Individuálny rozvoj(ontogenéza), periodizácia ontogenézy.
Všetky organizmy majú špecifický životný cyklus. U organizmov vyvíjajúcich sa pohlavne to začína od okamihu objavenia sa zygoty a končí prirodzenou smrťou organizmu.
Súhrn procesov, ktoré sa vyskytujú počas životného cyklu organizmov, je definovaný ako individuálny vývoj alebo ontogenéza.
Ontogenéza zahŕňa 3 obdobia:
1 obdobie. Preembryonálna alebo gametogenéza.
2 obdobie. Embryonálne.
3 obdobie. Postembryonálne.
1. Životné cykly organizmov. Vývoj je larválny a priamy.
Vývoj organizmov je priamy a nepriamy s premenou.
nepriamy vývoj prebieha cez larválne štádium. V larve sa vytvárajú určité zárodočné alebo dočasné orgány, ktoré zabezpečujú životnú aktivitu organizmu v tomto štádiu vývoja.
U vyšších stavovcov je vývin priamy, no provizórne orgány vznikajú aj počas embryonálneho vývoja. U cicavcov sú to embryonálne membrány (amnion, chorion, allantois, placenta) a žĺtkový vak.
2. Predembryonálne obdobie (preembryonálne, progenéza) vývoja. Etapy gametogenézy. Zmeny v oogenéze spojené so skorým vývojom embrya (amplifikácia génov, ooplazmatická segregácia, akumulácia živín).
Preembryonálne obdobie alebo gametogenéza zahŕňa niekoľko štádií: izoláciu, reprodukciu, rast, dozrievanie, tvorbu (posledné len v spermiách).
Počas oogenézy dochádza k najdôležitejším udalostiam, ktoré sú nevyhnutné pre vývoj budúceho organizmu.
1 udalosť. Počas oogenézy dochádza k amplifikácii rRNA génov alebo k zvýšeniu počtu kópií génov zodpovedných za rRNA. K tomuto procesu dochádza v profáze meiózy 1. Kópií génov rRNA môže byť až milión.
Potom sa tieto kópie oddelia od chromozómov, voľne plávajú v karyoplazme, okolo nich sa vytvoria jadierka a v jadierkach sa syntetizujú podjednotky ribozómov, ktoré vstupujú do cytoplazmy. Vo vajíčku sa teda počet ribozómov vopred prudko zvyšuje.
2 udalosť. Počas oogenézy sa počas profázy meiózy 1 syntetizujú rôzne typy mRNA. Transkripčné procesy prebiehajú na despiralizovaných úsekoch chromozómov. Chromozómy v profáznom štádiu meiózy 1 sa nazývajú chromozómy lampovej kefky.
3 udalosť.Živiny sú vo vajci uložené vo forme žĺtka.
4 udalosť. Vajíčko sa vyznačuje ooplazmatickou segregáciou, to znamená distribúciou látok v cytoplazme vajíčka, čo vedie k chemickej heterogenite cytoplazmy. Predpokladá sa, že je to nevyhnutné pre skorú diferenciáciu buniek.
5 udalosť. Pohlavné bunky sú špeciálne bunky tela, pretože majú totipotenciu, to znamená rovnakú dedičnosť. Iba zárodočné bunky, ako aj blastoméry u ľudí, v štádiu 2 blastomérov vedú k vzniku všetkých typov buniek. Napríklad experimenty so separáciou, zostrihom alebo zmiešaním blastomérov v štádiu štiepenia ukázali, že u druhov s radiálnym typom štiepenia vykazujú blastoméry niekoľkých generácií, ak sú izolované a umiestnené vo vhodných podmienkach, totipotenciu, t.j. vyvinie sa do kompletného organizmu. Pre rovnakú dedičnosť a totipotenciu buniek ľudských embryí až do štádia 2-4 blastomérov hovoria prípady narodenia dvoch, troch, štyroch jednovaječných dvojčiat.
3. Embryonálne obdobie vývoja, periodizácia.
Embryonálne obdobie ontogenézy zahŕňa niekoľko fáz:
1 stupeň oplodnenia.
2. štádium zygoty.
3 Štádium drvenia (tvorba jednovrstvového embrya).
4 Štádium gastrulácie (tvorba dvoj- a trojvrstvového embrya).
5 Etapa histo- a O rganogenéza (tvorba tkanív a orgánov).
4. Hnojenie a tvorba zygoty. Vlastnosti oplodnenia u cicavcov a ľudí.
štádium oplodnenia, toto je proces fúzie vajíčka a spermie, v dôsledku čoho sa vytvorí diploidná zygota, z ktorej sa vyvinie diploidný organizmus. V tomto procese existujú 3 fázy:
1 etapa- konvergencia gamét. V tomto zohrávajú dôležitú úlohu látky, ktoré sú vylučované vajíčkom a spermiou. Nazývajú sa gamóny (hormóny gamét, gynogamónov a androgamonov). Okrem toho existuje množstvo nešpecifických faktorov, ktoré zvyšujú pravdepodobnosť stretnutia a interakcie spermie s vajíčkom. Tie obsahujú
koordinácia nástupu pripravenosti na oplodnenie u mužov a žien,
správanie samcov a samíc, ktoré zabezpečuje kopuláciu a insemináciu,
nadmerná produkcia spermií
veľké vajíčko,
prítomnosť gamónov, ktoré prispievajú ku konvergencii a interakcii gamét,
prítomnosť kopulačných orgánov, ktoré zabezpečujú vnútornú insemináciu.
U cicavcov veľký význam má pobyt spermií v pohlavnom trakte samice, v dôsledku čoho získavajú samčie zárodočné bunky schopnosť oplodnenia, t.j. schopnosť akrozomálnej reakcie.
2. fáza - aktivácia gamét nastáva po ich kontakte. Aktivácia spermií sa nazýva akrozomálna reakcia. Aktivácia vajíčka je kortikálna reakcia.
Podstata akrozomálnej reakcie : Priepustnosť spermií v akrozóme sa mení mostík plazmatickej membrány a z akrozómu sa uvoľňujú enzýmy, spermatolyzíny. Tieto enzýmy uvoľňujú väzby medzi folikulárnymi bunkami, ktoré obklopujú vajíčko. Spermie prechádzajú cez vrstvu folikulárnych buniek, potom je zóna priezračnej bunky zničená a spermie prechádzajú cez túto zónu.
Podstata kortikálnej reakcie: Spočíva v zložitých štruktúrnych a fyzikálno-chemických zmenách. Vzhľadom na skutočnosť, že membrána spermií je priepustná pre ióny sodíka, tieto začnú vstupovať do vajíčka a menia sa membránový potenciál bunky. Potom vo forme vlny šíriacej sa od miesta kontaktu gamét dochádza k zvýšeniu obsahu iónov Ca 2+ (v hyaloplazme opúšťajú depot - EPS, retikulum) a vo vajíčku sa spúšťajú biochemické procesy, potom sa kortikálne granule tiež rozpustia vo vlne. Špecifické enzýmy uvoľnené súčasne prispievajú k oddeleniu žĺtkovej membrány; ona to stvrdne oplodňovacie puzdro.
Jedným z významov kortikálnej reakcie je prevencia polyspermie, t.j. prienik viac ako jednej spermie do vajíčka. U cicavcov kortikálna reakcia nespôsobuje tvorbu oplodňovacej membrány, ale jej podstata je rovnaká.
Aktivácia vajíčka sa končí začiatkom syntézy proteínov na translačnej úrovni, keďže mRNA, tRNA, ribozómy a energia boli uložené aj v oogenéze.
3 etapa- splynutie gamét, čiže syngamia. V tomto prípade sa v spermiách a vajíčku vytvorí spoločná plazmatická membrána. Ženské a mužské pronucleus sa zbiehajú a spájajú (synkaryón), čím vytvárajú spoločnú metafázovú platňu. Toto je moment konečného splynutia gamét – syngamie.
Vlastnosti hnojenia v rôznych druhoch organizmov.
1 príklad. U cicavcov a ľudí sa spermie viaže na vajíčko v oblasti, kde je receptor na zona pellucida. Po tejto interakcii sú zvyšné receptory zablokované.
2 príklad. o morský ježko po oplodnení sa vo vajíčku dramaticky zmení elektrický potenciál plazmatická membrána a potom sa vytvorí oplodňovacia membrána, ktorá zabraňuje polyspermii.
štádium zygoty. Po penetrácii sa mužské jadro nazýva mužské pronukleus. Chromatín sa v ňom uvoľňuje, dochádza k replikácii DNA. Ženské jadro sa nazýva ženské pronukleus. Odohrávajú sa v ňom rovnaké udalosti. U cicavcov a ľudí nedochádza k jadrovej fúzii, ale okamžite vzniká metafázová platňa.
5. Umelé oplodnenie vajíčka zvierat a ľudí, biologické a medicínske aspekty.
Umelé oplodnenie vajíčok zvierat má pre medicínu veľký vedecký význam, pretože v procese jeho štúdia sa vyvíjajú spôsoby a mechanizmy liečby neplodnosti u ľudí.
Umelé oplodnenie sa používa na rôzne formy neplodnosť, mužská aj ženská, ktorá sa ťažko lieči. Napríklad, keď má muž príliš málo spermií alebo sú prakticky nepohyblivé, keď má žena zhoršenú priechodnosť vajíčkovodov alebo došlo k inému poškodeniu vnútorných pohlavných orgánov s imunologickou inkompatibilitou partnerov.
6. všeobecné charakteristiky drvenie. Typy drvenia charakteristické pre rôzne druhy zvierat. Štiepenie a tvorba blastuly u placentárnych cicavcov.
štádium drvenia. Toto je štádium tvorby jednovrstvového embrya - blastuly. Vo vnútri blastuly je dutina nazývaná blastocoel.
Vlastnosti drvenia:
Bunky sa delia mitózou.
V predvečer každého delenia dochádza k replikácii DNA.
Deliace sa bunky nerastú.
Typ drvenia závisí od druhu vajíčka.
Kompletné rovnomerné drvenie v lancelete:
Prvá štiepna brázda prebieha vertikálne, tvoria sa dve blastoméry. Druhá brázda prebieha tiež vertikálne a tvoria sa štyri blastoméry. Tretia brázda prebieha horizontálne, vytvára sa osem blastomér a potom sa striedajú vertikálne a horizontálne brázdy. Po 12 cykloch sa drvenie stane asynchrónnym. V určitom štádiu vývoja je embryo zhlukom buniek alebo morulou. Potom sa medzi bunkami objavia medzery a vytvorí sa dutina - blastocoel. V lancelete pri drvení vzniká blastula, ktorá sa nazýva coeloblastula, teda jednovrstvová gulička.
Úplná nerovnomerná fragmentácia u obojživelníkov:
Bunky obojživelníkov sú stredne telolecitálne. Na živočíšnom póle bunky prebieha fragmentácia rýchlejšie ako na vegetatívnom póle. V dôsledku toho sú bunky na zvieracom póle menšie – mikroméry. Na vegetatívnom póle sú bunky väčšie – makroméry. Blastula obojživelníka sa nazýva amfiblastula. Blastocoel sa nachádza na zvieracom póle.
Vlastnosti drvenia u cicavcov a ľudí:
Štiepenie je od prvých štádií úplne nerovnomerné, asynchrónne, v určitom štádiu vývoja je embryo morula (hrudka buniek). Potom sa väčšie bunky oddelia na periférii, čím sa vytvorí trofoblast, a menšie bunky do stredu, čím sa vytvorí embryoblast. Blastula sa nazýva blastocysta. Blastocoel je veľmi malý. Trofoblast podporuje zavedenie embrya do sliznice maternice. Tento proces sa nazýva implantácia. Z embryoblastu vzniká samotné embryo a niektoré dočasné orgány.
7. Všeobecná charakteristika gastrulácie. Vlastnosti gastrulácie u obojživelníkov a vtákov. Gastrulácia u vyšších (placentárnych) cicavcov.
Štádium gastrulácie, alebo štádium tvorby dvojvrstvového embrya a následne trojvrstvového. Embryo v tomto štádiu sa nazýva gastrula.
Metódy tvorby dvojvrstvového embrya:
- Intususcepcia (invaginácia).
– Delaminácia (stratifikácia).
– Imigrácia (osídľovanie).
- Epiboly (znečistenie).
Intususcepcia alebo invaginácia. Táto metóda je typická pre lancelet. V určitej oblasti sa bunky blastuly invaginujú do blastocoelu, čo vedie k vytvoreniu dvojvrstvového embrya. Vonkajšia vrstva buniek sa nazýva ektoderm, vnútorná vrstva sa nazýva endoderm. Endoderm obmedzuje dutinu primárneho čreva alebo gastrocoel. Vstup do tejto dutiny sa nazýva primárne ústa alebo blastopór. Blatopór je obklopený perami.
Delaminácia alebo stratifikácia. Táto metóda je typická pre črevné zvieratá, u ktorých blastula vyzerá ako morula a blastocoel sa prakticky neexprimuje.
prisťahovalectvo alebo osídlenie. Niektoré bunky blastuly sú zavedené do blastocoelu, potom sa tieto bunky intenzívne delia. V dôsledku týchto buniek sa vytvára endoderm.
Epiboly (znečistenie). Mikroméry sú rozdelené a akoby navrstvené na makroméry. Mikroméry tvoria ektodermu, zatiaľ čo makroméry tvoria endodermu. Vo svojej čistej forme sa tieto metódy prakticky nenachádzajú, spravidla sa kombinujú. Obojživelníky kombinujú invagináciu a epibóliu. Vtáky a cicavce spájajú delamináciu a imigráciu.
Počnúc plochými červami sa v evolúcii objavuje tretia zárodočná vrstva, mezoderm.
Metódy tvorby mezodermu:
Teloblastická metóda je charakteristická pre protostomy. V oblasti pyskov blastopóru sa vylučujú 2 bunky, ktoré sa delia a vytvárajú mezoderm.
Metóda enterocoel je charakteristická pre deuterostómy (strunatcov). Od endodermu sú symetricky oddelené 2 sekcie buniek vo forme vreciek. Toto sú mezodermálne vrecká. Bunky mezodermálnych vreciek sa delia a dávajú vznik mezodermu. Mezoderm je zárodočná vrstva.
8. Všeobecná charakteristika histo- a organogenézy (tvorba tkanív a orgánov).
Štádium histo a organogenézy(štádium tvorby tkanív a orgánov). Tradične sa delí na dve obdobia.
1 obdobie. Obdobie vzdelávania axiálne orgány v embryu je to tvorba neurálnej trubice a chordy. Preto sa toto obdobie nazýva obdobie neurulácie a embryo v tomto štádiu sa nazýva neurula.
2 obdobie. Je charakterizovaná tvorbou iných tkanív a orgánov. Na chrbtovej strane embrya (dorzálnej) sa po celej dĺžke oddeľuje od ektodermy úsek buniek, z ktorých vzniká nervová platnička. Potom sa okraje neurálnej platničky dvíhajú, zhrubnú a vytvorí sa neurálna ryha, ktorá postupne klesá pod ektodermu. Potom sa okraje nervovej drážky uzavrú a vytvoria nervovú trubicu s dutinou vo vnútri, dutina sa nazýva neurocoel. U stavovcov sa predná časť nervovej trubice rozširuje a dáva vznik mozgu, zvyšok mieche. Súčasne je pod nervovou trubicou položený notochord, ktorý sa tvorí z endodermu a priľahlého mezodermu. Spočiatku je mezoderm homogénna bunková hmota, ale ako sa vyvíja, dochádza k jej segmentácii. Vznikajú štruktúry, ktoré sa nazývajú somity. Následne z nich vzniká pohybový aparát.
Deriváty zárodočných vrstiev:
Ektoderm - zubná sklovina, nervový systém a zmyslové orgány, epidermis kože a jej prídavky, epitel predného a zadného čreva.
Endoderm – epitel stredného čreva, tráviace žľazy a dýchací systém.
Mesoderm - pohybový aparát, urogenitálny systém, obehový a lymfatický systém, všetko spojivové tkanivo.
9. Charakteristika provizórnych orgánov embryí stavovcov. Dočasné orgány vyšších cicavcov.
Dočasné orgány fungujú v embryu a chýbajú v dospelom stave. Patria sem žĺtkový vak a takzvané embryonálne membrány - amnion, chorion a alantois.
Žĺtkový vak.Žĺtkový vak plní množstvo dôležitých funkcií: výživu, dýchanie, vylučovanie, krvotvorbu. Ale vzhľadom na nízky obsah žĺtka vo vajci nezohráva významnú úlohu vo výžive embrya.
Amnion. Na tvorbe amniónu sa podieľa amniová membrána, ktorá obmedzuje amniovú dutinu vyplnenú plodovou vodou, ktorá teraz obklopuje embryo zo všetkých strán. Vďaka tomu sa embryo vyvíja vo vodnom prostredí, ktoré ho chráni pred mechanickými traumatickými účinkami a priľnavosťou k škrupinám.
Allantois vytvorený ako výrastok zadného čreva. Hlavnou funkciou alantoisu je, že je zárodočným orgánom vylučovania. Zhromažďuje produkty rozpadu, ktoré vznikajú pri metabolizme v tele embrya.
Embryogenéza je komplexný holistický proces, ktorý je spojený s určitými javmi a mechanizmami. Mnohé z týchto javov nie sú úplne pochopené, hoci pre niektoré z nich boli získané určité údaje.
1. Molekulárne genetické zmeny v ranom vývoji.
2. Bunková proliferácia (bunkové delenie).
3. Diferenciácia buniek.
4. Tvorba alebo morfogenéza.
1. Molekulárne genetické zmeny v ranom vývoji (obdobie zygoty a štiepenia), úloha cytoplazmatických faktorov vo vajíčku.
Skorý vývoj zahŕňa štádiá zygoty a štiepenia.
Štúdiom týchto fáz sa vedci pokúsili odpovedať na otázky:
Po prvé, keď začnú fungovať vlastné gény embrya.
Po druhé, existujú kvalitatívne a kvantitatívne rozdiely v molekulách mRNA a proteínov rôzne časti embryo v počiatočných štádiách vývoja.
V zygote je génová aktivita nízka, pretože DNA je silne spojená s histónovými proteínmi. Prvé proteíny, ktoré sa syntetizujú v zygote, sú materského pôvodu, pretože ribozómy a molekuly mRNA sa vopred nahromadili vo vajíčku. Zistilo sa, že vlastné gény embrya u cicavcov začínajú pracovať v štádiu 2-4 blastomérov. U obojživelníkov je v štádiu blastuly. Ako prvé začnú fungovať gény zodpovedné za proliferáciu a celkový metabolizmus, neskôr začnú pôsobiť gény zodpovedné za diferenciáciu buniek a tkanív. Napríklad, keď sa jadro odstráni zo zygoty, dôjde k štiepeniu a embryo vo svojom vývoji dosiahne takmer štádium blastuly, po ktorom sa ďalší vývoj zastaví.
Zistilo sa, že neexistujú žiadne kvalitatívne rozdiely v molekulách mRNA a proteínov v rôznych častiach embrya v skorých štádiách vývoja. Existujú iba kvantitatívne rozdiely.
Dôležitú úlohu pri drvení hrá delenie cytoplazmy - cytotómiu. Má osobitný morfogenetický význam, pretože určuje typ drvenia. Po hraniciach medzi jednotlivými úsekmi ooplazmy prebiehajú štiepne ryhy, ktoré odrážajú fenomén ooplazmatickej segregácie. Preto sa cytoplazma rôznych blastomérov líši v chemickom zložení.
2. Bunková proliferácia, rast.
Bunková proliferácia alebo bunkové delenie prebieha počas embryogenézy. S tým je spojený rast tkanív a orgánov. Rast embrya ako celku.
3. Diferenciácia, molekulárne genetické mechanizmy diferenciácie.
Bunková diferenciácia je súbor procesov, v dôsledku ktorých bunky spoločného pôvodu nadobúdajú pretrvávajúce morfologické, fyziologické, biochemické rozdiely, čo vedie k špecializácii buniek. Špecifickosť buniek je určená proteínmi, ktoré sú v nich syntetizované a za proteíny sú zodpovedné zodpovedajúce gény. Preto môžeme konštatovať, že niektoré gény fungujú v niektorých bunkách a iné v iných. To je podstata hypotézy rozdielnej aktivity génov.
V skorých štádiách je diferenciácia buniek spojená s vplyvom cytoplazmatických látok na prácu zodpovedajúcich génov - to je epigenetická úroveň regulácie práce génov. Vo vajíčku prebieha fenomén ooplazmatickej segregácie, v dôsledku čoho rôzne časti cytoplazmy vajíčka obsahujú rôzne látky. Počas drvenia sa objavujú blastoméry, súbor génov v nich je rovnaký a zloženie cytoplazmy je odlišné. Následne tieto látky cytoplazmy zjavne vedú k rozdielnej aktivite génov.
Pri charakterizácii bunkovej diferenciácie sa používajú dva pojmy – determinácia a kompetencia.
rozhodnosť znamená, že diferenciácia buniek je geneticky predurčená a ireverzibilná.
V procese diferenciácie sa bunkový materiál embryonálnych anlage transformuje na určitý prvok dospelého organizmu. Uvažujme o diferenciácii na príklade mezodermálneho somitu, ktorý sa ďalej delí na dermatóm, sklerotóm a myotóm. Dermatómové bunky dermis, druhé - bunky chrupavky, tretie - priečne pruhované svalové vlákna. V dôsledku toho je konečný výsledok vývoja jednotlivých embryonálnych zmien vopred určený alebo určený.
kompetencie Ide o schopnosť buniek diferencovať sa v rôznych smeroch pod vplyvom faktorov prostredia. Napríklad notochord a priľahlý mezoderm pôsobia na ektodermu, čo vedie k vytvoreniu neurálnej trubice z ektodermy. Ak takýto účinok neexistuje, potom ektoderm vedie k vzniku epidermis kože.
4. Morfogenéza (tvarovanie), jej hlavné procesy:
Tvorba alebo morfogenéza. Morfogenéza je súbor procesov, v dôsledku ktorých embryo získava charakteristický vonkajší a vnútorná štruktúra. Morfogenéza je zase spojená s:
a) morfogenetický pohyb buniek
Počas embryogenézy sa jednotlivé bunky alebo skupiny buniek pohybujú. Bunky sa pohybujú na povrchu iných buniek (vďaka mechanizmu améboidného pohybu), kde sa nachádzajú špeciálne molekuly, ktoré udávajú smer pohybu. Niektoré typy buniek sa pohybujú pozdĺž chemického koncentračného gradientu (chemotaxia), ale tento mechanizmus je oveľa menej bežný.
Porušenie migrácie buniek počas embryogenézy vedie k nedostatočnému rozvoju orgánov alebo k zmene ich normálnej lokalizácie. Obidve sú vrodené chyby. Napríklad pri narušení migrácie buniek – neuroblastov sa v bielej hmote objavujú ostrovčeky šedej hmoty a zároveň bunky strácajú schopnosť diferenciácie.
Migrácia buniek je teda na jednej strane pod genetickou kontrolou a na druhej strane vplyvom okolitých buniek a tkanív.
b) embryonálna indukcia
Ide o vplyv jedného tkaniva (induktora) na iné tkanivo, v dôsledku čoho sa vývoj indukovaného tkaniva stáva kvalitatívne novým. Prvou a najvýznamnejšou indukciou je pôsobenie notochordu a mezodermu na ektodermu, výsledkom čoho je vytvorenie nervovej trubice. Bez neurálnej trubice sa celý ektoderm premení na epidermis. Ide o primárnu embryonálnu indukciu, prvý krok v reťazci po sebe nasledujúcich (sekundárnych, terciárnych) indukčných procesov v ďalšom vývoji.
Zistilo sa, že existujú "špecifické induktory", t.j. látky, ktoré majú indukčný účinok v zanedbateľných koncentráciách a líšia sa konečným výsledkom svojho pôsobenia. Výťažok z pečene cicavcov teda vyvoláva najmä vývoj mozgových štruktúr a výťažok z kostnej drene - mezodermálnych.
Schopnosť embryonálneho zárodku vnímať indukčný stimul sa nazýva kompetencia.
c) medzibunkové interakcie
Ide o interakciu buniek alebo vrstiev v kontakte alebo na diaľku. Interakcia na diaľku prebieha za účasti biologicky aktívnych látok (BAS).
Môžu to byť bielkoviny, hormóny atď. V počiatočných štádiách embryonálneho vývoja sú to hormóny matky, keďže embryo nemá vlastné endokrinné žľazy. Hormóny nespôsobujú novú diferenciáciu, ale posilňujú ju.
V dôsledku medzibunkových interakcií sa uskutočňujú také javy, ako je morfogenetický pohyb buniek, embryonálna indukcia a bunková adhézia.
d) priľnavosť- schopnosť buniek spájať sa. V experimente boli bunky ektodermu, mezodermu a endodermu oddelené a navzájom zmiešané. Potom sa opäť zhromažďujú do samostatných skupín, z ktorých každá je bunkovým agregátom homogénnych buniek. Opätovne sa vytvoria tri zárodočné vrstvy, ktoré sú voči sebe normálne umiestnené.
Na procese adhézie sa zúčastňujú špeciálne molekuly proteínov. Nazývajú sa bunkové adhézne molekuly (MAC) a existuje asi 100 typov.
Ďalšia hypotéza tvrdí, že kontakty medzi podobnými bunkami sú silnejšie ako medzi cudzími bunkami.
Selektívna adhézia buniek určitej zárodočnej vrstvy medzi sebou je nevyhnutnou podmienkou normálneho vývoja.
e) bunková smrť je nevyhnutný proces, pretože tvorba jednotlivých štruktúr (kanálikov, kanálikov, otvorov atď.) vyžaduje deštrukciu časti buniek.
Existujú dva zásadne odlišné typy bunkovej smrti: apoptóza(v preklade z gréčtiny „odpadnúť“) a nekróza.
Apoptóza je fyziologická, geneticky predurčená bunková smrť. Spolu s ďalšími mechanizmami morfogenézy prispieva k dosiahnutiu znakov jeho morfofunkčnej organizácie charakteristickej pre určitý biologický druh. Preto je apoptóza prirodzený, evolučne určený a geneticky riadený mechanizmus morfogenézy.
Nekróza - nefyziologická bunková smrť, v dôsledku vplyvu nepriaznivých faktorov (mechanických, chemických, fyzikálnych atď.). Nekróza je zvyčajne sprevádzaná zápalom a je patologickým procesom.
5. Integrácia vo vývoji, integrita ontogenézy. Úloha hormónov v koordinácii vývojových procesov.
V súčasnosti je známe množstvo látok, ktoré vyvolávajú delenie buniek, napríklad fytohemaglutinín, niektoré hormóny, ale aj komplex látok uvoľňovaných pri poškodení tkaniva. Boli objavené aj tkanivovo špecifické inhibítory bunkového delenia - keylony. Ich pôsobením je potlačiť alebo spomaliť rýchlosť delenia buniek v tkanivách, ktoré ich produkujú. Napríklad epidermálne kalóny pôsobia iba na epidermis. Keďže sú kalóny tkanivovo špecifické, nemajú druhovú špecifickosť. Epidermálny kalon tresky teda pôsobí aj na epidermis cicavca.
Hormóny sú organické zlúčeniny produkované určitými bunkami a určené na kontrolu telesných funkcií, ich reguláciu a koordináciu.
