23. Анатомичната структура на стъблата на тревисти и дървесни едносемеделни растения.
24. Анатомичната структура на стъблата на тревистите двудолни растения.
25. Анатомичната структура на стъблата на дървесните двусемеделни растения.
26. Анатомичната структура на иглолистните стъбла.
27. Анатомичната структура на коренищата на едносемеделни и двудолни растения.
31. Понятието за стъблото, бягството. Функции и метаморфози на стъблото. Морфологични характеристики на стъблото.
Стъблото е въздушен, вегетативен, аксиален орган на растение, което има радиална симетрия и има апикален растеж.
Стъблото с пъпки и листа - бягство, Стъблови участъци, носещи листни възли: между възли - пролуки, В зависимост от дължината на междувъзлите, издънките могат да бъдат удължени (зърнени култури), съкратени (глухарче).
От бъбрек се развива бягство.
апикален пъпка - горната част на стъблото, покрита с млади пъпки на листа: конус на растеж, в основата му има първични туберкули - от тях се формират първични листа. В синусите на първичните туберкули са вторични туберкули, които дават страничен или подмишничен бъбреците. Страничните клони също растат с върховете си и всеки клон завършва с апикална пъпка.
С настъпването на есента всички пъпки навлизат в сезонно състояние на покой. Те се наричат \u200b\u200bзимуващи. Те могат да бъдат вегетативната (образува листа или издънки) и вегетативно генеративно или цъфтеж (пролетно цъфтеж в цветя или съцветия). Защитени от люспи с твърда кожичка. Те имат склероиди, понякога корк. Цъфти поради резерви, депозирани в корена, стъблото (или в самите люспи).
Много аксиларни пъпки остават в покой дълго време и растат само с оста си на стъблото в съответствие с удебеляването му - спален бъбреците. Те започват да растат, ако премахнете апикалния бъбрек. Понякога те могат да бъдат потопени в дърво и да се събудят след почивка или къща от дърва.
Подчинени клаузи пъпки се полагат в възли и върху междувъзлия, корени, коренища, листа - резерв за вегетативно размножаване. Произход: от камбий, пелоген, велосипед, кора. Това са пъпки за подмладяване - те дават издънки, подобни на младите разсад.
Морфологични признаци на стъблото.
I. По позиция в пространството стъблата са: изправени, издигащи се, пълзящи, катерещи, катерещи и др.
II. Разклоняване на издънки.
1. Дихотомен - точката на растеж се разклонява на 2 нови (долен, папрат, мохоподобни).
2. Моноподиален - основната ос расте на върха си, страничните клони се развиват от страничните пъпки, които също се разклоняват моноподиално (иглолистни дървета - много силно се оценяват в горското и корабостроенето).
3. Симподиален - апикалната пъпка умира, вместо това се развива странична пъпка, расте често във вертикална посока, сякаш продължава растежа на главното стъбло и т.н. (бреза, върба, ябълково дърво, домат, картоф).
Известно е смесен разклоняване: първо моно-, а след това симподиален (в дървесни плодове, памук).
Интензивното цъфтене на голям брой пъпки прави издънката съкратена, което осигурява развитието на огромен брой листа по короната, т.е. голяма фотосинтетична повърхност.
4. Фалшиво дихотомично разклонение - под апикалния бъбрек се образуват два противоположно разположени аксиларни бъбрека; започвайки да растат, образуват вилица (имел, дроп, карамфил, люляк, конски кестен).
Изследването на разклонителните системи е от голямо практическо значение. Можете да регулирате реколтата.
III. Phyllotaxy: листата се монтират на стволови възли.
1. Друго - възелът има едно листо - ябълково дърво, бреза, клен.
2. Обратното - два листа на възел (лабиоекум, валериана, хортензия).
3. Набръчкани - три или повече листа - олеандър.
4. Базалната розета е иглика.
Редките листа са подредени на стъблото по спирала. Следвайки завоите си, можете да намерите два листа, разположени вертикално един над друг. Линията, свързваща листата в надлъжните редове на стъблото - ortostiha, Между краищата на ортостомията останалите листа са подредени в спирала, броят на завоите на спиралата между два листа, разположени на един и същ ортостих, се наричат \u200b\u200b- лист цикъл. Следващата подредба на листата е направена под формата на фракция, където броят на завоите на спиралата в цикъла на листа става числителят, а броят на листата в цикъла е знаменателят.
IV. Формата на стъблото в напречно сечение.
Формата на стъблото в напречното сечение е най-разнообразна: кръгла, плоска, оребрена, многостранна и др .; може да бъде куха и направена, гола и опушена.
Продължителност на живота и биоморфологични групи от растения. Кипарис - 3000 години, кестен - 2000 години, дъб - 1200 години, липа - 1000 години, топола - 300-600 години, ябълково дърво - 200 години, сладка череша - 40-100 години, бор - 50 години. В зависимост от продължителността на живота и вида на издънките, растенията се делят на:
дърветата - Големи растения със силно развити многогодишни стъбла. Има вторично увеличение.
храсти - основният багажник отсъства или е слабо изразен, разклоняването започва почти на почвената повърхност (4-6). Barberry, irga, дива роза. Има групи храсти не повече от 1 м - мащерка, саксаул.
храсти - издънките в долната част остават многогодишни, в горната част - едногодишните леторасти замръзват или изсъхват - пелин, астрагал, жълт кантарион;
тревист- смъртта на надземните стъбла се наблюдава в края на вегетационния период. Повечето полски растения са зърнени култури, грах, боб, лен, коноп;
двугодишно тревисто - през първата година се развива само розетка от кореновите листа, а цветята, плодовете и удълженото стъбло - през втората (цикория, кокошка, ружа, сладка детелина);
целогодишен - трайни подземни части и годишната смърт на надземните части (валериана, момина сълза, коприва).
Сред тревистите растения има зимни и пролетни форми:
а) ако семената покълнат през пролетта - пролетта;
б) зима - покълване на семената през есента (0-5 градуса).
Стволови функции:
1. Скелетът, свързващ листата и корените заедно, придава навика на растението;
2. Провеждане;
3. Складиране;
4. синтетичен;
5. Органът на вегетативно размножаване.
метаморфоза:
подземен: 1. Коренище - подземна издънка, листа под формата на люспи, размерът на коренищата се определя от функциите: ако за запаса - дебел (каламус, хвощ, купен), за размножаване - с пъпки за подновяване (житна трева, кит убиец).
2. Клубените - се образуват в краищата на подземни стъбла - сергии, имат бъбреци - очи (3 или повече всяка).
3. Луковици - съкратено стъбло - дъното, пъпките и листата.
земята: 4. Шипове - глог, обръщане.
5. Мустаци - тиква, краставица, пъпеш.
6. Филокладий - стрък под формата на листо - игла.
Стъблото - въздушен, вегетативен, аксиален орган на растението; има дълъг растеж на дължина, използвайки апикалната меристема в конуса на растежа или меристемата за вмъкване; носи листа, които се полагат в определен ред върху растежен конус под формата на туберкули; може да се разклонява поради пъпки, вградени в аксиларни листа; има радиална структура и няколко равнини на симетрия.
