Стебель — это осевая структура побега. И его анатомическое строение вытекает из главных функций. Во-первых, для стебля характерно развитие механической и проводящей тканей. Во-вторых, стебель имеет сложную систему меристем: верхушечные, боковые и вставочные, которые определяют его нарастание в течение длительного времени, а также возникновение новых органов. Возникает стебель из апикальной меристемы, из которой дифференцируются три слоя тканей: основная, покровная, проводящая.
Строение однодольных растений
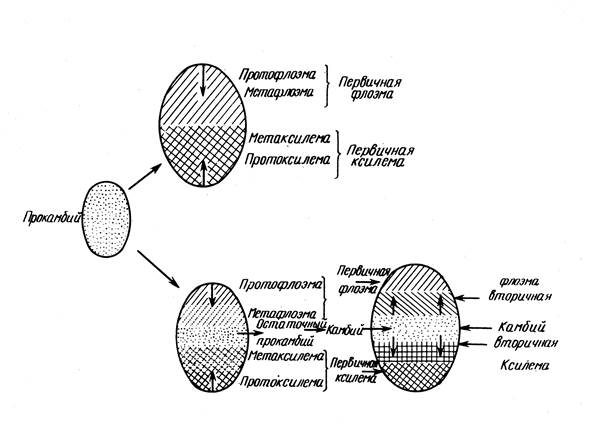
Весь прокамбий или первичная меристема у однодольных растений дифференцируется в элементы первичных проводящих тканей. Их стебли, особенно, если рассматривать травянистые (злаки), имеют более простое строение по сравнению со стеблями двудольных растений. Для них также в основном характерно первичное строение. У однодольных растений сосудисто-волокнистые проводящие пучки закрытые (без камбия), состоят только из первичных тканей ирасположены беспорядочно в основной паренхиме стебля.

Строение двудольных растений
В средней части прокамбиального тяжа у двудольных растений происходит образование камбия и начинается формирование вторичных проводящих тканей (метафлоэмы и метаксилемы). Объём этих вторичных проводящих тканей увеличивается за счёт деления клеток камбия, что в итоге приводит к сильному утолщению стебля.
Для двудольных характерны открытые пучки с камбием. Проводящие ткани в стеблях двудольных растений располагаются кольцом вокруг сердцевины. Пучки разделяются сердцевинными лучами, которые состоят из паренхимы и соединяют сердцевину или с перициклом, или с первичной корой. По периферии в стеблях двудольных растений располагаются механические ткани , при этом склеренхима входит в состав перицикла, колленхима , вместе с основной паренхимой, входит в состав первичной коры.
Строение центрального цилиндра двудольных может быть пучковым и непучковым .
По своей анатомической структуре однолетние стебли древесных растений сходны со стеблями травянистых растений: их проводящие системы имеют непучковое строение . Их отличительной чертой является активная деятельность камбия и раннее формирование вторичной покровной ткани - пробки .
Благодаря деятельности камбия в стебле образуются различные элементы вторичного происхождения, а первичные элементы при этом постепенно исчезают. Камбий формирует элементы ксилемы и флоэмы с различной скоростью: на одну клетку флоэмы камбий отделяет несколько клеток ксилемы. В результате этого ксилема (или древесина) нарастает намного быстрее, чем флоэма (луб), соответственно, почти вся масса ствола и ветвей дерева приходится на долю ксилемы. Флоэма, напротив, составляет сравнительно тонкий слой вторичной коры.

Сплошное камбиальное кольцо у большинства древесных растений (в качестве примера можно взять липу) образуется в самом начале формирования стебля.
Деление клеток камбия происходит тангентально, что определяет расположение клеток правильными рядами по радиусу. При нарастании ксилемы камбий сдвигается ближе к периферии и его окружность увеличивается за счёт радиального деления его клеток.
При вторичном утолщении вторичную ксилему или древесину с паренхимными лучами составляет все, что откладывается вовнутрь ствола, а все то, что откладывается наружу, т.е. к периферии ствола, составляет вторичную флоэму или луб с сердцевинными лучами.

Проводящая система ксилемы
состоит из трахеид и сосудов. У хвойных растений только трахеиды выполняют функцию проведения, а у древесных лиственных растений — и трахеиды, и сосуды.
Появление годичных колец
в ксилеме (древесине) происходит в результате периодической деятельности камбия. В весенний период, как правило, много воды и питательных веществ, поэтому камбий образует крупные элементы древесины
с большим просветом и тонкими стенками. Ближе к концу вегетационного периода происходит затухание деятельности камбия и в древесине начинают преобладать механические элементы
и узкие сосуды.