Fyziologické pôsobenie hormónov je zamerané na:
1) poskytovanie humorných, t.j. vykonávané krvou, regulácia biologických procesov;
2) udržiavanie integrity a stálosti vnútorného prostredia, harmonická interakcia medzi bunkovými zložkami tela;
3) regulácia procesov rastu, dozrievania a rozmnožovania. Hormóny regulujú činnosť všetkých telesných buniek. Ovplyvňujú bystrosť myslenia a fyzickú pohyblivosť, postavu a rast, podmieňujú vývoj znakov sexuálneho dimorfizmu a správania.
6. Úloha dedičnosti a prostredia v embryonálnom vývoji. Kritické obdobia vývoja. teratogénne faktory. Anomálie a malformácie.
V ktoromkoľvek štádiu ontogenézy organizmus existuje v jednote s životné prostredie. Embryogenéza nie je v tomto smere výnimkou. Rozsah podmienok potrebných pre život druhu môže byť široký. Pre organizmy akéhokoľvek druhu však existuje minimum, optimum a maximum nevyhnutné podmienky rozvoj. Vývoj embrya je ovplyvnený kolísaním prirodzene sa vyskytujúcich faktorov (teplota, vlhkosť, atmosférický tlak, žiarenie, plynné zloženie prostredia).
Takže v závislosti od teploty sa procesy vývoja spomaľujú alebo zintenzívňujú. Napríklad žabie vajcia z rovnakej znášky sa pri vyšších teplotách vyvíjajú rýchlejšie.
U škrkavky, keď je ukončený prístup kyslíka k embryu, sa vývoj zastaví.
Všeobecné pravidlo je, že pôsobením svetla z modrofialovej časti spektra sa embryonálny vývoj mnohých živočíšnych druhov zrýchli a z červenej sa spomalí.
Počas vnútromaternicového vývoja majú veľký význam faktory prostredia. Ak tieto faktory vedú k vzniku anomálií alebo vývojových chýb, potom sa nazývajú teratogénne. Teratogénne faktory môžu byť fyzikálne (vysoká teplota, ionizujúce žiarenie, RTG a pod.), chemické (lieky, soli ťažkých kovov a pod.) a biologické (vírusy, baktérie). Teratogénne faktory vedú k rozvoju anomálií v určitých obdobiach embryonálneho vývoja, ktoré sa nazývajú kritické. Tie obsahujú:
Obdobie tvorby zárodočných buniek (gametogenéza),
štádium oplodnenia,
štádium zygoty,
Implantácia embrya do steny maternice
Placentácia,
Obdobie histogenézy a organogenézy,
Malformácie.
Aplázia - absencia orgánu alebo jeho časti
Hypoplázia - nedostatočné rozvinutie orgánu
Hypotrofia - zníženie telesnej hmotnosti alebo hmotnosti orgánu
Hypertrofia - neúmerné zvýšenie hmoty orgánu
Gigantizmus - zvýšenie dĺžky tela
Heterotopia je atypická lokalizácia skupiny buniek alebo orgánu v tele.
Heteroplázia - porušenie diferenciácie tkanív
Stenóza - zúženie kanála alebo otvoru
Atrézia - absencia kanála alebo otvoru
Perzistencia – zachovanie embryonálnych štruktúr
V závislosti od príčiny sa vrodené chyby delia na:
dedičné spôsobené zmenou génov alebo chromozómov v gamétach rodičov, v dôsledku čoho zygota od samého začiatku nesie gén, chromozóm alebo genómovú mutáciu.
exogénne vznikajúce pod vplyvom teratogénnych faktorov: liečivá (talidomid), prídavné látky v potravinách, vírusy, priemyselné jedy atď. To všetko sú faktory prostredia, ktoré počas embryogenézy narúšajú vývoj tkanív a orgánov.
Multifaktoriálny defekty, ktoré sa vyvíjajú pod vplyvom exogénnych aj genetických faktorov.
1. Postnatálna ontogenéza, jej periodizácia.
Postembryonálny vývoj (pre človeka postnatálny) začína od okamihu narodenia a končí prirodzenou smrťou alebo smrťou.
Postembryonálny vývoj zahŕňa niekoľko období:
1. Predreprodukčné (juvenilné).
2. Reprodukčné (doba zrelosti).
3. Postreprodukčná (staroba).
2. Predreprodukčné obdobie, jeho charakteristika. Rast tela ako dôležitá charakteristika predreprodukčného obdobia.
Predreprodukčné obdobie začína bezprostredne po narodení. V tomto čase sa končia procesy morfogenézy, začínajú fungovať tie systémy, ktoré v embryogenéze nefungovali (respiračné, vylučovacie a mnohé iné).
Dôležitou charakteristikou predreprodukčného obdobia je rast organizmu. V tomto prípade dochádza k zvýšeniu veľkosti tela ako celku, jeho pozdĺžne rozmery sa zvyšujú; zvýšenie veľkosti tkanív a orgánov.
3. Charakter rastu organizmu a jeho jednotlivých častí.
Základom rastu organizmu sú tri hlavné procesy:
1. zvýšenie počtu buniek.
2. zväčšenie veľkosti buniek (hypertrofia).
3. hromadenie medzibunkovej látky.
Existujú dve možnosti rastu: obmedzené A neobmedzené. Neobmedzený rast pokračuje počas ontogenézy až do smrti.
Existuje niekoľko typov rastu:
Pomocný- rast, ktorý prebieha zväčšením veľkosti buniek.
proliferatívny- rast prebiehajúci množením buniek: multiplikatívne A akrečný.
Multiplikatívne rast sa vyznačuje tým, že obe bunky, ktoré vznikli delením rodičovskej bunky, opäť vstupujú do delenia. Multiplikatívny rast je veľmi efektívny, a preto sa takmer nikdy nevyskytuje vo svojej čistej forme alebo končí veľmi rýchlo (napríklad v embryonálnom období).
akrečný rast spočíva v tom, že po každom ďalšom delení sa len jedna z dcérskych buniek opäť rozdelí, zatiaľ čo druhá sa prestane deliť. V tomto prípade počet buniek rastie lineárne. Takýto rast je typický pre orgány, kde dochádza k obnove bunkového zloženia.
Je potrebné zdôrazniť, že zväčšenie pozdĺžnych rozmerov tela, ku ktorému dochádza najmä v dôsledku rastu dlhých tubulárnych kostí, má osobitný význam pri charakterizácii rastu. IN tubulárne kosti na hranici diafýzy a epifýzy je izolovaná rastová zóna. Tu sú bunky chrupavky, pri delení ktorých kosť rastie do dĺžky.
Konečná osifikácia každej kosti nastáva v určitom čase. U mužov rast zvyčajne končí vo veku 18-20 rokov, u žien - o 16-18 rokov. V tomto čase zmiznú posledné rastové zóny. Vtedy sa zastaví rast kostí do dĺžky.
Je potrebné zdôrazniť, že až do 30 rokov môže človek rásť o 3 cm v dôsledku zväčšenia stavcov.
Nárast lineárnych rozmerov osoby je opísaný krivkou v tvare S. Bezprostredne po narodení dochádza k zvýšenému rastu tela, potom sa znižuje a prudko zrýchľuje do 13-14-15 rokov. Ide o takzvaný pubertálny rastový spurt (počas puberty). Ďalej sa tempo rastu trochu spomaľuje a vo veku 30-40-45 rokov zostáva rast človeka konštantný. Podľa tejto schémy kosti, svaly a mnohé vnútorné orgány(pečeň, obličky, slezina).
So starnutím dochádza k miernemu poklesu rastu.
Niektoré orgány majú úplne odlišný charakter:
Tieto orgány zahŕňajú mozog a miechu, lymfoidné orgány, reprodukčné orgány.
Hmotnosť mozgu novorodenca je 25% konečnej hmotnosti mozgu (v dospelosti), o 5 rokov - 90%, o 10 rokov - 95%.
Rast týmusu (brzlík) je centrálnym orgánom imunitného systému. Relatívna hmotnosť týmusu (k telesnej hmotnosti) dosahuje maximum do 12. roku života. Absolútna hmotnosť dosahuje maximum do 30. roku života a potom dochádza k prudkému poklesu hmotnosti týmusu.
4. Genetická kontrola rastu. Úloha nervového a endokrinného systému v regulácii rastových procesov.
Výška sa vzťahuje na genetické vlastnosti, ktoré sú zdedené, ako je farba vlasov a pokožky, tvar očí atď. Preto majú vysokí rodičia väčšinou vysoké deti a naopak. Rast je polygénna vlastnosť, za jej prejav vo fenotype je zodpovedných niekoľko génov. Gény vykonávajú kontrolu nad rastom prostredníctvom vhodných hormónov. Najdôležitejším hormónom je rastový hormón alebo somatotropín produkovaný hypofýzou.
Somatotropín stimuluje tvorbu nových buniek chrupavky a čiastočne aj ich osifikáciu, podporuje syntézu bielkovín v bunkových štruktúr ah a tvorbu nových kapilár. Veľké množstvo tohto hormónu sa produkuje v noci. Vlastný somatotropín dieťaťa sa vyrába od 3 do 4 rokov.
Rast ovplyvňujú aj hormóny štítnej žľazy a pohlavné hormóny.
5. Interakcia biologického a sociálneho v detstve a mladosti.
Úloha dedičnosti pre rast je skvelá, ale nie je to jediný faktor. Dedičnosť treba považovať za približný program, podľa ktorého môže byť výška človeka napríklad v rozmedzí od 160 do 180 cm. Čo to v skutočnosti bude, závisí vo veľkej miere od vonkajších podmienok, ktoré môžu spomaliť dedičný program alebo prispieť k jeho realizácii. Podmienky prostredia, ktoré ovplyvňujú ľudský rast: výživa, fyzická aktivita, psychologický dopad fajčenie, alkohol.
To znamená, že populácia rastie (toto je zrýchlenie).
Jednou z jeho pravdepodobných príčin je zlepšenie životných podmienok (výživa). Všimli sme si, že počas rokov vojen a prírodných katastrof sa rast detí znižuje. Rast mierne ovplyvňuje klíma a geografické prostredie.
6. Formovanie konštitučných typov, telesné typy.
Formovanie konštitučných typov ľudí je spojené s rastom človeka. Malo by sa to chápať ako znaky vonkajších foriem tela, znaky funkcií tela, znaky správania táto osoba. V závislosti od štruktúry tela, v závislosti od vonkajších foriem tela, sa rozlišujú určité typy postavy. V súčasnosti existuje pomerne veľa klasifikácií. Jednou z nich je klasifikácia M.V. Chernorutsky. Podľa tohto
Každý organizmus, bez ohľadu na to, či je jednobunkový alebo mnohobunkový a do ktorej ríše života patrí, prechádza individuálny rozvoj, alebo ontogenézy(z gréčtiny. tov- tvor a genéza- narodenie). Študuje odbor biológia ontogenézy, sa volá vývinová biológia.
Ontogenéza - ide o obdobie života organizmu od zygoty (primárnej bunky) po smrť.
U mnohobunkových organizmov sa ontogenéza zvyčajne začína vytvorením zygoty a končí smrťou.
Telo zároveň nielen rastie, zväčšuje sa, ale prechádza aj množstvom rôznych životných fáz, z ktorých každá má špeciálnu štruktúru, inak funguje a v niektorých prípadoch aj radikálne odlišný spôsob života. .
U jednobunkových organizmov sa za začiatok ontogenézy považuje moment oddelenia od materskej alebo sesterskej bunky. Pokračuje až do ďalšieho rozdelenia alebo smrti. Vonkajšia ontogenéza jednobunkových organizmov sa zároveň zvyčajne prejavuje len miernym zväčšením veľkosti bunky, hoci v skutočnosti sa za tým ukrývajú úplne iné obdobia jej života. .
Každý druh má svoj vlastný program ontogenézy. A to nie je len súbor a postupnosť etáp vývoja, ktorými prechádza, ale aj trvanie každého z nich. Každý jedinec má zároveň individuálne znaky ontogenézy, ktoré však nepresahujú druh. , a tí sa zasa riadia zákonmi ontogenézy na kmeňovej, rodinnej, oddelenej a dokonca triednej úrovni .
program ontogenézy nie je nič iné ako realizácia dedičnej informácie zaznamenanej v génoch. Preto je špecifickosť ontogenézy na úrovni jednotlivých jedincov určená individuálnymi kombináciami génov a na úrovni druhov, rodov, rodín - špeciálnymi génmi, ktoré sú charakteristické iba pre každú systematickú skupinu organizmov.
Mechanizmus implementácie dedičnej informácie je v prvom rade v diferenciál(z angličtiny. líšiť- meniť sa) génová aktivita. To znamená, že v rôznych obdobiach vývoja a v rôznych tkanivách mnohobunkový organizmus aktívne sú gény, na ktorých nukleotidových sekvenciách sa syntetizujú určité mRNA. V dôsledku toho sa syntetizujú špecifické štrukturálne proteíny a enzýmy, ktoré v konečnom dôsledku určujú vlastnosti fungovania a správania buniek v určitých štádiách vývoja tela, vlastnosti ich metabolizmu.
U zvierat a kvitnúcich rastlín sa ontogenéza delí na dve obdobia. Embryonálny vývoj, alebo embryogenéza(z gréčtiny. embryo- plod a genéza) trvá od vytvorenia zygoty po pôrod alebo výstup z vajíčka a postembryonálne(z gréčtiny. rýchlo- po a embryo) vývoj pokračuje od narodenia alebo výstupu z vajíčka a končí smrťou organizmu.
Priebeh embryogenézy. Všeobecná schéma embryogenézy zahŕňa nasledujúce fázy: materiál zo stránky
- postupné delenia zygoty, ktoré vyvrcholia vytvorením mnohobunkového embrya, pozostávajúceho zo stoviek a dokonca tisícov identických buniek;
- diferenciácia (z angl. líšiť- rozlišovať) bunky, čo vedie k tvorbe tkanív;
- kladenie orgánov a rast embrya.
Počas ontogenézy organizmus prechádza množstvom fáz – stavov, v ktorých sa odlišuje štruktúrou, fungovaním a spôsobom života. Existujú dve etapy ontogenézy: embryogenéza - embryonálny vývoj a postembryonálny vývoj- obdobie života organizmu od narodenia (výstupu z vajíčka) po smrť.
Na tejto stránke sú materiály k témam:
Zhrnutie ontogenézy
Ontogenéza individuálneho vývinu abstrakt
Stručne pojem ontogenéza
-
Kapitola 7. PERIODIZÁCIA ONTOGENÉZY
Kapitola 7. PERIODIZÁCIA ONTOGENÉZY
7.1. etapy, obdobia a etapy ontogenézy
Individuálny vývoj je holistický nepretržitý proces, v ktorom sú jednotlivé udalosti prepojené v priestore a čase. Existuje niekoľko variantov periodizácie ontogenézy, z ktorých každá je najvhodnejšia na riešenie konkrétnych vedeckých alebo praktických problémov.
S všeobecné biologické uhly pohľadu významnú udalosť ontogenéza - sexuálnej reprodukcie. Ak korelujeme rôzne časové obdobia ontogenézy so schopnosťou jedinca vykonávať reprodukčnú funkciu, potom ju možno rozdeliť na tri obdobia: predreprodukčné, aktívne reprodukčné a postreprodukčné.
IN predreprodukčné obdobie jedinec nie je schopný reprodukcie. Jeho hlavný obsah spočíva vo vývoji sexuálne zrelého fenotypu. V tomto období dochádza k najvýraznejším štrukturálnym a funkčným premenám, realizuje sa hlavná časť dedičných informácií, telo je vysoko citlivé na všetky druhy vplyvov.
V aktívnom reprodukčné obdobie jedinec vykonáva funkciu sexuálnej reprodukcie, vyznačuje sa najstabilnejším fungovaním orgánov a systémov, ako aj relatívnou odolnosťou voči vplyvom.
Postreprodukčné obdobie spojené so starnutím tela a je charakterizované oslabením alebo úplným zastavením účasti na reprodukcii. Znížená adaptačná kapacita a odolnosť voči rôznym vplyvom. Vo vzťahu k ontogenéze človeka sú tieto obdobia navyše charakterizované špecifickými sociálnymi faktormi (vzdelanie, schopnosť pracovať, tvorivosť). Každé obdobie má svoje vlastné charakteristiky chorobnosti.
Predreprodukčné obdobie je rozdelené na ďalšie štyri: embryonálne, larválne, metamorfózne a juvenilné.
embryonálne, alebo zárodočný, bodka Ontogénia začína od okamihu oplodnenia a pokračuje až do uvoľnenia embrya z vaječných membrán. Toto obdobie sa vyznačuje závažnosťou procesov transformácie zygoty na organizmus schopný viac-menej samostatnej existencie. U väčšiny stavovcov zahŕňa štádiá (fázy) zygoty, štiepenia, gastrulácie, ako aj histo- a organogenézu. Jeho trvanie je rôzne. U placentárnych cicavcov je obzvlášť skrátená. Jedna škrupina vajíčka sa rozpustí pred implantáciou blastocysty do výstelky maternice. Do tejto doby má embryo len čas prejsť fázami zygoty a drvenia. Všetky ďalšie procesy prebiehajú pod ochranou a za účasti tela matky. Evolučný význam týchto vlastností je zvažovaný v časti 13.2.
Štiepeniu predchádzajú procesy gametogenézy a oplodnenia, ktoré priamo súvisia s individuálnym vývojom a nemusia k nemu ani viesť, ale do značnej miery určujú ďalší vývoj embrya, ak dôjde k počatiu. Tieto procesy sa nazývajú progenéza, ktorá predchádza vlastnej ontogenéze. Cytologicky procesy gametogenézy a oplodnenia predstavujú medzičlánok spájajúci ontogenézu rodičov s ontogenézou ich potomstva.
Larválne obdobie v typickom variante sa pozoruje vo vývoji tých stavovcov, ktorých embryá vychádzajú z vaječných membrán a začínajú viesť samostatný životný štýl bez toho, aby dosiahli definitívne (zrelé) znaky organizácie. Nachádza sa teda u niektorých predstaviteľov nižších stavovcov - mihule, väčšiny kostnatých rýb a obojživelníkov. Najcharakteristickejšími znakmi larvy sú: embryonálna povaha jej organizácie, prítomnosť dočasných (dočasných) orgánov, skorý nástup fungovania mnohých orgánov, čo umožňuje nezávislú existenciu. Vďaka aktívnemu kŕmeniu dostane larva možnosť dokončiť vývoj a vďaka aktívnemu pohybu má možnosť zvoliť si podmienky prostredia optimálne pre vývoj a vyhnúť sa tak konkurencii s vlastnými dospelými príbuznými. U stavovcov je trvanie larválneho obdobia oveľa dlhšie ako embryonálne obdobie.
Metamorfóza spočíva v premene larvy na juvenilnú formu. V procese metamorfózy dochádza k takým dôležitým morfogenetickým transformáciám, ako je čiastočná deštrukcia, reštrukturalizácia a novotvorba orgánov. Stupeň premeny je tým väčší, čím viac
rozdiely medzi biotopom larvy a dospelého organizmu, čo dobre ilustruje príklad vývoja bezchvostých obojživelníkov (porovnaj: pulec a žaba).
Mladistvý obdobie začína od okamihu dokončenia metamorfózy (u placentárnych cicavcov a ľudí - od narodenia) a končí pubertou a začiatkom reprodukcie. Znaky juvenilného obdobia sa prejavujú v originalite výživy mladého organizmu, jeho správaní a stupni závislosti od rodičov. Z morfologického hľadiska je toto obdobie charakteristické intenzívnym rastom, nastoľovaním konečných proporcií medzi rôzne časti tela, dokončenie vývoja kostry, kože, výmena chrupu, dokončenie vývoja pohlavných žliaz a hormonálna regulácia. Trvanie juvenilného obdobia u stavovcov sa pohybuje od minimálne 13 – 18 dní u hrabošov po maximálne 18 – 20 rokov u belugy, krokodíla, albatrosa a slona. U mnohých predstaviteľov stavovcov, najmä u ľudí, môže byť dosiahnutie puberty a začiatok reprodukcie oddelený značným časovým úsekom.
Aplikácia ekologické a embryologické prístup nám umožňuje rozdeliť ontogenézu do štádií, ktoré sa vyskytujú pred narodením, počas A po narodení jednotlivcov. Samotný pôrod, t.j. uvoľnenie vyvíjajúceho sa jedinca zo škrupín vajíčka alebo z tela matky, v odlišné typy sa vyskytuje v rôznych štádiách zrelosti. Zároveň je u všetkých druhov pred narodením telo chránené vaječnými membránami alebo telom matky a nie je schopné samo sa živiť a vykonávať iné dôležité funkcie. Ochrana skorých, morfofunkčne nezrelých štádií zabezpečuje prežitie druhu. Po narodení jedinec nadviaže spojenie s novým prostredím, začne sa sám živiť, pohybovať sa a vykonávať všetky ostatné funkcie.
Podrobnejšie predstavenie periodizácie ľudskej ontogenézy, ktorá má praktickú hodnotu v pôrodníckej a pediatrickej praxi pozri článok 7.6.1 a tabuľku. 7.3.
7.2. morfofyziologické a evolučné znaky vajíčok strunatcov
7.2.1. PÔVOD VAJEC
Vajíčka strunatcov pozostávajú z vajíčka, niekedy nazývaného aj vajíčko, a vaječných blán. Oocyty tvorené v ženských pohlavných orgánoch
žľaza - vaječník. Prechádzajú dlhou cestou vývoja, ktorá začína v embryonálnom a pokračuje v aktívnom reprodukčnom období ontogenézy ženských jedincov.
Primárne pohlavné bunky veľmi skoro v embryogenéze sú izolované zo somatických buniek. Takže u bezchvostých obojživelníkov sa gonocyty izolujú v štádiu blastuly, u vtákov sa určujú počas tvorby primárneho hypoblastu, u cicavcov v epiblaste v štádiu gastruly, u obojživelníkov chvostových sa tento proces vyskytuje neskôr, v štádiu gastruly alebo aj neurula (v mezoderme). Potom sa tieto bunky pohybujú a nakoniec dosiahnu základy pohlavných žliaz. Cicavčie gonocyty sa najskôr ocitnú v extraembryonálnej oblasti pri ústí žĺtkového vaku a potom migrujú do anlage gonád (obr. 7.1). Migračné mechanizmy sú uvedené v článku 6.5.3.
Keď sa dostanú do pohlavných žliaz, primárne zárodočné bunky sa začnú množiť. Delia sa mitózou a sú tzv ovo(oo)gonia. Vo väčšine nižších stavovcov si ovo(oo)gonia zachováva schopnosť deliť sa počas celého reprodukčného obdobia, napríklad ryby uvoľňujú tisíce ikier na jeden výter, obojživelníky - stovky. U vyšších stavovcov počet vajíčok, ktoré dozrievajú v rovnakom čase, zriedka dosahuje 15, zvyčajne je ich menej, niekedy jedno, čím sa dajú vysvetliť aj znaky ovo(oo)genézy.
V ľudských ženských embryách sa ovo(oo)gonia najintenzívnejšie rozmnožuje medzi 2. a 5. mesiacom vnútromaternicového
Ryža. 7.1. Lokalizácia primárnych zárodočných buniek v ľudskom embryu v štádiu 16 somitov (a) a ich migrácia do pohlavných žliaz (b): 1 - predžalúdočné črevo; 2 - zadné črevo; 3 - alantois; 4 - primárne zárodočné bunky; 5 - žĺtkový vak; 6 - srdce; 7 - vyvíjajúca sa gonáda (genitálny tuberkul); 8 - primárna oblička; 9 - kloaka
vývojové obdobie, kedy ich počet dosahuje približne 7 miliónov.Do 7. mesiaca mnohé ovo(oo)gónie odumierajú a zvyšné vstupujú do profázy prvého delenia meiózy a zastavujú sa v štádiu diakinézy. S nástupom puberty ovuluje každý mesiac jeden oocyt, ktorý dosiahne štádium metafázy druhého delenia meiózy. U niektorých vajíčok k tomu dochádza v čase puberty a u iných tesne pred menopauzou. Oocyt dokončí meiózu iba vtedy, ak dôjde k oplodneniu.
7.2.2. ŠPECIFIKÁCIA A VÝZNAM CHEMICKÉHO ZLOŽENIA CYTOPLAZMY VAJEC
Zrelé vajíčko je spravidla väčšie ako ovo(oo)gonia a akékoľvek iné somatické bunky. Počas ovo(oo)genézy v cytoplazme vajíčka, veľké množstvo látky potrebné na jeho dozrievanie a skorú embryogenézu. Počas obdobia rastu sa teda objem myšacieho ovocytu zväčší asi 40-krát a objem žabieho oocytu sa zväčší viac ako 400-krát.. Rýchlosť syntézy látok v oocytoch sa výrazne zvyšuje v dôsledku mnohých znakov. V dozrievajúcich vajíčkach dochádza k amplifikácii (pozri časť 2.4.3.4) jednotlivých génov (pozri časť 4), napríklad génov rRNA. Obojživelníky (Xenopus laevis) asi 450 génov rRNA sa nachádza v diploidnej bunke a viac ako milión v ovo(oo)cyte. Okrem toho sa zvyšuje transkripčná aktivita genómu týchto buniek. Rýchlosť transkripcie v rastúcich myších oocytoch je teda 10-krát vyššia ako v somatických bunkách.
Funkčná úloha skladovaných látok je rôzna.
po prvé, sú to zložky potrebné pre procesy replikácie, transkripcie a translácie, ako sú zodpovedajúce enzýmy, ribozómy, mRNA, tRNA a ich prekurzory.
po druhé, ide o súbor špecifických regulačných látok, ktoré zabezpečujú koordinované fungovanie všetkých uložených komponentov. Medzi tieto látky patrí faktor rozpadu jadrovej membrány (deštrukciou jadrovej membrány začína prometafáza 1. delenia meiózy); faktor, ktorý spôsobuje kondenzáciu chromozómov; faktor, ktorý premieňa jadro spermie na pronukleus a aktivuje v ňom syntézu DNA pred štiepením; cytostatický faktor csf, zodpovedný za blok meiózy v štádiu metafázy II (u mnohých stavovcov k odstráneniu tohto bloku dochádza až v dôsledku oplodnenia); cytoskeletálny mikrofilamentový systém zapojený do udržiavania bloku
meióza; cyklické nukleotidy (napr. cAMP), proteínkinázy a ďalšie látky potrebné na signalizáciu z okolitých ovo(oo)cyt buniek a extracelulárneho prostredia; zložky takzvaného inozitolfosfátového systému, ktoré zabezpečujú aktiváciu vajíčka po oplodnení; faktor zodpovedný za cytotómiu počas štiepenia. Niektoré z nich, ktoré sú v oocyte už v čase oplodnenia, začínajú pôsobiť až vo fáze gastrulácie.
po tretie, je to žĺtok, ktorý zahŕňa bielkoviny, fosfolipidy, neutrálne tuky, sacharidy, minerálne soli. Žĺtok je zásobárňou živín a energetických zdrojov potrebných na zabezpečenie embryonálneho obdobia. Mnohé látky produkované pečeňou vstupujú do ovo(oo)gónie počas svojho rastu cez folikulárne bunky vaječníka. To si od samice vyžaduje veľký energetický výdaj.
po štvrté, Ide o špecifické látky tzv morfogenetické determinanty, dosť rigidne určujúce osud buniek (blastomér), do ktorých spadajú pri štiepení. Podobné látky sa nenachádzajú v cytoplazme vajíčok u všetkých zvierat.
