Reduction in the number of gills.
Increase in respiratory surface due to the formation of gill filaments.
Formation of gill capillaries.
In the lancelet, the lateral walls of the pharynx are pierced by numerous (up to 150 pairs) obliquely located gill slits. The afferent branchial arteries approach the interbranchial septa, and the efferent branchial arteries depart. When water washes the interbranchial septa, gas exchange occurs between the passing water and the blood that flows through the thin vessels of the septa. The branchial arteries do not branch into capillaries. In addition, oxygen enters the animal’s body through the capillaries of the skin.
In proto-aquatic vertebrates (jawless and fish), as well as in lower chordates, gill slits are formed that connect the pharyngeal cavity with the external environment. In cyclostomes, gill sacs are formed from the endoderm lining the gill slits (in fish, gills develop from ectoderm). The inner surface of the bags is covered with numerous folds - gill filaments, in the walls of which a dense network of capillaries branches. The bag opens with an internal narrow channel into the pharynx (in adult lampreys - into the respiratory tube), and with an external one - on the lateral surface of the animal’s body. Hagfish have from 5 to 16 pairs of gill sacs; in the bdellostomidae family, each of them opens outward with an independent opening, and in the hagfish family, all external gill passages on each side merge into one canal, which opens outward with one opening located far behind. Lampreys have 7 pairs of gill pouches, each of which opens outwards with an independent opening. Breathing is carried out through rhythmic contractions and relaxations muscle wall gill region. In non-feeding lampreys, water enters the respiratory tube from the oral cavity, then washes the lobes of the gill sacs, providing gas exchange, and is removed through the external gill passages. In feeding cyclostomes, water enters and exits through the external openings of the gill sacs.
The respiratory system of fish has specialized gas exchange organs - ectodermal gills, which are either located on the interbranchial septa, as in cartilaginous fish, or directly extend from the gill arches, as in bony fish. The exchange of gases in the gills of vertebrates is built according to the type of “countercurrent systems”: during counter-flow, the blood comes into contact with oxygen-rich water, which ensures its effective saturation. An increase in the oxygen absorption surface due to the formation of gills was accompanied by a decrease in the number of gill slits in vertebrates compared to lower chordates. In whole-headed fish (from cartilaginous fish), a reduction in the interbranchial septa is observed and a leathery gill cover is formed, covering the outside of the gills. In bony fishes, a bony skeleton appears in the gill cover, and the intergill septa are reduced, which contributes to more intensive washing of the gill filaments with water. Along with gas exchange, fish gills participate in water and salt exchange, and in the removal of ammonia and urea from the body. The skin, swim bladder, suprapharyngeal labyrinths and specialized sections of the intestinal tube function as additional respiratory organs in certain groups of fish. Lungfishes and multi-feathered fish develop air breathing organs - lungs. The lungs arise as paired outgrowths of the abdominal part of the pharynx in the region of the last gill slit and are connected to the esophagus by a short canal. The walls of this outgrowth are thin and abundantly supplied with blood.
Directions of evolution of pulmonary type of breathing
Emergence and differentiation of the respiratory tract.
Differentiation of the lung and increase in respiratory surface.
Development of accessory organs ( chest).
In amphibians, the following are involved in the absorption of oxygen and the release of carbon dioxide: in larvae - the skin, external and internal gills, in adults - the lungs, skin and mucous membrane of the oropharyngeal cavity. In some species of tailed amphibians (sirens, proteas) and in adults, the gills are retained and the lungs are underdeveloped or reduced. The ratio of pulmonary and other types of gas exchange is not the same: in species of humid habitats, skin respiration dominates in gas exchange, in inhabitants of dry places most of oxygen enters through the lungs, but the skin plays a significant role in the release of carbon dioxide. The respiratory system of adult amphibians includes the oropharyngeal, laryngeal-tracheal cavities and sac-like lungs, the walls of which are intertwined with a dense network of capillaries. Tailless amphibians have a common laryngeal-tracheal chamber; in caudate amphibians, it is divided into the larynx and trachea. The arytenoid cartilages appear in the larynx, which support its wall and vocal cords. The lungs of tailed amphibians are two thin-walled bags without partitions. In tailless animals, inside the lung sacs there are partitions on the walls that increase the surface of gas exchange (cellular lungs). Amphibians do not have ribs, and the act of breathing occurs by pumping air during inhalation (due to an increase and then decrease in the volume of the oropharyngeal cavity) and pushing out air during exhalation (due to the elasticity of the walls of the lungs and abdominal muscles).