В нарастващия край стъблото носи апикална пъпка - растежен конус, облечен в млади, мутирали листа. Тя завършва с точка на растеж, зад която са разположени първичните туберкули - зародишите на листата. Още по-ниско в синусите им възникват вторични туберкули - рудиментите на страничните издънки.
Хистогенната теория на Хапщайн (1818 г.): дерматоген, пропускливост, плерома - не е вярно за стъблото. В началото на 20-ти век Шмид формулира теорията за туниката и тялото, според която растежният конус в покритосеменните растения и част от гимнаспермите се състои от две хистологично различни части с различна меристемична активност. Външната част е туниката, клетките й се разделят перпендикулярно на повърхността на стъблото и образуват епидермиса, понякога епидермиса и няколко слоя или цялата първична кора; вътрешната част - тялото - клетките се разделят във всички посоки и образуват останалите слоеве на първичната кора и центъра.
Първичната структура на стъблото се развива по време на диференциацията на апикалните клетки на меристемата.
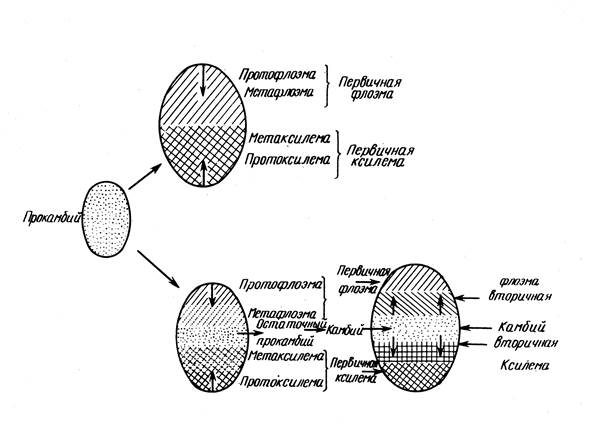
На нивото на първите примордии на листата клетките на туниката и корпуса престават да се делят; тяхната периферна част отива към образуването на първичната кора, вътрешната част към образуването на ядрото. Но между тях има няколко реда активни меристематични клетки, разположени в пръстен - образователен пръстен, Клетките му в основата на млади листни примордии пораждат първичната латерална меристема - прокамбия.
procambium представлява ембрионалната фаза в развитието на първичната проводяща меристема, следователно нейното местоположение до голяма степен определя последващото местоположение на проводящата система. Ако тя е положена с непрекъснат слой (пръстен), тогава възникват непрекъснати слоеве от флоема и ксилем, ако прокамбият е положен под формата на снопове (ленти), тогава първичните проводими тъкани са подредени под формата на снопове. Клетките на периферните слоеве на образователния пръстен, които не участват в образуването на прокамбий, образуват велосипед.
Прокамбиум възниква в основата на листния ембрион и оттук неговото развитие се простира в две посоки - акропетално, т.е. до върха на листния примордий, и базипетално, т.е. надолу по стъблото, където е прикрепен към други снопове, възникнали по-рано.
Флоемата се формира по-рано от ксилемата и се развива центропетално, първо протофлоем, след това протоксилем (пръстеновиден и спираловиден съд), след това метаксилем. Така се формира първичната структура на първичната структура.
На напречната секция на стъблото се разграничават три групи тъкани: покривна, първична кора и СТС.
Покривната тъкан на стъблото тревисто двудолно растение - епидермис. Клетките са леко сивкави, удължени по посока на стъблото, стомасите са малко или не.
Първичната кора се състои от коленхима под формата на кичури в ъглите на стъблото в лабиата или в ребрата му в зонтик, по-рядко под формата на пръстен в тиква, нощник. В периферията първичната кора се състои от редуващи се ленти от хлоренхим и механична тъкан. Във вътрешната част е безцветен паренхим. Най-вътрешният слой - ендодермата в някои случаи - е нишестена вагина, съдържаща малки зърна от защитено нишесте.
Периферната част на централния цилиндър е велосипедът. Провеждащите тъкани са подредени под формата на снопове или непрекъснат пръстен (в зависимост от местоположението на прокамбиума). Преходът към вторичната структура е свързан с полагането на камбий, в зависимост от което се разграничават следните видове структура на стъблата:
1. Прокамбий е положен в непрекъснат пръстен, първичната структура е несвързана; камбий се формира от прокамбий; второстепенната е не-група.
2. Прокамбий е положен с нишки - основната структура е греда; камбий се образува от прокамбий в снопове и от основната тъкан на СТС между сноповете. Образува се непрекъснат камбиален пръстен: вторичната структура е несвързана.
3. Първичната и вторичната структура са като лъчеви, защото междулъчевият камбий не се образува.
4. Първичната и вторичната структура са като лъчеви, защото прокамбият се полага в отделни шнурове и полученият междулъчеви камбий образува основната тъкан на C.O.C., а не флоемата и ксилема.
Стъблото на едносемеделни тревисти растения.Стъблото на едносемеделни тревисти растения се характеризира със снопчеста структура, сноповете са разположени на случаен принцип и отсъствието на вторичен растеж. Основните характеристики на анатомичната структура на стъблата се определят от системата на следи от листата: от основата на листа в стъблото са снопове, разположени около цялата обиколка, така че всички съдови снопове на монокотиледони са следи от листа (тип палма). SVPs на монокотиледони имат характерна форма: два големи точкови съда (симетрично) и 1-2 по-тесни с пръстеновидни и спираловидно-пръстеновидни удебеления. Въздушна кухина, съседна на крайната част от тях, се образува на мястото на разрушаване на елементите на протоксилема. Размерът на гредите се увеличава от периферията до центъра.
Има разлики в структурата на кората:
1. Монокотиледони с ясно изразена първична кора - (аспержи): се състои от няколко слоя клетки, носещи хлорофил; коленхима отсъства.
2. При други еднодолни (палма, зърнени култури) първичната кора не се изразява; непосредствено под епидермиса има участъци от хлоренхим, заобиколени от склеренхим, който се слива със склеренхима на SVP.
Голяма централна кухина (зърнена слама) се образува в междувъзлията на много монокотиледони, в тези случаи сноповете се изместват към периферията, но също така са разпръснати: по-малки към периферията, големи към кухината.
Структурата на стъблата на дървесните двудолни.Основната структура е без прах. Вторичният е свързан с полагането на пелогена и камбия. Камбият образува вторична флоема и ксилем. Флоем - лакът - твърд и мек е разположен под формата на трапец, разделен от върховете на първичните ядра лъчи; Функцията на Sapwood е 1 mm в камбий. Дърво под формата на дървени пръстени; звуковото дърво е механична функция.
Ядрото е ронливо, първо жива тъкан с запас от хранителни вещества. Свързани с челото и кората от основните лъчи (първични и вторични)
Фелогенът се образува поради епидермиса и слоевете на първичния кортекс, образува перидермата и кора. Под него първичната кора е запазена - колленхима, хлоренхим, тъкан за съхранение - ендодермата не се изразява (понякога се диференцира в нишестена вагина).
