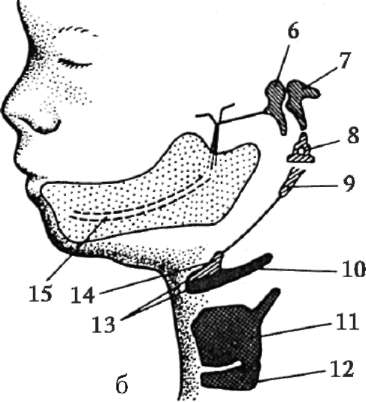
Ориз. 7.26.Глава и шия на човешки ембрион, страничен изглед: а - на 4-та седмица; b - на 24-та седмица: 1 - първата дъга; 2 - местоположение на развиващото се вътрешно ухо; 3 - втора дъга; 4 - трета дъга; 5 - четвърти ливади; 6 - чук. 7 - наковалня; 8 - стреме; 9 - стилоиден процес; 10 - голям рог на хиоидната кост; 11 - тироиден хрущял; 12 - крикоиден хрущял; 13 - тялото на хиоидната кост; 14 - малък рог; 15 - долна челюст; анлагите на висцералните дъги (а) и органите, развиващи се от тях (б), са обозначени със същото засенчване
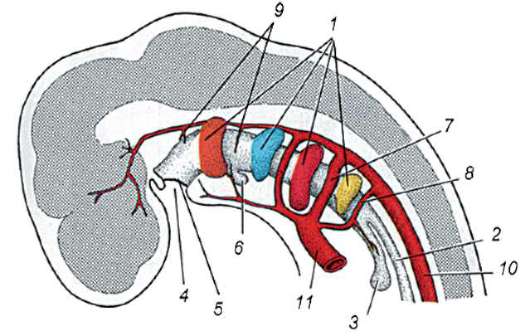
Ориз. 7.27.Област на хрилете на 5-седмичен човешки ембрион (отстранени покривна ектодерма и мезенхим). 1 - фарингеални джобове; 2 - хранопровод; 3 - трахея и белодробен бъбрек; 4 - стомодеум; 5 - стомодална (орална) плоча; 6 - рудимент на щитовидната жлеза; 7 - 4-та аортна дъга; 8 - 6-та аортна дъга; 9 - намаляване на 1-ва и 2-ра аортни дъги; 10 - дорзална низходяща аорта; 11 - коремна възходяща аорта (артериална торбичка)
расте настрани. Отвън, според фарингеалните израстъци, ектодермален хрилни джобове.
Ендодермата и ектодермата на първите фарингеални и хрилни джобове влизат в контакт на 4-та седмица от развитието (фиг. 7.28, а). Контактът не трае дълго, слепият край на фарингеалния джоб се отдалечава от повърхността и е заобиколен от мезенхим (фиг. 7.28, b). От дисталната част на фарингеалния джоб започва да се развива кухината на средното ухо (тимпаничната кухина), а от проксималната част се образува слуховата (евстахиевата) тръба. От мезенхима допълнително се диференцират анлагите на слуховите осикули, които служат за провеждане на звук (фиг. 7.28, c). Малеусът и наковалнята произлизат от клетъчния материал на първата висцерална дъга, стремето - от клетъчния материал на дорзалната част на втората висцерална дъга. От клетъчния материал на областта на първата и втората висцерални дъги се формират мускулите и нервите, свързани със звукопроводящия осикуларен апарат чрез осикулите на средното ухо.
В края на вътреутробния живот и в рамките на няколко месеца след раждането ембрионалната съединителна тъкан в тъпанчевата кухина се резорбира. Едва след това слуховите осикули придобиват подвижност и вибрациите на тимпаничната мембрана могат свободно да се предават на мембраната на овалния прозорец на вътрешното ухо.
При формирането на структурите областта на лицетоИ устната кухинаучастват екто- и мезодермни клетки. Повечето отструктура
Ориз. 7.28.Развитие на средното ухо, фронтална част: а - на 4-та седмица; b - на 5-та седмица; c - по-късни етапи; 1 - първа хрилна плоча; 2 - първи хрилен джоб; 3 - първата висцерална (хрилна) дъга; 4 - слухов везикул; 5 - първи фарингеален джоб; 6 - производни на първата висцерална дъга; 7 - производно на втората висцерална дъга; 8 - кухина на тръбата; 9 - запушалка на външния слухов канал. 10 - първичен външен слухов канал; 11 - чук; 12 - наковалня; 13 - стреме; 14 - тимпанична кухина; 15 - слухова (евстахиева) тръба
обиколката на лицевата област се развива от маси от мезенхимни клетки от ектодермален произход, мигриращи от невралния гребен (4-ти зародишен слой). От този мезенхим се развиват съединителна, хрущялна и костна тъкан и тъкан на зъбната пулпа. Набраздените мускули на мускулите на лицето и фаринкса се образуват от мезенхима с мезодермален произход. Тук е важно да разберем как е протекъл процесът на „разширяване и промяна на функциите“ в антропогенезата.
Устната кухина се образува от ектодермална инвагинация стомо-деум,в съседство със сляпо завършващото предстомашие. Те са разделени от тънки стомодална пластина,състоящ се от ектодермален (от страната на стомодеума) и ендодермален (от страната на червата) листове (виж Фиг. 7-26). Тази пластина се счупва в края на 4-та седмица от развитието и устната кухина е свързана с първоначално сляпо затворената чревна тръба.
Паротидните слюнчени жлези се развиват от ектодермата на стомодеума, а сублингвалните и субмандибуларните слюнчени жлези се развиват от ендодермата на вентралната част на фаринкса.