In reptiles, there is further differentiation of the respiratory tract and a significant increase in the functional surface of gas exchange in the lungs. The airways are divided into the nasal cavity (it is combined with the oral cavity, but in crocodiles and turtles these cavities are separated by the bony palate), the larynx, the trachea and two bronchi. The walls of the larynx are supported by paired arytenoid and unpaired cricoid cartilages. In lizards and snakes, the inner walls of the lung sacs have a folded cellular structure. In turtles and crocodiles a complex system The septum protrudes into the internal cavity of the lung so deeply that the lung acquires a spongy structure. The chest is formed: the ribs are movably connected to the spine and sternum, the intercostal muscles develop. The act of breathing is carried out due to a change in the volume of the chest (costal type of breathing). Turtles retain the oropharyngeal type of air injection. In aquatic turtles in water, additional respiratory organs are the capillary-rich outgrowths of the pharynx and cloaca (anal bladders). Reptiles do not have cutaneous respiration.
In birds, the airways are represented by the nasal cavity, the larynx, which is supported by the arytenoid and cricoid cartilages, the long trachea and the bronchial system. The lungs are small, dense and poorly extensible, attached to the ribs on the sides of the spinal column. The primary bronchi are formed by dividing the lower part of the trachea and enter the tissue of the corresponding lung, where they break up into 15–20 secondary bronchi, most of which end blindly, and some communicate with the air sacs. The secondary bronchi are interconnected by smaller parabronchi, from which many thin-walled cellular bronchioles arise. The bronchioles, entwined with blood vessels, form the morphofunctional structure of the lung. Associated with the lungs of birds are air sacs - transparent, elastic, thin-walled outgrowths of the mucous membrane of the secondary bronchi. The volume of the air sacs is approximately 10 times the volume of the lungs. They play a very important role in the implementation of the peculiar respiratory act of birds: air with a high oxygen content enters the lungs both during inhalation and exhalation - “double breathing”. In addition to intensifying breathing, air bags prevent the body from overheating during intense movement. An increase in intra-abdominal pressure during exhalation promotes defecation. Diving birds, by increasing the pressure in the air sacs, can reduce the volume and thereby increase the density, which makes it easier to dive into water. There is no skin respiration in birds.
In mammals, further differentiation of the respiratory tract is observed. The nasal cavity, nasopharynx is formed, the entrance to the larynx is covered by the epiglottis (in all terrestrial vertebrates except mammals, the laryngeal fissure is closed by special muscles), thyroid cartilage appears in the larynx, then comes the trachea, which branches into two bronchi going into the right and left lungs. In the lungs, the bronchi branch repeatedly and end in bronchioles and alveoli (the number of alveoli is from 6 to 500 million), this significantly increases the respiratory surface. Gas exchange occurs in the alveolar ducts and alveoli, the walls of which are densely intertwined with blood vessels. The morphofunctional unit of the mammalian lung is the pulmonary acinus, which is formed as a result of the branching of the terminal bronchiole. The chest is formed, which is separated by the diaphragm from the abdominal cavity. The number of respiratory movements is from 8 to 200. Respiratory movements are carried out in two ways: due to changes in the volume of the chest (costal breathing) and due to the activity of the diaphragmatic muscle (diaphragmatic breathing). Higher mammals have developed cutaneous respiration through a system of skin capillaries, which plays an important role in gas exchange.
The evolution of breathing.