Според Глобалния фонд X1 (1987), кора във фармацевтичната практика - Това е външната част на стволовете, клоните и корените на дърветата и храстите, разположени до периферията на камбиума. По външни знаци цялата кора има вид на тръбни, набраздени или плоски парчета с различни размери. Външната повърхност на кората с кафяви или сиви тапи обикновено е гладка или с надлъжни (или напречни) бръчки, понякога с пукнатини. Кората на клоните и стволовете има заоблени или продълговати лещи, понякога върху нея могат да бъдат листни лишеи (те трябва да бъдат отстранени по време на прибиране на реколтата). Вътрешната повърхност на кората обикновено е по-лека, гладка или оребрена. Напречната извивка обикновено е неравномерна: цепеща се, влакнеста или гранулирана.
На микропрепаратобърнете внимание на външната кора, разположена към периферията от края на основните лъчи и състояща се от перидермата и остатъците от първичната кора и вътрешната, състояща се от флоема. Дебелината, цветът и естеството на коркът, наличието на колленхима, съотношението на дебелината на първичната и вторичната кора, ширината на основните лъчи, броя, разположението и структурата на голени влакна, каменисти клетки, както и включването на калциев оксалат, млечни клетки и клетки с етерично масло имат диагностична стойност.
Структурата на иглолистните стъбла.Основният е сноп, след това се образуват непрекъснат камбиален пръстен и пръстени от флоема и ксилем. Полага се пелоген.
В кравата: няма коленхима, има катранени пасажи.
2. Във флоемата няма придружителни клетки, голени влакна, паренхим.
3. Камбият е многосекционен.
Дървесината се състои от трахеиди, механични влакна и дървен паренхим отсъстват.
Основната структура е лъч.
Структурата на стъблата на дървесни едносемеделни растения.Особени за дървесни представители на семейство лилии (драцена, юка, алое). Изградено е младо стръкче в близост до растежния конус, както при всички монокотиледони: покривна тъкан - епидермиса; под него е първичната кора, представена от няколко слоя ламеларен коленхима; Центърът започва с велосипед и съдържа SVPs със затворен обект, разположени на случаен принцип. Сгъстяването възниква поради образуването на удебеляващ пръстен, образуван от велосипеда или от вътрешния слой на първичната кора. Образува центробежен центроксилен концентричен SVP. Покривната тъкан е корк.
Структурата на коренищата на едносемеделни растения.Подобно на стъбло на дърво. Покривната тъкан е лигифицирана екзодерма; първичната кора винаги е развита, представена от тъкан за съхранение; ендодерма, характерна за петна от Каспари и удебелявания във формата на подкова. В CSC - колатерални снопове (V) и концентричен центрофлоем. Образува се от уплътняващ пръстен.
Структурата на коренищата на двудолни.Покривната тъкан е корк, перидермата се формира от фалогена; първична кора - паренхим за съхранение; ендодерма с петна от Каспари или подкова; в центъра местоположението на тъканите е подобно на земното стъбло; в типа греда - греда; в преходни и непукисти - непукисти.
Еволюцията на стелата.В процеса на еволюция се наблюдава промяна в структурата и относителното положение на различни растителни тъкани, особено на проводящите. Установени са няколко еволюционни типа стволови структури, свързани с центъра или стелата (Ван Тихем - френски ботаник). По-късно звездната теория получи блестящо потвърждение при изследването на изчезнали растения (риниофити). Най-древният тип е протостела. Тежестта на ксилема, покрита със слой от флоема, актиностела - ксилемата има формата на звезда (плуно, хвощ), заобиколена от флоема
Сифонна стена - появява се сърцевината.
Диктиостела - сифоностела се нарязва на отделни снопове (папрати).
Eustela - отворени колатерални снопове (двудолни).
Atactostela - в монокотиледони затворени колатерални снопове, разположени на случаен принцип.
Заключение.За макро- и микроскопска диагностика на лекарствени растителни материали трябва да са известни различни структурни типове на стъблото и коренищата.
Стъблото е част от лечебния растителен материал - трева (Herba). С макроскопска диагноза билки в съответствие с Държавната фармакопея X1 (1987) отбелязват следните характеристики на стъблото: вид разклоняване, форма на напречно сечение, опушен, размери (дължина и диаметър в основата), разположение на листата. Относно микропрепаратите напречното сечение на стъблото се отбелязва от структурните особености на епидермалните клетки, разположението на проводящите снопове, наличието и естеството на местоположението на механичните тъкани, кристалните включвания, съдовете, секреторните канали, млечните канали и други диагностични характеристики.
Ризоми (Rhizomata) в анализа по външни знаци те се различават по форма (проста или разклонена, цилиндрична или овална, ясно изразена, права, извита или усукана и др.), характеристики на външната повърхност (плоска или набръчкана със следи от отстранени корени) и характер на счупването (гладка, зърнеста, цепеща се или влакнеста). Понякога с просто око естеството на подреждането на проводящите греди се разглежда на почивка. С микроскопска диагноза обърнете внимание на структура без възел или сноп, характеристики на местоположението и видове снопове, структурата на вътрешната тъкан, съхранението и сърцевината, наличието на различни контейнери, механични елементи, канали, запаси от мляко, кристали на калциев оксалат, резервни хранителни вещества (нишесте, слуз, инулин, мастни масла и др. е.).
В анализ луковици (Bulbi), грудки (Tubera) и коренчета (Bulbotubera), форма (сферична, яйцевидна, овална, удължена, сплескана, с вретенообразна форма и др.), размер, повърхностен характер, структурни особености на люспите (суха или сочна) материя. С микроскопски диагностиката отбелязва характеристиките на тъканта за съхранение и местоположението на сноповете.
Структурата на стъблото на двудолни тревисти растения
В ранните стадии на развитие двудолното стъбло има първична структура. В резултат на дейността на първичните меристеми на конуса на растежа се образуват епидермиса, първичната кора, централният цилиндър и сърцевината.
епидермисстъблото има малък брой стомаси, обикновено покрити с кожички. Под епидермиса се намират тъкани първична кора.Външният му слой (екзодерма) често се формира от коленхима, която се намира или в непрекъснат пръстен (слънчоглед), или в отделни секции. При растения с оребрени стъбла (тиква) колеленхимата запълва издатините на стъблото. В тънките стебла на коленхимата обикновено е ъглов, в стъбла с активно вторично удебеляване - ламелен. Основната част от първичната кора (мезодерма) е съставена от хлоренхим и паренхим. Тук могат да се развият дихателни пътища и контейнери с секрети. Клетките на вътрешния слой на първичния кортекс (ендодерма) обикновено се пълнят с големи нишестени зърна (нишестени влагалища).
Централен цилиндър(стела) се състои от велосипед и проводима система. Ядрото еот паренхима, който често се разрушава и на негово място се образува въздушна кухина. Перикълът е представен от един или повече реда склеренхим или отделните му връзки. Проводимата система има формата на кух цилиндър, който разделя първичната кора и сърцевината. Проводимите снопове, които изграждат този цилиндър, са разделени от междупанелен паренхим, основните лъчи, които свързват сърцевината и първичната кора. Повечето двудолни се характеризират със специален тип централен цилиндър - еустелът, на напречното сечение на който гредите са подредени в пръстен (фиг. 4).
Фигура 4. Дикотиледони на Eustel.