Kvôli opísaným vlastnostiam chemického zloženia cytoplazmy vajíčka embryo počas obdobia drvenia v niektorých prípadoch nevyužíva genetickú informáciu jadier zygoty a blastomérov na vývoj. Špecifické chemické zloženie a pravidelná distribúcia látok v cytoplazme vajíčka majú veľký význam pre počiatočné fázy embryogenézy. Zásobné živiny a energetické látky zabezpečujú embryonálny vývoj bez ich dodatočného príjmu zvonku.
7.2.3. VEĽKOSŤ VAJEC A ICH ÚLOHA VO EVOLÚCII. TYPY VAJEC
V procese vývoja sa odhaľuje pravidelnosť, ktorá spočíva v tom, že čím dlhšie je embryonálne obdobie, tým viac žĺtka by sa malo nahromadiť vo vajci. Trvanie embryonálneho obdobia závisí od štádia, v ktorom embryo prechádza do samostatnej existencie vo vonkajšom prostredí. Ak je postembryonálny vývoj priamy, t.j. bez larvy a metamorfózy, potom by malo byť vo vajci viac žĺtka. Podľa počtu žĺtkov sa vajíčka strunatcov (tab. 7.1) delia na alecitálny, oligo-, mezo- A polylecitálny, tie. so zanedbateľným malým, malým, stredným a veľkým množstvom žĺtka (z gr. lecitos- žĺtok) (tabuľka 7.2).
Tabuľka 7.1.Systematika typu Chordata
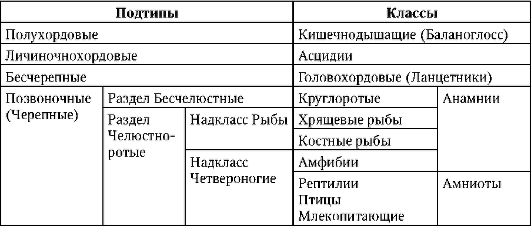
Tabuľka 7.2.Typy oocytov nachádzajúce sa v chordátoch

o lancelet, zástupca nižších strunatcov, vajce oligolecitálny. U väčšiny stavovcov obsahujú vajíčka značné množstvo žĺtka. Medzi nižšími stavovcami (Anamnia) najväčšie ikry sa nachádzajú v hagfishes (trieda Cyclostomes), v žralokoch a chimérach (trieda chrupavkovité ryby) a v niektorých obojživelníkoch. Zvyšok obojživelník, ako aj vajíčka jesetera mesolecitálny, tie. mať priemerné množstvo žĺtka. U vyšších stavovcov (Amniota) ako sú plazy, vtáky a vajcorodé cicavce, polylecitál, tie. vo vajci je veľa žĺtka. Ich embryonálny vývoj trvá obzvlášť dlho.
Tento vzor je porušený u vačnatcov a placentárne cicavce, ktoré majú oligo- A alecitál vajcia, resp. U vačkovcov embryo vychádza z vaječných membrán a maternice s neúplnou organogenézou, je prenesené do vačku, kde pokračuje vo vývoji. V placentách, vrátane ľudí, embryo vychádza
z vaječných membrán ešte skôr, v štádiu blastocysty, ale potom prechádza do vnútromaternicovej existencie, kde absolvuje všetky hlavné obdobia vývoja, ktoré ju pripravujú na pôrod. Znížiť množstvo žĺtka vo vajciach cicavcov možno tzv sekundárny, pretože ich predkovia, ktorí ovládali pozemné prostredie, mal, ako všetky amnioty, polylecitálne vajíčka.
7.2.4. POLARITA vajíčok
Polarita oocytov plánuje sa ešte v štádiu akumulácie žĺtka v ovo(oo)cytoch pri ich rýchlom (veľkom) raste a fixuje sa pri izolácii polárnych (redukčných) teliesok. Po izolácii druhého redukčného telesa sa polarita stáva stabilnou a nezvratnou, čo dokazujú Guerrierove experimenty na odstreďovaní vajíčok v rôznych štádiách ich dozrievania. Pól, pri ktorom vystupujú redukčné telesá, sa nazýva zviera, a opak je vegetatívny. Póly vaječnej bunky sa líšia mnohými spôsobmi: koncentráciou rôzne látky, počet organel, aktivita vnútrobunkových procesov a množstvo ďalších. Áno, experimenty v posledných rokoch pomocou vibračných elektród odhalené elektrické polia okolo oocytov a vajíčok množstva živočíchov a rastlín a tok elektrických prúdov cez ich cytoplazmu. Predpokladá sa, že je to spôsobené rôznymi koncentráciami iónových kanálov a púmp na opačných póloch oocytov. Vo vajíčku žaby pazúrikovanej je koncentrácia iónových kanálov vyššia na zvieracom póle a koncentrácia púmp je vyššia na vegetatívnom póle.
Hromadenie žĺtka vajcom- prvý prejav jeho polarizácie. S malým množstvom žĺtka vo vajci je zvyčajne rovnomerne distribuovaný v cytoplazme a jadro je umiestnené približne v strede. Tieto vajíčka sú tzv izolecitál(z gréčtiny. isos- rovný). Väčšina stavovcov má veľa žĺtka a je nerovnomerne distribuovaný v cytoplazme vajíčka. Toto anisolecithal bunky. Väčšina žĺtka sa hromadí v vegetatívny pól. Tieto vajíčka sú tzv telolecitálny(z gréčtiny. telos- koniec). Smerom opačne zviera aktívna cytoplazma bez žĺtka je odsunutá nabok k pólu. Ak je žĺtok stále ponorený v cytoplazme a nie je z nej izolovaný ako samostatná frakcia, ako u jeseterov a obojživelníkov, vajíčka sa nazývajú stredne telolektálny. Ak je žĺtok úplne oddelený od cytoplazmy, ako u amniotov, potom toto ostro telolecitálne vajcia.
Vlastnosti veľkosti a polarity vajíčok strunatcov sú schematicky zoskupené v tabuľke. 7.2.
V procesoch polarizácie vajíčka sa zjavne zúčastňuje a kôra- Ide o povrchovú vrstvu cytoplazmy vajíčka, ktorá sa nachádza priamo pod plazmatickou membránou. Obsahuje mikrofilamenty a kortikálne granuly. Tie obsahujú celý rad látok podieľajúcich sa na tvorbe oplodňovacej membrány a zabraňujúcich polyspermii. Zhromažďovanie a redistribúcia prvkov cytoskeletu kortikálnej vrstvy pozorovaná po oplodnení zabezpečuje jej získanie kontraktility, ktorá je potrebná na rozdelenie zygotového štiepenia. U väčšiny živočíchov prvé dve štiepne brázdy prebiehajú po vzájomne kolmých živočíšno-vegetatívnych rovinách (meridiány spájajúce živočíšne a vegetačné póly).
Vo všeobecnosti platí, že na zvieracom póle vajíčka je spravidla viac voľnej cytoplazmy, organel, uloženej RNA, zvyčajne sa jadro nachádza aj na zvieracom póle alebo bližšie k nemu. Vegetatívny pól je charakterizovaný prevahou žĺtkových granúl.
Polarizáciu vajíčka sprevádza vzhľad ovo(oo)-plazmatická segregácia vajcia, t.j. vytvorenie vnútornej heterogenity rezov cytoplazmy vajíčka.
Živočíšno-vegetatívna polarizácia vajíčka má rozhodujúci význam pre všetky nasledujúce procesy embryogenézy, pretože určuje budúcu priestorovú organizáciu embrya. U dospelých zvierat sa predozadná os tela zhoduje so zvieravo-vegetatívnou osou vajíčka (napríklad u stavovcov) alebo je na ňu kolmá (napríklad u máloštetinavcov a niektorých článkonožcov).
7.2.5. VAJEČNÉ škrupiny
Vajíčka sú z vonkajšej strany pokryté jednou alebo viacerými membránami, ktoré neskôr plnia okrem iného aj funkciu ochrany vyvíjajúceho sa embrya.
Rozlišovať primárny plášť, tvorené samotným vajíčkom sekundárny plášť- produkt činnosti ovariálnych folikulárnych buniek, a terciárne škrupiny, ktorým je vajíčko pri prechode vajcovodom obklopené.
primárny plášť, niekedy tzv žĺtok, nachádza vo vajciach všetkých zvierat. U stavovcov, vrátane cicavcov,
Primárny obal je súčasťou hustého obalu, ktorý tvorí jeho vnútornú časť. Vonkajšia časť hustého plášťa produkujú folikulárne bunky sekundárny plášť. Hustá škrupina je preniknutá zvnútra mikroklkami vajíčka a zvonka mikroklkami folikulárnych buniek. Pre ich optické vlastnosti u cicavcov je tzv lesklá škrupina(zona pellucida). Takže táto šupka kombinuje primárne a sekundárne. Na vrchole zona pellucida je žiarivá koruna(corona radiata), vytvorený z folikulárnych buniek, ktoré priľnú k vajíčku, kým je vo folikule vaječníka (obr. 7.2).
Zona pellucida je komplexný komplex extracelulárnych glykoproteínov, ktoré sa u cicavcov označujú ZP. U myší sa skladá z troch rôznych sulfátovaných glykoproteínov – ZP1, ZP2, ZP3. Používaním ZP3 spermie sa viažu na zona pellucida a po penetrácii jednej spermie je tento glykoproteín modifikovaný a penetrácia ďalšej
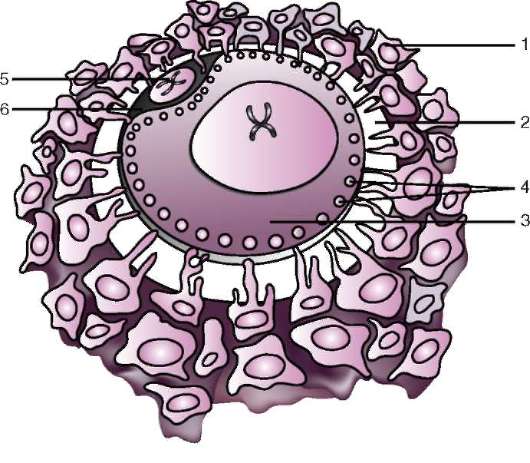
Ryža. 7.2. Vajíčko (ovocyt I. rádu) cicavcov: 1 - žiarivá koruna; 2 - lesklá škrupina; 3 - cytoplazma; 4 - kortikálne granule; 5 - polárne teleso; 6 - periyolkový priestor
gih spermie sa stáva nemožným. Okrem toho sú glykoproteíny druhovo špecifické, čo zabraňuje medzidruhovému oplodneniu. U ostatných strunatcov je druhová špecifickosť oplodnenia určená interakciou spermií s receptormi. žĺtková membrána. Zona pellucida neprilieha priamo k blane vajíčka, ale oddeľuje sa perivitelínový (periyotelový) priestor. Po preniknutí prvej spermie do vajíčka sa do tohto priestoru dostane kortikálna granula ovoperoxidáza. Predpokladá sa, že pôsobenie tohto enzýmu sa mení ZP3 A Zp2,čo vedie k tvrdnutiu zona pellucida. Pretrváva okolo embrya počas celého predimplantačného obdobia alebo vo významnej časti tohto obdobia. Lesklá škrupina zabraňuje zlepeniu susedných embryí a prilepeniu embryí na steny vajcovodu a maternice. Je známe, že v počiatočných štádiách drvenia až po blastocystu majú blastoméry vysokú priľnavosť. Ak sa embryo bez zona pellucida transplantuje do vajcovodu, blastoméry sa nevyhnutne prilepia na stenu vajcovodu a embryo zomrie. Navyše vďaka zona pellucida sú blastoméry kompaktné a usporiadané, čo podporuje vytváranie kontaktov a interakciu medzi nimi a zabezpečuje normálny vývoj embrya v tomto štádiu. Ak sa škrupina odstráni, fragmentácia bude pokračovať, ale blastoméry budú usporiadané v reťazci a ich zhutnenie bude úplne narušené alebo bude veľmi neskoro.
Treťohorné mušle dobre vyvinuté u chrupavčitých rýb a obojživelníkov, ale zvláštnu zložitosť nadobúdajú u suchozemských stavovcov - plazov, vtákov a nižších cicavcov. Formovanie zo sekrétov žliaz vajcovodu, tieto schránky nemajú bunkovú štruktúru. U všetkých stavovcov plnia funkcie ochrany embrya pred mechanickým poškodením a pôsobením škodlivých biotických faktorov, ako sú bakteriálne, plesňové a protozoálne faktory. Okrem toho sa u suchozemských stavovcov objavujú zásadne nové funkcie skladovania vody a živín, aby uspokojili potreby embrya. U plazov škrupinová membrána funguje ako čerpadlo, odoberá vodu z pôdy a vzduchu. Vtáky majú zásobu vody proteínová škrupina. Absorpcia a odparovanie vody je regulované pórmi v membráne plášťa. Škrupina obsahuje veľa minerálnych solí potrebných na vývoj kostry embrya.
7.3. HNOJENIE A PARTENOGENÉZA
Hnojenie- ide o proces fúzie zárodočných buniek, ktorý vyvrcholí zjednotením ich genetického materiálu. Diploidná bunka, ktorá je výsledkom oplodnenia zygota- predstavuje počiatočné štádium vývoja nového organizmu.
Proces hnojenia pozostáva z troch po sebe nasledujúcich fáz:
Vzdialená interakcia a konvergencia gamét;
Kontaktná interakcia gamét a aktivácia vajíčka;
Fúzie gamét alebo syngamie.
Aproximácia spermií s vajíčkom poskytovaná kombináciou nešpecifických faktorov, ktoré zvyšujú pravdepodobnosť ich stretnutia a interakcie. Patrí medzi ne koordinácia nástupu pripravenosti na oplodnenie u samcov a samíc, správanie samcov a samíc, ktoré zabezpečuje kopuláciu a insemináciu, nadmerná produkcia spermií, veľká veľkosť vajíčka, ako aj spermií produkovaných vajíčkami a spermiami. chemických látok, čo prispieva k zbližovaniu a interakcii zárodočných buniek. Tieto látky, tzv gamóny(hormóny gamét) na jednej strane aktivujú pohyb spermií a na druhej strane ich väzbu. Pri pohybe cicavčích spermií pozdĺž horných častí vajcovodu je fenomén reotaxia: ich schopnosť pohybovať sa proti prichádzajúcemu toku tekutiny vo vajíčkovodoch.
U cicavcov má pobyt spermií v ženskom pohlavnom trakte veľký význam, v dôsledku čoho kapacita spermií- ich nadobudnutie schopnosti hnojenia. Ihneď po vstupe do pohlavného traktu nie sú ženské spermie schopné preniknúť do vajíčok. Glykoproteíny a proteíny semennej plazmy sa odstraňujú z plazmolemy spermií v oblasti akrozómu pôsobením látok tajomstva ženského pohlavného traktu, ktoré blokujú aktívne centrá receptorových molekúl spermiovej plazmolemy, rozpoznávajúc povrch samice. reprodukčná bunka. Okrem toho sa molekuly albumínu nachádzajúce sa v ženskom pohlavnom trakte viažu na cholesterol membrány spermií. To vedie k destabilizácii plazmalemy spermií a jej akrozomálnej membrány, čo uľahčuje následné uvoľňovanie akrozomálnych enzýmov. Zistilo sa tiež, že v procese kapacity sa menia vlastnosti povrchu spermií, ako napríklad ich náboj. Okrem toho dochádza k zmene mobility (hyperaktivácii) spermií a aktivácii akrozomálnych enzýmov. U ľudí trvá kapacita kapacity asi 7 hodín.
Vo fáze kontaktná interakcia spermie ničia membrány vajíčka (u cicavcov - žiarivá korunka, priehľadná membrána) a cytoplazmatickú membránu oocytu v dôsledku akrozómová reakcia. Pri kontakte so schránkou ženskej zárodočnej bunky pôsobením jej aktivačných látok (jednou z nich je oplodnenie) sa spustí aktívny tok vápenatých katiónov do hlavičky spermie. Výsledkom je fokálna fúzia bunkových a akrozomálnych membrán spermií a ich čiastočná deštrukcia. Prostredníctvom vytvorených mikrootvorov sa uvoľňujú spermie enzýmy - hyaluronidáza, penetráza a iné, ktoré rozpoja kontakty medzi bunkami žiarivej korunky, ako aj medzi nimi a oocytom. Akrozomálny enzým akrozín ničí časť zona pellucida ženskej zárodočnej bunky a spermie prenikajú do periyolkového priestoru. V mieste kontaktu hlavičky spermie s plazmatickou membránou oocytu dochádza k fúzii a následnej deštrukcii membrán ženských a mužských gamét. Prostredníctvom výsledného cytoplazmatického mostíka sa spája cytoplazma oboch gamét. Potom jadro a centriol spermie prechádzajú do cytoplazmy vajíčka a membrána spermie je zabudovaná do membrány vaječnej bunky. Chvostová časť spermie buď zostane vonku, alebo tiež vstúpi do vajíčka, ale potom sa oddelí a rozloží bez toho, aby zohrávala akúkoľvek úlohu v ďalšom vývoji.
Sodné ióny začnú aktívne vstupovať do cytoplazmy vajíčka cez membránu spermií, v dôsledku čoho sa membránový potenciál oocytu dramaticky zmení a ženská zárodočná bunka sa stane imúnnou voči kontaktu s inými spermiami - rýchly blok polyspermie. Prílev sodíkových iónov spôsobuje uvoľňovanie iónov vápnika z vnútrobunkových depotov a zvýšenie jeho obsahu v cytoplazme vajíčka, ktoré sa šíri vo forme vlny z miesta kontaktu gamét. Toto začína kortikálna reakcia: membrány kortikálnych granúl splývajú s membránou vajíčka a z nich uvoľnené proteolytické enzýmy sa dostávajú do periyolkového priestoru. Pod vplyvom enzýmov sa škrupina vajíčka zahusťuje, zahusťuje, stráca receptorové proteíny pre spermie a mení sa na oplodňovacia škrupina. Okrem toho glykoproteín vylučovaný z kortikálnych granúl prispieva k oddeleniu žĺtkovej membrány od plazmolemy vajíčka. V dôsledku všetkých vyššie uvedených skutočností je penetrácia iných spermií nemožná - pomalý blok polyspermie. U cicavcov
kortikálna reakcia nespôsobí tvorbu oplodňovacej membrány, ale jej podstata je rovnaká.
V dôsledku kontaktu spermie s vajíčkom je to aktivácia. Spočíva v zložitých štrukturálnych a fyzikálno-chemických zmenách. Počiatočným štádiom aktivácie je kortikálna reakcia opísaná vyššie. U zvierat, ako sú ostnokožce, kostnaté ryby a obojživelníky, sú zmeny v cytoplazme vajíčka sprevádzané viditeľnými morfologickými prestavbami. Tieto javy sa nazývajú stratifikácia resp plazmová segregácia. Jeho význam pre ďalší embryonálny vývoj bude diskutovaný nižšie. U mnohých druhov anuránov vedie prienik spermie do vajíčka k pohyb pigmentových granúl zvieracieho pólu a proti miestu prieniku spermií sa objaví slabo sfarbená oblasť v tvare polmesiaca, tzv "sivý kosák"(obr. 7.3).
Aktivácia vajíčka končí začiatkom syntézy proteínov na translačnej úrovni, keďže mRNA, tRNA, ribozómy a energia boli uložené späť v ovo(oo)genéze. Aktivácia vajíčka môže začať a pokračovať až do konca bez jadra spermie a bez jadra vajíčka, čo bolo dokázané experimentmi na enukleácii zygoty.
Vajíčko je v čase stretnutia so spermiou zvyčajne v jednom zo štádií meiózy, blokované špecifickým faktorom. U väčšiny stavovcov sa tento blok vykonáva
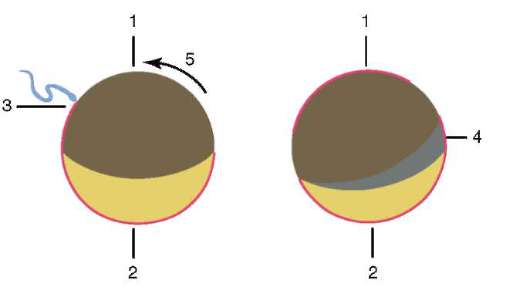
Ryža. 7.3. Segregácia cytoplazmy vajíčka obojživelníka po preniknutí spermiou: 1 - zvierací pól; 2 - vegetatívny pól; 3 - miesto zavedenia spermií; 4 - sivý kosák; 5 - smer pohybu cytoplazmy s pigmentovými granulami
v štádiu metafázy II; u mnohých bezstavovcov, ako aj u troch druhov cicavcov (kone, psy a líšky) sa blok vyskytuje v štádiu diakinézy (profáza I). Vo väčšine prípadov sa meiotický blok odstráni po aktivácii vajíčka v dôsledku oplodnenia. Kým je vo vajíčku dokončená meióza, modifikuje sa jadro spermie, ktoré do neho preniklo. Má formu medzifázového a potom profázneho jadra. Počas tejto doby sa DNA zdvojnásobí a množstvo dedičný materiál V mužský pronukleus sa stáva p2s, tie. obsahuje haploidný súbor reduplikovaných chromozómov.
Jadro vajíčka, ktoré dokončilo meiózu, sa stáva ženský pronucleus, aj nadobúdanie p2s. Obe pronukleá robia zložité pohyby, potom sa k sebe priblížia a splynú. (syncarion), tvoriaci spoločnú metafázovú platňu. Toto je v skutočnosti moment konečnej fúzie gamét - syngamia. Prvé mitotické delenie zygoty vedie k vytvoreniu dvoch embryonálnych buniek (blastomérov) so sadou chromozómov 2p2s vo všetkých.
V niektorých prípadoch sa vývoj vyskytuje bez oplodnenia - partenogenéza(z gréčtiny. parthenoe- panna). Kedy prirodzená partenogenéza vývoj prebieha na základe cytoplazmy a pronuklea vajíčka. Jedinci, ktorí sa tvoria z vajíčka, majú buď haploidnú alebo diploidnú sadu chromozómov, pretože sa spúšťa jeden z mechanizmov zdvojnásobenia počtu chromozómov. V niektorých prípadoch počas meiózy ženskej zárodočnej bunky vypadne štádium zníženia počtu chromozómov a vajíčko sa získa s diploidným pronukleom. V iných prípadoch dochádza k diploidizácii počas prvého delenia štiepenia, pri ktorom nedochádza k cytotómii.
prirodzená partenogenéza, spravidla nie je jediným spôsobom reprodukcie druhu. Buď sa strieda s normálnym pohlavným rozmnožovaním, alebo sa vyskytuje u jedinca Preteky. Prirodzená partenogenéza bola zistená v letných generáciách niektorých kôrovcov a vírnikov, u včiel, ôs a mnohých Lepidoptera. Medzi stavovcami bola opísaná partenogenetická reprodukcia v troch rasách skalného jaštera z Arménska, ktoré pozostávali iba zo samíc. 40% morčacích vajec znesených v neprítomnosti samca sa môže začať vyvíjať, ale tento vývoj sa málokedy dotiahne do konca, častejšie sa zastaví kvôli anomáliám. U iných druhov stavovcov nie je prirodzená partenogenetická reprodukcia známa.
umelá partenogenéza Zdá sa, že je to možné u všetkých zvierat. Dôležitý je vývoj metód pre partenogenetický vývoj
problém vo vedeckých a aplikovaných vzťahoch. Veľký prínos k tomuto problému mali domáci výskumníci A.A. Tichomirov, B.L. Astaurov, V.A. Strunnikov. Zistilo sa, že aktivácia vajíčka spermiou nie je špecifická. Ako aktivátory môžu pôsobiť mnohé fyzikálne a chemické faktory. Na priadke morušovej sa ukázalo, že pomocou umelej partenogenézy je možné regulovať pomer mužského a ženského pohlavia v populácii, čím sa dosiahne veľký ekonomický efekt.
K prirodzenej partenogenéze dochádza najčastejšie pri neúplnom oplodnení, t.j. v tých prípadoch, keď došlo k aktivácii vajíčka, ale jadro spermie sa nezúčastnilo oplodnenia. V aktivovaných vajíčkach sa využíva iba informácia zo samičieho pronuklea. Tento typ partenogenézy sa nazýva gynogenéza. Pri umelej partenogenéze je možné odstrániť ženský pronukleus a potom sa vývoj uskutoční iba na úkor mužských pronukleov. Toto an-drogenéza. Pri špeciálnych pokusoch na ježovkách sa zistilo, že potomstvo pri gynogenéze dedí buď len črty matky, alebo počas androgenézy len črty otca. To naznačuje, že dedičné vlastnosti jedinca sú určené hlavne jadrom a nie cytoplazmou.
7.4. embryonálny vývoj
7.4.1. ROZDELENIE SA
7.4.1.1. Podstata štádia drvenia
Rozdelenie- ide o sériu postupných mitotických delení zygoty a výsledných blastomér, ktoré končia tvorbou mnohobunkové jednovrstvové embryo- blastula. Prvé rozdelenie štiepenia začína po spojení dedičného materiálu pronuklea a vytvorení spoločnej metafázovej platničky. Bunky vzniknuté pri štiepení sa nazývajú blastoméry(z gréčtiny. výbuch- klíčok, klíčok). Znakom mitotických cyklov drvenia je, že s každým delením sa bunky zmenšujú a zmenšujú, až kým nedosiahnu pomer objemov jadra a cytoplazmy, ktorý je obvyklý pre somatické bunky. Napríklad u morského ježka to vyžaduje 6 delení a embryo pozostáva zo 64 buniek. Medzi po sebe nasledujúcimi deleniami nedochádza k rastu buniek (obdobie G1 chýba), ale DNA sa nevyhnutne syntetizuje. Všetky prekurzory DNA a potrebné
Stmievateľné enzýmy sa hromadia v procese oogenézy. V dôsledku toho sa mitotické cykly skracujú a delenia nasledujú po sebe oveľa rýchlejšie ako v bežných somatických bunkách. Po niekoľkých cykloch delenia vzniknuté blastoméry na seba nadväzujú a vytvárajú zhluk buniek tzv morula. Potom sa medzi bunkami vytvorí dutina - blastocoel, naplnené kvapalinou. Bunky sú vytlačené na perifériu a tvoria stenu blastuly - blastoderm. V dôsledku absencie rastu blastomér počas obdobia štiepenia celková veľkosť embrya v štádiu blastuly nepresahuje veľkosť zygoty.
7.4.1.2. Morfológia drvenia
Spravidla sú blastoméry usporiadané v prísnom poradí voči sebe navzájom a voči polárnej osi vajíčka. Poradie alebo spôsob drvenia závisí od množstva, hustoty a rozloženia žĺtka vo vajci. Podľa pravidiel Sachs-Hertwigovej má bunkové jadro tendenciu byť umiestnené v strede cytoplazmy bez žĺtka a vreteno bunkového delenia - v smere najväčšieho rozsahu tejto zóny.