1) Diffuse breathing- This is the process of equalizing the concentration of oxygen inside the body and in its environment. Oxygen penetrates the cell membrane in single-celled organisms.
2) Skin breathing- this is the exchange of gases through the skin in lower worms and in vertebrates (fish, amphibians), which have special respiratory organs.
Gill breathing
PINUS GILLS(skin outgrowths on both sides of the body) appear in marine annelids, aquatic arthropods, and in mollusks in the mantle cavity.
GILLS- respiratory organs of vertebrate animals, formed as invaginations of the digestive tube.
In the lancelet, the gill slits penetrate the pharynx and open into the circumbranchial cavity with frequent changes of water.
Fish have gills from gill arches with gill filaments pierced by capillaries. The water swallowed by the fish enters the oral cavity, passes through the gill filaments to the outside, washes them and supplies the blood with oxygen.

4) Tracheal and pulmonary breathing- more effective, since oxygen is absorbed directly from the air, and not from water. Characteristic of terrestrial mollusks (sac-like lungs), arachnids, insects, amphibians, reptiles, birds, and mammals.
Arachnids have lung sacs (scorpions), tracheas (mites), and spiders have both.

INSECTS have tracheas - the respiratory organs of terrestrial arthropods - a system of air tubes that open with respiratory openings (stigmas) on the lateral surfaces of the chest and abdomen.
AMPHIBIANS They have 2/3 cutaneous respiration and 1/3 pulmonary respiration. The airways appear for the first time: larynx, trachea, bronchial rudiments; lungs are smooth-walled bags.
REPTILES have developed airways; the lungs are cellular, there is no skin respiration.
BIRDS have developed airways, spongy lungs. Some of the bronchi branch outside the lungs and form air sacs.

Air bags- air cavities connected to the respiratory system, 10 times larger than the volume of the lungs, serving to enhance air exchange in flight, do not perform the function of gas exchange. Breathing at rest is carried out by changing the volume of the chest.
Breathing in flight:
1. When the wings rise, air is sucked through the nostrils into the lungs and rear air sacs (I gas exchange in the lungs);
Front air bags← light - rear air bags
2. When the wings lower, the air sacs are compressed, and air from the rear air sacs enters the lungs (II gas exchange in the lungs).
Front air bags - light ← rear air bags
Double Breathing- This is the exchange of gases in the lungs during inhalation and exhalation.
MAMMALS- gas exchange is almost entirely in the lungs (through the skin and alimentary canal -2%)

Airways: nasal cavity → nasopharynx → pharynx → larynx → trachea → bronchi (bronchi branch into bronchioles, alveolar ducts and end with alveoli - pulmonary vesicles). The lungs have a spongy structure and consist of alveoli intertwined with capillaries. The respiratory surface is increased 50-100 times compared to the surface of the body. The type of breathing is alveolar. The diaphragm, which separates the chest cavity from the abdominal cavity, as well as the intercostal muscles, provide ventilation to the lungs. Complete separation of the oral and nasal cavities. Mammals can breathe and chew at the same time.
The set of processes that ensure the consumption of O 2 and the release of CO 2 in the body is called breathing. There are processes of external and internal respiration. External respiration ensures the exchange of gases between the body and the external environment, internal respiration ensures the consumption of O2 and the release of CO 2 by the cells of the body.
The factor ensuring the diffusion of gases through the respiratory surfaces is the difference in their concentrations. The movement of dissolved gases occurs in the direction from an area of high concentration to an area of low concentration.
U small organisms Gas exchange, as a rule, occurs diffusely over the entire surface of the body (or cell). In larger animals, gases are transported to the tissues either directly (the tracheal system of insects) or through special vehicles (blood, hemolymph).
The amount of oxygen entering the animal’s tissues depends on the area of the respiratory surface and the difference in oxygen concentration on them. Therefore, growth of the respiratory epithelium is observed in all respiratory organs. To maintain a high gradient of oxygen diffusion on the exchange membrane, movement of the medium (ventilation) is necessary. It is provided by the respiratory rhythmic movements of the entire body of the animal (the oligochaete worm, leeches) or certain parts of it (crustaceans), as well as the work of the ciliated epithelium (molluscs, lancelet).