При двудолни растения слой от прокамбиеви клетки между първичната флоема и ксилем запазва способността да се дели и се превръща във вторична образователна тъкан - т.е. камбий, която е разделена чрез поставяне на вътрешни елементи на вторичния ксилем, а към периферията - вторичната флоема. Гроздовете от двудолни растения са отворени, благодарение на работата на камбия, букетът расте, диаметърът му се увеличава. Активността на камбиума осигурява появата на вторични елементи в стъблото, т.е. прехода от първичната към вторичната анатомична структура.
Вторичната структура е характерна за всички двусемеделни и иглолистни растения - треви и дървета. Разнообразие от видове структура се дължи на първо място на местоположението на проводими тъкани, което се определя от полагането на Прокамбий и активността на камбий.
Прокамбиумът в процеса на диференциация на растежния конус се полага под формата на шнурове. Ако шнурите са разделени от достатъчно широки редове паренхим, тогава се образува сноп или преходна структура на стъблото; ако прокамбиевите шнурове са толкова близо, че се сливат в цилиндър, се образува несвързана (твърда) структура.
1. Структурата на гредата на стъблото намира се в някои тревисти растения (копър, детелина, пеперуда, грах, люцерна). Прокамбиални шнурове, положени в конус на растеж в един кръг около периферията на централния цилиндър, Всеки прокамбиален шнур се превръща в отворен колатерален сноп, състоящ се от първичен ксилем, първична флоема и ивица камбий между тях. Камбиалните клетки, разделящи се, дават нови (вторични) елементи на проводящия лъч: навътре - ксилема към периферията - флоема. В този случай най-младите участъци от флоемата и ксилема прилежават към камбиума, докато по-старите се придвижват към периферията на лъча. Най-крайните позиции ще бъдат заети от първичната флоема и ксилем. Провеждащите лъчи са разделени от широки междулъчеви зони и обграждат мощно ядро.
В повечето растения камбият работи активно и диаметърът на сноповете значително се увеличава.Активността на камби с лъч стимулира паренхима, който отделя снопчетата, който започва да се дели, давайки вторична образователна тъкан. Снопчетата и междинните снопчета меристеми се затварят и образуват непрекъснат камбиален пръстен, чиято активност осигурява равномерно удебеляване на стъблото. Ако снопът камбий дава елементи от вторичен ксилем и флоема, тогава междукамерен камбий произвежда паренхим (в копър, зърнен зор и т.н.). В детелина, люцерна и други растения междубиещият камбий снася предимно склеренхим-подобни клетки в посока на ксилема. Тези клетки са удължени, дебелостенни лигнифицирани. Следователно забавянето на събирането на детелината намалява качеството на сеното. В тези растения е запазена отчетлива структура на гредите през целия живот на стъблото.
Структурата на гредата има стъбло детелина, Под епидермиса се намира първичната кора, представена от паренхим, носещ хлорофил. Външният ред на клетките му е ламеларният вътрешен коленхим - клетките на нишестената вагина (ендодерма). Границата между първичния кортекс и централния цилиндър е ясно видима поради склерихима от перицикличен произход.
Склеренхимът образува половин дъги над отворени колатерални снопове, подредени в кръг. Пакетът съдържа първична флоема, съседна на склеренхима, вторична флоема, камбий, вторичен ксилем и първичен ксилем. Камбиятът на лъча се слива с междулъчевия лъч, образувайки плътен меристематичен пръстен. Камбиятът между лъчевите лъчи полага вътрешният паренхим, който свързва проводящите снопове. Има пръстен (цилиндър) от лигнифицирани тъкани, осигуряващ висока здравина на стъблото. Проводимите тъкани обаче остават фрагментирани под формата на снопове. Стъблото ядро \u200b\u200bе представено от паренхим.
Свързващата структура на стъблото може да бъде заместена с непрекъснато твърдо вещество с възрастта. 2. Преходна структура на стъблотохарактерно за слънчоглед, Първоначално стъблото се оформя по същия начин, както в детелината, но междукамерен камбиум не поставя паренхима, а ксилемата и флоемата. Образуват се допълнителни проводими греди. Постепенно всички снопове могат да се слеят в непрекъснат трислоен (ксилем, камбий, флоем) цилиндър. В горната (млада) част стъблото има структура на гредата, в основата (в старата част) е без куп.
3. Структура без лъч характерно за стъблата на много билки и дървета. В растежния конус на тези растения прокамбиалните шнурове са толкова близки, че образуват почти непрекъснат цилиндър (в напречното сечение изглежда като пръстен). Диференциращ се, прокамбиалният цилиндър (пръстен) привежда първичния ксиломен цилиндър към центъра, първичният флоелен цилиндър към периферията и между тях се образува камбиален цилиндър.
Вторичните промени са свързани с работата на камбия. Камбиевите клетки се разделят успоредно на повърхността на стъблото, докато 10 ... 20 пъти повече клетки се отлагат навътре, отколкото навън. Вътре камбият образува елементите на вторичния ксилем (дърво), външно вторичната флоема (лапа) се отлага. На някои места камбият полага от двете страни паренхимните клетки на основните лъчи.
Особености на несъбирателната структура на тревните стъбла могат да бъдат разгледани на примера на стъбло лен, Стъблото е покрито с епидермис, под който има хлабав хлоренхим на първичната кора. Голям брой стомаси в епидермиса и отсъствието на колленхима в първичната кора са от съществено практическо значение, тъй като по време на техническото промиване с лен не среща пречки и лесно прониква в стъблото. Клетките на нишестената вагина (ендодерма) са по-големи и съдържат добре изразени нишестени зърна.
Централният цилиндър в съседство с ендодермата започва с групи от склеренхимни клетки - голи влакна, между които са разположени клетките на паренхима. В основата и в първичната флоема се срещат лубни влакна. Те се различават по много дебели, обикновено нелигинизирани стени, състоящи се от 75 ... 90% целулоза. Дължината на най-голите влакна е 4 ... 60 мм, диаметърът е 12 ... 37 микрона. По време на растежа на склеренхимна клетка нейното ядро \u200b\u200bсе разделя, но цитокинезата не се проявява и протопластът е многоядрен. Клетъчната стена е много удебелена, кухината на клетката е подобна на тесен канал. Клетъчната стена се състои от много слоеве целулоза и е почти лишена от пори. Клетката от зрели влакна е мъртва. Протопласт след сгъстяване на клетъчната стена се унищожава. Лобните влакна имат необичайно висока якост. Якостта на опън в тях съответства на съпротивлението на стоманата, а еластичността е по-висока. Клетките от луковични влакна са много тясно свързани. Лененото влакно е отличен пределен материал, използван в текстилната промишленост.
След това в последователен ред са цилиндрите (пръстените): вторичната флоема, камбият, вторичният ксилем с ясно видими ядрови лъчи. Основният ксилем е разположен в области, съседни на ядрото. Паренхимът на сърцевината бързо се разрушава (в процеса на удължаване на стъблото), образувайки централна кухина.
Стъблата на двудолни тревисти растения се характеризират с:
1. вторична структура, много рано след следващата първична;
2. наличието на първична покривна тъкан - епидермиса;
3. развита първична кора, разделяща се на колленхима (екзодерма), паренхим (мезодерма) и скорбяла вагина (ендодерма);
4. лъчева или без лъчева (плътна) структура;
5. правилното разположение на обезпечаващи или двустранни проводящи снопове в един кръг около периферията на централния цилиндър;
6. наличието на камбий (отворени гроздове),
7. Ядрото е добре развито.