Сердцевинные лучи состоят из крупных паренхимных клеток, которые имеют прямоугольную форму. По своему происхождению эти лучи могут быть первичными и вторичными. Первичные лучи тянутся от первичной коры до сердцевины, и они длиннее вторичных. Основная функция лучей — проведения воды и органических веществ в горизонтальном направлении.
Механическая ткань в ксилеме (древесине) состоит из толстостенных узких и уже одревесневших клеток.
В состав вторичной флоэмы тоже входят 3 типа тканей: основная, механическая и проводящая. Флоэмные сердцевинные лучи проходят от камбия наружу, и стенки их клеток не одревесневают. В своей периферийной части лучи сильно расширяются. Механическую ткань вторичной флоэмы называют вторичной склеренхимой, она представлена лубяными волокнами (твёрдый луб ). Проводящая ткань представляет собой ситовидные трубки с клетками-спутницами (мягкий луб ).

На ранних этапах развития стебель двудольных имеет первичное строение. В результате деятельности первичных меристем конуса нарастания формируются эпидерма, первичная кора, центральный цилиндр и сердцевина.
Эпидерма стебля имеет небольшое число устьиц, обычно покрыта кутикулой.
Под эпидермой находятся ткани первичной коры. Наружный слой ее часто образован колленхимой, которая располагается либо сплошным кольцом (подсолнечник), либо отдельными участками. Основная часть первичной коры сложена хлоренхимой и паренхимой. Здесь могут развиваться воздухоносные полости и вместилища выделений. Клетки внутреннего слоя первичной коры обычно заполнены крупными крахмальными зернами (крахмалоносное влагалище).
Центральный цилиндр состоит из перицикла и проводящей системы. Сердцевина - из паренхимы, которая часто разрушается, а на ее месте образуется воздухоносная полость. Перицикл представлен одним или несколькими рядами склеренхимы или отдельными ее тяжами. Проводящая система имеет вид полого цилиндра, который разделяет первичную кору и сердцевину. Проводящие пучки, составляющие этот цилиндр, разделены прослойками межпучковой паренхимы – сердцевинными лучами, которые связывают сердцевину и первичную кору. Большинству двудольных свойственен особый тип центрального цилиндра, на поперечном срезе которого пучки расположены кольцом (рис.) .
Рисунок. Стебель двудольных растений.
У двудольных растений слой клеток прокамбия между первичными флоэмой и ксилемой сохраняет способность к делению и превращается во вторичную образовательную ткань - камбий , который делится, откладывая внутрь элементы вторичной ксилемы, а к периферии - вторичной флоэмы. Пучки двудольных растений открытые, благодаря работе камбия пучок растет, диаметр его увеличивается. Деятельность камбия обеспечивает возникновение вторичных элементов в стебле, т. е. переход от первичного к вторичному анатомическому строению.
Вторичное строение характерно для всех двудольных и хвойных растений - трав и деревьев. Разнообразие типов строения обусловлено, прежде всего, расположением проводящих тканей, которое определяется заложением прокамбия и деятельностью камбия.
Прокамбий в процессе дифференциации конуса нарастания закладывается в виде тяжей. Если тяжи разделены достаточно широкими рядами паренхимы, то формируется пучковое или переходное строение стебля ; если тяжи прокамбия сближены настолько, что сливаются в цилиндр, то формируется непучковое (сплошное) строение .
Пучковое строение стебля встречается у некоторых травянистых растений (укроп, клевер, лютик, горох, люцерна). Заложенные в конусе нарастания прокамбиальные тяжи располагаются в один круг по периферии центрального цилиндра . Каждый прокамбиальный тяж превращается в открытый коллатеральный пучок, состоящий из первичной ксилемы, первичной флоэмы и полоски камбия между ними. Клетки камбия, делясь, дают новые (вторичные) элементы проводящего пучка: внутрь - ксилему к периферии - флоэму. Проводящие пучки разделены широкими межпучковыми зонами и окружают мощную сердцевину.