A number of fairly large animals do not have specialized respiratory organs. In them, gas exchange is carried out through moist skin, equipped with an abundant network of blood vessels (earthworm). Cutaneous respiration as an additional method is characteristic of animals with specialized respiratory organs. For example, in eels that have gills, 60% of their oxygen requirement is met through skin respiration; in frogs that have lungs, this value is more than 50%.
The respiratory organs in the aquatic environment are gills, in the land-air environment - the lungs and trachea.
Gills are organs located outside the body cavity in the form of epithelial surfaces penetrated by a dense network of blood capillaries. Gill respiration is characteristic of polychaete annelids, most mollusks, crustaceans, fish, and amphibian larvae. Gill respiration is most effective in fish. It is based on counterflow phenomenon: Blood in the capillaries of the gill filaments flows in the direction opposite to the flow of water washing the gills.
Lungs, as a rule, are internal organs and are protected from drying out. There are two types: diffusion And ventilation. In the first type of lung, gas exchange occurs only by diffusion. Relatively small animals have such lungs: pulmonary mollusks, scorpions, spiders. Only terrestrial vertebrates have ventilation lungs.
The complication of the structure of the lungs in the series from amphibians to mammals is associated with an increase in the area of the respiratory epithelium. Thus, in amphibians, 1 cm 3 of lung tissue has a total gas exchange surface of 20 cm 2. A similar indicator for human lung epithelium is 300 cm 2.
Simultaneously with the increase in the respiratory surface, the mechanism of lung ventilation is improved, which, starting with reptiles, is carried out due to changes in the volume of the chest, and in mammals - with the participation of the diaphragm muscles. These adaptations allowed warm-blooded animals (birds and mammals) to dramatically increase their metabolic rate.
The third type of respiratory organs is trachea. They are air-filled, thin-walled, branching, non-collapsing invaginations into the body. The trachea communicates with the external environment through openings in the cuticle - spiracles. In insects there are most often 12 pairs of them: 3 pairs on the chest and 9 pairs on the abdomen. The spiracles can close or open depending on the amount of oxygen. At high degree development of the tracheal system (in insects), its numerous branches intertwine everything internal organs And directly provide gas exchange in tissues. The fundamental difference between tracheal breathing and pulmonary and gill breathing is that it does not require the participation of blood as a transport intermediary in gas exchange.
The tracheal system is capable of supporting sufficient high level tissue respiration, thereby ensuring high physiological activity of the insect.
Ventilation of the trachea in insects in the absence of flight is most often carried out by rhythmic contractions of the abdomen, and during flight it is enhanced by movements of the chest.
The aquatic larvae of some insects breathe using tracheal gills. In this case, the tracheal system is devoid of spiracles, i.e. it is closed and filled with air. The branches of the closed tracheal system extend into the “gills” - appendages with a large surface and a thin cuticle that allows gas exchange between water and air of the tracheal system. Such tracheal gills are found, for example, in mayfly larvae. In the larvae of some dragonflies, the tracheal gills are located in the rectal cavity, and the insect ventilates them by drawing water into the intestine and pushing it back out.
Evolution of the respiratory system
Stages of the breathing process
Breath– a set of processes that ensure the supply of oxygen to the body from environment, necessary for oxidation organic matter in the mitochondria of the cell, and the release of carbon dioxide
Types of breathing:

Breathing type:
Cellular.
Organisms: single-celled animals (amoeba, green euglena, slipper ciliates); coelenterates (jellyfish, coral polyps); some worms.
Single-celled organisms absorb oxygen dissolved in water over the entire surface of the body by diffusion.
Oxygen is involved in the breakdown of complex organic substances, resulting in the release of energy that is necessary for the life of the animal.
Formed by respiration carbon dioxide is also secreted out through the entire surface of the body.