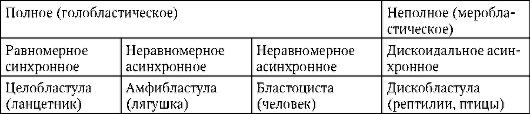
V oligo- a mesolecitálnych vajíčkach štiepenie kompletný, alebo holoblastický. Tento typ drvenia sa vyskytuje u lanceletov, mihule, niektorých rýb, všetkých obojživelníkov, ako aj u vačnatcov a placentárnych cicavcov. Pri úplnom rozdrvení zodpovedá rovina prvého delenia rovine obojstrannej symetrie. Rovina druhého delenia prebieha kolmo na rovinu prvého. Obe brázdy prvých dvoch divízií sú poludníkové, t.j. začínajú na zvieracom póle a šíria sa k vegetatívnemu pólu. Vaječná bunka je rozdelená na štyri blastoméry, ktoré majú viac-menej rovnakú veľkosť. Rovina tretieho delenia prebieha kolmo na prvé dve v smere zemepisnej šírky. U zvierat s izolecitálnymi vajíčkami je všetkých osem vytvorených blastomér približne rovnakých - rovnomerné drvenie. V mezolecitálnych vajíčkach sa objavuje nezrovnalosť drvenie. Na zvieracom póle sú štyri menšie blastoméry - mikrometre, na vegetatívnom - štyri väčšie - makroméry. Potom rozdelenie opäť ide v rovinách poludníka a potom znova v rovinách zemepisnej šírky.
V polylecitálnych oocytoch kostnatých rýb, plazov, vtákov a tiež monotrémnych cicavcov štiepenie čiastočný, alebo meroblastický, tie. pokrýva iba cytoplazmu bez žĺtka. Nachádza sa vo forme tenkého disku pri zvieracom póle, preto sa tento typ drvenia nazýva tzv diskoidný.
Pri charakterizácii typu drvenia sa berie do úvahy aj relatívna poloha a rýchlosť delenia blastomérov. Ak sú blastoméry usporiadané v radoch nad sebou pozdĺž polomerov, nazýva sa drvenie radiálne. Je typický pre strunatce a ostnatokožce. V prírode existujú aj iné varianty priestorového usporiadania blastomér pri drvení, ktoré určuje jej typy, ako napr. špirála u mäkkýšov bilaterálne pri škrkavke, anarchický pri medúze.
Bol zaznamenaný vzťah medzi distribúciou žĺtka a stupňom synchronizácie pri delení živočíšnych a vegetatívnych blastomér. V oligolecitálnych vajíčkach ostnatokožcov a hlavonožcov je štiepenie takmer synchrónne, u mezolecitálnych vajec napríklad u obojživelníkov je synchronizácia narušená po treťom delení, pretože vegetatívne blastoméry sa delia pomalšie kvôli veľkému množstvu žĺtka. V organizmoch s čiastočným štiepením (plazy, vtáky) sú delenie od samého začiatku asynchrónne a blastoméry zaujímajúce centrálnu polohu sa delia rýchlejšie.
Na konci drvenia, blastula. Typ blastuly závisí od typu drvenia, a teda od typu vajíčka. Niektoré typy drvenia a blastuly sú znázornené na obr. 7.4 a schéma (v tabuľke) 7.1. Viac Detailný popisštiepenie u cicavcov vrátane ľudí, pozri časť 7.5.1.
Tabuľka 7.3.Druhy drvenia a druhy strunatcových blastúl
7.4.1.3. Vlastnosti molekulárno-genetických a biochemických procesov pri drvení
Ako je uvedené vyššie, mitotické cykly počas obdobia štiepenia sú značne skrátené, najmä na samom začiatku. Napríklad celý cyklus štiepenia u vajíčok morského ježka trvá 30 – 40 minút s trvaním S-fázy len 15 min.
Gj-, a u mnohých zvierat G 2- obdobia prakticky chýbajú, keďže potrebná zásoba všetkých látok sa vytvorila v cytoplazme vajíčka a čím väčšia, tým väčšia. Ukázalo sa, že po odstránení zo zygoty
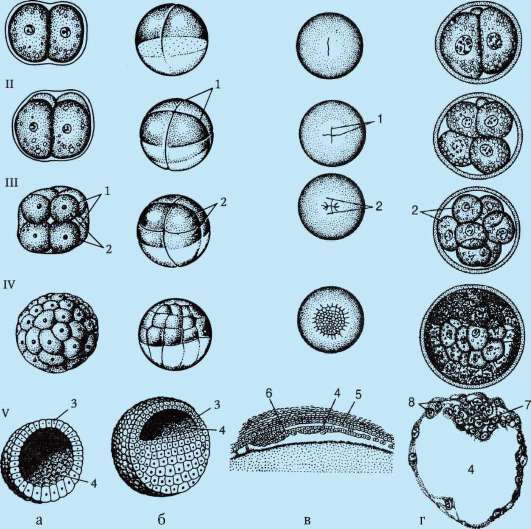
Ryža. 7.4.Štiepenie u strunatcov s rôznymi druhmi vajec: a - lancelet; b - žaba; c - vták; d - cicavec: 1 - 2 blastoméry; II - 4 blastoméry; III - 8 blastomérov; IV - morula; V - blastula; 1 - drvenie brázd; 2 - blastoméry; 3 - blastoderm; 4 - blastocoel; 5 - epiblast; 6 - hypoblast; 7 - embryoblast; 8 - trofoblast; veľkosť embryí na obrázku neodráža skutočnú veľkosť embryí
dochádza k odštiepeniu jadra a embryo sa dostáva vo svojom vývoji takmer do štádia blastuly. Ďalší vývoj sa zastaví.
Pred každým delením dochádza k syntéze DNA a histónov, avšak trvanie S-obdobie skrátené. Počas fragmentácie počas replikácie DNA je rýchlosť postupu replikačnej vidlice normálna. Zároveň je v blastomérovej DNA viac iniciačných bodov ako v
somatické bunky, preto sú replikóny kratšie. Syntéza DNA prebieha vo všetkých replikónoch súčasne (synchrónne). Preto sa čas replikácie DNA v jadre zhoduje s časom zdvojenia jedného, navyše skráteného, replikónu.
Na začiatku štiepenia prakticky chýbajú iné typy jadrovej aktivity, ako je transkripcia. V rôznych typoch vajíčok začína génová transkripcia a syntéza RNA v rôznych štádiách. V prípadoch, keď je v cytoplazme veľa rôznych látok, ako napríklad u obojživelníkov, sa transkripcia neaktivuje okamžite. Syntéza RNA v nich začína v štádiu ranej blastuly. Naopak, u cicavcov sa syntéza RNA začína už v štádiu dvoch blastomér.
Počas obdobia štiepenia sa tvorí RNA a proteíny, podobné tým, ktoré sa syntetizujú počas oogenézy. Väčšinou históny, bielkoviny bunkové membrány a enzýmy potrebné na delenie buniek. Tieto proteíny sa okamžite použijú spolu s proteínmi predtým uloženými v cytoplazme vajíčok. Spolu s tým je počas obdobia štiepenia možná syntéza proteínov, ktoré predtým neboli možné, čo dokazujú údaje o prítomnosti regionálnych rozdielov v syntéze RNA a proteínov medzi blastomérmi. Niekedy tieto látky začnú pôsobiť v neskorších štádiách.
Dôležitú úlohu pri drvení hrá delenie cytoplazmy - cytotómiu. Má osobitný morfogenetický význam, pretože určuje typ drvenia. V procese cytotómie sa najskôr vytvorí zúženie pomocou kontraktilného prstenca mikrofilamentov. Zostavenie tohto prstenca prebieha pod priamym vplyvom pólov mitotického vretienka. Po cytotómii zostávajú blastoméry oligolecitálnych vajíčok navzájom spojené len tenkými mostíkmi. Práve v tomto čase sa dajú najľahšie oddeliť. Je to preto, že cytotómia vedie k zníženiu oblasti kontaktu medzi bunkami v dôsledku obmedzeného povrchu membrán.
Ihneď po cytotómii začína syntéza nových častí bunkového povrchu, zvyšuje sa kontaktná zóna a blastoméry sa začínajú tesne dotýkať, čo poskytuje možnosť medzibunkovej interakcie. Po hraniciach medzi jednotlivými úsekmi ovoplazmy prebiehajú štiepne ryhy, preto sa cytoplazma rôznych blastomér líši chemickým zložením. To odráža tento jav ovoplazmatická segregácia.
Hlavným výsledkom zdrvujúceho obdobia je premena jednobunkového embrya – zygoty – na mnohobunkový A jednovrstvový embryo - blastula. Okrem toho pri drvení dochádza k tvorbe
ing medzibunkové kontakty a zväčšenie ich plochy, distribúcia rôznych úsekov cytoplazmy zygoty medzi rôzne blastoméry embrya, postupná aktivácia vlastného genómu embrya, obnovenie jadrovo-cytoplazmatického pomeru charakteristického pre somatické bunky. V štádiu drvenia sú všetky bunky embrya homogénne vo vzťahu k funkčnému stavu genetického aparátu. Rozdiel v cytoplazme blastomér a medzibunkové interakcie určujú (predurčujú) smer ďalšieho vývoja bunky.
7.4.2. GASTRULATION
7.4.2.1. Podstata štádia gastrulácie
Podstatou štádia gastrulácie je, že jednovrstvové embryo – blastula – sa mení na viacvrstvový- dva- alebo trojvrstvový, volal gastrula(z gréčtiny. gaster- žalúdok a zdrobnená prípona „st“).
U primitívnych strunatcov, napríklad v lancelete, sa homogénna jednovrstvová blastoderm počas gastrulácie premení na vonkajšiu zárodočnú vrstvu - ektodermu- a vnútorná zárodočná vrstva - endoderm. Endoderm tvorí primárne črevo s dutinou vo vnútri - gastrocoel. Otvor vedúci ku gastrocoelu je tzv blastopor alebo primárne ústa. Dve zárodočné vrstvy definujú morfologický znak gastrulácia. Ich existencia v určitom štádiu vývoja u všetkých mnohobunkových živočíchov, od koelenterátov až po vyššie stavovce, nám umožňuje uvažovať o homológii zárodočných vrstiev a jednote pôvodu všetkých týchto živočíchov.
U stavovcov sa pri gastrulácii tvorí okrem dvoch spomínaných zárodočných vrstiev aj tretia - mezoderm, nachádza sa medzi ekto- a endodermou. Vývoj strednej zárodočnej vrstvy, ktorá je chordomesoderm,- evolučná komplikácia fázy gastrulácie u stavovcov, spojená s urýchlením ich vývoja v raných štádiách embryogenézy. U primitívnejších strunatcov, ako je lancelet, sa chordomesoderm zvyčajne vytvára na začiatku fázy nasledujúcej po gastrulácii - organogenéza. Posun v čase vývoja niektorých orgánov v porovnaní s inými u potomkov v porovnaní so skupinami predkov je prejavom heterochrónia. Zmeny v načasovaní formovania najdôležitejších orgánov v priebehu evolúcie sú celkom bežné.
Charakteristický je proces gastrulácie dôležité bunkové premeny, ako sú riadené pohyby skupín a jednotlivých buniek, selektívne rozmnožovanie a triedenie buniek, začiatok cytodiferenciácie a indukčné interakcie. Uvedené bunkové mechanizmy ontogenézy sú podrobne diskutované v časti 8.2.
Metódy gastrulácie sú rôzne. Rozlišujú sa štyri typy priestorovo usmernených pohybov buniek, ktoré vedú k premene embrya z jednej vrstvy na viacvrstvovú (dve alebo tri vrstvy).
Intususcepcia- invaginácia jedného z úsekov blastodermu dovnútra ako celej vrstvy. V lancelete invaginujú bunky vegetatívneho pólu, u obojživelníkov dochádza k intususcepcii na hranici medzi zvieracím a vegetatívnym pólom v oblasti sivého polmesiaca. Proces invaginácie je možný iba vo vajciach s malým alebo stredným množstvom žĺtka.
epiboly- zanášanie malými rýchlo sa deliacimi bunkami živočíšneho pólu väčších, v rýchlosti delenia zaostávajúcich a menej pohyblivých buniek vegetatívneho pólu. Tento proces je jasne vyjadrený u obojživelníkov.
Delaminácia- rozvrstvenie buniek blastodermy do dvoch vrstiev ležiacich nad sebou. Delamináciu možno pozorovať v diskoblastule embryí s neúplným rozdrvením zvierat, ako sú plazy, vtáky a vajcorodé cicavce. Delaminácia sa prejavuje v embryoblastoch placentárnych cicavcov, čo vedie k vytvoreniu hypoblastu a epiblastu.
imigrácia- pohyb skupín alebo jednotlivých buniek, ktoré nie sú spojené do jednej vrstvy. Imigrácia sa vyskytuje vo všetkých embryách, ale je najcharakteristickejšia pre druhú fázu gastrulácie u vyšších stavovcov (plazy, vtáky a cicavce).
V každom konkrétnom prípade embryogenézy sa spravidla kombinuje niekoľko metód gastrulácie.
7.4.2.2. Morfológia gastrulácie
Podrobnejšie vyšetrenie gastrulácie u lanceletu, žaby, sliepky a cicavcov pomôže lepšie pochopiť evolučné vzťahy a pochopiť zákonitosti individuálneho vývoja.
7.4.2.2-a. Gaetrulácia lanceletu
gastrulácia lancelet znázornené na obr. 7.5. Rôzne markery v štádiu blastuly (obr. 7.5, a) označené predpokladaný(údajne) základy. Sú to oblasti blastuly, z bunkového materiálu
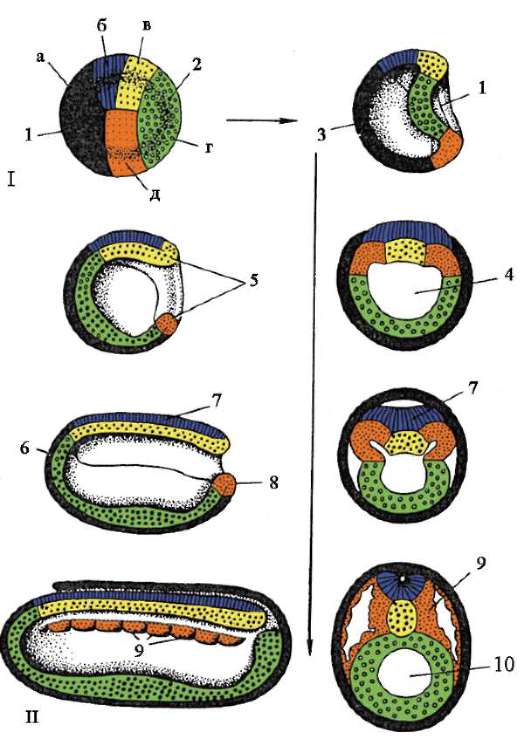
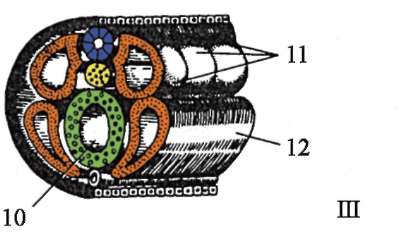
Ryža. 7.5.Predpokladané základy, gastrulácia a neurulácia v lancelete. I - predpokladané rudimenty v štádiu blastuly (vonkajší pohľad) a ranej gastruly (rezový pohľad); II - neskorá gastrula a neurulácia na sagitálnych (ľavý rad) a priečnych (pravý rad) rezoch; III - plastický model embrya na konci neurulačného obdobia:
1 - zvieracia tyč; 2 - vegetatívny pól; 3 - blastocoel; 4 - gastro-gól; 5 - dorzálne a ventrálne pysky blastopóru; 6 - hlavový koniec embrya; 7 - medulárna doska; 8 - chvostový koniec embrya; 9 - dorzálna časť mezodermu; 10 - dutina sekundárneho čreva; 11 - segmentované somity; 12 - brušná časť mezodermu; a, b, c, d, e - označenia predpokladaných a vyvíjajúcich sa orgánov: a - kožný ektoderm; b - nervová trubica; v - akord; d - endoderm, črevný epitel; d - mezoderm
z ktorých sa pri gastrulácii a včasnej organogenéze (neurulácii) zvyčajne vytvoria úplne definované zárodočné vrstvy a orgány (obr. 7.5, b a c).
Invaginácia začína na vegetatívnom póle. Vďaka rýchlejšiemu deleniu bunky živočíšneho pólu rastú a vytláčajú bunky vegetatívneho pólu do blastuly. To je uľahčené zmenou stavu cytoplazmy v bunkách, ktoré tvoria pery blastopóru a priľahlé k nim. V dôsledku invaginácie klesá blastocoel a zvyšuje sa gastrocoel. Súčasne s vymiznutím blastocoelu sa ektoderm a endoderm dostanú do úzkeho kontaktu. Lancelet, ako každý iný deuterostómy(patrí sem kmeň Echinodermata, kmeň Chordata a niektoré ďalšie relatívne malé druhy živočíchov), oblasť blastopóru sa mení na chvostovú časť organizmu, na rozdiel od protostómov, v ktorých blastopor zodpovedá hlavovej časti. Otváranie úst v deuterostómoch vytvorený na konci embrya oproti blastopóru.
7.4.2.2-6. Gaetrulácia u obojživelníkov
Gastrulácia u obojživelníkov má veľa spoločného s gastruláciou lanceletu, ale keďže žĺtok v ich vajciach je oveľa väčší a nachádza sa hlavne na vegetatívnom póle, veľké blastoméry amfiblastuly sa nedokážu vydúvať dovnútra. Intususcepcia ide trochu inak. Na hranici medzi zvieracím a vegetatívnym pólom v oblasti sivého kosáka sa bunky najprv silne natiahnu dovnútra, pričom majú tvar „baňového tvaru“ (obr. 7.6, a) a potom vytiahnu bunky povrchovej vrstvy blastuly spolu s nimi. Objavuje sa polmesiačiková ryha a dorzálny (dorzálny) pysk blastopóru (obr. 7.6, b).
Súčasne sa menšie bunky zvieracieho pólu, ktoré sa delia rýchlejšie, začnú pohybovať smerom k vegetatívnemu pólu. V oblasti chrbtovej pery sa vytáčajú a invaginujú a väčšie bunky rastú po stranách a na opačnej strane kosákovitej ryhy. Potom proces epiboly vedie k vytvoreniu laterálnych (laterálnych) a ventrálnych (ventrálnych) pyskov blastopóru. Blatopór sa uzatvára do prstenca, vo vnútri ktorého sú nejaký čas viditeľné veľké svetelné bunky vegetatívneho pólu v podobe tzv. žĺtka. Neskôr sú úplne ponorené dovnútra a blastopór sa zužuje.
Pomocou metódy značenia vitálnymi (vitálnymi) farbivami u obojživelníkov boli podrobne študované pohyby buniek
Ryža. 7.6.Tvorba blastopóru včasnej gastruly obojživelníkov: a - baňovité bunky v oblasti dorzálneho pysku blastopóru; b - všeobecná forma ryha polmesiaca a chrbtový pysk; 1 - bunky v tvare banky; 2 - dorzálny pysk blastopóru
blastula pri gastrulácii sa zistilo, že špecifické oblasti blastodermu, tzv predpokladaný(z lat. praesumptio- predpoklad), pri normálnom vývine sa objavujú najskôr v zložení určitých rudimentov orgánov a potom v zložení orgánov samotných (obr. 7.7). Je známe, že u bezchvostých obojživelníkov materiál predpokladaného notochordu a mezodermu v štádiu blastuly neleží na jeho povrchu, ale vo vnútorných vrstvách steny amfiblastuly, avšak približne v rovnakých úrovniach, ako je znázornené na obrázku.
Určitá podobnosť medzi procesmi gastrulácie a oblasťami predpokladaných orgánov u obojživelníkov a lancelet, t.j. homológia hlavných orgánov, ako je nervová trubica, notochorda, sekundárne črevo, naznačuje ich fylogenetický vzťah.
7.4.2.2-c. Gastrulácia u vtákov
Gastrulácia v embryách s meroblastickým typom štiepenia a vývoja má svoje vlastné charakteristiky. o vtákov začína po rozštiepení a vytvorení blastuly pri prechode embrya cez vajcovod. V čase, keď je vajíčko znesené, embryo už pozostáva z niekoľkých vrstiev: horná vrstva sa nazýva epiblast, nižšie - primárny hypoblastóm(pozri obr. 7.4, c). Medzi nimi je úzka medzera - blastocoel. Potom sa vytvoril sekundárny hypoblast, spôsob vzniku nie je celkom jasný. Existujú dôkazy, že v primár
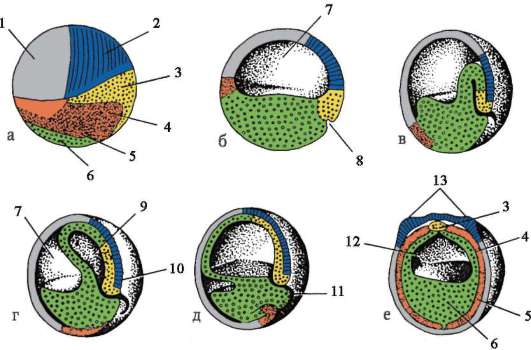
Ryža. 7.7. Mapa oblastí predpokladaných orgánových rudimentov v skorých štádiách embryonálneho vývoja obojživelníkov: a - štádium blastuly (pohľad vľavo); b-e - postupné štádiá gastrulácie (sagitálne rezy); e - začiatok neurulácie (prierez); 1 - kožný ektoderm; 2 - nervová trubica; 3 - akord; 4 - somitový mezoderm; 5 - mezoderm splanchnotómov; 6 - endoderm; 7 - blastocoel; 8 - polmesiaca drážka; 9 - gastrocoel; 10 - blastopór dorzálnej pery; 11 - žĺtka; 12 - dutina sekundárneho čreva; 13 - nervové valčeky
V oblasti vtákov vznikajú primárne zárodočné bunky a sekundárne tvoria extraembryonálny endoderm. Vznik primárneho a sekundárneho hypoblastu sa považuje za jav predchádzajúci gastrulácii.
Hlavné udalosti gastrulácie a konečnej tvorby troch zárodočných vrstiev sa začínajú po kladení vajíčok so začiatkom inkubácie. Dochádza k hromadeniu buniek v zadnej časti epiblastu v dôsledku nerovnomernej rýchlosti bunkového delenia a ich pohybu z laterálnych častí epiblastu do stredu, smerom k sebe. Takzvaný primárna línia, ktorý sa tiahne smerom k hlavovému koncu. V strede primárneho pruhu sa tvorí primárna brázda, a pozdĺž okrajov primárne rolky. Na hlavovom konci primárneho prúžku sa objaví zhrubnutie - Hensenov uzol, a v ňom - primárna jama(obr. 7.8).
Keď bunky epiblastu vstúpia do primárnej drážky, ich tvar sa zmení. Tvarom sa podobajú na bunky „baňovitého tvaru“ gastruly obojživelníkov. Tieto bunky sa potom stanú hviezdicovitými a ponoria sa pod epiblast, aby vytvorili mezoderm. Z časti migrujúcich buniek, uložených v hypoblaste, neskôr vzniká germinálny endoderm (obr. 7.9). Endoderm je teda tvorený na báze primárneho a sekundárneho hypoblastu s pridaním novej generácie endodermálnych buniek migrujúcich z horných vrstiev blastodermu. Prítomnosť niekoľkých generácií endodermálnych buniek naznačuje predĺženie doby gastrulácie v čase.
Časť buniek migrujúcich z epiblastu cez Hensenov uzol tvorí budúci notochord. Súčasne s anlážou a predĺžením akordu sa Hensenov uzol a primárny pruh postupne posúvajú v smere od predného ku kaudálnemu koncu a miznú. Tomu zodpovedá zúženie a uzavretie blastopóru. Keď sa primárny pruh zmršťuje, zanecháva za sebou vytvorené časti osových orgánov embrya v smere od hlavy k častiam chvosta. Zdá sa rozumné považovať pohyby buniek v kuracom embryu za homológne s pohybmi u obojživelníkov a primárny pruh a Hensenov uzol za homológne s blastopórom a dorzálnym (dorzálnym) pyskom gastruly týchto zvierat.
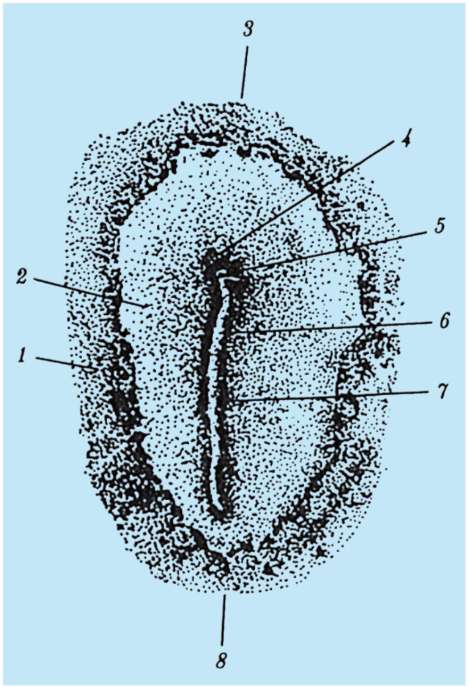
Ryža. 7.8.Kuracie embryo v štádiu primárneho pruhu (dorzálny pohľad): 1 - tmavá oblasť; 2 - priesvitná oblasť zárodočného disku; 3 - hlavový koniec; 4 - Hensenov uzol; 5 - primárna fossa; 6 - primárna drážka; 7 - primárny valec; 8 - koniec chvosta
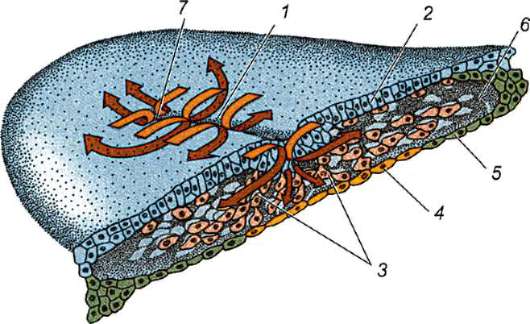
Ryža. 7.9.Kuracie embryo v štádiu primárneho pásika (prierez): 1 - primárna drážka; 2 - epiblast (ektoderm); 3 - migrujúce bunky (mezoderm); 4 - germinálny endoderm; 5 - hypoblast (endoderm); 6 - blastocoel; 7 - Hensenov uzol
7.4.2.2-d. Gastrulácia u cicavcov
Je zaujímavé poznamenať, že bunky embryí cicavcov (pozri odsek 7.5.1), napriek skutočnosti, že u týchto zvierat majú vajíčka malé množstvo žĺtka a fragmentácia je úplná, vo fáze gastrulácie si zachovávajú pohyby charakteristické pre embryá plazov a vtákov. To potvrdzuje myšlienku pôvodu cicavcov zo skupiny predkov, ktorých vajcia boli bohaté na žĺtok.
7.4.2.3. Vlastnosti štádia gastrulácie
Gastrulácia je charakterizovaná rôznymi bunkovými procesmi. Mitotic pokračuje reprodukcia buniek, navyše má v rôznych častiach embrya rôznu intenzitu. Medzifázy mitotických cyklov zahŕňajú všetky obdobia (G 1, S, G 2), preto sa od štádia gastrulácie pozoruje rast vyvíjajúceho sa organizmu. Avšak najviac charakteristický gastrulácia pozostáva z pohyb bunkových hmôt. To vedie k zmene štruktúry embrya a jeho premene z blastuly na gastrulu. deje triedenie bunky svojou príslušnosťou k rôznym zárodočným vrstvám, vo vnútri ktorých sa navzájom „spoznávajú“.
Začína sa fáza gastrulácie diferenciácia buniek, čo znamená prechod k aktívnemu využívaniu biologickej informácie vlastného genómu. Jedným z regulátorov genetickej aktivity je rozdielne chemické zloženie cytoplazmy embryonálnych buniek, ktoré vzniká v dôsledku ovo(oo)plazmatickej segregácie, pričom obe pochádzajú z vegetatívneho pólu vajíčka.