У большинства растений камбий работает активно и диаметр пучков значительно увеличивается Деятельность пучкового камбия стимулирует паренхиму, разделяющую пучки, которая начинает делиться, давая вторичную образовательную ткань. Пучковая и межпучковая меристемы смыкаются и образуют сплошное камбиальное кольцо , деятельность которого обеспечивает равномерное утолщение стебля. Если пучковый камбий дает элементы вторичных ксилемы и флоэмы, то межпучковый камбий производит паренхиму (у укропа, звездчатки злаковидной и др.). У клевера, люцерны и других растений межпучковый камбий откладывает преимущественно склеренхимоподобные клетки в сторону ксилемы. Эти клетки удлиненные, толстостенные одревесневающие. Поэтому задержка с уборкой клевера снижает качество сена. Отчетливое пучковое строение сохраняется у этих растений в течение всей жизни стебля.
Пучковое строение стебля может с возрастом смениться непучковым сплошным. Переходное строение стебля характерно для подсолнечника . Вначале стебель формируется так же, как у клевера, но межпучковый камбий откладывает не паренхиму, а ксилему и флоэму. Формируются добавочные проводящие пучки. Постепенно все пучки могут слиться в сплошной трехслойный (ксилема, камбий, флоэма) цилиндр. В верхней (молодой) части стебель имеет пучковое строение, у основания (в старой части) - непучковое.
Непучковое строение свойственно стеблям многих трав и деревьев. В конусе нарастания этих растений прокамбиальные тяжи настолько сближены, что образуют почти сплошной цилиндр (на поперечном срезе он имеет вид кольца). Прокамбиальный цилиндр (кольцо), дифференцируясь, дает к центру цилиндр первичной ксилемы, к периферии - цилиндр первичной флоэмы, а между ними образуется камбиальный цилиндр.
Вторичные изменения связаны с работой камбия. Клетки камбия делятся параллельно поверхности стебля, при этом внутрь откладывается в 10...20 раз больше клеток, чем наружу. Внутрь камбий формирует элементы вторичной ксилемы (древесины), наружу откладывается вторичная флоэма (луб). Местами камбий откладывает в обе стороны паренхимные клетки сердцевинных лучей.
Особенности непучкового строения стеблей трав можно рассмотреть на примере стебля льна . Стебель покрыт эпидермой, под которой располагается рыхлая хлоренхима первичной коры. Большое количество устьиц в эпидерме и отсутствие колленхимы в первичной коре имеют существенное практическое значение, так как во время технической мочки льна вода не встречает препятствий и легко проникает внутрь стебля. Клетки крахмалоносного влагалища (эндодермы) более крупные, содержат хорошо заметные крахмальные зерна.
Примыкающий к эндодерме центральный цилиндр начинается группами склеренхимных клеток – лубяных волокон, между которыми расположены клетки паренхимы. Лубяные волокна возникают в перицикле и в первичной флоэме. Они отличаются очень толстыми, обычно неодревесневшими стенками, состоящими на 75...90 % из целлюлозы. Созревшая клетка лубяного волокна мертвая. Протопласт после утолщения клеточной стенки разрушается. Лубяные волокна обладают необыкновенно высокой прочностью. Сопротивление разрыву у них соответствует сопротивлению стали, а упругость выше. Клетки лубяного волокна очень тесно соединены между собой. Льняное волокно - великолепное прядильное сырье, используемое в текстильной промышленности.
Далее в последовательном порядке располагаются цилиндры (кольца): вторичной флоэмы, камбия, вторичной ксилемы с хорошо заметными сердцевинными лучами. Первичная ксилема расположена участками, примыкающими к сердцевине. Паренхима сердцевины быстро разрушается (в процессе удлинения стебля), образуя центральную полость.
Стебли двудольных травянистых растений характеризуются:
1. вторичным строением, очень рано возникающим вслед за первичным;
2. наличием первичной покровной ткани - эпидермы;
3. развитой первичной корой, делящейся на колленхиму (экзодерма), паренхиму (мезодерма) и крахмалоносное влагалище (эндодерма);
4. пучковым или непучковым (сплошным) строением;
5. правильным расположением коллатеральных или биколлатеральных проводящих пучков в один круг по периферии центрального цилиндра;
6. наличием камбия (пучки открытые).
Метаморфозы побега.
Если дополнительные функции начинают преобладать над главными, или орган в процессе развития приобретает какие-то новые функции, он очень сильно изменяется и морфологически, и анатомически. Такие видоизменения органов называют метаморфозами (от греч. metamorphosis - превращение, преобразование). Органы, имеющие одинаковое происхождение, но выполняющие разные функции называют гомологичными . Гомологичными органами являются корневища, клубни, луковицы, колючки боярышника, кладодии, филлокладии, усики винограда и тыквы, т.к. все они исторически развились из листостебельного побега.