Tracheal breathing is breathing using a system of united tracheal tubes that permeate the entire body.
Organisms: class Insects (beetles, butterflies, grasshoppers, flies)
The insect's abdomen is divided into 5–11 parts (segments). Each of them has a pair of small holes - spiracle. Branching tubes extend inward from each spiracle - trachea, which permeate the entire body of the insect. Watching the cockchafer, you can notice how its abdomen either decreases in volume or increases in size. These are breathing movements. When you inhale, air containing oxygen enters the body through the spiracles, and when you exhale, air saturated with carbon dioxide comes out.
In spiders (class Arachnids), the respiratory organs are represented not only by tracheas, but also by pulmonary sacs, which communicate with the external environment through the respiratory openings.
Gill respiration is breathing using specialized structures with a dense network of blood vessels.
Organisms: many aquatic inhabitants (fish, crayfish, mollusks)
Fish breathe oxygen dissolved in water with the help of special branched skin outgrowths called gills.
Fish constantly swallow water. From the oral cavity, water passes through the gill slits, washes the gills and comes out from under the gill covers. Gills consist of gill arches And gill filaments, which are penetrated by many blood vessels. From the water that washes the gills, oxygen enters the blood, and carbon dioxide is removed from the blood into the water. The gills located inside the body are called internal gills.
Some animals, such as amphibians, have dense tufts of gills on the surface of their bodies. These gills are called - external. This is the structure of Proteus, a blind cave animal from the western regions of Yugoslavia, and axolotls (which are similar in general appearance to newts) - their homeland is Mexico.
Amphibians have two types of respiratory organs (not counting the skin): gills and lungs. The weakening of gill respiration and the emergence of pulmonary respiration is already observed in Dipnoi; changes in this direction are observed in Polypterus and Lepidosteus. In amphibians, gill breathing is retained primarily in larvae, and then in those Urodela that spend their entire lives in water (Perennibranchiata in previous systems). Gill slits are inherited by amphibians from fish-like ancestors. Gill arches are found in stegocephalians, larvae and some adults (Branchiosauridae). All modern amphibians in the larval state breathe through gills. Normally, they develop 5 visceral sacs and a 6th underdeveloped one. But not all of them open outward: 4 or even fewer gill slits are formed. Sometimes there are significantly fewer gaps than arcs. The presence of cracks and arcs is evidence of the origin of amphibians from fish. Internal gills, homologous to the gills of fish, are found, however, only in Anura larvae in the form of short outgrowths of integument on the arches separating the gill slits. The gill region is covered from the outside with a soft gill operculum, growing from the side of the hyoid arch. The gill covers of the right and left sides merge with each other on the lower side, leaving paired openings in some Anura, and one unpaired one on the left side of the body in most.
In the early stages of development, the larvae of Anura and all other amphibians have only external gills, apparently homologous to the external gills of the larvae of Polypterini and Dipnoi. In Apoda and Anura, external gills exist only in the larval period, in the early stages of development, while in Urodela, which have returned to aquatic life for the second time, they remain throughout life. Hence the name for these amphibians is Perennibranchiata, although this name, as has been said, embraces groups of amphibians of different origins. External gills are probably inherited by amphibians from lobe-finned fish.
Light amphibians look like long cylindrical bags with thin walls (in Urodela) or shorter ones (in Anura). In legless people, the right lung is much more developed than the left. Lungs appeared in the ancestors of tetrapods long before they set foot on land. We see the same lungs in lungfishes. They apparently appeared as an additional respiratory organ due to the insufficient development of gill respiration, on the one hand, and possibly unfavorable conditions for breathing in dry and spoiled waters, on the other. The posterior part of the gill cavity has developed into an additional respiratory organ. Initially, this organ, which had the appearance of a bilobed sac that opened on the underside of the pharynx, was imperfect: its walls should have been thin, although abundantly supplied with blood, with poorly developed or almost undeveloped septa. Like all gill protrusions (slits), it had smooth visceral muscles and was innervated by the vagus first.