Počas gastrulácie je úloha veľmi veľká embryonálna indukcia- interakcie medzi bunkovými komplexmi (časťami) vyvíjajúceho sa embrya. Ukázalo sa, že výskyt primárneho pruhu u vtákov je výsledkom indukčnej interakcie medzi hypoblastom a epiblastom. Hypoblast má polaritu. Zmena polohy hypoblastu voči epiblastu spôsobuje zmenu orientácie primitívneho pruhu. Všetky tieto procesy sú podrobne popísané v kapitole 8.
7.4.3. TVORBA ORGÁNOV A TKANIV 7.4.3.1. Podstata štádia organogenézy
organogenéza, spočívajúce vo vytváraní jednotlivých orgánov, tvoria hlavnú náplň embryonálneho obdobia. Pokračujú v larve a končia v juvenilnom období. Organogenéza sa vyznačuje najzložitejšími a najrozmanitejšími morfogenetickými (tvarovacími) premenami. Nevyhnutným predpokladom prechodu k organogenéze je dosiahnutie štádia gastruly zárodkom, a to vytvorenie zárodočných vrstiev. Zárodočné vrstvy, ktoré vo vzťahu k sebe zaujímajú určitú pozíciu, poskytujú kontaktom a interakciou také vzťahy medzi rôznymi bunkovými skupinami, ktoré stimulujú ich vývoj v určitom smere. Tento tzv embryonálna indukcia- najdôležitejší dôsledok vzájomného pôsobenia medzi zárodočnými vrstvami.
V priebehu organogenézy sa mení tvar, štruktúra a chemické zloženie buniek, izolujú sa bunkové skupiny, ktoré sú základom budúcich orgánov. Postupne sa vyvíja určitá forma orgánov, vytvárajú sa medzi nimi priestorové a funkčné spojenia. Procesy morfogenézy sú sprevádzané diferenciáciou
diferenciácia buniek a tvorba tkanív, diferenciácia tkanív a štruktúr, ako aj ich selektívny a nerovnomerný rast. Predpokladom organogenézy spolu s reprodukciou buniek, migráciou a triedením je ich selektívna smrť (pozri kapitolu 8).
7.4.3.2. Neuralizácia
Samotný začiatok organogenézy sa nazýva neurulácia. Neuralizácia pokrýva procesy od objavenia sa prvých príznakov tvorby nervovej platničky až po jej uzavretie v nervovej trubici (obr. 7.10). V rovnakom čase, akord A sekundárne črevo, a mezoderm ležiaci po stranách chordy sa rozdeľuje v kraniokaudálnom smere na segmentované párové štruktúry - somites(vzniká komplex osových orgánov - primárna organogenéza).
Nervový systém stavovcov, vrátane ľudí, sa vyznačuje stabilitou hlavného štrukturálneho plánu v celom rozsahu evolučná história podtyp. Pri tvorbe neurálnej trubice majú všetky strunatce veľa spoločného. Spočiatku nešpecializované dorzálny ektoderm, reagujúc na indukčný efekt z chordomesodermy sa mení na nervová platnička, predložené neuroepiteliálne cylindrické bunky.
Nervová platnička nezostáva dlho sploštená. Čoskoro sa jeho bočné okraje zdvíhajú a vytvárajú sa nervové záhyby, ktoré ležia po oboch stranách plytkého pozdĺžnika nervová drážka. Okraje nervových záhybov sa potom uzavrú a vytvoria uzavretú nervovú trubicu s kanálom vo vnútri - neurokéla. Po prvé, k uzavretiu nervových záhybov dochádza na úrovni začiatku (hranica medzi miechou a mozgom) miechy a potom sa šíri do mozgu a chvosta.
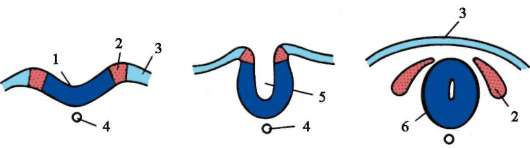
Ryža. 7.10. Postupné štádiá tvorby neurálnej trubice a neurálnej lišty (prierez embrya): 1 - neurálna platnička; 2 - nervový hrebeň; 3 - ektoderm; 4 - akord; 5 - nervová drážka; 6 - neurocoel; 7 - nervové záhyby
inštrukcie. Ukázalo sa, že mikrotubuly a mikrofilamenty neuroepiteliálnych buniek hrajú dôležitú úlohu v morfogenéze nervovej trubice. Zničenie týchto bunkových štruktúr kolchicínom a cytochalazínom IN spôsobí, že nervová platnička zostane otvorená. Neuzavretie nervových záhybov vedie k vrodeným malformáciám nervovej trubice.
Po uzavretí nervových záhybov sa vytvoria bunky, ktoré sa pôvodne nachádzali medzi nervovou platničkou a budúcim kožným ektodermom. neurálny hrebeň. Bunky neurálnej lišty sa vyznačujú schopnosťou rozsiahlej, ale prísne regulovanej migrácie v tele (pozri časť 8.2.2, obr. 8.9) a tvoria dva hlavné prúdy. Bunky jedného z nich - povrchný- sú zahrnuté v epidermis alebo dermis kože, kde sa diferencujú na pigmentové bunky. Ďalší prúd migruje ventrálnym smerom a tvorí citlivé spinálne gangliá, sympatické gangliá, dreň nadobličiek, parasympatické gangliá. Bunky z lebečnej neurálnej lišty vedú k vzniku nervových buniek a mnohých ďalších štruktúr, ako je žiabrová chrupavka, niektoré krycie kosti lebky.
Endoderm vo všetkých embryách nakoniec tvorí epitel sekundárneho čreva a mnohých jeho derivátov. Samotné sekundárne črevo sa vždy nachádza pod akordom.
V procese neurulácie teda komplex osové orgány: nervová trubica, chorda, črevo, ktoré sú najcharakteristickejším znakom organizácie tela všetkých strunatcov. Rovnaký pôvod, vývoj a vzájomné usporiadanie osových orgánov prezrádza ich úplnú homológiu a evolučnú kontinuitu.
Hĺbkové vyšetrenie a porovnanie neurulačných procesov u konkrétnych predstaviteľov strunatcovitého typu odhaľuje niektoré rozdiely, ktoré súvisia najmä so znakmi, ktoré závisia od štruktúry vajíčok, spôsobu drvenia a gastrulácie (obr. 7.11). Pozornosť upriamuje na odlišný tvar embryí a posun v čase kladenia osových orgánov voči sebe, t.j. heterochrónia opísaná vyššie.
7.4.3.3. mezodermálna diferenciácia
Mezoderm, ktorý zaberá miesto po stranách notochordu a siaha ďalej medzi kožným ektodermom a endodermou sekundárneho čreva, sa delí na dorzálnu a ventrálnu oblasť. dorzálny
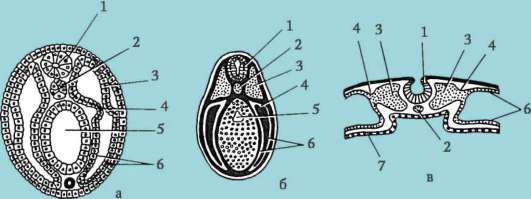
Ryža. 7.11. Neuruly rôznych strunatcov: a - lancelet; b - žaba; c - kuracie mäso; 1 - nervová trubica; 2 - akord; 3 - somit; 4 - noha somit; 5 - sekundárne črevo; 6 - bočná doska; 7 - endoderm
časť je segmentovaná a prezentovaná v pároch somites. Kladenie somitov prebieha od hlavy po chvost. Ventrálna nesegmentovaná časť mezodermu, ktorá vyzerá ako tenká vrstva buniek, je tzv. bočná doska. Somity sú spojené s laterálnou platňou intermediárnym mezodermom vo forme segmentov somite nohy.
Všetky oblasti mezodermu sa postupne diferencujú. Na začiatku tvorby majú somity konfiguráciu charakteristickú pre epitel s dutinou vo vnútri. Pod indukčným efektom vychádzajúcim z chordy a neurálnej trubice ventromediálne časti somitov - sklerotómy- premieňajú sa na sekundárny mezenchým, sú vypudzované zo somitu a obklopujú notochord a ventrálnu časť nervovej trubice. Nakoniec sa z nich vytvoria stavce, rebrá a lopatky.
Dorzolaterálna časť somitov sa diferencuje v dvoch smeroch. Formujú sa oblasti somitov smerujúce k krycej ektoderme dermatómy, z ktorých vzniká vnútorná vrstva kože – dermis. Vznikajú bunky umiestnené na vnútornej strane dorzolaterálnej časti somitov myotómy, z ktorého sa vyvinie priečne pruhované kostrové svalstvo tela a končatín. Z oblasti nôh somitov s rudimentami nefrotóm A gonotóm tvoria sa vylučovacie orgány a pohlavné žľazy.
Nesegmentované bočné dosky rozdelené na dva listy, ktoré obmedzujú sekundárnu telesnú dutinu - všeobecne. vnútorná plachta, priľahlý k endodermu sa nazýva viscerálny. Obklopuje črevo zo všetkých strán a tvorí mezentérium, pokrýva pľúcne
parenchýmu a srdcového svalu. Vonkajší list laterálna platnička susedí s ektodermou a je tzv parietálny. V budúcnosti tvorí vonkajšie listy pobrušnice, pohrudnice a osrdcovníka.
7.4.3.4. Deriváty zárodočných vrstiev
Ektoderm, mezoderm a endoderm sa v priebehu ďalšieho vývoja, ktoré sa navzájom ovplyvňujú, podieľajú na tvorbe špecifických orgánov (súkromná alebo lokálna organogenéza). Vznik rudimentu orgánu je spojený s miestnymi zmenami v určitej oblasti zodpovedajúcej zárodočnej vrstvy. takže, z ektodermy vzniká epidermis kože a jej derivátov (perie, vlasy, nechty, koža a mliečne žľazy), zložky orgánov zraku, sluchu, čuchu, epitel ústnej dutiny a zubná sklovina. Najdôležitejšími ektodermálnymi derivátmi sú nervová trubica, nervový hrebeň a všetky nervové štruktúry z nich vytvorené.
Endodermové deriváty sú epitel žalúdka a čriev, pečeňové bunky, sekrečné bunky pankreasu, črevné a žalúdočné žľazy. Endoderm prednej časti embryonálneho čreva tvorí epitel pľúc a dýchacích ciest, ako aj secernujúce bunky predného a stredného laloka hypofýzy, štítnej žľazy a prištítnych teliesok.
mezoderm, okrem už popísaných kostrových štruktúr kostrové svaly, dermis kože, orgány vylučovacej a reprodukčnej sústavy tvorí kardiovaskulárny systém, lymfatický systém, pohrudnicu, pobrušnicu a osrdcovník. Od mezenchým, ktoré majú zmiešaný pôvod v dôsledku buniek troch zárodočných vrstiev, vyvíjajú sa všetky typy spojivového tkaniva, hladkého svalstva, krvi a lymfy (podrobnejšie pozri odsek 8.2.5).
Rudiment konkrétneho orgánu sa spočiatku tvorí zo špecifickej zárodočnej vrstvy, ale potom sa orgán stáva zložitejším a v dôsledku toho sa na jeho tvorbe podieľajú dve alebo tri zárodočné vrstvy.
7.4.4. PROVIZORNÉ ORGÁNY EMBRYA STAVOVCOV
provizórne, alebo dočasné, orgány sa tvoria v embryogenéze radu zástupcov stavovcov na zabezpečenie životných funkcií embrya, ako je dýchanie, výživa, vylučovanie, pohyb a iné. Nedostatočne vyvinuté orgány zvierat, ktoré sa najviac vyvíjajú, ešte nie sú schopné fungovať podľa predstáv, hoci musia
hrajú určitú úlohu v systéme vyvíjajúceho sa celého organizmu. Len čo embryo dosiahne potrebný stupeň zrelosti, keď je väčšina orgánov schopná vykonávať životne dôležité funkcie, dočasné orgány sa resorbujú alebo zlikvidujú.
Čas vytvorenia provizórnych orgánov závisí od toho, aké zásoby živín sa nahromadili vo vajíčku a v akých podmienkach prostredia sa embryo vyvíja. Napríklad u bezchvostých obojživelníkov v dôsledku dostatočného množstva žĺtka vo vajci a skutočnosti, že vývoj prebieha vo vode, embryo uskutočňuje výmenu plynov a uvoľňuje produkty disimilácie priamo cez škrupiny vajíčka a dosahuje štádium pulca. V tomto štádiu sa vytvárajú provizórne orgány dýchania (žiabre), trávenia a pohybu prispôsobené vodnému životnému štýlu. Uvedené larválne orgány umožňujú pulcovi pokračovať vo vývoji. Po dosiahnutí stavu morfologickej a funkčnej zrelosti orgánov dospelého typu miznú dočasné orgány v procese metamorfózy.
Plazy a vtáky majú vo vajci viac zásob žĺtka, ale embryo sa nevyvíja vo vode, ale na súši. V tomto ohľade je veľmi skoro potrebné zabezpečiť dýchanie a vylučovanie, ako aj ochranu pred vysychaním. U nich sa už v ranej embryogenéze, takmer súbežne s neuruláciou, začína tvorba provizórnych orgánov, ako napr. amnion, chorion A žĺtkový vak. O niečo neskôr sa vytvorí alantois. U placentárnych cicavcov sa tieto isté dočasné orgány vytvárajú ešte skôr, pretože vo vajci je veľmi málo žĺtka. Vývoj takýchto zvierat sa vyskytuje in utero, tvorba provizórnych orgánov v nich sa časovo zhoduje s obdobím gastrulácie.
Prítomnosť alebo neprítomnosť amniónu a iných dočasných orgánov je základom rozdelenia stavovcov do dvoch skupín: Amniota A Anamnia(pozri tabuľku 7.1). Evolučne staršie stavovce, ktoré sa vyvíjajú výlučne vo vodnom prostredí a sú zastúpené takými triedami, ako sú cyklostómy, ryby a obojživelníky, nepotrebujú ďalšiu vodu a iné schránky embrya a tvoria skupinu anamnium. Do skupiny amniotov patria primárne suchozemské stavovce, t.j. tie, ktorých embryonálny vývoj prebieha v pozemských podmienkach. Toto sú tri triedy: plazy, vtáky a cicavce. Patria medzi vyššie stavovce, keďže majú koordinované a vysoko výkonné orgánové systémy, ktoré zabezpečujú ich existenciu v naj. ťažké podmienky aké sú pôdne podmienky. Títo
triedy zahŕňajú veľké množstvo druhov, ktoré sekundárne prešli do vodného prostredia. Vyššie stavovce si teda dokázali osvojiť všetky biotopy. Takáto dokonalosť by bola nemožná, a to aj bez vnútorného oplodnenia a vytvorenia špeciálnych provizórnych embryonálnych orgánov, tzv embryonálne membrány. Amniotické membrány zahŕňajú amnion, chorion (serosa), žĺtkový vak a alantois.
V štruktúre a funkciách provizórnych orgánov rôznych amniotov je veľa spoločného. Pri najvšeobecnejšom opise provizórnych orgánov embryí vyšších stavovcov treba poznamenať, že všetky sa vyvíjajú z bunkového materiálu už vytvorených zárodočných vrstiev. Vo vývoji embryonálnych membrán placentárnych cicavcov existujú určité znaky, o ktorých sa bude diskutovať nižšie.
Amnion je vak obsahujúci embryo a naplnený plodovou vodou. Amniotická membrána je tvorená extraembryonálnym ektodermom a somatopleurou. Jeho ektodermálna časť je špecializovaná na sekréciu a vstrebávanie plodovej vody, ktorá obmýva embryo. Amnion zohráva primárnu úlohu pri ochrane embrya pred vysychaním a mechanickým poškodením a vytvára preň najpriaznivejšie a najprirodzenejšie vodné prostredie. Mezodermálna časť amniónu dáva vznik hladkým svalovým vláknam. Kontrakcie týchto svalov spôsobujú pulzovanie amniónu a pomalé oscilačné pohyby, ktoré sú pri tomto procese embryu oznámené, zjavne pomáhajú zabezpečiť, aby sa jeho rastúce časti navzájom nerušili.
Chorion (serosa)- vonkajšia zárodočná membrána susediaca so schránkou (serosa) alebo materskými tkanivami (chorion), vychádzajúca podobne ako amnion z ektodermy a somatopleury. Táto škrupina slúži na výmenu medzi embryom a prostredím. U vajcorodých druhov je hlavnou funkciou serózy účasť na dýchaní (výmena plynov); u cicavcov plní chorion oveľa rozsiahlejšie funkcie, podieľa sa okrem dýchania aj na výžive, vylučovaní, filtrácii a syntéze látok, ako sú hormóny.
Žĺtkový vak vytvorený z extraembryonálneho endodermu a viscerálneho mezodermu a je priamo spojený s črevnou trubicou embrya. U embryí s veľkým množstvom žĺtka sa podieľa na výžive. U vtákov sa napríklad vyvinie vaskulatúra v splanchnopleuróne žĺtkového vaku. Žĺtok neprechádza cez žĺtok, ktorý spája vačok s črevom. Najprv sa preloží do
rozpustná forma pôsobením tráviacich enzýmov produkovaných endodermálnymi bunkami steny vaku. Potom vstúpi do ciev a šíri sa krvou po celom tele embrya.
Cicavce nemajú zásoby žĺtka a zachovanie žĺtkového vaku môže byť spojené s dôležitými sekundárnymi funkciami. Mezoderm žĺtkového vaku produkuje krvinky embrya. Okrem toho je žĺtkový vak cicavcov naplnený tekutinou s vysokou koncentráciou aminokyselín a glukózy, čo naznačuje možnosť metabolizmu bielkovín v žĺtkovom vaku.
Osud žĺtkového vaku u rôznych zvierat je trochu odlišný. U vtákov sú na konci inkubačnej doby zvyšky žĺtkového vaku už vo vnútri embrya, potom rýchlo zmizne a úplne ustúpi do konca 6 dní po vyliahnutí. U cicavcov je žĺtkový vak vyvinutý rôznymi spôsobmi. U dravcov je pomerne veľký, s vysoko vyvinutou sieťou ciev, zatiaľ čo u primátov sa rýchlo zmenšuje a pred pôrodom mizne bez stopy.
Allantois sa vyvíja o niečo neskôr ako ostatné dočasné orgány. Je to vakovitý výrastok ventrálnej steny zadného čreva. Preto ho tvorí endoderm zvnútra a splanchnopleura zvonka. U plazov a vtákov alantois rýchlo rastie na chorion a vykonáva niekoľko funkcií. V prvom rade je to rezervoár pre močovinu a kyselinu močovú, ktoré sú konečnými produktmi metabolizmu obsahujúceho dusík. organickej hmoty. Alantois má dobre vyvinutú cievnu sieť, vďaka čomu sa spolu s chorionom podieľa na výmene plynov. Pri vyliahnutí sa vonkajšia časť alantois zahodí, zatiaľ čo vnútorná časť sa zachová ako mechúr.
U mnohých cicavcov je alantois tiež dobre vyvinutý a spolu s chorionom tvorí chorioalantoickú placentu. Termín placenta znamená úzke prekrytie alebo splynutie zárodočných membrán s tkanivami rodičovského organizmu. U primátov a niektorých iných cicavcov je endodermálna časť alantois rudimentárna a mezodermálne bunky tvoria hustú šnúru siahajúcu od kloakálnej oblasti po chorion. Cievy rastú pozdĺž mezodermu alantois až po chorion, cez ktorý placenta vykonáva vylučovacie, dýchacie a nutričné funkcie.
Je dostupnejšie a ľahšie študovať tvorbu a štruktúru embryonálnych membrán na príklade kuracieho embrya. V štádiu neuruly prechádzajú tri zárodočné vrstvy priamo z embrya do extra-
dýchacia časť, bez akéhokoľvek ohraničovania. Keď sa embryo formuje, vytvorí sa okolo neho niekoľko záhybov, ktoré embryo akoby podrežú, oddelia od žĺtka a vytvoria jasné hranice medzi embryom a extraembryonálnymi oblasťami. Volajú sa kmeňové záhyby(obr. 7.12).
Najprv sa vytvorí záhyb hlavy. Zospodu odreže hlavovú časť. Zadné konce tohto záhybu prechádzajú do bočných kmeňových záhybov, ktoré ohraničujú telo embrya zo strán. Chvostový záhyb ohraničuje zadný koniec embrya. Postupne sa zužuje noha spájajúca stredné črevo a žĺtkový vak, vytvorený
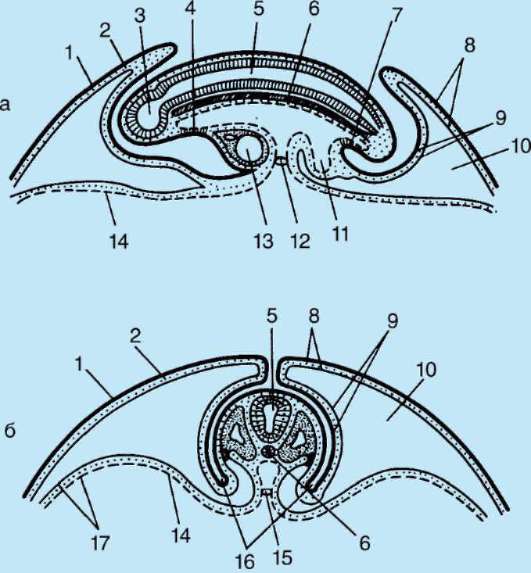
Ryža. 7.12. Tvorba kmeňových záhybov a embryonálnych membrán v kuracom embryu: a - pozdĺžny rez; b - prierez; 1 - ektoderm; 2 - mezoderm; 3 - základ mozgu; 4 - faryngálna membrána; 5 - nervová trubica; 6 - akord; 7 - kloakálna membrána; 8 - chorion; 9 - amnion; 10 - exocoel; 11 - alantois; 12 - oblasť pupka; 13 - základ srdca; 14 - endoderm; 15 - záložka čreva; 16 - záhyby trupu; 17 - žĺtkový vak
predné a zadné črevá. Zároveň sa z ektodermy a k nej priľahlej somatopleury najskôr vytvorí hlavový záhyb (obr. 7.13), ktorý ako kapucňa prirastá na embryo zhora. Na bokoch sa tvoria konce záhybu hlavy amniotické záhyby. Rastú na vrchu embrya smerom k sebe a rastú spolu, pričom bezprostredne tvoria steny amniónu, susediace s embryom, a chorion, ktorý leží vonku.
Neskôr vzniká alantois (obr. 7.14). Celkový pohľad na kuracie embryo v 6. deň inkubácie je znázornený na obr. 7.15. U rôznych cicavcov sú procesy tvorby provizórnych orgánov viac-menej podobné tým, ktoré sú opísané vyššie u vtákov. Charakteristiky ich vývoja u primátov vrátane človeka sú uvedené v nasledujúcej kapitole.
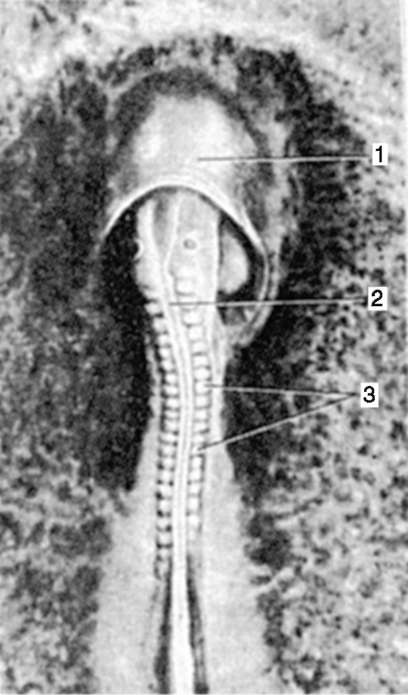
Ryža. 7.13.Kuracie embryo asi 40 hodín inkubácie: 1 - záhyb amniónovej hlavy; 2 - nervová trubica; 3 - somity
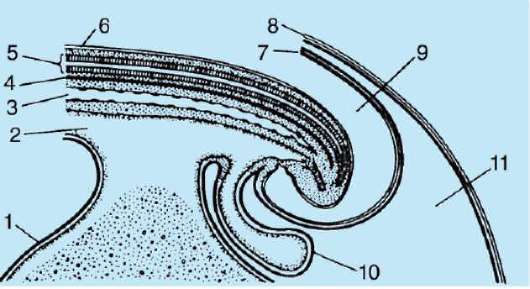
Ryža. 7.14.Tvorba alantoisu v kuracom embryu (pozdĺžny rez chvostom): 1 - žĺtkový vak; 2 - stredné črevo; 3 - aorta; 4 - akord; 3 - nervová trubica; 6 - ektoderm; 7 - amnion; 8 - chorion; 9 - amniová dutina; 10 - alantois; 11 - exocoelom
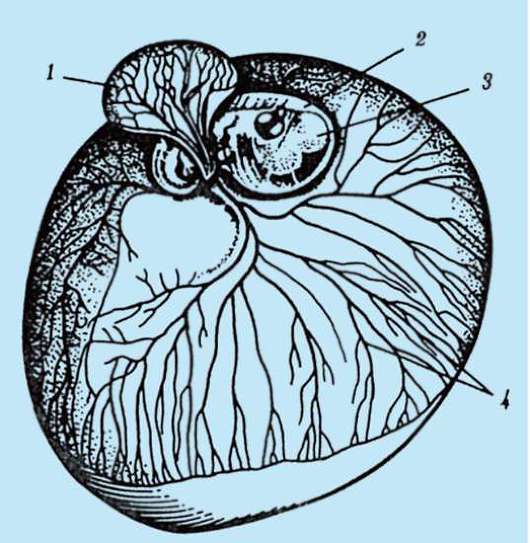
Ryža. 7.15. Kuracie embryo na 6. deň inkubácie (bielkoviny a chorion odstránené, allantois posunuté nahor): 1 - allantois; 2 - amnion; 3 - embryo; 4 - cievy žĺtkového vaku
7.5. embryonálny vývoj cicavcov
7.5.1. PERIODIZÁCIA A VČASNÝ VÝVOJ EMBRYA
Štúdium prenatálneho a najmä embryonálneho (vnútromaternicového) vývoja človeka je veľmi dôležité, pretože pomáha lepšie pochopiť vzťah medzi orgánmi, ako aj mechanizmy vzniku vrodených vývojových chýb. V embryonálnom vývoji rôznych druhov cicavcov existujú spoločné znaky, ale existujú aj rozdiely. Vo všetkých placentách sa napríklad procesy skorej embryogenézy výrazne líšia od procesov predtým opísaných u iných stavovcov. Zároveň medzi placentárnymi existujú špecifické znaky.