При плод на 5 седмици отваряне на устатаограничена от следните структури (фиг. 7.29, а): в средната линия над устната кухина има заоблена надвиснала област - челен (челен) перваз;отстрани има подковообразни възвишения, заобикалящи обонятелните ями. Средните процеси на тези възвишения
Ориз. 7.29.Последователни етапи на формиране на лицето (изглед отпред): а - на 5-та седмица; b - на 10-та седмица. 1 - стомодеум; 2 - челен (челен) перваз; 3 - медиален назален процес; 4 - страничен назален процес; 5 - обонятелна ямка; 6 - максиларен процес; 7 - полагане на долночелюстната дъга; 8 - филтрум; 9 - око
Наречен медиален назален,и страничните страничен
назални процеси.От горните странични ъгли на устната кухина към средната линия растат максиларни процеси.
Ограничаване на устната кухина отдолу Долна челюстобразуван от сдвоени отметки на долночелюстната дъга.Първоначално те се появяват от всяка страна на средната линия, след това се увеличават един към друг и растат заедно.
През 6-та седмица има бързо развитие горна челюст.Максиларните процеси нарастват към средната линия. В същото време носните процеси се увеличават, избутвайки долната част на челната издатина, разположена между тях. Средните назални процеси растат особено интензивно, които се изместват към средната линия и се сливат един с друг, след което се сливат с максиларните процеси, като по този начин образуват пълна челюстна арка на 10-та седмица от развитието. Образува се кожната ектодерма над слети медиални процеси филтрум- надлъжен жлеб над горната устна под носа (фиг. 7.29, b).
До края на втория месец от развитието, когато се формира горната челюст, тя започва да се развива плътно небе.В нейното формиране, както и във формирането на горната челюстна дъга, участват средни носни и максиларни процеси(Фиг. 7.30, а). От средната област на горната челюст, възникнала в резултат на сливането на средните носни процеси, се образува малка триъгълна част - първично небце.Главна част - вторично небце- се развива от странични палатинални израстъци (процеси),които се появяват на горната челюст (фиг. 7.30, b). Отначало те са насочени надолу, а след това краищата на палатинните процеси се издигат нагоре и към средната линия. Те контактуват и се сливат, за да образуват шева на небцето. Образуваната основна част на небцето отделя най-горната част на оригиналната стомодална кухина. Вторичното небце расте заедно с първичното, а носната преграда расте до тяхната черепна повърхност (фиг. 7.30, c). По този начин, едновременно с отделянето на носната кухина от устната кухина, тя се разделя на дясна и лява половина.
В началния период на неговото формиране (на 5-та седмица от ембриогенезата) езикпредставлява торба, образувана от лигавицата, пълна с нарастваща мускулна маса. Смята се, че лигавицата на езика възниква от стомодеум ектодерма,и мускулите идват от тилни миотоми.
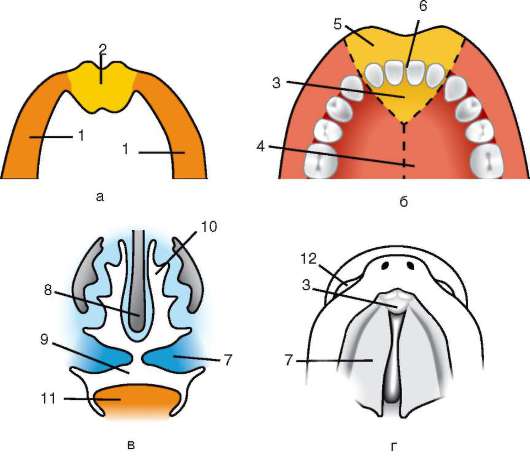
Ориз. 7.30 ч.Образуване на твърдото небце: а - схематично изображение на горната челюст (изглед от страната на устната кухина); b - 10-та седмица на развитие (изглед от устната кухина); c - 8-ма седмица на развитие (челен участък); d - 8-ма седмица на развитие (изглед от устната кухина): 1 - максиларен процес; 2 - слети медиални назални процеси; 3 - първично небе; 4 - вторично небце (странични палатинови израстъци); 5 - филтрум на устните; 6 - горна челюст с резци; 7 - страничен палатинов израстък; 8 - носна преграда; 9 - устна кухина; 10 - носна кухина; 11 - език; 12 - око
Няколко структури на пода на устната кухина участват в образуването на лигавицата на езика. Тялото на езика се образува от сливането на сдвоени странични удебеления - странични езични туберкулии разположено между тях малко средно възвишение, т.нар tuberculum impar(фиг. 7.31). Смята се, че тялото на езика е производно на клетъчния материал на областта на първата висцерална дъга. Участва в образуването на корена на езика корула(скоба)- средно възвишение, разположено отзад tuberculum imparи включете
Ориз. 7.31.Езиково развитие: а - 5-седмичен ембрион; б - 5-месечен плод; 1 - страничен езиков туберкул; 2- tuberculum impar 3 - копула (скоба); 4 - foramen coecum; 5 - крайна бразда; 6 - тялото на езика; 7 - коренът на езика; 8 - полагане на епиглотичния хрущял; 9 - глотис; 10 - аритеноидна туберкулоза; 11 - епиглотис, палатинална сливица. I-IV - номера на висцералните дъги
клетъчния материал на областите на втората, третата и четвъртата висцерална дъга. След сливането на всички зачатъци и окончателното формиране на езика, тялото му се отделя от корена терминална бразда(sulcus terminalis),в центъра на който е сляпа дупка(форамен коекум)- следа от инвагинация на дъното на фаринкса, от която се появява щитовидната жлеза (виж фиг. 7.27). Именно тя се счита за границата между тялото и корена на езика.
По-късно езикът почти напълно се отделя от дъното на устната кухина, като връзката между тях се запазва под формата на гънка на лигавицата - френулума на езика.