Органы, выполняющие одинаковые функции и сходные морфологически, но имеющие разное происхождение, называют аналогичными . Аналогичны корневые шишки и побеговые клубни, побеговые колючки боярышника и листовые колючки барбариса, листовые усики гороха и побеговые усики тыквы.
Видоизменения подземных побегов.
В природе широко распространены подземные побеги: корневища, клубни, клубнелуковицы, луковицы. Они участвуют в запасе веществ и вегетативном размножении растений.
Корневище представляет собой побег с чешуевидными листьями, почками и придаточными корнями. На поверхности корневища хорошо заметны рубцы от прошлогодних низовых листьев и отмерших побегов. Корневище имеет много придаточных корней. Обычно корневища подземные. Почки, развивающиеся в надземные побеги. Корневища характерны для пырея, ириса, кувшинки.
Столон - удлиненный тонкий побег с недоразвитыми листьями. В отличие от корневища он обычно недолговечен. У мускусницы, кислицы столоны участвуют в накоплении питательных веществ, откладывающихся в сильно утолщающихся низовых листьях, но основная функция столона - участие в вегетативном размножении.
Клубень в отличие от корневища и столона сильно укорочен и утолщен. Запасные вещества в нем локализуются в стеблевой паренхиме.
Клубни могут развиваться на корневищах, столонах, главном побеге и других частях растений. Они могут быть подземными и надземными.
Мощные корневищные клубни с большим количеством крахмала характерны для таро, или колоказии съедобной из семейства ароидных, которую культивируют в тропических и субтропических странах.
У картофеля клубни формируются на конце подземных столонах - беловатых хрупких побегах с чешуевидными листьями, развивающихся из пазушных почек, находящихся в основании олиственных побегов. На поверхности клубня располагаются пазушные почки - глазки , сидящие в небольших ямках, обрамленных с одной стороны бровками - остатками оснований листьев.
Клубни топинамбура , или земляной груши , тоже образуются на столонах. Проводящие ткани в них слабо развиты. Клубни богаты инулином, который откладывается преимущественно в стеблевой паренхиме.
Клубни, формирующиеся на столонах и корневищах, недолговечные. Они сохраняются только один год, между двумя вегетационными периодами, отмирая после развития из их почек олиственных побегов, потребляющих питательные вещества до тех пор, пока не образуются корни, и растения не перейдут к самостоятельному питанию.
Луковица , как и клубень, представляет собой специализированный, видоизмененный укороченный побег, служащий не только для хранения питательных веществ, но и для перенесения неблагоприятных периодов года, вегетативного возобновления и размножения растений. Луковицы свойственны однодольным растениям из семейств лилейных, амариллисовых и других. Как исключение, они встречаются у двудольных: некоторых видов кислицы и жирянки.
Стебель, называемый донцем, в луковице сильно редуцирован и более или менее уплощен. Он несет листья в виде чешуй, а в нижней части - придаточные корни. В отличие от корневища и клубня запасающими органами луковицы служат мясистые чешуи. Луковицы образуются у тюльпана, рябчика нарцисса, гиацинта, пролески, подснежника и других растений.
В зависимости от развития боковых почек луковицы бывают простыми , когда почки невелики (лук, лилия и др.), и сложными – почки в пазухах сухих чешуй крупные, сильно развитые (чеснок).
Клубнелуковица - подземное видоизменение побега, сочетающее признаки корневища и луковицы. Она развивается из побега с сильно укороченным стеблем, от нижней части которого отходят придаточные корни. На верхушке клубнелуковицы или в ее основании находится почка, из которой образуется цветоносный побег. Стебель клубнелуковицы покрыт основаниями отмерших прошлогодних листьев, имеющих вид сухих пленчатых чешуй. Из почек, находящихся в их пазухах, развиваются новые клубнелуковицы - детки. Клубнелуковицы имеют шафран, гладиолус, безвременник.
Видоизменения надземных побегов
Кочан капусты представляет собой, по существу, почку гигантских размеров: среднюю часть кочана занимает мясистый стебель (кочерыга ), вдоль которого расположены сочные мясистые листья. Наиболее мелкие и молодые из них находятся внутри кочана, вокруг конуса нарастания, которым заканчивается кочерыга. У брюссельской капусты образуются мелкие кочаны из боковых почек.
В кочане оба органа побега участвуют в накоплении питательных веществ: водорастворимых сахаров и других биологически ценных соединений, в том числе витаминов, но основная роль принадлежит листьям.
Надземные клубни , развивающиеся из нижней части главного побега, характерны для капусты кольраби.