RozdelenieĽudská zygota sa vyznačuje nasledujúcimi znakmi. Rovina prvého delenia prechádza cez póly vajíčka, t.j. ako u iných stavovcov je to poludník. V tomto prípade je jedna z výsledných blastomér väčšia ako druhá, čo naznačuje nezrovnalosť drvenie. Do ďalšej divízie vstupujú prvé dve blastoméry asynchrónne. Brázda prebieha pozdĺž poludníka a je kolmá na prvú brázdu. Existuje teda štádium troch blastomér. Pri delení menšej blastoméry sa pár menších blastomér otočí o 90°, takže rovina štiepnej brázdy je kolmá na prvé dve
Ryža. 7.16.Skoré štádiá štiepenia králičej zygoty: I - rovina prvej štiepnej brázdy; IIa - rovina druhej brázdy drviaca jednu z prvých dvoch blastomér; IIb - rovina druhej štiepnej brázdy druhej z prvých dvoch blastomér
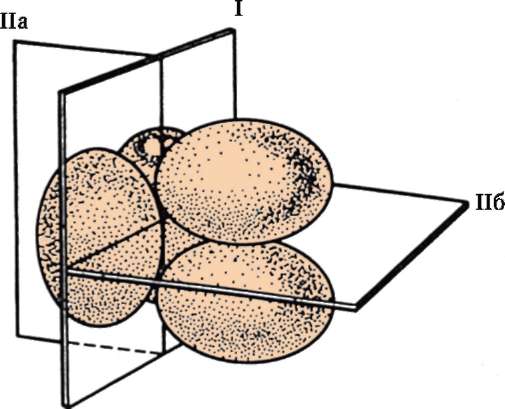
brázdy. Podobné usporiadanie blastomérov v 4-bunkovom štádiu bolo opísané u myší, králikov, norkov a opíc (obr. 7.16). V dôsledku asynchrónneho štiepenia môžu existovať štádiá s nepárnym počtom blastomérov - 3, 5, 7, 9, ktoré sa v dôsledku absencie rastového štádia líšia svojou veľkosťou. Takže drvenie človeka plne asynchrónne nejednotné.
V dôsledku drvenia sa vytvára akumulácia buniek - morula. Povrchovo umiestnené blastoméry tvoria bunkovú vrstvu a blastoméry ležiace vo vnútri moruly sú zoskupené do centrálneho bunkového uzla. Čoskoro sa vo vnútri moruly objaví tekutina, vytvorí sa dutina (blastocoel) a embryo sa zmení na blastocysta. Tvorba moruly nastáva v štádiu 16 blastomérov a kavitácia (tvorba dutiny) od štádia 32 blastomér.
IN blastocysta rozlíšiť vonkajšiu vrstvu buniek (trofoblast) a vnútorná bunková hmota (zárodočný uzlík, príp embryoblast). Vnútorná bunková hmota je tekutinou vytlačená nabok k jednému z pólov blastocysty. Neskôr od trofoblast vyvinie sa vonkajšia zárodočná membrána – chorion, a od embryoblast- samotné embryo a niektoré mimoembryonálne orgány. Ukázalo sa, že skutočné embryo sa vyvíja z veľmi malého počtu buniek zárodočného uzlíka.
Fáza drvenia prebieha pod lesklou škrupinou. Obrázok 7.17 ukazuje rané štádiá ľudskej embryogenézy, ukazuje, kde sa embryo nachádza v tele matky. Fragmentácia ľudskej zygoty a vznik blastocysty sú schematicky znázornené na obr. 7.18 a 7.19.
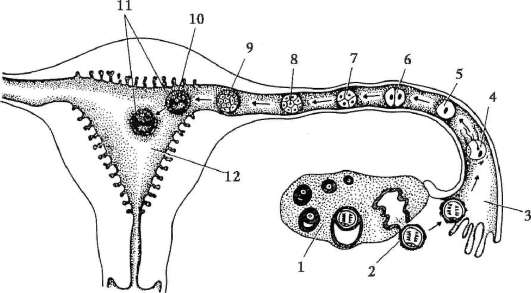
Ryža. 7.17.Ovulácia, oplodnenie a ľudské embryo v 1. týždni vývoja: 1 - vaječník; 2 - oocyt II poradia (ovulácia); 3 - vajcovod; 4 - hnojenie; 5 - zygota; 6 - embryo v štádiu dvoch blastomérov; 7 - embryo v štádiu štyroch blastomérov; 8 - embryo v štádiu ôsmich blastomérov; 9 - morula; 10, 11 - blastocysta; 12 - zadná stena maternice
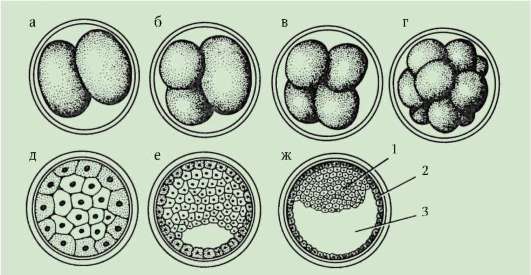
Ryža. 7.18.Štiepenie ľudskej zygoty: a - dve blastoméry; b - tri blastoméry; c - štyri blastoméry; g - morula; e - úsek moruly; f, g - úsek skorej a neskorej blastocysty: 1 - embryoblast; 2 - trofoblast; 3 - blastocoel
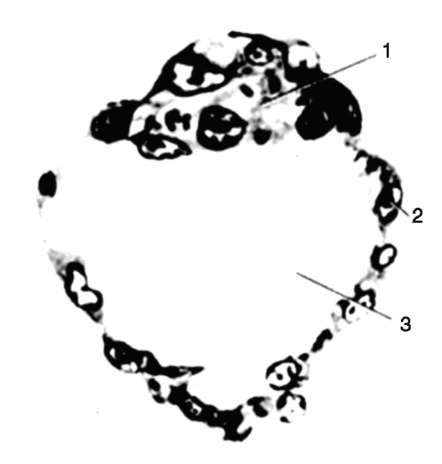
Ryža. 7.19.Ľudská embryoblastocysta (rez): 1 - embryoblast; 2 - trofoblast; 3 - blastocoel
Približne na 6.-7.deň po oplodnení je embryo, ktoré 2-3 dni voľne plávalo v dutine maternice, pripravené na implantáciu, t.j. k ponoreniu do jeho sliznice. Pôsobením enzýmov vylučovaných trofoblastom je zona pellucida čiastočne zničená a embryo sa z nej vykľuje ("vyliahne"). Za účasti integrínových proteínov syntetizovaných tak epitelom maternice, ako aj bunkami trofoblastu je blastula pripojená k stene maternice svojim embryonálnym pólom. Po kontakte s materskými tkanivami sa bunky trofoblastu rýchlo množia a uvoľnením proteolytických enzýmov ich (tieto tkanivá) zničia. To spôsobuje reakciu steny maternice sprevádzanú aktívnou tvorbou ciev v mieste implantácie. Porušenie syntézy integrínov a enzýmov potrebných na implantáciu tento proces znemožňuje, čo následne vedie k hypoxii a smrti embrya. Pri implantácii sa trofoblast diferencuje na dve vrstvy: vnútorná vrstva, tzv cytotrofoblastóm, pretože šetrí bunkovej štruktúry, a ten vonkajší, tzv syncytio-trofoblastóm, pretože je to syncytium. Obrázok 7.20 ukazuje ľudské embryo v procese implantácie.
gastrulácia u cicavcov úzko súvisí s inými embryonálnymi premenami. Súčasne s rozdelením trofoblastu na dve vrstvy sa zárodočný uzlík splošťuje a mení sa na dvojvrstvový zárodočný štít. Spodná vrstva štítu - hypoblast, alebo primárny endoderm, podľa väčšiny autorov, vzniká delamináciou vnútornej bunkovej hmoty, približne ako sa vyskytuje v embryonálnom disku vtákov. Primárny
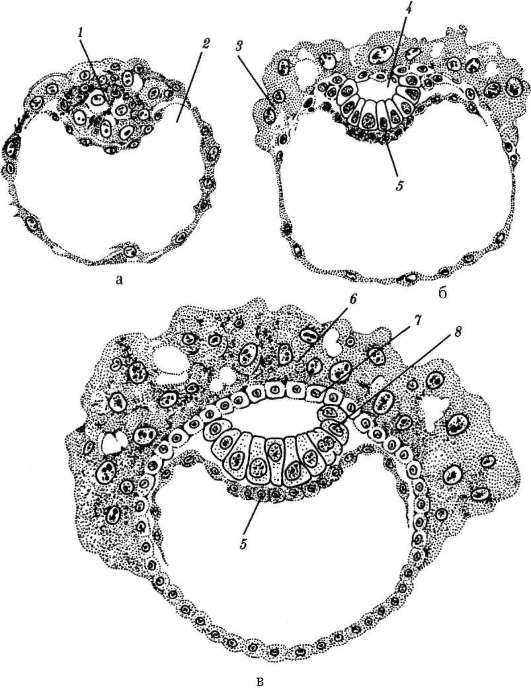
Ryža. 7.20.Postupné fázy implantácie a vývoja ľudského embrya na konci 1. a 2. týždňa: a - blastocysta; b - blastocysta na samom začiatku implantácie (7. deň vývoja); c - čiastočne implantovaná blastocysta (8. deň vývoja)
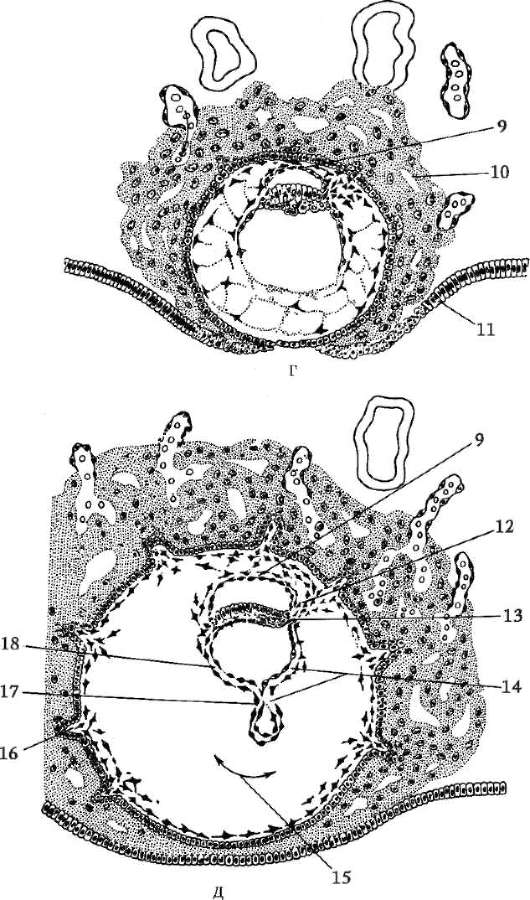
Ryža. 7.20. Koniec: d - embryo na 9-10 deň vývoja; e - embryo na 13. deň vývoja: 1 - embryoblast, 2 - blastocoel, 3 - trofoblast, 4 - amniová dutina, 5 - hypoblast, 6 - syncytiotrofoblast, 7 - cytotrofoblast, 8 - epiblast, 9 - amnion, 10 - lakuna trofoblast, 11 - epitel maternice, 12 - stopka tela, 13 - oblička alantois, 14 - žĺtkové znamienko, 15 - extradýchací coelom, 16 - choriový villus, 17 - primárny žĺtkový vak, 18 - sekundárny žĺtkový vak
naya endoderm sa úplne vynakladá na tvorbu extraembryonálneho endodermu. Vystiela dutinu trofoblastu a tvorí primárny žĺtkový vak cicavcov.
Horná bunková vrstva epiblast- zdroj budúceho ektodermu, mezodermu a sekundárneho endodermu. V 3. týždni sa tvorí epiblast primárna línia, ktorého vývoj je sprevádzaný takmer rovnakými pohybmi bunkových hmôt ako pri tvorbe primárneho pruhu vtákov (obr. 7.21). Na konci primárnej série, Hensenov uzol A primárny otvor, homológne s dorzálnym blastopórovým pyskom iných stavovcov. Bunky, ktoré sa pohybujú v oblasti primárnej jamky, sú nasmerované pod epiblast smerom k prechordálnej platni.
predkordálna platňa nachádza sa na hlavovom konci embrya a označuje miesto budúcej orofaryngeálnej membrány. Bunky pohybujúce sa pozdĺž centrálnej osi tvoria základ notochordu a mezodermu a tvoria chordomesodermový proces. Hensenov uzol sa postupne posúva smerom ku kaudálnemu koncu embrya, primárny pruh sa skracuje a rudiment notochordu sa predlžuje. Po stranách chordomeso-dermálneho výbežku sa vytvárajú mezodermálne platničky, ktoré sa rozširujú v oboch smeroch. Obrázok 7.22 predstavuje zovšeobecnený diagram niektorých procesov skorého embryonálneho vývoja.
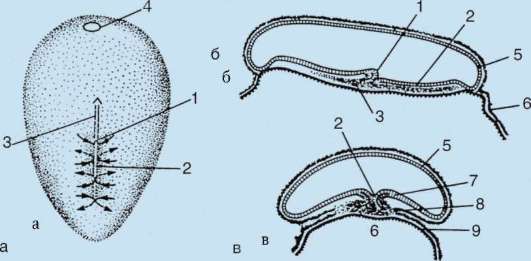
Ryža. 7.21.Vývoj ľudského embrya v štádiu primárneho pruhu (15-17 dní): a - pohľad na embryo zhora (amnion je odstránený); b - pozdĺžny rez; c - priečny rez cez primárny pás: 1 - Hensenov uzol; 2 - primárny pás; 3 - akord; 4 - prechordálna doska; 5 - amnion; 6 - žĺtkový vak; 7 - ektoderm; 8 - mezoderm; 9 - endoderm
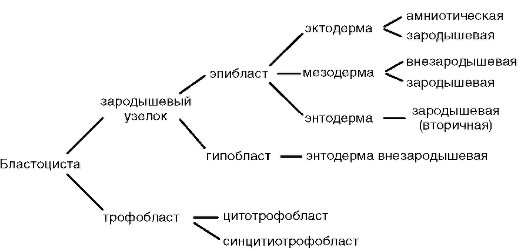
Ryža. 7.22. Diferenciácia zárodočných vrstiev cicavcov
Do konca 3. týždňa v ektoderme embrya nad rudimentom notochordu, nervová platnička. Skladá sa z vysokých cylindrických buniek. V strede nervovej platničky sa vo forme vytvorí vychýlenie nervový žľab, a po jej stranách stúpajú nervové záhyby. Toto je začiatok neurulácie. V strednej časti embrya dochádza k uzavretiu nervových záhybov – a neurálnej trubice. Potom sa uzáver rozšíri v smere hlavy a chvosta. Nervová trubica a priľahlé časti ektodermy, z ktorej sa následne vyvíja neurálny hrebeň,úplne ponorené a oddelené od ektodermy zrastajúcej nad nimi (pozri obr. 7.10). Pás buniek ležiacich pod nervovou trubicou sa mení na strunu. Po stranách notochordu a nervovej trubice v strednej časti embrya sa objavujú segmenty dorzálneho mezodermu - somity. Do konca 4. týždňa sa rozširujú na končeky hlavy a chvosta, ich počet dosahuje asi 40 párov.
Začiatok tvorby sekundárneho čreva, analizácie srdca a vaskulatúry žĺtkového vaku patrí do rovnakého obdobia. Obrázok 7.23 ukazuje pomer veľkosti embrya a extraembryonálnych orgánov v 21. deň vývoja. Podrobnejšie oddelenie tela embrya od embryonálnych membrán a uloženie orgánov je možné vidieť na obr. 7.24, ktorá zobrazuje nielen celkový pohľad na embryo, ale aj rezné plány. Pozornosť sa venuje rýchlemu (do 7 dní od 4. týždňa) formovaniu embrya vo forme predĺženého a zakriveného tela, zdvihnutého a odrezaného záhybmi trupu od žĺtkového vaku. vzadu
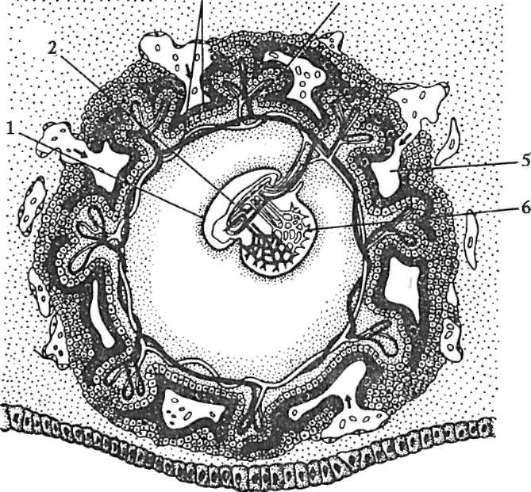
Ryža. 7.23.Embryo a extraembryonálne ľudské orgány na 21. deň vývoja: 1 - amnion; 2 - embryo; 3 - chorion; 4 - terciárny vilus; 5 - materská krv; 6 - žĺtkový vak
v tomto čase sú položené všetky somity, štyri páry žiabrových oblúkov, srdcová trubica, obličky končatín, stredné črevo, ako aj „vrecká“ predného a zadného čreva.
V nasledujúcich štyroch týždňoch embryonálneho vývoja sú položené všetky hlavné orgány. Porušenie vývojového procesu počas tohto obdobia vedie k najzávažnejším a viacnásobným vrodeným malformáciám.
Ako je uvedené vyššie, vývoj extraembryonálnych provizórnych orgánov u cicavcov a ľudí má špecifické črty. Tieto orgány sa tvoria veľmi skoro, súčasne s gastruláciou a trochu inak ako v iných amniotoch. Začiatok vývoja chorionu a amnia pripadá na 7.-8.deň, t.j. sa zhoduje so začiatkom implantácie. Chorion vzniká z trofoblastu, ktorý sa už rozdelil na cytotrofoblast a syncytiotrofoblast. Ten pod vplyvom kontaktu so sliznicou maternice rastie a ničí ju. Do konca 2. týždňa
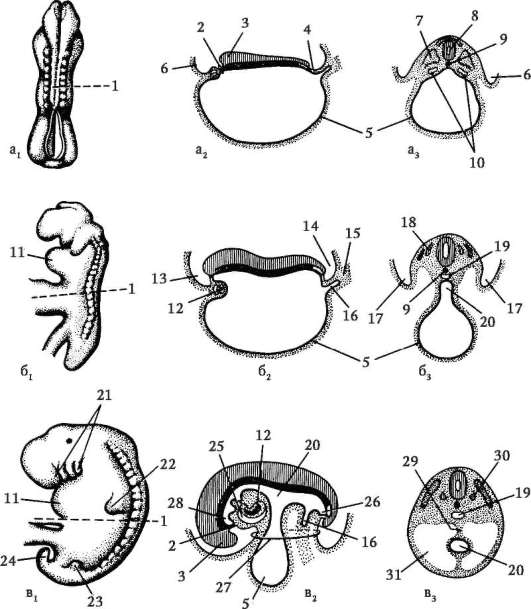
Ryža. 7.24.Vývoj ľudského embrya v 4. týždni: a 1 b 1 v 1 - všeobecný pohľad; a 2 b 2 c 2 - pozdĺžny rez; a 3 b 3 c 3 - prierez; a 1 a 2 a 3 - 22 dní; b 1 b 2 b 3 - 24 dní; v 1 v 2 za 3 - 28 dní: 1 - úroveň priečneho rezu; 2 - orofaryngeálna membrána; 3 - mozog; 4 - kloakálna membrána; 5 - žĺtkový vak; 6 - amnion; 7 - somity; 8 - nervová trubica; 9 - akord; 10 - spárované záložky brušnej aorty; 11 - srdcová rímsa; 12 - srdce; 13 - záhyb kmeňa hlavy; 14 - záhyb chvosta; 15 - noha tela; 16 - alantois; 17 - bočné záhyby kmeňa; 18 - nervový hrebeň; 19 - dorzálna aorta; 20 - stredné črevo; 21 - žiabrové oblúky; 22 - oblička prednej končatiny; 23 - oblička zadnej končatiny; 24 - chvost; 25 - osrdcovník; 26 - vrecko zadného čreva; 27 - pupočná šnúra; 28 - vrecko predného čreva; 29 - dorzálne mezenterium; 30 - nervový uzol zadného koreňa; 31 - intraembryonálny celok
primárne klky chorionu sa tvoria vo forme akumulácie cytotrofoblastových epitelových buniek. Začiatkom 3. týždňa do nich prerastá mezodermálny mezenchým a vznikajú sekundárne klky a keď sa do konca 3. týždňa objavia vo vnútri jadra spojivového tkaniva cievy, nazývajú sa terciárne klky. Oblasť, kde tesne susedia tkanivá chorionu a sliznice maternice, sa nazýva placenta.
U ľudí, ako aj u iných primátov, strácajú cievy materskej časti placenty svoju kontinuitu a choriové klky sa vlastne kúpajú v krvi a lymfe materského organizmu. Táto placenta sa nazýva hemochoriálny. Ako tehotenstvo postupuje, klky sa zväčšujú, rozvetvujú, ale krv plodu je od začiatku až do konca izolovaná od krvi matky placentárnou bariérou.
Placentárna bariéra pozostáva z trofoblastu, spojivového tkaniva a cievneho endotelu plodu. Táto bariéra je priepustná pre vodu, elektrolyty, živiny a produkty disimilácie, ako aj pre fetálne erytrocytové antigény a materské protilátky, toxické látky a hormóny. Bunky placenty produkujú štyri hormóny vrátane ľudského chorionického gonadotropínu, ktorý sa nachádza v moči tehotnej ženy od 2. – 3. týždňa tehotenstva.
Amnion vzniká divergenciou epiblastových buniek vnútornej bunkovej hmoty. Ľudský amnion sa nazýva schizamnion(pozri obr. 7.20) na rozdiel od pleuramnion vtáky a niektoré cicavce. Plodová dutina je určitý čas obmedzená epiblastovými bunkami a čiastočne miestom trofoblastu. Potom bočné steny epiblastu tvoria záhyby smerujúce nahor, ktoré sa následne spájajú. Dutina je úplne vystlaná epiblastickými (ektodermálnymi) bunkami. Vonku je amniotický ektoderm obklopený extraembryonálnymi mezodermálnymi bunkami. Plodová voda obsahuje celý rad látok, ako sú bielkoviny, sacharidy, soli, stopové prvky, hormóny atď Jej zloženie sa počas vývoja mení, navyše sa neustále aktualizuje. V procese vývoja embrya hrá dôležitú úlohu pri tvorbe nosovej a ústnej dutiny, tráviacich a dýchacích orgánov.
Žĺtkový vak sa objaví, keď sa tenká vrstva hypoblastu oddelí od vnútornej bunkovej hmoty a jej extraembryonálne endodermálne bunky, pohybujúce sa, lemujú povrch tro-
foblast. Výsledný primárny žĺtkový vak sa zrúti na 12. až 13. deň a premení sa na sekundárny žĺtkový vak spojený s embryom pomocou žĺtkového kanálika. Endodermálne bunky sú zvonka pokryté extraembryonálnym mezodermom. V 3. týždni vývoja sa v stene žĺtkového vaku začínajú zisťovať primárne zárodočné bunky, krvinky a cievy. Až do 7. – 8. týždňa vývoja je tento žĺtkový vak hlavným krvotvorným orgánom embrya.
Allantois vzniká v ľudskom embryu, podobne ako u iných amniot, vo forme vrecka vo ventrálnej stene zadného čreva, ale jeho endodermálna dutina zostáva rudimentárnou štruktúrou. Napriek tomu sa v jeho stenách vyvíja bohatá sieť ciev, ktoré sa spájajú s hlavnými krvnými cievami embrya. Mezoderm alantois sa spája s mezodermom chorionu a dáva do neho krvné cievy, ktoré poskytujú výživu vyvíjajúcemu sa embryu približne do druhého mesiaca embryogenézy. Dochádza tak k vaskularizácii chorioalantoickej placenty.
Niektoré štádiá a termíny vývoja orgánov v ľudských embryách sú uvedené v tabuľke. 7.4.
Tabuľka 7.4. Hlavné obdobia a udalosti v ranej ľudskej ontogenéze
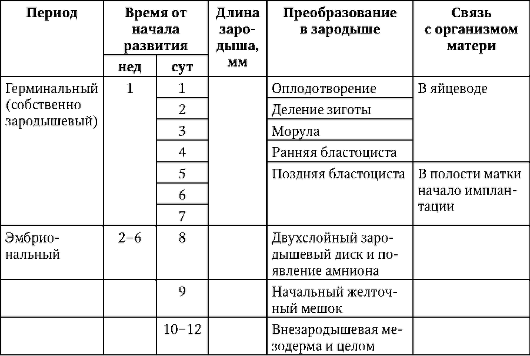
Pokračovanie tabuľky. 7.4
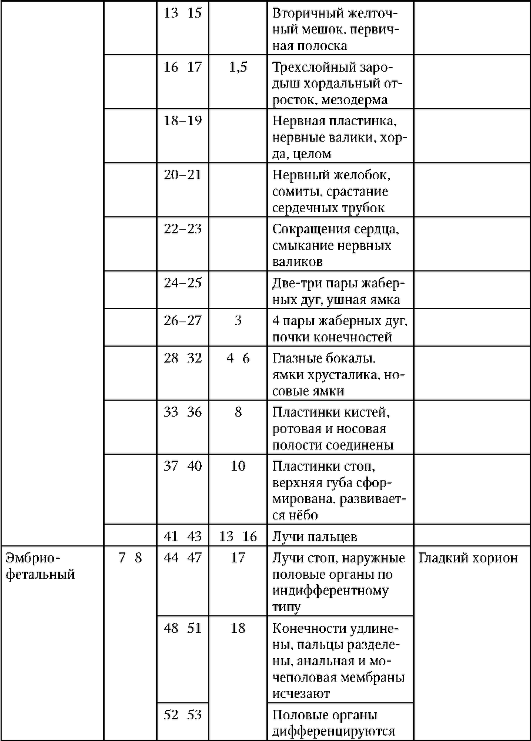
Koniec tabuľky. 7.4
7.5.2. PRÍKLADY ĽUDSKEJ ORGANOGENÉZY ODRAZUJÚCE VÝVOJ DRUHU
Táto časť nemá za cieľ popísať vývoj všetkých ľudských orgánov. Zohľadnia sa len niektoré morfogenetické procesy, ktoré ilustrujú nasledujúce všeobecné biologické body:
Význam medzibunkových, tkanivových a medziorgánových interakcií v morfogenéze;
Odraz evolučne starodávnejších znakov organizácie stavovcov v morfogenéze človeka;
Vlastnosti organogenézy, umožňujúce posúdiť štádium a mechanizmy vývojových porúch pri určitých vrodených malformáciách.
Segmentová štruktúra chrbtice odráža svoj pôvod zo somitov embrya. Okrem toho tvorba stavcov úzko súvisí s notochordom a miechou. Vertebrálne telá sú tvorené zhlukami mezenchymálnych buniek migrujúcich smerom k notochordu a obklopujúcich ho, ktoré pochádzajú zo sklerotómov. Na tvorbe tiel stavcov sa podieľajú mezenchymálne bunky dvoch susedných párov somitov, a preto sa telá stavcov nenachádzajú na úrovni somitov, ale medzi nimi. Zároveň sa svaly vyvíjajúce z myotómov somi-
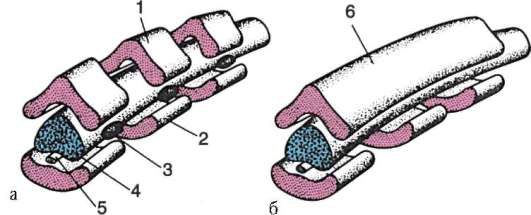
Ryža. 7.25. Vývoj chrbtových oblúkov stavcov u kuracieho embrya: a - normálny vývoj; b - po odstránení spinálnych ganglií: 1 - chrbtový oblúk; 2 - telo stavca; 3 - miechový uzol; 4 - miecha; 5 - akord; 6 - nesegmentované chrbtové oblúky stavcov
comov, akoby prehodený cez medzistavcové kĺby a zabezpečiť pohyb stavcov.