Интересен пример за органогенеза е образуването зъби,в който участва покривната ектодерма - епител на стомодеума (емайл) и мезенхимни клетки, произхождащи от невралния гребен (дентин, цимент и пулпа) (фиг. 7.32). В хода на развитието се отбелязват индукционни взаимодействия между мезенхимните и епителните структури.
По време на живота се развиват две поколения зъби. Смяната на зъбите се случва в детството, първите зъби се наричат млечни продукти.Има 20 от тях: 10 в горната и 10 в долната челюст. Млечните зъби никнат напълно на около 2-годишна възраст. Този набор от зъби служи на детето през следващите 4 години, след което постепенно млечните зъби
Ориз. 7.32.Човешки постоянни зъби: 1 - резци; 2 - зъби; 3 - малки молари; 4 - големи молари
но падат и се сменят постоянни зъби(второ поколение зъби), които функционират в човека до края на живота. Периодът на смяна на зъбите продължава приблизително от 6 до 12 години. 32 постоянни зъба: 16 горни и 16 долни (добавени са големи молари) (виж фиг. 7.32). По форма те са подобни на млечните, но са по-големи по размер.
Bookmark и образование зъбни микробиМлечните зъби започват от 6-7 седмици от вътрематочното развитие. Епителът на устната кухина е потопен в подлежащия мезенхим под формата на плътна нишка, т.нар. зъбна пластина(фиг. 7.33). На зъбната пластина се появяват малки епителни издатини - зъбни пъпки,от които ще се развият млечните зъби. С нарастването на зъбната пластинка всеки зъбен зародиш увеличава размера си, навлиза по-дълбоко в мезенхима и приема формата на обърната купа. Тази структура формира емайл орган,и се образува подлежащият мезенхим, който изпълва кухината на чашата зъбна папила.При по-нататъшно развитие последният образува пулпата на зъба. На 3-ия месец от вътрематочното развитие органът на емайла се увеличава по размер, променя формата си и постепенно се отделя от зъбната пластина. Формата на бъдещия зъб се определя от мезенхимния компонент. При комбиниране инвитромезенхим на полагането на моларния зъб (молар) с епителния компонент на полагането на резеца, моларът се развива. Обратно, в резултат на комбинацията от ектодермата на молара и мезенхима на резеца се развива резец.
По време на хистогенезата на зъбните тъкани, която започва в края на 4-ия месец, дентин, емайл и пулпа(Фиг. 7.33, d). Емайл
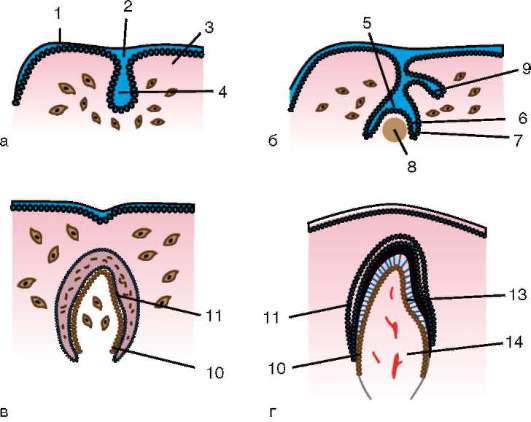
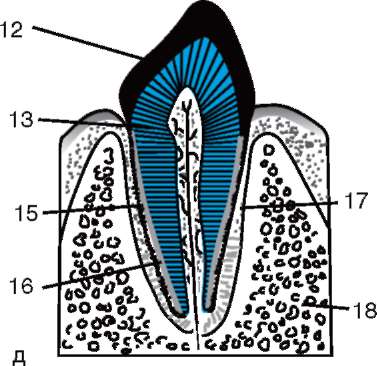
Ориз. 7.33.Развитие на зъбите: а - 8-седмичен ембрион; b - 10-седмичен плод; в - 3-месечен плод; g - 6-месечен плод; d - след изригване; 1 - епител на устната кухина; 2 - зъбна пластина; 3 - мезенхим; 4 - зародиш на млечен зъб; 5 - емайлов орган; 6 - вътрешният слой на емайловия орган; 7 - външен слой на емайловия орган; 8 - зъбна папила; 9 - зародиш на постоянен зъб; 10 - одонтобласти; 11 - амелобласти; 12 - емайл; 13 - дентин; 14 - зъбна пулпа; 15 - цементобласти; 16 - цимент; 17 - пародонта; 18 - алвеоларна кост
произвеждат клетки от вътрешния слой на емайловия орган - бластиране на емайла.Техните процеси отделят органичната основа на емайла - емайлови призми, които след това се калцират. Източник на развитие дентин- одонтобласти (дентинобласти),повърхностни пулпни клетки. Върхът на дентинобластите също има процеси, които секретират органични вещества с фибриларна структура, т.нар. предентини образуване на матрицата на дентина. От края на 5-ия месец в предентина се отлагат калциеви и фосфорни соли и се образува окончателният дентин. Образуването на дентин и емайл се различава от развитието на костта по това, че клетките не са заградени междуклетъчно вещество, но се отдалечават: амелобластите - навън, одонтобластите - навътре. В допълнение, съдържанието на органични вещества в тези тъкани е различно: костта съдържа приблизително 45% органична материя, дентинът - 28%, а емайлът - по-малко от 5%.
На външната повърхност на дентина от мезенхима около зъба (торбичка за зъби)се появяват малко преди никненето на зъбите цементобласти.Тези клетки отделят колагенови влакна и междуклетъчно вещество, образувайки цимент.Образува се едва 4-5 месеца след раждането по време на развитието на корените на зъбите. От външния слой на зъбната торбичка се образува зъбът. лигамент (периодонтиум)(Фиг. 7.33, д). Така в пренаталното развитие се развиват само короните на млечните зъби.