Мелкие клубеньки могут развиваться и в области соцветия. У горца живородящего, например, они образуются в пазухах нижних кроющих листьев соцветия. Их почки начинают прорастать уже на материнском растении. После опадения они укореняются, образуя новые растения.
Метаморфизированные побеги могут служить не только для хранения питательных веществ, воспроизведения и размножения растений, но выполнять и другие функции.
У некоторых растений побеги в процессе развития теряют листья, и фо-тосинтезирующим органом становится стебель, называемый в этом случае кладодием . Обычно он уплощен и обладает способностью к длительному росту (у тропического растения мюленбекии, или гомалокладуса, из семейства гречишных, опунция). В узлах хорошо выражены перетяжки, поэтому кладодий выглядит членистым. Во влажных условиях кладодии могут быть олиственными, но в сухую погоду листья опадают. Как орган фотосинтеза кладодий имеет хорошо развитую хлорофиллоносную ткань, расположенную под эпидермой.
Кладодии, по внешнему виду сходные с листьями, называют филлокладиями . Они развиваются из боковых почек, поэтому всегда находятся в пазухе небольшого пленчатого или чешуевидного листа. Филлокладии очень разнообразны. У спаржи перистой, или щетинистой, они мелкие, игольчатые, у спаржи Шпренгера, или густоцветковой, - уплощенные, линейные. Широколанцетные или яйцевидные в очертаниях кожистые филлокладии характерны для видов иглицы, встречающихся в Средиземноморье, Закавказье, Крыму. Летом на нижней стороне филлокладия, в его средней части, развивается соцветие, расположенное в пазухе мелкого кроющего листа, осенью и зимой иглица цветет, а весной на филлокладиях созревают оранжевые ягоды.
Довольно распространенный тип изменения побегов - колючка . Это сильно одревесневающий безлистный укороченный побег с острой верхушкой. У боярышника побег, развивающийся весной в пазухе листа, сначала имеет мелкие листья и верхушечную почку. Вскоре верхушечный рост прекращается, листья опадают, стебель сильно одревесневает и превращается в колючку. Так же развиваются колючки у терна.
Цепляющиеся растения снабжены усиками , которые, как и колючки, представляют собой у некоторых растений видоизмененные боковые побеги. У пассифлоры усики простые, неветвистые, с закрученной верхушкой. Они находятся в пазухах листьев, как и цветки. Ветвистые усики винограда представляют собой видоизмененные соцветия. У девичьего винограда окончания разветвленных усиков дисковидно расширены и играют роль присосок, помогающих растению взбираться на опору. Усики побегового происхождения образуются также у тыквы, арбуза, огурца и других представителей семейства тыквенных.
Первичное анатомическое строение стебля.
Определение и функции стебля.
Стебель – осевой, обычно радиально симметричный орган, обладающий длительным верхушечным (апикальным) ростом.
Функции стебля:
1. осуществляет двустороннее передвижение веществ между корнями и листьями,
2. поддерживает крону растения,
3. способствует увеличению общей ассимиляционной поверхности растения благодаря ветвлению,
4. участвует в хранении запасных веществ,
5. в молодом возрасте – осуществляет также фотосинтез.
Эти функции определяют наличие в стебле покровных, хорошо развитых проводящих, механических тканей и функционально развитой паренхимы.
Строение конуса нарастания.
В конусе нарастания стебля покрытосеменных растений выражены две зоны: наружная – туника , и внутренняя – корпус, которые различаются плоскостями клеточных делений (теория А. Шмидта и Дж. Будера) (рис. 1).
Туника может состоять из одного или нескольких слоев клеток, делящихся перпендикулярно поверхности, у большинства двудольных растений она двухслойная. Наружный слой туники дает начало протодерме, из которой затем образуется эпидерма, покрывающая листья и стебель. Если туника многослойная, то внутренний слой образует ткани первичной коры.
При наличии однослойной туники всю первичную кору и стелу образует корпус. Корпус образован массой клеток, которые делятся во всех плоскостях .
Зачатки листьев чаще всего возникают во втором слое туники, а пазушные почки закладываются в корпусе.
Рисунок 1. б) апекс покрытосеменных растений.
В результате деятельности первичных меристем апекса формируется первичное анатомическое строение стебля: эпидерма, первичная кора, центральный цилиндр и сердцевина.
Вторичное утолщение происходит за счет деятельности камбия.
Первичное анатомическое строение стебля.