Първичната анатомична структура на стъблото.
Определение и функция на стъблото.
Стъблото е аксиален, обикновено радиално симетричен орган с дълъг апикален (апикален) растеж.
Функции на стъблото:
1. осъществява двупосочно движение на вещества между корените и листата,
2. поддържа короната на растението,
3. насърчава увеличаване на общата асимилационна повърхност на растението поради разклоняване,
4. участва в съхранението на резервни вещества,
5. в млада възраст - извършва и фотосинтеза.
Тези функции определят наличието на вътрешности, добре развити проводящи, механични тъкани и функционално развит паренхим в стъблото.
Структурата на конуса за растеж.
В конуса на растежа на стъблото на покритосеменните растения се изразяват две зони: външна - туника, а вътрешната - жилище които се различават по равнини на клетъчните деления (теория на А. Шмид и Дж. Будър) (фиг. 1).
Туниката може да се състои от един или повече слоеве клетки, които се делят перпендикулярно повърхност, в повечето двусемеделни растения е двуслоен. Външният слой на туниката поражда protoderma,след това се образува епидермиса, покриващ листата и стъблото. Ако туниката е многопластова, тогава вътрешният слой образува тъканите на първичната кора.
При наличието на еднослойна туника тялото образува цялата първична кора и стела. жилищеобразувана от маса от клетки които са разделени във всички равнини.
Зародишите на листата най-често се срещат във втория слой на туниката, а аксиларните пъпки се полагат в тялото.
Фигура 1. б) върха на покритосеменните растения.
В резултат на дейността на първичните меристеми на върха се формира първичната анатомична структура на стъблото: епидермиса, първичната кора, централния цилиндър и сърцевината.
Вторичното сгъстяване възниква поради активността на камбий.
Първичната анатомична структура на стъблото.
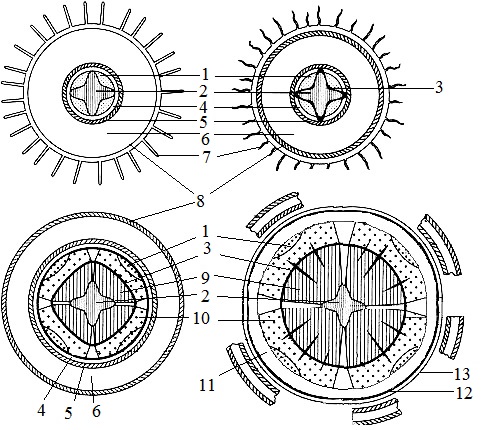
Структурата на стъблото, образувана в резултат на активността на първичните меристеми на върха, се нарича първичен, В стъблото с първичната структура могат да се разграничат три анатомични и топографски зони:
1. покривна тъкан,
2. първична кора
3. Централният цилиндър.
Епидермисът предпазва стъблото от изпаряване.
Под епидермиса се намира външният слой на първичния кортекс - екзодерма -представена от хлоренхим и (в двуколестни) коленхима. Във вътрешния граничен слой с централния цилиндър, ендодерма -нишестените зърна обикновено се натрупват и след това той се превръща в т.нар нишестена вагина (скорбялен слой).Нишестените зърна под влияние на гравитацията могат да се заселят в клетките. Поради това ендодермата играе важна роля в геотропните (от гръцки trpos - въртене, посока), растежните движения на стъблото на растението поради насоченото действие на земната гравитация)
В периферията на централния цилиндър в pericycleразположен склеренхим. Склеренхимните влакна образуват непрекъснат слой или се събират близо до проводими снопове, извън основната флоема. Основният елемент на централния цилиндър (стела) са проводящи снопове. Те се формират от прокамбиални шнурове и се състоят от първичен ксилем и флоема, Диференциацията на прокамбиума е центропетна: флоемата се образува от външната страна на лъча към центъра му, а ксилемата - към него (фиг. 2). Procambium може напълно да се превърне в първични проводящи комплекси, образувайки затворен колатерален сноп (в монокотиледони) или запазени клетки в центъра на снопа образуват камбий. Такива отворени гроздове са характерни за двудолни растения. Работата на камбията осигурява формирането на вторичен ксилем и флоема.
Фигура 2
Полагането на прокамбиум определя структурата на диригентската система:
1. Ако прокамбият образува непрекъснат кухи цилиндър, тогава от него се образуват ксилеми и флоеми цилиндри (в някои иглолистни и двусемеделни); структура без лъч.
2. Ако прокумбията се състои от отделни нишки (за повечето монокотиледони, много двудолни и иглолистни), тогава получените първични проводими тъкани запазват същото структура на лъча.
Прокамбиумът се появява в основата на листния анлаж и неговото развитие протича в две посоки: до върха на листния анлаж и надолу по стъблото, където се свързва с лентите, възникнали по-рано. В същата последователност е разграничаването на флоема и ксилем. Обикновено флоемите се формират по-рано, след това ксилеми. Провеждащите лъчи са разположени в паренхима на централния цилиндър.
В центъра на стъблото се развива ясно видимо ядро. Паренхимните клетки с тънки стени често се разхлабват, умират и се пълнят с въздух, тогава цялата тъкан изглежда бяла (при слънчоглед, царевица, бъз). Понякога ядрото умира много рано, когато разширението на междувъзлия все още не е приключило. Това води до разкъсване на сърцевината и образуване на кухо стъбло.
Първичната структура на стъблото се запазва в едносемеделни растения през целия живот, а в двудолни и гимнастични, само в ранните фази на неговото развитие.
Анатомичната структура на стъблото на двусемеделна структура на растителни лъчи
Стъблото е оста на издънката. Заедно с листото е основната структурна част на издънката. Основните функции са опорни и проводими. Чрез стъблото се осъществява връзка между корените и листата и обменът на продукти от въздушно и минерално хранене. В допълнение, резервните хранителни вещества често се отлагат в стъблото. Стъблото, подобно на целия издънка като цяло, е „отворена“ система за растеж, т.е. тя расте дълго време и върху нея се появяват нови листа.
Стъблото като част от издънката има система от меристеми, поддържащи растежа на тъканите по дължина и дебелина. Растежът в дължина се дължи на апикалните и интеркалярните меристеми, а на дебелината - в двудолни, дължащи се на странични вторични меристеми - камбий и пелоген.
Първичната структура на стъблото се развива, когато клетките на апикалната издънка меристема се диференцират. Апикалната меристема на двусемеделната издънка доста рано се диференцира в няколко групи клетки, които се различават по характеристиките на делене и степента на меристематична активност. Външните му слоеве се трансформират в протодерма, клетките на която по-късно образуват първичната обвивна тъкан - епидермиса.