Изригването на зъбите се осигурява от три фактора. Първият и най-важен е растежът и развитието на корена, което води до изхвърляне на короната през лигавицата. Вторият фактор е нарастването на пулпата, което води до повишаване на налягането вътре в зъбния зародиш. Освен това се получава допълнително отлагане на дъното на зъбната алвеола. костна тъканслоеве.
Полагането на постоянните зъби започва много рано, в началото на 5-ия месец от вътреутробното развитие. Зародишът на постоянния зъб се намира зад зародиша на млечния, като развиващият се корен на последния през първите две години след раждането има индукционен ефект върху развитието на зародиша на постоянния зъб. Хистологичните процеси, които водят до образуването на млечни и постоянни зъби, са еднакви. Когато млечните зъби никнат, в зачатъците на постоянните зъби се образуват емайл и дентин. За известно време те остават в латентно състояние и се активират едва след като челюстта достигне достатъчно голям размер. В процеса на заместване растежът на постоянен зъб и натискът на емайла му върху корена на млечния водят до резорбция.
остеокласти (клетки, които разрушават костната тъкан) на по-меката тъкан - дентина на млечния зъб. Последният се изтласква и се заменя с постоянен.
Развитие на отдела храносмилателнасистеми е трудно да се разгледат в детайли, тъй като чревната тръба е силно удължена, диференцирана и свързана в развитието си с много други системи: ендокринна, дихателна, отделителна, кръвоносна, нервна и вторична кухина - целом. Нека разгледаме само най-ранните и често срещани процеси, протичащи в чревната тръба, както и промените в връзката й с жълтъчната торбичка.
Чревната тръба възниква от ендодермата на покрива на жълтъчната торбичка и съседния висцерален лист на мезодермата в процеса на отделяне на тялото на ембриона от екстраембрионалните части с помощта на главата, опашката и страничните гънки на багажника (виж фиг. 7.24, b 2, b 3). На 4-та седмица тя е представена от сравнително проста тръба, състояща се от предното черво, стомодалната (орална) пластина, сляпо затворена отпред, средното черво, свързано с жълтъчната торбичка
през жълтъчното стъбло (канала) и задното черво, сляпо затворено в края на опашката от клоакална (анална) мембрана
(фиг. 7.34).
През периода от 4-та до края на 8-та седмица от ембрионалното (вътрематочно) развитие червата се удължават и започват да се диференцират. От предната му част се образуват фаринкса, хранопровода, стомаха и дванадесетопръстника с неговите производни (черен дроб и панкреас), преди да се влее в него общият жлъчен канал.
Оформя се средното черво U-образно-разнибримка, която е свързана с жълтъчната торбичка през жълтъчния канал (фиг. 7.35, а). Частта от червата между дръжката на жълтъка и стомаха ще се превърне в тънките черва и секцията
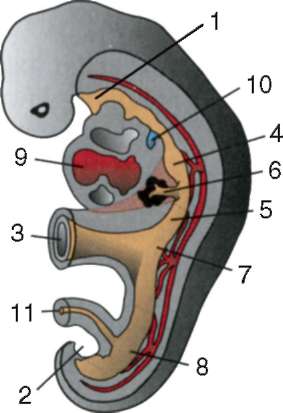
Ориз. 7.34.Храносмилателна система на 4-седмичен ембрион: 1 - стомо-делова плоча; 2 - клоакална мембрана; 3 - жълтъчен канал; 4 - стомаха; 5 - дванадесетопръстника; 6 - черен дроб; 7 - средно черво; 8 - задно черво; 9 - сърце; 10 - полагане на дихателните органи; 11 - урахус
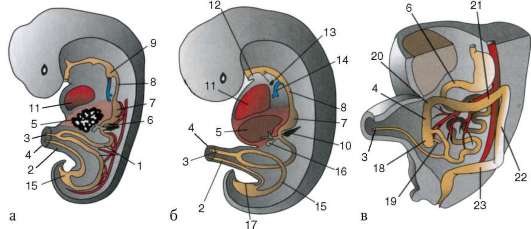
Ориз. 7.35.Развитие на човешката храносмилателна система: а - на 32-ия ден; b - на 36-ия ден; c - по-късни етапи; 1 - U-образна бримка на червата; 2 - урахус; 3 - жълтъчен канал; 4 - коремна дръжка (пъпна връв); 5 - черен дроб; 6 - дванадесетопръстника; 7 - стомаха; 8 - хранопровод; 9 - ларинкса; 10 - панкреас; 11 - сърце; 12 - език; 13 - маркер на щитовидната жлеза; 14 - рудимент на трахеята и белите дробове; 15 - задно черво; 16 - жлъчен мехур; 17 - клоакална мембрана; 18 - цекума; 19 - апендикс; 20-23 - участъци от дебелото черво
притискайки каудално към жлъчния канал, - в дебелото черво. Средното черво се удължава, част от него изпъква в кухината на коремната дръжка, образувайки херниална издатина на коремната стена на ембриона.
През същото време някои части на червата, въртящи се, променят позицията си (фиг. 7.35, b). До 10-та седмица изпъкналата част на чревната примка се изтегля назад през пъпния ръб, заемайки окончателното си положение в коремната кухина. Малко по-рано, на мястото на прехода на тънките черва в дебелото черво, се развива цекума. Първоначално той се увеличава по размер, но до 3-ия месец дисталната му част започва да изостава от растежа на другите части (неравномерен растеж), в резултат на което диаметърът на тази част е много по-малък и се образува апендикс. образувани. От задното черво дебелото черво се образува под ъгъла на далака, включително ректума (фиг. 7.35, c). Развитието на клоакалния край на задното черво е тясно свързано с развитието на урогениталния отвор (фиг. 7.36). Уроректалната преграда, която разделя клоаката на урогениталния синус и ректума, се формира на 5-6-та седмица. Пробивът на аналната мембрана настъпва през 8-та седмица.