Stavcové oblúky sú tvorené nahromadením mezenchymálnych sklerotomických buniek šíriacich sa dorzálnym (dorzálnym) smerom, zatiaľ čo priečne výbežky a rebrá sú tvorené nahromadením sklerotomických buniek migrujúcich do strán. Miecha a miechové gangliá v príslušných štádiách vnútromaternicového vývoja sa podieľajú na morfogenéze chrbtových oblúkov stavcov. Ak sa teda odstránia spinálne gangliá, vytvorí sa chrupka chrbtových oblúkov, ale vyzerá ako nesegmentovaná tyčinka (obr. 7.25).
Postupne sa mení histologická štruktúra stavcov. Počas 4.-5. týždňa sa tvoria mezenchymálne stavce, v 6. týždni vznikajú ložiská chondrogenézy a následne v 8. týždni začína osifikácia, ktorá trvá až takmer 25 rokov. Úseky chordy umiestnené vo vnútri tiel stavcov postupne miznú. V rámci medzistavcových platničiek je notochord zachovaný ako štruktúra podobná hlienu známa ako nucleus pulposus.
Tvorenie stredného ucha spojené s ontogenetickými premenami prvého a druhého viscerálne (žiabrové) oblúky. Obrázok 7.26 schematicky znázorňuje oblasti hlavy a krku v 4. a 24. týždni (koniec prvého a druhého trimestra prenatálneho vývoja) týždni ľudského vývoja. V 4. týždni sú viscerálne (žiabrové) oblúky položené v oblasti hlavy a krku embrya vpravo a vľavo. Z vnútra, zo strany hltana, medzi nimi sú vytvorené hltanové vrecká(obr. 7.27). Sú to výbežky (alebo výbežky) endodermu,
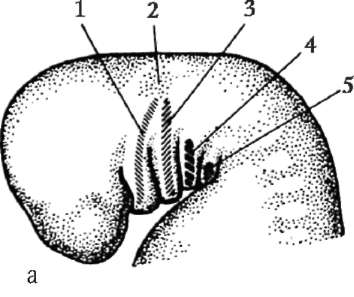
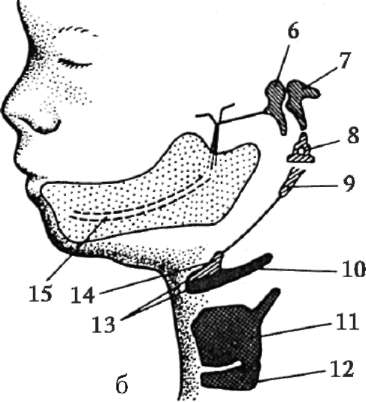
Ryža. 7.26.Hlava a krk ľudského embrya, bočný pohľad: a - v 4. týždni; b - v 24. týždni: 1 - prvý oblúk; 2 - umiestnenie vyvíjajúceho sa vnútorného ucha; 3 - druhý oblúk; 4 - tretí oblúk; 5 - štvrté lúky; 6 - kladivo. 7 - kovadlina; 8 - strmeň; 9 - styloidný proces; 10 - veľký roh hyoidnej kosti; 11 - chrupavka štítnej žľazy; 12 - kricoidná chrupavka; 13 - telo hyoidnej kosti; 14 - malý roh; 15 - spodná čeľusť; úpony viscerálnych oblúkov (a) a orgány z nich vyvíjajúce (b) sú označené rovnakým tieňovaním
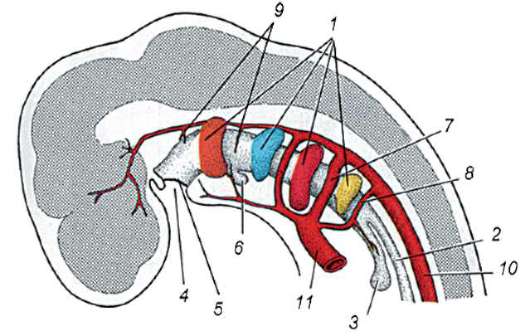
Ryža. 7.27.Žiabrová oblasť 5-týždňového ľudského embrya (odstránená krycia ektoderma a mezenchým). 1 - hltanové vrecká; 2 - pažerák; 3 - priedušnica a pľúcna oblička; 4 - stomodeum; 5 - stomodálna (orálna) doska; 6 - rudiment štítnej žľazy; 7 - 4. oblúk aorty; 8 - 6. oblúk aorty; 9 - zmenšenie 1. a 2. oblúka aorty; 10 - dorzálna zostupná aorta; 11 - brušná vzostupná aorta (arteriálny vak)
rastúce bokom. Vonku podľa výrastkov hltana ektodermálne žiabrové vrecká.
Endoderm a ektoderm prvých hltanových a žiabrových vačkov prichádzajú do kontaktu v 4. týždni vývoja (obr. 7.28, a). Kontakt netrvá dlho, slepý koniec hltanovej kapsy sa vzďaľuje od povrchu a je obklopený mezenchýmom (obr. 7.28, b). Z distálnej časti hltanovej kapsy sa začína vyvíjať stredoušná dutina (bubienková dutina) a z proximálnej časti vzniká sluchová (Eustachovská) trubica. Z mezenchýmu sa ďalej rozlišujú analáže sluchových kostičiek, ktoré slúžia na vedenie zvuku (obr. 7.28, c). Kladivo a kovadlina pochádzajú z bunkového materiálu prvého viscerálneho oblúka, strmeňa - z bunkového materiálu dorzálnej časti druhého viscerálneho oblúka. Z bunkového materiálu oblasti prvého a druhého viscerálneho oblúka sú tvorené svaly a nervy spojené so zvukovovodným kostným aparátom pomocou kostičiek stredného ucha.
Na konci vnútromaternicového života a v priebehu niekoľkých mesiacov po narodení sa embryonálne spojivové tkanivo v bubienkovej dutine resorbuje. Až potom získajú sluchové kostičky pohyblivosť a vibrácie bubienka sa môžu voľne prenášať na membránu oválneho okienka vnútorného ucha.
Pri tvorbe štruktúr oblasť tváre A ústna dutina sa zúčastňujú ekto- a mezodermálne bunky. Väčšina zštruktúru
Ryža. 7.28. Vývoj stredného ucha, frontálny úsek: a - v 4. týždni; b - v 5. týždni; c - neskoršie štádiá; 1 - prvá žiabrová doska; 2 - prvé žiabrové vrecko; 3 - prvý viscerálny (žiabrový) oblúk; 4 - sluchová vezikula; 5 - prvé hltanové vrecko; 6 - deriváty prvého viscerálneho oblúka; 7 - derivát druhého viscerálneho oblúka; 8 - dutina potrubia-slave; 9 - zátka vonkajšieho zvukovodu. 10 - primárny vonkajší sluchový meatus; 11 - kladivo; 12 - kovadlina; 13 - strmeň; 14 - bubienková dutina; 15 - sluchová (Eustachovská) trubica
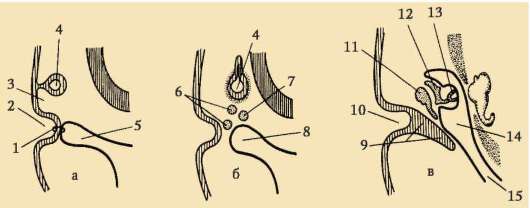
prehliadka oblasti tváre sa vyvíja z masy mezenchymálnych buniek ektodermálneho pôvodu migrujúcich z neurálnej lišty (4. zárodočná vrstva). Z tohto mezenchýmu sa vyvíjajú spojivové, chrupavkové a kostné tkanivá a tkanivo zubnej drene. Z mezenchýmu mezodermálneho pôvodu sa tvoria priečne pruhované svaly svalov tváre a hltana. Tu je dôležité pochopiť, ako prebiehal proces „expanzie a zmeny funkcií“ v antropogenéze.
Ústna dutina vzniká ektodermálnou invagináciou stomo-deum, susediace so slepo končiacim predžalúdkom. Sú oddelené tenkými stomodálna platnička, pozostávajúce z ektodermálnych (zo strany stomodea) a endodermálnych (zo strany čreva) plátov (pozri obr. 7-26). Táto platnička sa na konci 4. týždňa vývinu zlomí a ústna dutina sa napojí na pôvodne slepo uzavretú črevnú trubicu.
Príušné slinné žľazy sa vyvíjajú z ektodermy stomodea a sublingválne a submandibulárne slinné žľazy sa vyvíjajú z endodermy ventrálnej časti hltana.
U 5 týždňového plodu otvorenie úst ohraničené nasledujúcimi štruktúrami (obr. 7.29, a): v strednej čiare nad ústnou dutinou je zaoblená previsnutá oblasť - čelná (čelná) rímsa; po jeho stranách sú podkovovité vyvýšeniny obklopujúce čuchové jamky. Stredné procesy týchto vyvýšenín
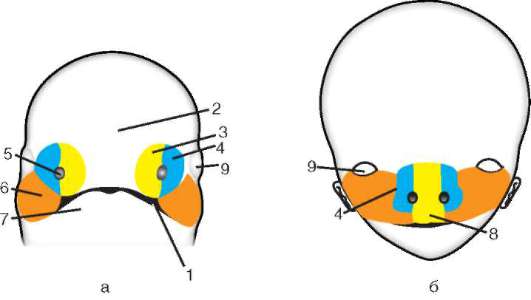
Ryža. 7.29. Postupné fázy formovania tváre (predný pohľad): a - v 5. týždni; b - v 10. týždni. 1 - stomodeum; 2 - čelná (čelná) rímsa; 3 - mediálny nazálny proces; 4 - laterálny nazálny proces; 5 - čuchová jamka; 6 - maxilárny proces; 7 - položenie mandibulárneho oblúka; 8 - filtrum; 9 - oko
volal mediálny nos, a tie vedľajšie bočné
nosové procesy. Z horných bočných rohov ústnej dutiny smerom k stredovej línii vyrastajú maxilárne procesy.
Obmedzenie ústnej dutiny zospodu spodná čeľusť vytvorený z párové záložky mandibulárneho oblúka. Najprv sa objavujú na každej strane stredovej čiary, potom sa zväčšujú smerom k sebe a rastú spolu.
Počas 6. týždňa dochádza k rýchlemu vývoju Horná čeľusť. Maxilárne procesy rastú smerom k strednej čiare. Súčasne sa zvyšujú nosové výbežky, ktoré tlačia nahor spodnú časť čelného výčnelku umiestneného medzi nimi. Obzvlášť intenzívne rastú mediálne nazálne výbežky, ktoré sa posúvajú k strednej línii a navzájom sa spájajú, a potom sa spájajú s čeľustnými výbežkami, čím sa v 10. týždni vývoja vytvorí kompletný čeľustný oblúk. Vytvára sa kožný ektoderm nad zlúčenými mediálnymi výbežkami filtrum- pozdĺžna ryha nad hornou perou pod nosom (obr. 7.29, b).
Do konca druhého mesiaca vývoja, keď sa tvorí horná čeľusť, sa začína rozvíjať pevná obloha. Na jej vzniku, ako aj na tvorbe oblúka hornej čeľuste sa podieľajú stredné nosové a maxilárne procesy(Obr. 7.30, a). Zo strednej oblasti hornej čeľuste, ktorá vznikla v dôsledku fúzie stredných nosových výbežkov, sa vytvorí malá trojuholníková časť - primárne podnebie. Hlavná časť - sekundárne podnebie- sa vyvíja z bočné palatinové výrastky (procesy), ktoré sa objavujú na hornej čeľusti (obr. 7.30, b). Najprv sú nasmerované nadol a potom okraje palatinových procesov stúpajú nahor a smerom k stredovej čiare. Dotýkajú sa a spájajú sa a vytvárajú steh podnebia. Vytvorená hlavná časť podnebia oddeľuje najvrchnejšiu časť pôvodnej stomodálnej dutiny. Sekundárne podnebie rastie spolu s primárnym a nosová priehradka prirastá k ich kraniálnemu povrchu (obr. 7.30, c). Tak sa súčasne s oddelením nosovej dutiny od ústnej dutiny rozdelí na pravú a ľavú polovicu.
V počiatočnom období svojho vzniku (v 5. týždni embryogenézy) Jazyk je vak tvorený sliznicou, vyplnený rastúcou svalovou hmotou. Predpokladá sa, že sliznica jazyka vzniká z stomodeum ektoderm, a svaly pochádzajú okcipitálne myotómy.
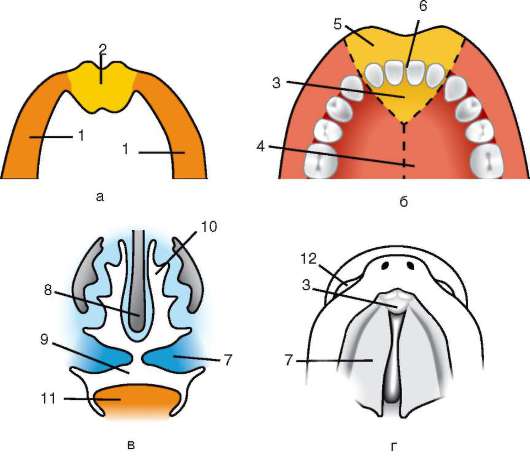
Ryža. 7.30. Tvorba tvrdého podnebia: a - schematické znázornenie hornej čeľuste (pohľad zo strany ústnej dutiny); b - 10. týždeň vývoja (pohľad z ústnej dutiny); c - 8. týždeň vývinu (čelný rez); d - 8. týždeň vývoja (pohľad z ústnej dutiny): 1 - čeľustný výbežok; 2 - zlúčené mediálne nazálne procesy; 3 - primárna obloha; 4 - sekundárne podnebie (laterálne palatinové výrastky); 5 - lip filtrum; 6 - horná čeľusť s rezákmi; 7 - bočný palatínový výrastok; 8 - nosná priehradka; 9 - ústna dutina; 10 - nosná dutina; 11 - jazyk; 12 - oko
Na tvorbe sliznice jazyka sa podieľa viacero štruktúr dna ústnej dutiny. Telo jazyka je tvorené fúziou párových bočných zhrubnutí - bočné lingválne tuberkulózy a nachádza sa medzi nimi malá stredová eminencia, tzv tuberculum impar(obr. 7.31). Predpokladá sa, že telo jazyka je derivátom bunkového materiálu oblasti prvého viscerálneho oblúka. Podieľa sa na tvorbe koreňa jazyka coula(zátvorka)- stredová eminencia, umiestnená vzadu tuberculum impar a zapnite
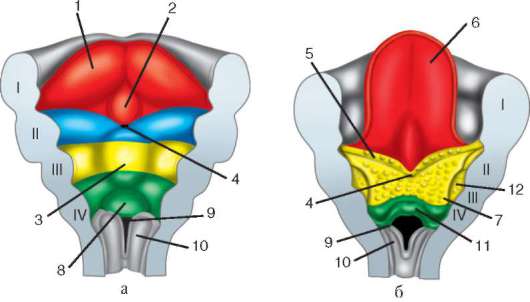
Ryža. 7.31. Vývoj jazyka: a - 5-týždňové embryo; b - 5-mesačný plod; 1 - bočný lingválny tuberkul; 2- tuberculum impar 3 - kopula (zátvorka); 4 - foramen coecum; 5 - koncová brázda; 6 - telo jazyka; 7 - koreň jazyka; 8 - položenie epiglotickej chrupavky; 9 - hlasivková štrbina; 10 - arytenoidný tuberkul; 11 - epiglottis, palatinová mandľa. I-IV - počty viscerálnych oblúkov
bunkový materiál oblastí druhého, tretieho a štvrtého viscerálneho oblúka. Po fúzii všetkých analáží a konečnej formácii jazyka sa jeho telo oddelí od koreňa terminálna brázda(sulcus terminalis), v strede ktorého je slepá diera(foramen coecum)- stopa po invaginácii dna hltana, z ktorého vystupuje štítna žľaza (pozri obr. 7.27). Je to to, čo sa považuje za hranicu medzi telom a koreňom jazyka.
Neskôr sa jazyk takmer úplne oddelí od dna ústnej dutiny a spojenie medzi nimi sa zachová v podobe záhybu sliznice – uzdičky jazyka.
Zaujímavým príkladom organogenézy je formácia zuby, na ktorom sa podieľa integumentárny ektoderm - epitel stomodea (sklovina) a mezenchymálne bunky pochádzajúce z neurálnej lišty (dentín, cement a pulpa) (obr. 7.32). V priebehu vývoja sú zaznamenané indukčné interakcie medzi mezenchymálnymi a epitelovými štruktúrami.
Počas života sa vyvinú dve generácie zubov. K zmene chrupu dochádza v detskom veku, prvé zúbky sú tzv mliekareň. Je ich 20: 10 v hornej a 10 v dolnej čeľusti. Mliečne zuby úplne prerezávajú vo veku približne 2 rokov. Tento chrup slúži dieťaťu ďalšie 4 roky, potom mliečny chrup postupne
Ryža. 7.32.Ľudské trvalé zuby: 1 - rezáky; 2 - tesáky; 3 - malé stoličky; 4 - veľké stoličky
ale vypadnú a sú nahradené trvalé zuby(druhá generácia zubov), ktoré fungujú v človeku po celý život. Obdobie výmeny zubov trvá približne od 6 do 12 rokov. 32 stálych zubov: 16 horných a 16 dolných (pridávajú sa veľké stoličky) (pozri obr. 7.32). Tvarom sú podobné mliečnym, no sú väčšie.
Záložka a vzdelávanie zubné zárodky mliečne zuby začínajú od 6-7 týždňov vnútromaternicového vývoja. Epitel ústnej dutiny je ponorený do podkladového mezenchýmu vo forme hustého vlákna tzv. zubná platnička(obr. 7.33). Na zubnej platničke sa objavujú malé epiteliálne výbežky - zubné puky, z ktorých sa vyvinú mliečne zuby. Ako zubná vrstva rastie, každý zubný zárodok sa zväčšuje, preniká hlbšie do mezenchýmu a nadobúda tvar obrátenej misky. Táto štruktúra sa tvorí smaltovaný organ, a vytvorí sa spodný mezenchým, ktorý vypĺňa dutinu pohárika zubná papila. V ďalšom vývoji tvorí ten druhý zubnú dreň. V 3. mesiaci vnútromaternicového vývoja sa orgán skloviny zväčšuje, mení tvar a postupne sa oddeľuje od zubnej platničky. Tvar budúceho zuba je určený mezenchymálnou zložkou. Pri kombinácii in vitro mezenchým uloženia črenového zuba (molára) s epiteliálnou zložkou uloženia rezáka, vzniká molár. Naopak, v dôsledku spojenia ektodermy moláru a mezenchýmu rezáka vzniká rezák.
Počas histogenézy zubných tkanív, ktorá začína koncom 4. mesiaca, dentín, sklovinu a dreň(Obr. 7.33, d). Smalt
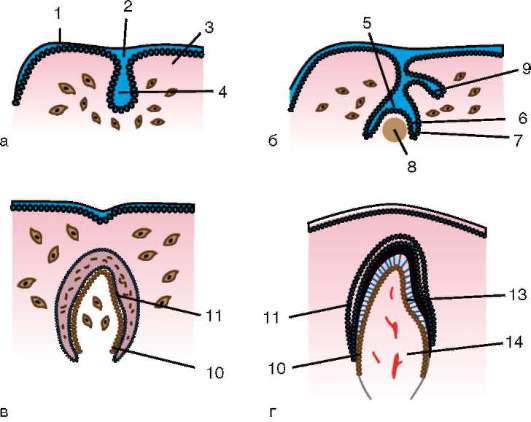
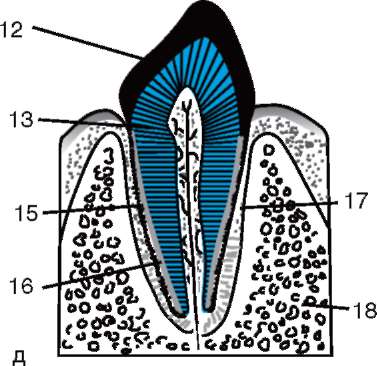
Ryža. 7.33.Vývoj zubov: a - 8-týždňové embryo; b - 10-týždňový plod; c - 3-mesačný plod; g - 6-mesačný plod; d - po erupcii; 1 - epitel ústnej dutiny; 2 - zubná doska; 3 - mezenchým; 4 - zárodok mliečneho zuba; 5 - smaltovaný orgán; 6 - vnútorná vrstva orgánu skloviny; 7 - vonkajšia vrstva orgánu skloviny; 8 - zubná papila; 9 - zárodok trvalého zuba; 10 - odontoblasty; 11 - ameloblasty; 12 - smalt; 13 - dentín; 14 - zubná dreň; 15 - cementoblasty; 16 - cement; 17 - parodontálna; 18 - alveolárna kosť
produkujú bunky vnútornej vrstvy orgánu skloviny - smalt výbuchy. Ich procesy vylučujú organický základ skloviny – sklovinové hranoly, ktoré následne kalcifikujú. Zdroj rozvoja dentín- odontoblasty (dentinoblasty), bunkami povrchovej miazgy. Vrchol deninoblastov má tiež procesy, ktoré vylučujú organické látky fibrilárnej štruktúry, tzv predentin a vytvorenie matrice dentínu. Od ukončeného 5. mesiaca sa v predentíne ukladajú soli vápnika a fosforu a vzniká finálny dentín. Tvorba dentínu a skloviny sa líši od vývoja kosti tým, že bunky nie sú zakryté medzibunková látka, ale vzdiaľte sa: ameloblasty - smerom von, odontoblasty - dovnútra. Okrem toho sa obsah organických látok v týchto tkanivách líši: kosť obsahuje približne 45% organickej hmoty, dentín - 28% a sklovina - menej ako 5%.
Na vonkajšom povrchu dentínu z mezenchýmu obklopujúceho zub (zubný vačok) sa objavia krátko pred prerezávaním zúbkov cementoblasty. Tieto bunky vylučujú kolagénové vlákna a tvoria sa medzibunkové látky cement. Tvorí sa až 4-5 mesiacov po narodení počas vývoja koreňov zubov. Z vonkajšej vrstvy zubného vaku vzniká zub. väzivo (parodont)(Obr. 7.33, e). V prenatálnom vývoji sa teda vyvíjajú iba korunky mliečnych zubov.
Erupcia zubov je zabezpečená tromi faktormi. Prvým a najdôležitejším je rast a vývoj koreňa, vedúci k vypudeniu korunky cez sliznicu. Druhým faktorom je rast zubnej drene, čo vedie k zvýšeniu tlaku vo vnútri zubného zárodku. Okrem toho dochádza k ďalšiemu ukladaniu na dne zubnej alveoly. kostného tkaniva vrstvy.
Kladenie trvalých zubov začína veľmi skoro, na začiatku 5. mesiaca vnútromaternicového vývoja. Zárodok trvalého zuba sa nachádza za zárodkom mliečneho zuba a jeho vyvíjajúci sa koreň v prvých dvoch rokoch po narodení má indukčný vplyv na vývoj zárodku stáleho zuba. Histologické procesy, ktorých výsledkom je tvorba mliečnych a trvalých zubov, sú rovnaké. Pri prerezávaní mliečnych zubov sa v základoch trvalých zubov tvorí sklovina a dentín. Po určitú dobu zostávajú v latentnom stave a aktivujú sa až potom, čo čeľusť dosiahne dostatočne veľkú veľkosť. V procese výmeny vedie rast trvalého zuba a tlak jeho skloviny na koreň mlieka k resorpcii.
osteoklasty (bunky, ktoré ničia kostné tkanivo) mäkšieho tkaniva – dentínu mliečneho zuba. Ten je vytlačený a nahradený trvalým.
Rozvoj oddelenia tráviaci Systémy je ťažké podrobne zvážiť, pretože črevná trubica je značne predĺžená, diferencovaná a vo svojom vývoji spojená s mnohými ďalšími systémami: endokrinným, respiračným, vylučovacím, obehovým, nervovým a sekundárnou dutinou - coelom. Uvažujme len o najskorších a najbežnejších procesoch vyskytujúcich sa v črevnej trubici, ako aj o zmenách v jej spojení so žĺtkovým vakom.
Črevná trubica vychádza z endodermu stropu žĺtkového vaku a priľahlej viscerálnej vrstvy mezodermu v procese oddeľovania tela embrya od extraembryonálnych častí pomocou záhybov hlavy, chvosta a bočných kmeňov (pozri obr. 7.24, b 2, b 3). V 4. týždni je reprezentovaná relatívne jednoduchou trubicou pozostávajúcou z predžalúdka, spredu slepo uzavretej stomodálnej (ústnej) platničky, stredného čreva spojeného so žĺtkovým vakom.
cez stopku žĺtka (kanál) a zadné črevo, slepo uzavreté na konci chvosta kloakálnou (análnou) membránou
(obr. 7.34).
V období od 4. do konca 8. týždňa embryonálneho (vnútromaternicového) vývoja sa črevo predlžuje a začína sa diferencovať. Z jeho prednej časti sa tvorí hltan, pažerák, žalúdok a dvanástnik s jeho derivátmi (pečeň a pankreas) skôr, ako do neho vtečie spoločný žlčovod.
Vytvára sa stredné črevo V tvare U-rôzne slučku, ktorá je spojená so žĺtkovým vakom cez žĺtkový kanál (obr. 7.35, a). Úsek čreva medzi stopkou žĺtka a žalúdkom sa zmení na tenké črevo a úsek
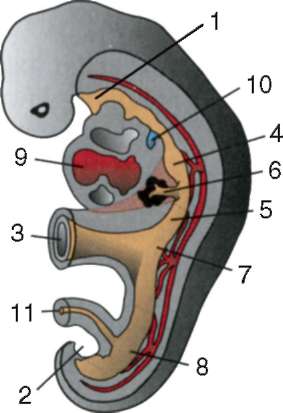
Ryža. 7.34.Tráviaci systém 4-týždňového embrya: 1 - stomo-deal tanier; 2 - kloakálna membrána; 3 - žĺtkový kanál; 4 - žalúdok; 5 - dvanástnik; 6 - pečeňový rudiment; 7 - stredné črevo; 8 - zadné črevo; 9 - srdce; 10 - položenie dýchacích orgánov; 11 - urachus
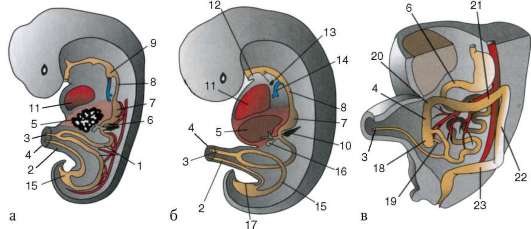
Ryža. 7.35.Vývoj tráviaceho systému človeka: a - na 32. deň; b - na 36. deň; c - neskoršie štádiá; 1 - slučka čreva v tvare U; 2 - urachus; 3 - žĺtkový kanál; 4 - brušná stopka (pupočná šnúra); 5 - pečeň; 6 - dvanástnik; 7 - žalúdok; 8 - pažerák; 9 - hrtan; 10 - pankreas; 11 - srdce; 12 - jazyk; 13 - záložka štítnej žľazy; 14 - rudiment priedušnice a pľúc; 15 - zadné črevo; 16 - žlčník; 17 - kloakálna membrána; 18 - slepé črevo; 19 - príloha; 20-23 - úseky hrubého čreva
priliehajúce kaudálne k vitelinovému kanáliku, - do hrubého čreva. Stredné črevo sa predlžuje, jeho časť vyčnieva do dutiny brušnej stopky a tvorí herniálny výbežok brušnej steny embrya.