Фиг. 1. Вид на гредите с междинни камби в стъблото на кирказона (Aristolochia clematitis) в напречно сечение (I) и диаграма на структурата на стъблото на различни нива (II)
А е филийка на нивото на поява на прокамбиум; Б - на нивото на поява на камбий; Б - на нивото на образуваната структура. 1 - прокамбиум, 2 - епидермис, 3 - колленхима, 4 - кортикален паренхим, 5 - ендодерма (3-5 - първичен кортекс), 6 - склеренхим на велосипеда, 7 - флоема, 8 - ксилема, 9 - сноп камбий (7-9 - отворен обезпечителен сноп), 10 - междулъчеви камбий,11 - основен лъч, 12 - паренхим на сърцевината (6-12 - централен цилиндър)
На базовото ниво на първия примордий на листата, апикалните меристемни клетки, разположени към периферията и в центъра на върха, престават активно да се делят, увеличават се по размер и се вакуолизират. От тези клетки се образуват първичната кора и ядрото. Много двудолни в началния пръстен диференцират кръга от изолирани прокамбиални въжета. Клетките на началния пръстен, разположени между тези нишки, по-късно се диференцират в паренхимни елементи. На напречните сечения тези лъчи имат формата на радиални ленти, свързващи ядрото с първичната кора.
Прокамбиумът в редица двудолни също може да бъде положен под формата на непрекъснат пръстен. Той може да се развие по цялата дебелина на началния пръстен или да се образува от неговата част. Procambius е предшественик на първичните проводими тъкани: първичната флоема и първичната ксилема. Флоемата започва да се образува по-рано и се полага във външните части на прокамбиалните шнурове или прокамбиални пръстени. Флоемата се развива центропетално, т.е. първите елементи заемат външното положение, а най-новите - вътрешното. Xylem е положен във вътрешността на Procambius и се развива центробежно. Така флоемата и ксилемата се оформят една към друга. Първите елементи на ксилема са проксимален, тесен, сравнително тънкостенни съдове или трахеиди със спирални или пръстеновидни вторични удебеления. Метаксилемът се образува малко по-късно от протоксилема и се състои от стълбищни и порести съдове. Основният фактор, контролиращ диференциацията както на флоемата, така и на ксилема, е ауксиновият фитохормон, произведен от листни примордии и се движи по прокамбиума от върха към основата. Основната флоема и ксилем, образувани от Прокамбий, съставляват основата на аксиалния цилиндър, или стелата.
Стелата, която заема централната част на стъблото, се състои от проводими тъкани, сърцевина, велосипед и онези постоянни тъкани, които възникват от него. Отвън на рециклирането е първичната кора, която се състои от паренхим, често коленхима и понякога секреторни елементи. Ядрото обикновено се състои от сравнително тънкостенни паренхимни клетки. Резервните хранителни вещества често се отлагат в ядрото. Тук често се срещат идиобласти, т.е. отделни клетки, пълни с таниди, кристали, слуз и др. Понякога част от сърцевината се разрушава и се образува кухина. Периферната част на сърцевината, съседна на ксилемата, се нарича перимедуларна зона.
Вторичното удебеляване на стъблото при повечето двудолни се случва доста рано, което води до формирането на вторичното тяло на растението. Тези промени са свързани най-вече с активността на страничната вторична меристема - камбийът и отчасти друга друга вторична меристема - фалогенът. Поради появата на вторични тъкани, двудолни растения растат в дебелина. Вторичните промени в централния цилиндър започват с полагането на камбий. Камбий възниква от останките на Прокамбий, на границата на първичния ксилем и флоема.
Камбиевите клетки са силно вакуулирани и удължени във вертикална или хоризонтална посока. Първите - фузиформни инициали дават проводящи елементи на проводими тъкани, вторите - инициали - образуват хоризонтално ориентирани радиационни клетки на основните лъчи. Камбиевите начални клетки са способни на два вида деление - периклинално и антиклинално. В първия случай клетъчната плоча се полага успоредно на повърхността на стъблото, във втория - перпендикулярно. Резултатът е непрекъсната серия от производни клетки, простиращи се от камбията радиално навън и навътре. Клетките, отложени към външната повърхност и стъблото, постепенно се диференцират във вторичната флоема, към сърцевината във вторичния ксилем.
Активността на камбий се активира от фитохормоните гъберилин и ауксин, идващи от бъбреците и младите листа. В крайна сметка в структурата на стъблото на годишния издъх на двудолно растение е възможно да се разграничи модифициран централен цилиндър, който включва постоянни тъкани, произтичащи от велосипеда, останките от първичната и вторичната флоема, камбий, вторичните и остатъците от първичния ксилем и сърцевината. Модифицираният централен цилиндър е заобиколен от остатъците от първичната кора.
Вторичен коренКоренът е основният орган на висше растение. Той изпълнява функцията на хранене с минерали и вода. Друга важна функция е фиксирането, „закотвянето“ на растението в почвата. Чрез корена растенията абсорбират вода и йони на минерални соли, разтворени в него от почвата. Корените също извършват биосинтеза на редица вторични метаболити, по-специално алкалоиди. Коренът е способен на метаморфоза. Най-често това е място за съхраняване на резервни хранителни вещества. Понякога той играе ролята на дихателен орган, може да служи като орган на вегетативно размножаване.
Фиг. 2. Преходът от първичната структура на корена към вторичната
Образуването на вторичната структура на корена се свързва преди всичко с активността на камбий, което осигурява растежа на корена в дебелина. Камбият първоначално възниква от тънкостенни паренхимни клетки под формата на фрагментирани участъци от вътрешната страна на флоемовите връзки между лъчите на първичния ксилем. Някои части на велосипеда, които са разположени навън от лъчите на първичната ксилема, скоро придобиват камбиална активност. В резултат на това се формира непрекъснат камбиален слой.
Клетките на вторичния ксилем лежат до центъра на камбия, а клетките на вторичната флоема - към периферията. Клетките на камбия, които са вградени в перикла, образуват широки радиални светлинни лъчи на паренхима, разположени между шнурите на вторичната проводима тъкан. Тези лъчи, понякога наричани първични лъчи на сърцевината, осигуряват физиологична връзка между централната част на корена и първичната кора. По-късно могат да бъдат положени и вторични лъчи, които „свързват“ вторичния ксилем и флоема.
В резултат на активността на камбия първичната флоема се изтласква навън, а „звездата“ на първичния ксилем остава в центъра на корена. Неговите "лъчи" се задържат дълго време, понякога до края на живота на корена. В допълнение към вторичните промени, настъпващи в централния цилиндър, се наблюдават значителни промени в първичната кора. Поради бързия растеж на вторичните тъкани отвътре, причинявайки силно удебеляване на корена, първичната кора често се разкъсва. По това време клетките на велосипеда, разделящи се по цялата обиколка на аксиалния цилиндър, образуват широка зона от паренхимни клетки, във външната част на която е положен фалогенът, поставящ корк отвън, а фалодермата вътре. Коркът изолира първичната кора от проводимите тъкани, кората умира и се изхвърля. Клетките на фалодермата и паренхима, образувани вследствие на велосипеда, впоследствие растат и образуват паренхимната зона, обграждаща проводимите тъкани. Понякога тази зона се нарича вторична кора. Отвън корените на двудолни, които имат вторична структура, са покрити от перидермата. Кората рядко се образува, само по корени на старо дърво.
В ранните стадии на развитие двудолното стъбло има първична структура. В резултат на дейността на първичните меристеми на конуса на растежа се образуват епидермиса, първичната кора, централният цилиндър и сърцевината.
епидермисстъблото има малък брой стомаси, обикновено покрити с кожички.