На 2-ия месец от вътрематочното развитие започва бърза пролиферация на епитела на хранопровода и дванадесетопръстника, което
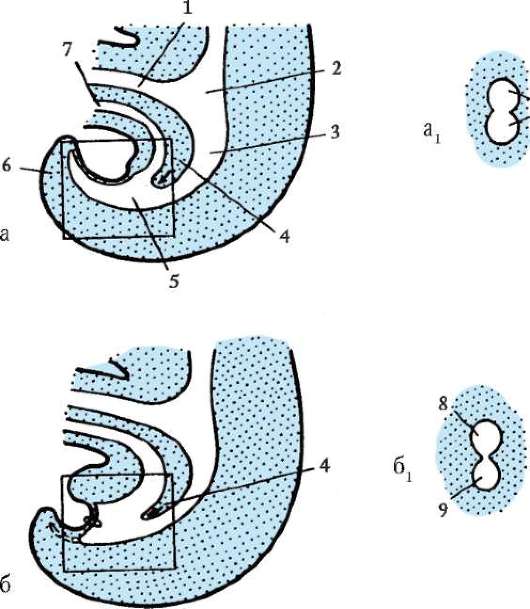
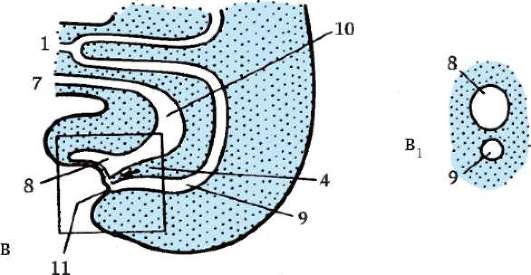
Ориз. 7.36.Развитие на клоакалната област на задното черво в човешкия ембрион: а - на 4-та седмица; b - на 6-та седмица; c - на 7-та седмица; a 1 b 1 и c 1 - напречни разрези през клоаката (нивата на изрязване са обозначени с квадратчета на фиг. a; b; c). 1 - стъбло жълтък; 2 - средно черво; 3 - задно черво; 4 - урогенитална преграда; 5 - клоака; 6 - опашка; 7 - алантоис; 8 - урогенитален синус; 9 - ректума; 10 - пикочен мехур; 11 - анална мембрана; стрелките показват посоката на растеж
води до временно затваряне на лумена им. До края на 2-ия месец настъпва реканализация на тези органи, т.е. възстановяване на пропускливостта.
сърцечовек започва да се развива от края на 3-та - началото на 4-та седмица от splanchnomesoderm под формата на сдвоени рудименти, разположени под фаринкса (фиг. 7.37). Тъй като тялото на самия ембрион е ограничено от гънките на багажника и предното черво е затворено от вентралната страна, сдвоените ендокардни тръби се затварят в една, разположена по средната линия (виж фиг. 7.37). За земноводните е доказано наличието на индуктивно действие на ендодермата върху прекордиалната мезодерма. Това се доказва и от експеримент върху пилешки ембрион, когато след отстраняване на дъното на предстомашието се получава ембрион с двойно сърце. Сдвоените зародиши на целомичните кухини се сливат от вентралната страна, образувайки перикардната кухина.
Регионалната диференциация на сърцето започва с бързо удължаване на първичната сърдечна тръба, което води до нейното огъване и придобиване на 5-образна форма (фиг. 7.38). Отбелязва се, че образуването на завой с локални промени във формата на клетките по дължината на сърдечната тръба се стимулира от самата сърдечна тръба. Експлантация (ситуация „извън тялото“ или ex vivo),сърдечната тръба запазва способността да образува 5-образна структура.
Ориз. 7.37.Развитие на човешкото сърце на 21-22-ия ден: а - общ изглед на ембриона; b - надлъжен разрез на конуса на главата на ембриона; c - напречно сечение на ембриона: 1 - ниво на разрез; 2 - нервен валяк; 3 - сомити; 4 - първична лента; 5 - мозък; 6 - предстомашие; 7 - сърдечна тръба; 8 - перикард; 9 - амнион; 10 - орофарингеална мембрана; 11 - невронна бразда; 12 - акорд; 13 - сдвоени маркери на дорзалната аорта; 14 - сливащи се сърдечни тръби; 15 - жълтъчна торбичка
Ориз. 7.38.Развиващото се човешко сърце: а - на 21-22-ия ден; b - на 23-ия ден; c - на 24-ия ден (изглед от коремната страна); г - на 28-ия ден (надлъжен разрез): 1 - първата двойка аортни дъги; 2 - сливащи се сърдечни тръби; 3 - несливащи се сърдечни тръби; 4 - артериален ствол; 5 - крушка на сърцето; 6 - вентрикула; 7 - атриум; 8 - венозен синус; 9 - канал на Кювие; 10 - пъпна вена; 11 - жълтъчна вена; 12 - атриовентрикуларен канал
В първоначално каудалния край е венозният синус, в който се вливат големи вени: каналите на Кювие, пъпната и жлъчката. По-късно венозният синус ще стане част от дясното предсърдие. Краниално към венозния синус, атриумът се образува от разширената част на сърдечната тръба, а вентрикулът се образува от извитата средна част. Преходната област, където вентрикулът се стеснява, се нарича конус или луковица. По-късно ще бъде включен в стената на дясната камера. Конусът преминава в артериалния ствол, от който се отклоняват корените на коремната аорта.