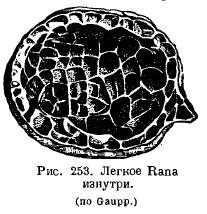
The lungs of amphibians have changed little in comparison: in aquatic Urodela, the lungs serve rather as a hydrostatic apparatus and have a smooth internal surface; their height of organization is even lower than that of Dipnoi. Normally, in amphibians, the inner surface of the lungs is cellular due to the fact that a system of crossbars protrudes into the lung cavity (Fig. 253). It is very interesting that the more terrestrial a particular species is, the more developed the crossbars in the lungs are: in a toad the lung is more cellular than in frogs. In the genus Ascaphus, living in mountain streams, in water rich in oxygen, skin respiration is highly developed, while the lungs, on the contrary, are small and poorly supplied with blood. A number of amphibians from the suborder Salamandroidea (Salamandrina, Plethodon, Spelerpes, Batrachoseps, etc.) completely lost their lungs, in replacement of which pharyngeal and cutaneous respiration developed greatly. .
In the simplest case, the pulmonary sacs are connected to each other in front, opening directly into the pharynx with a longitudinal slit supported on the sides by cartilaginous strips. These cartilaginous strips, with the help of muscles attached to them, can expand and narrow the laryngeal opening.
These cartilages originate from the last gill arch and are found in their simplest form in some Urodela. Cartilages called cricoid cartilages can separate from these cartilages. They can be compared with the arytenoid cartilages (cartilagines arythenoidea) of higher vertebrates. Some Urodela, as well as Apoda, have a rather long windpipe supported by cartilaginous rings. In Anura, the mucous membrane in the larynx forms the vocal cords. The larynx has complex muscles. At the bottom or corners of the mouth there are resonators that inflate when croaking.
The breathing mechanism of terrestrial amphibians is rooted in reflexes observed in fish and aquatic amphibians. Closest to the respiration of fish is the respiration of Anura larvae, which have internal gills, an opercular fold and a gill cavity formed by their fusion, which opens outward with one opening. In addition, in amphibian larvae the oral cavity is abundantly supplied with blood. By taking water into the mouth and pushing it by lifting the jaws through the nostrils, the larvae increase gas exchange in the oral cavity. When the larvae grow, they rise to the surface, where they swallow air like ceratodes, and by raising the bottom of the oropharyngeal cavity push air into the lungs. A similar act is observed in aquatic Urodela. When the bottom of the oropharyngeal cavity is lowered and the gill openings are closed at the back, water is sucked into the oral cavity through the mouth or nostrils or both. By subsequently raising the floor of the mouth with the nostrils closed, water is pushed out through the gill slits. Thanks to these movements, the mucous membrane of the mouth and pharynx comes into contact with new masses of water, and the gills experience a movement that renews the respiratory environment.
In terrestrial amphibians, the breathing mechanism is the act of swallowing air due to the lowering of the muscular floor of the oral cavity and pushing it into the lungs due to the raising of the floor. Thus, the respiration of terrestrial amphibians is an act carried out according to the type of pressure pump that predominates in lower fish. The immediate basis on which it develops is the breathing mechanism of permanent branchial amphibians. This latter, observed, for example, in Necturus, must have evolved in the distant fish-like ancestors of amphibians. From it a more complex type of terrestrial breathing has already developed - Anura.
In lungless salamanders, gas exchange in the intraoral and pharyngeal cavities is highly developed, which occurs with the help of frequent, up to 120-170 vibrations per minute of the mouth diaphragm (in frogs there are 30).
In general, it should be said that pulmonary respiration in amphibians in general is an auxiliary method of breathing. This also indicates its phylogenetic origin.
The breathing of modern amphibians could in no way be the source of the development of breathing in higher Tetrapoda (breathing by raising the ribs, expanding the chest and thus sucking in air). The latter type could have arisen, or at least emerged, among the most ancient extinct amphibians that had long ribs.