Под епидермиса се намират тъкани първична кора.Външният му слой често се формира от коленхима, която е разположена или в непрекъснат пръстен (слънчоглед), или в отделни секции. Основната част от първичната кора е съставена от хлоренхим и паренхим. Тук могат да се развият дихателни пътища и контейнери с секрети. Клетките на вътрешния слой на първичната кора обикновено са изпълнени с големи скорбяла на зърна (нишестени влагалища).
Централен цилиндърсе състои от велосипед и проводима система. Ядрото еот паренхима, който често се разрушава и на негово място се образува въздушна кухина. Перикълът е представен от един или повече реда склеренхим или отделните му връзки. Проводимата система има формата на кух цилиндър, който разделя първичната кора и сърцевината. Проводимите снопове, които изграждат този цилиндър, са разделени от междупанелен паренхим, основните лъчи, които свързват сърцевината и първичната кора. Повечето двудолни се характеризират със специален тип централен цилиндър, на напречното сечение на който гредите са подредени в пръстен (фиг.).
Фигура. Стъблото на двудолни растения.
При двудолни растения слой от прокамбиеви клетки между първичната флоема и ксилем запазва способността да се дели и се превръща във вторична образователна тъкан - т.е. камбий, която е разделена чрез поставяне на вътрешни елементи на вторичния ксилем, а към периферията - вторичната флоема. Гроздовете от двудолни растения са отворени, благодарение на работата на камбия, букетът расте, диаметърът му се увеличава. Активността на камбиума осигурява появата на вторични елементи в стъблото, т.е. прехода от първичната към вторичната анатомична структура.
Вторичната структура е характерна за всички двусемеделни и иглолистни растения - треви и дървета. Разнообразие от видове структура се дължи на първо място на местоположението на проводими тъкани, което се определя от полагането на Прокамбий и активността на камбий.
Прокамбиумът в процеса на диференциация на растежния конус се полага под формата на шнурове. Ако шнурите са разделени от достатъчно широки редове паренхим, след това се оформят сноп или преходна структура на стъблото; ако прокамбиевите шнурове са толкова близки, че се сливат в цилиндър, тогава несвързана (твърда) структура.
Гредина структура на стъблото намира се в някои тревисти растения (копър, детелина, пеперуда, грах, люцерна). Прокамбиални шнурове, положени в конус на растеж в един кръг около периферията на централния цилиндър, Всеки прокамбиален шнур се превръща в отворен колатерален сноп, състоящ се от първичен ксилем, първична флоема и ивица камбий между тях. Камбиалните клетки, разделящи се, дават нови (вторични) елементи на проводящия лъч: навътре - ксилема към периферията - флоема. Провеждащите лъчи са разделени от широки междулъчеви зони и обграждат мощно ядро.
В повечето растения камбият работи активно и диаметърът на сноповете значително се увеличава.Активността на камби с лъч стимулира паренхима, който отделя снопчетата, който започва да се дели, давайки вторична образователна тъкан. Снопът и междинните връзки меристеми се сливат и образуват непрекъснат камбиален пръстен, чиято активност осигурява равномерно удебеляване на стъблото. Ако снопът камбий дава елементи от вторичен ксилем и флоема, тогава междукамерен камбий произвежда паренхим (в копър, зърнен зор и т.н.). В детелина, люцерна и други растения междубиещият камбий снася предимно склеренхим-подобни клетки в посока на ксилема. Тези клетки са удължени, дебелостенни лигнифицирани. Следователно забавянето на събирането на детелината намалява качеството на сеното. В тези растения е запазена отчетлива структура на гредите през целия живот на стъблото.
Свързващата структура на стъблото може да бъде заместена с непрекъснато твърдо вещество с възрастта. Преходна структура на стъблотохарактерно за слънчоглед, Първоначално стъблото се оформя по същия начин, както в детелината, но междукамерен камбиум не поставя паренхима, а ксилемата и флоемата. Образуват се допълнителни проводими греди. Постепенно всички снопове могат да се слеят в непрекъснат трислоен (ксилем, камбий, флоем) цилиндър. В горната (млада) част стъблото има структура на гредата, в основата (в старата част) е без куп .
Структура без лъч характерно за стъблата на много билки и дървета. В растежния конус на тези растения прокамбиалните шнурове са толкова близки, че образуват почти непрекъснат цилиндър (в напречното сечение изглежда като пръстен). Диференциращ се, прокамбиалният цилиндър (пръстен) привежда първичния ксиломен цилиндър към центъра, първичният флоелен цилиндър към периферията и между тях се образува камбиален цилиндър.
Вторичните промени са свързани с работата на камбия. Камбиевите клетки се разделят успоредно на повърхността на стъблото, докато 10 ... 20 пъти повече клетки се отлагат навътре, отколкото навън. Вътре камбият образува елементите на вторичния ксилем (дърво), външно вторичната флоема (лапа) се отлага. На някои места камбият полага от двете страни паренхимните клетки на основните лъчи.
Особености на несъбирателната структура на тревните стъбла могат да бъдат разгледани на примера на стъбло лен, Стъблото е покрито с епидермис, под който има хлабав хлоренхим на първичната кора. Голям брой стомаси в епидермиса и отсъствието на колленхима в първичната кора са от съществено практическо значение, тъй като по време на техническото промиване с лен не среща пречки и лесно прониква в стъблото. Клетките на нишестената вагина (ендодерма) са по-големи и съдържат добре изразени нишестени зърна.
Централният цилиндър в съседство с ендодермата започва с групи от склеренхимни клетки - голи влакна, между които са разположени клетките на паренхима. В основата и в първичната флоема се срещат лубни влакна. Те се различават по много дебели, обикновено нелигинизирани стени, състоящи се от 75 ... 90% целулоза. Клетката от зрели влакна е мъртва. Протопласт след сгъстяване на клетъчната стена се унищожава. Лобните влакна имат необичайно висока якост. Якостта на опън в тях съответства на съпротивлението на стоманата, а еластичността е по-висока. Клетките от луковични влакна са много тясно свързани. Лененото влакно е отличен пределен материал, използван в текстилната промишленост.
След това в последователен ред са цилиндрите (пръстените): вторичната флоема, камбият, вторичният ксилем с ясно видими ядрови лъчи. Основният ксилем е разположен в области, съседни на ядрото. Паренхимът на сърцевината бързо се разрушава (в процеса на удължаване на стъблото), образувайки централна кухина.
Стъблата на двудолни тревисти растения се характеризират с:
1. вторична структура, много рано след следващата първична;
2. наличието на първична покривна тъкан - епидермиса;
3. развита първична кора, разделяща се на колленхима (екзодерма), паренхим (мезодерма) и скорбяла вагина (ендодерма);
4. лъчева или без лъчева (плътна) структура;
5. правилното разположение на обезпечаващи или двустранни проводящи снопове в един кръг около периферията на централния цилиндър;
6. наличието на камбий (отворени гроздове).
Бягство от метаморфози.