След образуването на атриума и вентрикула се появяват външни признаци на предстоящото разделяне на сърцето на дясната и лявата половина, появява се средната бразда. В този момент в тъканите на развиващото се сърце се отбелязва фокална клетъчна смърт и взаимодействие на извънклетъчни и клетъчни елементи на фактори. След известно време на вътрешната повърхност на вентрикула, съответстваща на нивото на външния жлеб, се образува преграда от мускулни връзки, израстващи от върха на сърцето към атриума. На дорзалната и вентралната стена на стеснения атриовентрикуларен канал се образуват ендокардни възглавнички от рехав мезенхим. По-късно се трансформират в плътни съединителната тъкан, растат заедно и разделят канала на десен и ляв канал. В същото време се появява първична средна преграда, която по-късно се заменя с вторична. В него има отвор, наречен овал, през който
кръвта от дясното предсърдие се влива в лявото. Необходим е за кръвообращението на плода.
Едновременно с промените в основната част на сърцето като самостоятелен орган, артериалният ствол се разделя на два канала. Този процес започва от корена на коремната аорта между четвъртата и шестата висцерална дъга. Разделянето възниква поради образуването на надлъжни гънки. Гънките са подредени спираловидно, растат вътре в артериалния ствол и, срещайки се, го разделят на аортата и белодробния ствол. В допълнение, гънките се разпространяват към конуса, където се образуват полулунните клапи на аортата и белодробния ствол от специализирани области, и по-нататък във вентрикулите, където се срещат с движещата се ендокардна тъкан на атриовентрикуларния канал и предсърдната преграда. Това води до пълно разрастване на интервентрикуларния отвор.
До края на вътрематочния живот остават отворени само клапата в овалния отвор на междупредсърдната преграда и артериалният (боталов) канал, който свързва белодробния ствол с аортата. Ще бъде разгледано по-подробно по-долу. Артериалният канал служи за отклоняване на част от кръвта от дясната камера към аортата, заобикаляйки белите дробове, докато те достигнат необходимото развитие и не получат функционален стимул във връзка с началото на дихателната функция (фиг. 7.39).
Големи артерииразвиват се в комбинация със сърцето, започвайки от 4-та седмица. Първичната кръвоносна система на ембриона функционира от края на 5-та седмица, до края на 8-та седмица се изпълнява основният план на структурата. артериална система. Сдвоените висцерални дъги, образувани на 4-та седмица, получават подходящо артериално захранване под формата на аортни дъги. Тези аортни дъги се издигат от артериалния ствол или по-скоро от разширената му част - артериалната торбичка (наричана още коремна или възходяща аорта) - и завършват в корените на дорзалната или низходящата аорта (виж фиг. 7.27).
Въпреки факта, че са положени шест двойки аортни (висцерални) дъги, те не съществуват едновременно. През периода, когато се формира шестата двойка дъги, първите две двойки вече се израждат. Петата дъга се появява само на кратко времепод формата на рудиментарен съд.
При образуването на главните съдове съществена роля играят корените на коремната и дорзалната аорта, както и третата, четвъртата и шестата двойка аортни дъги (фиг. 7.40).
Проксималните части на третата двойка образуват общите каротидни артерии (фиг. 7.41). Лявата четвърта дъга образува част от дъгата на аортата.
Ориз. 7.39.Схема на кръвообращението на плода: 1 - овален прозорец; 2 - междупредсърдна преграда; 3 - белодробна вена; 4 - горна празна вена; 5 - артериален (ботал) канал; 6 - белодробна артерия; 7 - низходяща аорта; 8 - пъпна артерия; 9 - пъпна вена; 10 - долна празна вена; единадесет - дуктус венозус; 12 - портална вена на черния дроб. I - черен дроб; II - долна празна вена; III - дясно предсърдие; IV - ляво предсърдие; V - низходяща аорта
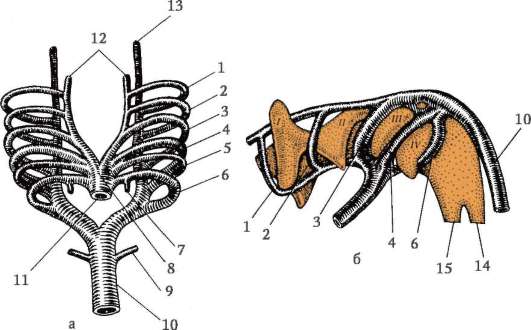
Ориз. 7.40.Развитие на аортните дъги: а - изглед от вентралната страна: 1-6 - аортни дъги; 7 - белодробна артерия; 8 - артериална торбичка, коремна аорта; 9 - седма междусегментна артерия, субклавиална; 10 - дорзална низходяща аорта; 11 - артериален ствол; 12 - външна каротидна артерия, корен на коремната възходяща аорта; 13 - вътрешна каротидна артерия, дорзален аортен корен
Дясната четвърта дъга става проксималната част на дясната субклавиална артерия. Дисталната част на тази артерия произлиза от дясната дорзална аорта. Проксималната част на лявата шеста дъга се превръща в проксималната част на лявата белодробна артерия, а дисталната част в артериалния (боталов) канал. Проксималната част на дясната шеста дъга образува проксималната част на дясната белодробна артерия, а дисталната част е намалена. Корените на дорзалната аорта се сливат много рано в несдвоената дорзална аорта, но на 7-та седмица от развитието дисталната част на десния дорзален аортен корен обикновено се свързва с дясната субклавиална артерия, както вече беше споменато по-горе. Това става възможно поради обратното развитие на сайта, свързващ десния корен с левия. Представянето на материал, свързан с развитието на сърцето и големите артерии, дава основание за съпоставяне
Ориз. 7.41.Образуване на най-големите артерии, производни на артериалния ствол, аортна торбичка, аортни дъги и дорзална аорта: а - 6-седмичен плод; б - 7-седмичен плод; c - 6-месечен плод: 1-6 - аортни дъги; 7 - артериален ствол; 8 - белодробна артерия; 9 - субклавиална артерия; 10 - дорзална низходяща аорта; 11 - аортна торбичка; 12 - вътрешна каротидна артерия; 13 - артериален (ботал) канал; 14 - възходяща аорта; 15 - белодробен ствол; 1b - външна каротидна артерия; 17 - обща каротидна артерия
на онтогенезата на човека с филогенетичното развитие на гръбначните животни (виж Глава 14), а също така ни позволява да се доближим до разбирането на механизмите на възникване на вродени малформации на сърцето и кръвоносните съдове.