Počas toho istého času niektoré časti čreva, ktoré sa otáčajú, menia svoju polohu (obr. 7.35, b). V 10. týždni sa vyčnievajúca časť črevnej kľučky stiahne späť cez pupočný okraj a zaujme svoju konečnú polohu v brušnej dutine. O niečo skôr, v mieste prechodu tenkého čreva do hrubého čreva, sa vyvíja slepé črevo. Najprv sa zväčšuje, ale do 3. mesiaca jej distálna časť začína zaostávať za rastom ostatných častí (nerovnomerný rast), v dôsledku čoho je priemer tejto časti oveľa menší a slepé črevo je tvorené. Zo zadného čreva sa tvorí hrubé črevo pod slezinným uhlom vrátane konečníka (obr. 7.35, c). Vývoj kloakálneho konca zadného čreva úzko súvisí s vývojom urogenitálneho otvoru (obr. 7.36). Urorektálna priehradka, ktorá oddeľuje kloaku na urogenitálny sínus a konečník, sa vytvára v 5. – 6. týždni. Prielom análnej membrány nastáva v 8. týždni.
V 2. mesiaci vnútromaternicového vývoja začína rýchla proliferácia epitelu pažeráka a dvanástnika, čo
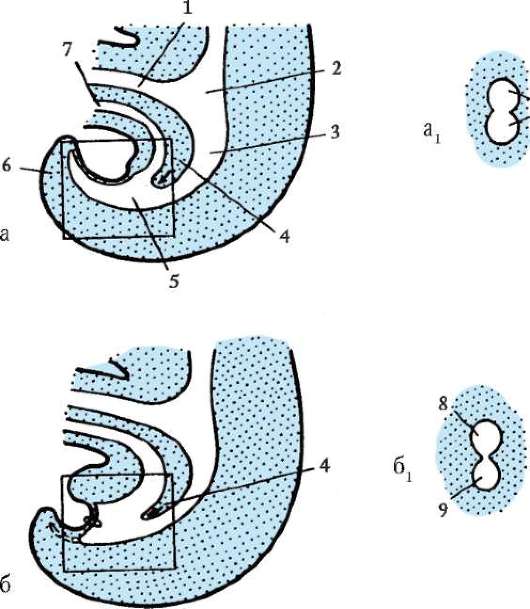
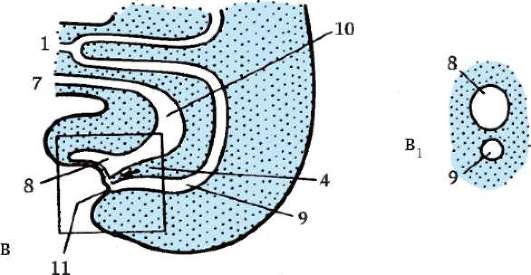
Ryža. 7.36. Vývoj kloakálnej oblasti zadného čreva v ľudskom embryu: a - v 4. týždni; b - v 6. týždni; c - v 7. týždni; a 1 b 1 a c 1 - priečne rezy kloakou (úrovne rezu sú označené štvorčekmi na obr. a; b; c). 1 - stopka žĺtka; 2 - stredné črevo; 3 - zadné črevo; 4 - urogenitálna priehradka; 5 - kloaka; 6 - chvost; 7 - alantois; 8 - urogenitálny sínus; 9 - konečník; 10 - močového mechúra; 11 - análna membrána; šípky ukazujú smer rastu
vedie k dočasnému uzavretiu ich lúmenu. Do konca 2. mesiaca nastáva rekanalizácia týchto orgánov, t.j. obnovenie priepustnosti.
Srdcečlovek sa začína vyvíjať od konca 3. - začiatku 4. týždňa zo splanchnomesodermu vo forme párových rudimentov umiestnených pod hltanom (obr. 7.37). Keďže telo samotného embrya je ohraničené záhybmi trupu a predžalúdočné črevo je uzavreté z ventrálnej strany, párové endokardiálne trubice sa uzatvárajú do jednej, ležiacej pozdĺž stredovej čiary (pozri obr. 7.37). U obojživelníkov bola dokázaná existencia indukčného pôsobenia endodermy na prekordiálnu mezodermu. Dokazuje to aj pokus na kuracom embryu, kedy sa po odstránení spodnej časti predžalúdka získalo embryo s dvojitým srdcom. Párové úpony coelomických dutín sa spájajú na ventrálnej strane a vytvárajú perikardiálnu dutinu.
Regionálna diferenciácia srdca začína rýchlym predĺžením primárnej srdcovej trubice, čo vedie k jej ohybu a získaniu tvaru 5 (obr. 7.38). Je potrebné poznamenať, že tvorba ohybu s lokálnymi zmenami tvaru buniek pozdĺž srdcovej trubice je stimulovaná zo samotnej srdcovej trubice. Byť explantovaný (situácia „mimo tela“, príp ex vivo), srdcová trubica si zachováva schopnosť vytvárať štruktúru v tvare 5.
Ryža. 7.37. Vývoj ľudského srdca na 21.-22. deň: a - celkový pohľad na embryo; b - pozdĺžny rez hlavovým kužeľom embrya; c - prierez embrya: 1 - úroveň rezu; 2 - nervový valec; 3 - somity; 4 - primárny pás; 5 - mozog; 6 - predžalúdky; 7 - srdcová trubica; 8 - osrdcovník; 9 - amnion; 10 - orofaryngeálna membrána; 11 - nervová drážka; 12 - akord; 13 - spárované záložky dorzálnej aorty; 14 - spájanie srdcových rúrok; 15 - žĺtkový vak
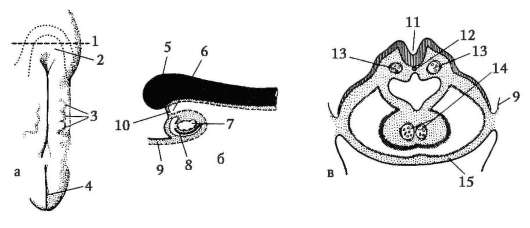
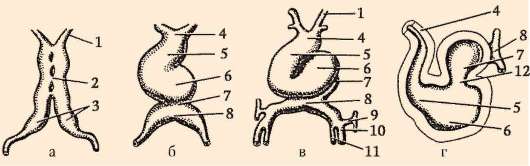
Ryža. 7.38. Vyvíjajúce sa ľudské srdce: a - na 21.-22. deň; b - na 23. deň; c - na 24. deň (pohľad z ventrálnej strany); d - na 28. deň (pozdĺžny rez): 1 - prvý pár aortálnych oblúkov; 2 - spájanie srdcových rúrok; 3 - nezlučujúce sa srdcové trubice; 4 - arteriálny kmeň; 5 - žiarovka srdca; 6 - komora; 7 - átrium; 8 - venózny sínus; 9 - Cuvierov kanál; 10 - pupočná žila; 11 - žĺtková žila; 12 - atrioventrikulárny kanál
Na pôvodne kaudálnom konci je venózny sínus, do ktorého prúdia veľké žily: Cuvierove vývody, pupočník a vitelin. Venózny sínus sa neskôr stane súčasťou pravej predsiene. Kraniálny k venóznemu sínusu je predsieň vytvorená z rozšírenej časti srdcovej trubice a komora je vytvorená zo zakrivenej strednej časti. Prechodná oblasť, kde sa komora zužuje, sa nazýva kužeľ alebo žiarovka. Neskôr sa začlení do steny pravej komory. Kužeľ prechádza do arteriálneho kmeňa, z ktorého odchádzajú korene brušnej aorty.
Po vytvorení predsiene a komory sa objavia vonkajšie príznaky nadchádzajúceho rozdelenia srdca na pravú a ľavú polovicu, objaví sa stredný sulcus. V tejto chvíli je v tkanivách vyvíjajúceho sa srdca zaznamenaná fokálna bunková smrť a interakcia extracelulárnych a bunkových prvkov faktorov. O nejaký čas neskôr sa na vnútornom povrchu komory, zodpovedajúcej úrovni vonkajšej drážky, vytvorí prepážka zo svalových povrazov rastúcich od srdcového hrotu smerom k predsieni. Na dorzálnych a ventrálnych stenách zúženého atrioventrikulárneho kanála sa z uvoľneného mezenchýmu tvoria endokardiálne vankúšiky. Neskôr sa premenia na husté spojivové tkanivo, rastú spolu a rozdeľujú kanál na pravý a ľavý kanál. Súčasne sa objaví primárna stredná priehradka, ktorá je neskôr nahradená sekundárnou. Má otvor, nazývaný ovál, cez ktorý
krv z pravej predsiene prúdi do ľavej. Je nevyhnutný pre krvný obeh plodu.
Súčasne so zmenami v hlavnej časti srdca ako nezávislého orgánu je arteriálny kmeň rozdelený na dva kanály. Tento proces začína pri koreni brušnej aorty medzi štvrtým a šiestym viscerálnym oblúkom. K oddeleniu dochádza v dôsledku tvorby pozdĺžnych záhybov. Záhyby sú usporiadané do špirály, rastú vo vnútri arteriálneho kmeňa a pri stretnutí ho rozdeľujú na aortu a pľúcny kmeň. Okrem toho sa záhyby šíria smerom ku kužeľu, kde sa zo špecializovaných oblastí tvoria semilunárne chlopne aorty a pľúcneho kmeňa a ďalej do komôr, kde sa stretávajú s pohyblivým endokardiálnym tkanivom atrioventrikulárneho kanála a predsieňového septa. To vedie k úplnému prerastaniu medzikomorového otvoru.
Do konca vnútromaternicového života zostáva otvorená iba chlopňa vo foramen ovale interatriálneho septa a arteriálny (botall) vývod, ktorý spája kmeň pľúcnice s aortou. Podrobnejšie sa o tom bude diskutovať nižšie. Arteriálny kanál slúži na odvádzanie časti krvi z pravej komory do aorty, obchádzajúc pľúca, kým nedosiahnu potrebný vývoj a nedostanú funkčný stimul v súvislosti s nástupom respiračnej funkcie (obr. 7.39).
Veľké tepny vyvíjať v kombinácii so srdcom, počnúc 4. týždňom. Primárny obehový systém embrya funguje od konca 5. týždňa, do konca 8. týždňa sa realizuje základný plán štruktúry arteriálny systém. Párové viscerálne oblúky vytvorené v 4. týždni dostávajú primerané arteriálne zásobenie vo forme aortálnych oblúkov. Tieto oblúky aorty vychádzajú z kmeňa tepny, alebo skôr z jeho rozšírenej časti - arteriálneho vaku (nazývaného aj brušná alebo vzostupná aorta) - a končia v koreňoch dorzálnej alebo zostupnej aorty (pozri obr. 7.27).
Napriek tomu, že je položených šesť párov aortálnych (viscerálnych) oblúkov, nekoexistujú súčasne. V období, keď sa tvorí šiesty pár oblúkov, prvé dva páry už degenerujú. Piaty oblúk sa zobrazí iba na krátky čas vo forme rudimentárnej nádoby.
Pri tvorbe hlavných ciev zohrávajú podstatnú úlohu korene brušnej a dorzálnej aorty, ako aj tretí, štvrtý a šiesty pár aortálnych oblúkov (obr. 7.40).
Proximálne časti tretieho páru tvoria spoločné krčné tepny (obr. 7.41). Ľavý štvrtý oblúk tvorí časť oblúka aorty.
Ryža. 7.39. Schéma fetálnej cirkulácie: 1 - oválne okno; 2 - interatriálna priehradka; 3 - pľúcna žila; 4 - horná dutá žila; 5 - arteriálny (botalový) kanál; 6 - pľúcna tepna; 7 - zostupná aorta; 8 - pupočníkové tepny; 9 - pupočná žila; 10 - dolná dutá žila; jedenásť - ductus venosus; 12 - portálna žila pečene. I - pečeň; II - dolná dutá žila; III - pravá predsieň; IV - ľavá predsieň; V - zostupná aorta
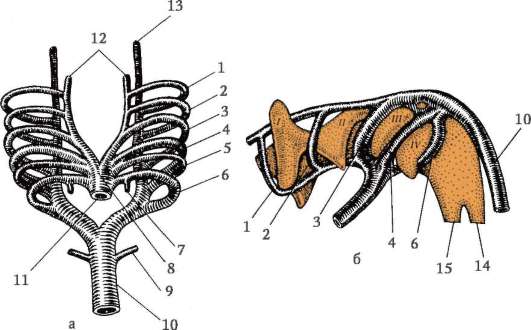
Ryža. 7,40. Vývoj aortálnych oblúkov: a - pohľad z ventrálnej strany: 1-6 - aortálne oblúky; 7 - pľúcna tepna; 8 - arteriálny vak, brušná aorta; 9 - siedma intersegmentálna artéria, subclavian; 10 - dorzálna zostupná aorta; 11 - arteriálny kmeň; 12 - vonkajšia krčná tepna, koreň brušnej vzostupnej aorty; 13 - vnútorná krčná tepna, dorzálny koreň aorty
Pravý štvrtý oblúk sa stáva proximálnou časťou pravej podkľúčovej tepny. Distálna časť tejto tepny vychádza z pravej dorzálnej aorty. Proximálna časť ľavého šiesteho oblúka sa mení na proximálnu časť ľavej pľúcnej tepny a distálna časť na arteriálny (botal) kanál. Proximálna časť pravého šiesteho oblúka tvorí proximálnu časť pravej pľúcnej tepny a distálna časť je redukovaná. Korene dorzálnej aorty veľmi skoro splývajú do nepárovej dorzálnej aorty, ale v 7. týždni vývoja sa distálna časť pravého koreňa dorzálnej aorty zvyčajne spája do pravej podkľúčovej tepny, ako už bolo uvedené vyššie. To je možné vďaka opačnému vývoju miesta spájajúceho pravý koreň s ľavým. Prezentácia materiálu týkajúceho sa vývoja srdca a veľkých tepien poskytuje základ pre porovnanie
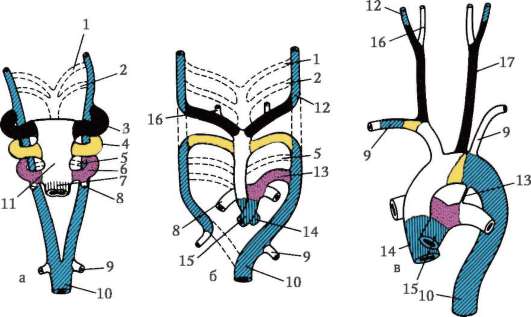
Ryža. 7.41. Tvorba najväčších tepien, deriváty arteriálneho kmeňa, aortálneho vaku, aortálnych oblúkov a dorzálnej aorty: a - 6-týždňové embryo; b - 7-týždňový plod; c - 6-mesačný plod: 1-6 - aortálne oblúky; 7 - arteriálny kmeň; 8 - pľúcna tepna; 9 - podkľúčová tepna; 10 - dorzálna zostupná aorta; 11 - aortálny vak; 12 - vnútorná krčná tepna; 13 - arteriálny (botalový) kanál; 14 - vzostupná aorta; 15 - pľúcny kmeň; 1b - vonkajšia krčná tepna; 17 - spoločná krčná tepna
ontogenézy človeka s fylogenetickým vývojom stavovcov (pozri kapitolu 14), a tiež nám umožňuje priblížiť sa k pochopeniu mechanizmov vzniku vrodených vývojových chýb srdca a ciev.
Otázky na sebaovládanie
1. Aké typy ontogenézy existujú? Vymenujte a opíšte hlavné štádiá ontogenézy.
2. Aké sú znaky stavby a funkcie zárodočných buniek?
3. Ako prebieha hnojenie a aké fázy tento proces zahŕňa?
4. Opíšte podstatu a morfologické formy štádií embryonálneho vývoja u predstaviteľov rôznych tried
strunatcov (drvenie, gastrulácia, organogenéza a tvorba profyzorických orgánov).
5. Aké sú znaky embryonálneho vývoja cicavcov?
6. Uveďte príklady embryonálneho vývoja ľudských orgánov, odrážajúce vývoj rodových skupín strunatcov.
Ontogenéza je individuálny vývoj organizmu, súbor postupných morfologických, fyziologických a biochemických premien, ktorým organizmus prechádza od oplodnenia (počas pohlavného rozmnožovania) alebo od okamihu odlúčenia od jedinca matky (počas asexuálna reprodukcia) až do konca života.
Prideľte preembryonálne, embryonálne a postembryonálne obdobia. U placentárnych zvierat a ľudí sa rozlišuje prenatálne (predpôrodné) a postnatálne (postnatálne) obdobie.
Súhrn udalostí vyskytujúcich sa v ovo(oo)genéze, ale v záujme procesu individuálneho vývoja nového organizmu, tvorí obsah obdobia progenézy - preembryonálne obdobie individuálneho rozvoja. Začína sa pred oplodnením a vytvorením zygoty a spája sa hlavne so ženskou zárodočnou bunkou. Ten v priebehu gametogenézy (ovogenézy) získava niektoré vlastnosti, ktoré nevyužije sám, ale jedinec novej generácie, ktorá začala individuálny vývoj. Jednou z týchto, azda najznámejšou, je tvorba väčšieho alebo menšieho množstva žĺtka v cytoplazme vajíčka v závislosti od druhu živočícha, ktorý sa používa ako živná látka pri vývoji potomstva. . S obdobím pred oplodnením časovo súvisí aj funkčná genetická aktivita množstva génov, ktorá sa prejavuje ich transkripciou a post(post)transkripčnými zmenami v primárnom a (m)RNA transkripte. i(m)RNA vytvorená ako výsledok tejto aktivity organizuje syntézu proteínov dôležitých pre skoré štádiá embryogenézy.
Druhy vajec.
Sú klasifikované podľa množstva a rozloženia žĺtka. Môže byť:
1. Alecitálne vajcia – žiadny alebo veľmi malý žĺtok
Príklad: u cicavcov
2. Isolecitál – žĺtka je málo, je rovnomerne rozmiestnený v cytoplazme
Príklad: lancelet
3. Polylecital: a) telolecital - žĺtok je posunutý smerom k vegetatívnemu pólu
b) centrolecitál - žĺtok v strede.
Hnojenie
Fúzia spermie s vajíčkom. Míľniky proces oplodnenia: 1) prienik spermie do vajíčka; 2) aktivácia rôznych syntetických procesov vo vajci; 3) fúzia jadier vajíčka a spermie s obnovením diploidnej sady chromozómov.
Životnosť a schopnosť oplodniť vajíčka a spermie sú obmedzené. U väčšiny cicavcov si vajíčko zachováva schopnosť oplodnenia 24 hodín a u ľudí 12-24 hodín po ovulácii. Spermie si zachovávajú svoju schopnosť oplodnenia v ženskom pohlavnom trakte aj niekoľko hodín.
Priamy kontakt spermie a vajíčka spôsobuje pravidelné zmeny v oboch bunkách, ktoré spôsobujú prienik jadra a centriolu mužskej gaméty do vajíčka a aktiváciu metabolizmu vajíčka. Tieto zmeny na strane spermií sa nazývajú akrozomálna reakcia a na strane vajíčka kortikálna reakcia. Podstata akrozomálnej reakcie je nasledovná. V momente kontaktu s vajíčkom v hornej časti hlavičky spermie sa plazmatická membrána a časť membrány akrozomálneho vezikula priľahlá k nej rozpustia. V dôsledku uvoľnenia enzýmov z akrozomálnej granule sa priľahlá časť vaječnej membrány rozpustí. Akrozomálna membrána vyčnieva smerom von a vytvára výrastok vo forme dutej trubice. Ten sa predlžuje, prechádza cez vaječné membrány a prichádza do kontaktu s plazmatickou membránou vajíčka. V oblasti takéhoto kontaktu sa objaví výčnelok alebo tuberkulum oplodnenia, po ktorom sa plazmatické membrány oboch gamét zlúčia a začne sa spojenie ich obsahu. Od tohto momentu sú spermie a vajíčko jedna bunka – zygota. Akrozomálna reakcia je extrémne rýchla.
Aktivácia vajíčka alebo kortikálna reakcia, ktorá sa vyvíja v dôsledku kontaktu so spermiou. Zmeny v povrchovej kortikálnej vrstve ooplazmy a tvorba oplodňovacej membrány. Táto blana, nazývaná aj žĺtková, vzniká odlupovaním z povrchu vajíčka. Pod ním sa vytvorí periyolkový priestor, do ktorého sa naleje obsah granúl kortikálnej vrstvy cytoplazmy vajíčka. Oplodňovacia membrána chráni vajíčko pred prienikom nadpočetných spermií, t.j. poskytuje "blok polyspermie". Tekutina, ktorá sa hromadí v periyolku, slúži ako špecifické prostredie, v ktorom vývoj embrya prebieha až do okamihu, keď opustí vaječné membrány.
IN embryonálny V tomto období existujú tri hlavné štádiá: drvenie, gastrulácia a primárna organogenéza. Embryonálne alebo zárodočné obdobie ontogenézy začína okamihom oplodnenia a pokračuje, kým sa embryo nevynorí z vaječných membrán. U väčšiny stavovcov zahŕňa štádiá zygoty, štiepenia, gastrulácie, histo- a organogenézy.
Zygota je jednobunkové embryo. V zygote dochádza k procesu diferenciácie cytoplazmy. Pozoruje sa pohyb ooplazmy. Objavujú sa v nej oblasti, z ktorých sa následne vytvárajú predpokladané (domnelé) rudimenty budúcich orgánov.
Štiepenie - séria postupných mitotických delení oplodneného alebo iniciovaného vajíčka na vývoj. Štiepenie je prvé obdobie embryonálneho vývoja, ktoré je prítomné v ontogenéze všetkých mnohobunkových živočíchov a vedie k vytvoreniu embrya nazývaného blastula (jednovrstvové embryo). Hmotnosť embrya a jeho objem sa zároveň nemení, to znamená, že zostávajú rovnaké ako u zygoty a vajíčko sa delí na stále menšie bunky - blastoméry. Po každom delení štiepenia sa bunky embrya zmenšujú a zmenšujú, to znamená, že vzťahy medzi jadrom a plazmou sa menia: jadro zostáva rovnaké a objem cytoplazmy sa zmenšuje. Proces pokračuje, kým tieto ukazovatele nedosiahnu hodnoty charakteristické pre somatické bunky. Typ drvenia závisí od množstva žĺtka a jeho umiestnenia vo vajci. Ak je žĺtka málo a je rovnomerne rozmiestnený v cytoplazme (izolecitálne vajíčka: ostnatokožce, ploskokožce, cicavce), potom sa drvenie postupuje podľa typu úplnej uniformy: blastoméry sú rovnako veľké, rozdrví sa celé vajíčko. Ak je žĺtok rozmiestnený nerovnomerne (telelektálne vajcia: obojživelníky), tak drvenie prebieha podľa typu úplne nerovnomerného: blastoméry sú rôznej veľkosti, tie, ktoré obsahujú žĺtok, sú väčšie, vajce sa drví ako celok. Pri neúplnom rozdrvení je vo vajciach toľko žĺtka, že ho drviace ryhy nedokážu úplne oddeliť. Štiepenie vajíčka, pri ktorom sa rozdrví iba „čiapka“ cytoplazmy sústredená na zvieracom póle, kde sa nachádza jadro zygoty, sa nazýva neúplné diskoidálne (telolecitálne vajíčka: plazy, vtáky). Pri neúplnom rozdrvení povrchu v hlbinách žĺtka dochádza k prvým synchrónnym jadrovým deleniam, ktoré nie sú sprevádzané tvorbou medzibunkových hraníc. Jadrá, obklopené malým množstvom cytoplazmy, sú v žĺtku rovnomerne rozložené. Keď ich je dostatok, migrujú do cytoplazmy, kde potom po vytvorení medzibunkových hraníc vzniká blastoderm (centrolecitálne vajíčka: hmyz).
· Gastrulácia – proces bunkového pohybu, sprevádzaný rastom, rozmnožovaním, diferenciáciou. Embryo v tomto štádiu sa nazýva gastrula.
Spočiatku sa vytvárajú vonkajšie (ektodermické) a vnútorné (endodermické) listy. Neskôr sa objaví tretia, stredná zárodočná vrstva (mezoderm), ktorá sa nachádza v tele embrya medzi ektodermou a endodermou.
Existujú 4 hlavné spôsoby formovania vonkajších a vnútorných listov. Často však dochádza ku kombinácii viacerých spôsobov gastrulácie.
1 spôsob invaginácie, ktorá spočíva v tom, že určitý úsek blastodermu sa pri zachovaní štruktúry vrstvy zaskrutkuje do blastocoelu. Potom blastocoel zmizne a objaví sa gastrocoel. Otvor, cez ktorý táto dutina komunikuje s vonkajším prostredím, sa nazýva primárne ústa alebo blastopór.
2-cestná epibola– zanášanie makromérov rýchlo sa deliacimi mikromérmi zvieracieho pólu. V embryách takýchto zvierat sa spočiatku netvorí blastopór a chýba gastrocoel.
3-cestná imigrácia– výpočet časti buniek steny blastuly vo vnútri blastocoelu.
4-cestná delaminácia - bunky vonku sa premenia na vrstvu
a-ektoderm; b-endoderm; v-blastocoel.
Pokladanie tretej zárodočnej vrstvy prebieha dvoma spôsobmi: teloblastické a enterocoelózne.
Teloblastická - metóda tvorby mezodermu v protostómoch oddelením dvoch mezodermálnych pásikov od dvoch primárnych embryonálnych buniek (teloblastov).
Enterocoelous - spočíva v tom, že z endodermu primárneho čreva sa na oboch stranách vytvárajú výbežky - vrecká (coelomické vaky), ktoré sa neskôr šnurujú a rastú medzi ekto - a endodermom, čím sa vytvára mezoderm; ich dutiny navzájom splývajú a vzniká sekundárna telová dutina, čiže coelom.
Histo- a organogenéza
Možno rozlíšiť dve fázy.
1 neurulácia, spočíva vo vytvorení komplexu axiálnych orgánov - nervovej trubice, chordy. Embryo v štádiu neurulácie sa nazýva neurula. Najprv dochádza k splošteniu bunkovej vrstvy, čo vedie k vytvoreniu nervovej platničky, ktorej okraje, stúpajúc, tvoria nervové záhyby. V dôsledku pohybu buniek pozdĺž strednej čiary nervovej platničky dochádza k depresii. V tomto štádiu neurulácie je záložka nervový systém nazývaná nervová drážka. Súčasne sa zdá, že nervová platňa sa prehýba pozdĺž strednej čiary a o niečo neskôr sa jej okraje zatvoria. V dôsledku týchto procesov sa objavuje nervová trubica s dutinou - neurocoel. k uzáveru hrebeňov dochádza najskôr v strede a potom v zadnej časti neurálnej drážky. To sa deje ako posledné v časti hlavy, ktorá je širšia ako ostatné oblasti. Predný, rozšírený úsek v ďalšom vývoji tvorí mozog, zvyšok nervovej trubice - miecha.
2 histo- a organogenéza.
Z ektodermy sa tvorí kožná epidermis a jej deriváty, žľazy, epitel ústnej dutiny, vaginálny epitel, zubná sklovina a receptorové nervové bunky.
Z endodermu - epitelu dýchacích ciest a pľúc vzniká časť buniek pankreasu, secernujúce bunky, epitel žalúdka, čreva.
Z mezodermu - kostra chrupaviek a kostí, svaly, obličky, cievy.