Ако допълнителни функции започнат да надделяват над основните или орган придобие някои нови функции в процеса на развитие, той се променя много морфологично и анатомично. Такива модификации на органите се наричат метаморфози(от гръцки. метаморфоза - преобразуване, трансформация). Извикват се органи с един и същ произход, но изпълняващи различни функции хомологичен . Хомологичните органи са коренища, грудки, луковици, тръни от глог, кладода, филокладий, сухожилие на грозде и тиква, както всички те са исторически развити от листно-стъблена издънка.
Наричат \u200b\u200bсе органи, които изпълняват едни и същи функции и са сходни морфологично, но с различен произход сходен , Коренови шишарки и стрелящи грудки, стреляйте шипове на глог и листни шипове на берберис, листни пъпки на грах и издънки на издънки от тиква са подобни.
Модификации на подземни издънки.
Подземните издънки са широко разпространени в природата: коренища, грудки, костури, луковици. Те участват в доставката на вещества и вегетативното размножаване на растенията.
коренищее бягство с люспести листа, пъпки и корени на придатка. На повърхността на коренището ясно се виждат белези от миналогодишните долни листа и мъртви издънки. Кореневото има много подчинени корени. Обикновено подземни коренища. Бъбреците се развиват във въздушни издънки. Ризомите са характерни за житна трева, ирис, водни лилии.
ластун - удължен тънък издънка с недоразвити листа. За разлика от коренищата, той обикновено е краткотраен. В мускат, кисели оксиди, столони участват в натрупването на хранителни вещества, отложени в много уплътнени низове, но основната функция на столона е да участва във вегетативно размножаване.
грудказа разлика от коренището и столона, той значително се скъсява и уплътнява. Резервните вещества в него са локализирани в стволовия паренхим.
Клубените могат да се развият върху коренища, столони, основната издънка и други части на растенията. Те могат да бъдат подземни и надземни.
Мощните коренищни грудки с много нишесте са характерни за таро или ядлива колоказия от семейството на ароидите, което се култивира в тропически и субтропични страни.
в картоф клубените се образуват в края на подземните столончета - белезникави крехки издънки с люспести листа, развиващи се от аксиларни пъпки, разположени в основата на листните леторасти. На повърхността на грудката са разположени аксиларни бъбреци - очиседнали в малки ями, рамкирани от едната страна вежди -останките от основите на листата.
грудки ерусалимски артишокили земляна крушасе образуват и върху столоните. Проводимите тъкани в тях са слабо развити. Грудките са богати на инулин, който се отлага главно в паренхима на стъблото.
Клубените, които се образуват на столони и коренища, са краткотрайни. Те продължават само една година, между два вегетационни периода, отмират след развитието на листни издънки от пъпките си, консумират хранителни вещества, докато се образуват корените, а растенията не преминават към самостоятелно хранене.
крушка, подобно на грудката, тя е специализирана, модифицирана съкратена издънка, която служи не само за съхраняване на хранителни вещества, но и за пренасяне на неблагоприятни периоди от годината, вегетативно обновяване и размножаване на растенията. Луковиците са характерни за едносемеделни растения от семейства лилии, амарилиси и други. Като изключение те се срещат в двудолни: някои видове кисело и джинджифил.
Извика стъбло дъното,в луковицата е силно намалена и повече или по-малко сплескана. Тя носи листа под формата на люспи, а в долната част - принадлежащи корени. За разлика от коренищата и грудките, месестите люспи служат като органи за съхранение на луковицата. Луковиците се оформят в лале, лешник, нарцис, зюмбюл, синя клечка, кокиче и други растения.
В зависимост от развитието на страничните бъбреци са луковиците просткогато бъбреците са малки (лук, лилии и т.н.), и комплекс - бъбреците в синусите на сухите люспи са големи, силно развити (чесън).
Луковицата- Подземна модификация на издънката, съчетаваща признаците на коренища и луковици. Развива се от издънката със силно съкратено стъбло, от долната част на което отклоняват адвентивните корени. В горната част на кората или в основата му е разположен бъбрек, от който се формира издънка с цветя. Стъблото на кората е покрито с основи на мъртви миналогодишни листа, които приличат на сухи мембранозни люспи. От бъбреците в синусите им се развиват нови корички - деца. Коремите имат шафран, гладиолус, колхикум.
Модификации на надземните издънки
Глава навънзелето е по същество бъбрек с гигантски размери: месото стъбло заема средната част на главата на зелето ( пънове), по протежение на която са разположени сочни месести листа. Най-малките и най-младите от тях са вътре в главата на главата, около конуса на растежа, който завършва с пънчето. Брюкселското зеле произвежда малки глави зеле от страничните пъпки.
В главата на зелето и двата органа за бягство участват в натрупването на хранителни вещества: водоразтворими захари и други биологично ценни съединения, включително витамини, но основната роля играят листата.
Надземни грудки, развиваща се от дъното на основната издънка, характерна за зеленото зеле.
Малки възли могат да се развият и в областта на съцветие. В живописния хайландър например се образуват в синусите на долните покриващи листа на съцветието. Пъпките им започват да покълват вече по майчиното растение. След като паднат, те се вкореняват, образувайки нови растения.
Метаморфозираните издънки могат да служат не само за съхраняване на хранителни вещества, размножаване и размножаване на растенията, но и да изпълняват други функции.
При някои растения издънките по време на процеса на развитие губят листата си и стъблото се превръща в фотосинтезиращ орган, наречен в този случай platyclade . Обикновено тя е сплескана и има способността да расте дълго време (в тропическо растение, мулбенекия или хомалокладус от семейство елда, бодлива круша). В възлите стесненията са добре изразени, така че кладода изглежда съчленен. При влажни условия кладодите могат да бъдат зеленина, но при сухо време листата окапват. Като орган на фотосинтеза, кладоди има добре развита тъкан, носеща хлорофил, разположена под епидермиса.
Cladody, на външен вид подобен на листа, се нарича phylloclades , Те се развиват от страничните пъпки, така че винаги са в пазвите на малко мембранозно или люспесто листо. Филокладиите са много разнообразни. При аспержите пенисти или настръхнали, те са малки, с игловидна форма, в аспержи Sprenger, или гъсто цъфтящи, те са сплескани, линейни. Кожени ланцетни или яйцевидни очертания кожени фалокладии са характерни за иглените видове, открити в Средиземноморието, Закавказието и Крим. През лятото от долната страна на филокладия в средната му част се развива съцветие, разположено в пазвата на малкия покривен лист, през зимата и зимата иглата цъфти, а през пролетта оранжевите плодове узряват на филокладията.
Доста често срещан вид смяна на издънки е трън , Това е много буйно съкратено издънка без листа с остър връх. При глог, издънка, която се развива през пролетта в листния синус, първо има малки листа и апикална пъпка. Скоро апикалният растеж престава, листата окапват, стъблото е много лигнифицирано и се превръща в трън. Развиват се и тръните на завоя.
Прилепналите растения са оборудвани мустак , които, подобно на бодли, са в някои растения модифицирани странични издънки. Passiflora имат прости, неразклонени антени, с усукан връх. Те са в аксилите на листата, като цветя. Разклонените пъстрички на гроздето са модифицирани съцветия. При момичешкото грозде краищата на разклонените антени са с форма на диск и играят ролята на вендузи, за да помогнат на растението да се изкачи до опората. Антените с произход на издънки също се формират в тикви, дини, краставици и други представители на тиквеното семейство.