Въпроси за самоконтрол
1. Какви видове онтогенеза съществуват? Избройте и опишете основните етапи на онтогенезата.
2. Какви са характеристиките на структурата и функциите на зародишните клетки?
3. Как се извършва оплождането и какви етапи включва този процес?
4. Опишете същността и морфологичните форми на етапите на ембрионалното развитие при представители на различни класове
хордови (раздробяване, гаструлация, органогенеза и образуване на профизорни органи).
5. Какви са особеностите на ембрионалното развитие на бозайниците?
6. Дайте примери за ембрионалното развитие на човешките органи, отразяващи еволюцията на прародителските групи хордови.
Онтогенезата е индивидуалното развитие на организма, съвкупност от последователни морфологични, физиологични и биохимични трансформации, претърпени от организма, от оплождането (по време на половото размножаване) или от момента на отделяне от майчиния индивид (по време на безполово размножаване) до края на живота си.
Разпределете предембрионални, ембрионални и постембрионални периоди. При плацентарните животни и хората се разграничават пренатален (антенатален) и постнатален (постнатален) период.
Съвкупността от събития, протичащи в ово(оо)генезата, но в интерес на процеса на индивидуално развитие на нов организъм, съставлява съдържанието на периода на прогенезата - предембрионаленпериод на индивидуално развитие. Започва преди момента на оплождането и образуването на зигота и се свързва главно с женската зародишна клетка. Последният в хода на гаметогенезата (овогенезата) придобива някои характеристики, които ще бъдат използвани не от само себе си, а от индивида на новото поколение, което е започнало индивидуално развитие. Една от тези характеристики, може би най-известната, е образуването в цитоплазмата на яйцето на по-голямо или по-малко количество жълтък, в зависимост от вида на животното, което се използва като хранителен материал при развитието на потомството. . Функционалната генетична активност на редица гени, която се проявява в тяхната транскрипция и пост(пост)транскрипционни промени в първичния и(м)РНК транскрипт, също е свързана във времето с периода преди оплождането. Образуваната в резултат на тази активност i(m)RNA организира синтеза на протеини, важни за ранните етапи на ембриогенезата.
Видове яйца.
Те се класифицират според количеството и разпределението на жълтъка. Може да бъде:
1. Алецитални яйца - никакъв или много малко жълтък
Пример: при бозайници
2. Изолецитален - има малко жълтък, разпределен е равномерно в цитоплазмата
Пример: ланцет
3. Полилецитални: а) телолецитални - жълтъкът е изместен към вегетативния полюс
б) центролецитален - жълтък в центъра.
Оплождане
Сливане на сперматозоид с яйцеклетка. Важни етапипроцес на оплождане: 1) проникване на спермата в яйцето; 2) активиране на различни синтетични процеси в яйцето; 3) сливане на яйцеклетката и спермата с възстановяване на диплоидния набор от хромозоми.
Продължителността на живота и способността за оплождане на яйцеклетки и сперма са ограничени. При повечето бозайници яйцеклетката запазва способността си да опложда 24 часа, а при човека 12-24 часа след овулацията. Сперматозоидите запазват оплождащата си способност в женските генитални пътища също няколко часа.
Директният контакт на сперматозоида и яйцеклетката причинява закономерни промени в двете клетки, които предизвикват проникването на ядрото и центриола на мъжката гамета в яйцеклетката и активирането на метаболизма на последната. Тези промени от страна на сперматозоида се наричат акрозомна реакция, а от страна на яйцеклетката кортикална реакция. Същността на акрозомалната реакция е следната. В момента на контакт с яйцеклетката в горната част на главата на сперматозоида, плазмената мембрана и частта от мембраната на акрозомния везикул, съседна на нея, се разтварят. Поради освобождаването на ензими от акрозомалната гранула, съседната част на яйчната мембрана се разтваря. Акрозомалната мембрана изпъква навън и образува израстък под формата на куха тръба. Последният се удължава, преминава през мембраните на яйцето и влиза в контакт с плазмената мембрана на яйцето. В областта на такъв контакт възниква издатина или туберкул на оплождане, след което плазмените мембрани на двете гамети се сливат и започва обединяването на тяхното съдържание. От този момент нататък спермата и яйцеклетката са една клетка - зигота. Акрозомната реакция е изключително бърза.
Активиране на яйцеклетката или кортикална реакция, която се развива в резултат на контакт със сперматозоид. Промени в повърхностния кортикален слой на ооплазмата и образуването на оплодителната мембрана. Тази мембрана, наричана още жълтъчна мембрана, възниква чрез отлепване на повърхността на яйцето. Под него се образува периолково пространство, в което се излива съдържанието на гранулите на кортикалния слой на цитоплазмата на яйцето. Оплождащата мембрана предпазва яйцето от проникване на излишни сперматозоиди, т.е. осигурява "блок на полиспермия". Течността, която се натрупва в околоплодното пространство, служи като специфична среда, в която протича развитието на ембриона до момента, в който той напусне мембраните на яйцата.
IN ембрионаленпериод, има три основни етапа: раздробяване, гаструлация и първична органогенеза. Ембрионалният или зародишният период на онтогенезата започва от момента на оплождането и продължава, докато ембрионът излезе от мембраните на яйцето. При повечето гръбначни животни той включва етапите на зигота, разцепване, гаструлация, хисто- и органогенеза.
Зиготата е едноклетъчен ембрион. В зиготата протича процесът на диференциация на цитоплазмата. Наблюдава се движение на ооплазмата. В него се появяват области, от които впоследствие се формират предполагаеми (предполагаеми) зачатъци на бъдещи органи.
Разцепване - поредица от последователни митотични деления на оплодена или инициирана за развитие яйцеклетка. Разцепването е първият период от ембрионалното развитие, който присъства в онтогенезата на всички многоклетъчни животни и води до образуването на ембрион, наречен бластула (еднослоен ембрион). В същото време масата на ембриона и неговият обем не се променят, тоест те остават същите като тези на зиготата, а яйцето се разделя на все по-малки клетки - бластомери. След всяко разделяне на разцепването клетките на ембриона стават все по-малки и по-малки, т.е. ядрено-плазмените отношения се променят: ядрото остава същото, а обемът на цитоплазмата намалява. Процесът продължава, докато тези показатели достигнат стойностите, характерни за соматичните клетки. Видът на разбиване зависи от количеството жълтък и местоположението му в яйцето. Ако има малко жълтък и той е равномерно разпределен в цитоплазмата (изолецитални яйца: бодлокожи, плоски червеи, бозайници), тогава раздробяването протича според вида на пълната униформа: бластомерите са с еднакъв размер, цялото яйце е смачкано. Ако жълтъкът е разпределен неравномерно (телолецитални яйца: земноводни), тогава раздробяването протича според вида на пълната неравномерност: бластомерите са с различни размери, тези, които съдържат жълтъка, са по-големи, яйцето се раздробява като цяло. При непълно раздробяване в яйцата има толкова много жълтък, че браздите за раздробяване не могат да го отделят изцяло. Разцепването на яйцеклетка, при което се раздробява само "шапката" на цитоплазмата, концентрирана върху анималния полюс, където се намира ядрото на зиготата, се нарича непълно дискоидно (телолецитални яйца: влечуги, птици). При непълно повърхностно смачкване в дълбините на жълтъка се появяват първите синхронни ядрени деления, които не са придружени от образуването на междуклетъчни граници. Ядрата, заобиколени от малко количество цитоплазма, са равномерно разпределени в жълтъка. Когато ги има достатъчно, те мигрират в цитоплазмата, където след образуването на междуклетъчните граници се появява бластодермата (центролецитни яйца: насекоми).
· Гаструлация – процесът на движение на клетките, съпроводен с растеж, размножаване, диференциация. Ембрионът на този етап се нарича гаструла.
Първоначално се образуват външните (ектодерма) и вътрешните (ендодерми) листове. По-късно се появява трети, среден зародишен лист (мезодерма), разположен в тялото на ембриона между ектодерма и ендодерма.
Има 4 основни начина за оформяне на външните и вътрешните листове. Често обаче има комбинация от няколко начина на гаструлация.
1 начин инвагинация, което се състои във факта, че определен участък от бластодермата, като запазва структурата на слоя, се завинтва в бластоцела. След това бластоцелът изчезва и се появява гастроцелът. Отворът, през който тази кухина комуникира с външната среда, се нарича първична уста или бластопор.
2 начин epiboly– обрастване на макромери с бързо делящи се микромери на анималния полюс. В ембрионите на такива животни първоначално бластопорът не се образува и гастроцелът липсва.
3-посочна имиграция– изчисляване на част от клетките на стената на бластулата вътре в бластоцела.
4-посочно разслояване -клетките отвън се трансформират в слой
а-ектодерма; b-ендодерма; v-бластоцел.
Полагането на третия зародишен слой става по два начина: телобластни и ентерокоелни.
Телобласт - метод за образуване на мезодерма в протостоми чрез отделяне на две мезодермални ленти от две първични ембрионални клетки (телобласти).
Enterocoelous - се състои в това, че от ендодермата на първичното черво се образуват издатини от двете страни - джобове (целомични торбички), които по-късно се завързват и растат между екто - и ендодермата, образувайки мезодермата; техните кухини се сливат една с друга и възниква вторична телесна кухина или целом.
Хисто- и органогенеза
Могат да се разграничат две фази.
1 неврулация, се състои в образуването на комплекс от аксиални органи - невралната тръба, хордата. Ембрионът на етапа на неврулация се нарича неврула. Първо се получава сплескване на клетъчния слой, което води до образуването на невралната пластина, чиито ръбове, издигайки се, образуват невралните гънки. Поради движението на клетките по средната линия на невралната пластина възниква депресия. На този етап на неврулация, отметката нервна системанаречена неврална бразда. В същото време невралната плоча изглежда се сгъва по средната линия и малко по-късно краищата й се затварят. В резултат на тези процеси се появява неврална тръба с кухина - невроцел. затварянето на гребените става първо в средата и след това в задната част на невралната бразда. Това се случва последно в частта на главата, която е по-широка от другите области. Предната, разширена част в по-нататъшното развитие образува мозъка, останалата част от невралната тръба - гръбначния мозък.
2 хисто- и органогенеза.
От ектодермата се образуват кожният епидермис и неговите производни, жлези, епител на устната кухина, вагинален епител, зъбен емайл и рецепторни нервни клетки.
От ендодермата - се образува епител на дихателните пътища и белите дробове, част от клетките на панкреаса, секретиращи клетки, епител на стомаха, червата.
От мезодермата - хрущялният и костен скелет, мускулите, бъбреците, кръвоносните съдове.