Строение стебля, сформировавшегося в результате деятельности первичных меристем апекса, называется первичным . В стебле, при первичном строении можно различить три анатомо-топографические зоны:
1. покровную ткань,
2. первичную кору
3. центральный цилиндр.
Эпидерма, защищает стебель от испарения.
Под эпидермой находится наружный слой первичной коры - экзодерма - представленная хлоренхимой и (у двудольных) колленхимой. Во внутреннем, пограничном с центральным цилиндром слое - эндодерме - обычно накапливаются крахмальные зерна, и тогда он превращается в так называемое крахмалоносное влагалище (крахмалоносный слой). Крахмальные зерна под действием силы тяжести могут оседать в клетках. Благодаря этому эндодерма играет важную роль в геотропической ((от греч. trpos - поворот, направление), ростовые движения стебля растений обусловленные направленным действием силы земного тяготения) реакции стеблей
По периферии центрального цилиндра в перицикле расположена склеренхима. Склеренхимные волокна образуют сплошной слой или собраны около проводящих пучков, кнаружи от первичной флоэмы. Главный элемент центрального цилиндра (стелы) - проводящие пучки. Они образованы из прокамбиальных тяжей и состоят из первичных ксилемы и флоэмы . Дифференциация прокамбия проходит центростремительно: флоэма формируется от внешней стороны пучка к его центру, а ксилема - навстречу ей (рис. 2). Прокамбий может полностью превратиться в первичные проводящие комплексы, образовав закрытый коллатеральный пучок (у однодольных), или сохранившиеся клетки в центре пучка формируют камбий. Такие открытые пучки характерны для двудольных растений. Работа камбия обеспечивает формирование вторичных ксилемы и флоэмы.
Рисунок 2.
Заложение прокамбия предопределяет строение проводящей системы:
1. Если прокамбий образует сплошной полый цилиндр, то из него формируются вложенные друг в друга цилиндры ксилемы и флоэмы (у некоторых хвойных и двудольных) – непучковое строение .
2. Если же прокамбий состоит из отдельных тяжей (у большинства однодольных, многих двудольных и хвойных), то возникающие первичные проводящие ткани сохраняют такое же пучковое строение .
Прокамбий возникает у основания листового зачатка, и его развитие идет в двух направлениях: к верхушке листового зачатка и вниз по стеблю, где он соединяется с тяжами, возникшим ранее. В той же последовательности идет и дифференциация флоэмы и ксилемы. Обычно раньше образуются элементы флоэмы, затем ксилемы. Проводящие пучки располагаются в паренхиме центрального цилиндра.
В центре стебля развивается хорошо заметная сердцевина. Паренхимные тонкостенные клетки сердцевины нередко разрыхляются, отмирают и заполняются воздухом, тогда вся ткань кажется белой (у подсолнечника, кукурузы, бузины). Иногда сердцевина отмирает очень рано, когда удлинение междоузлий еще не закончилось. Это влечет за собой разрыв сердцевины и образование полого стебля.
Первичное строение стебля сохраняется у однодольных растений в течение всей жизни, а у двудольных и голосеменных - лишь в ранние фазы его развития.
Строение стебля.
Стебель представляет собой ось побега, несущая листья и почки. Основные функции стебля - опорная и проводящая. Стебель осуществляет связь между корнями и листьями. Кроме того, в стебле нередко откладываются запасные питательные вещества. Иногда стебель - ассимилирующий орган.
В результате деятельности прокамбия и остальной первичной меристемы конуса нарастания образуется первичное строение стебля. В первичном стебле обычно различают первичную кору и стелу (центральный цилиндр). В отличие от корня первичная кора снаружи покрыта эпидермой.
У голосеменных и большинства двудольных покрытосеменных рост стебля в толщину осуществляет камбий, образующий вторичные ткани. Он возникает в виде цилиндра между первичной ксилемой и первичной флоэмой и остается в относительно том же положении неопределенно долго, откладывая по направлению к центру оси вторичную ксилему, а кнаружи - вторичную флоэму.
Вторичное утолщение происходит также в результате деятельности феллогена (пробкового камбия).
У древесных и кустарниковых двудольных, а также у хвойных вторичные утолщения могут продолжаться многие годы. В итоге в стебле выделяют три основные части: кору, древесину и сердцевину.
Отличия строения стебля однодольных и двудольных растений.
У однодольных стебель травянистый, не способен к вторичному утолщению, ветвится редко. Проводящие пучки без камбия, разбросаны по всему стеблю.
У двудольных стебель травянистый или одревесневший, способен к вторичному утолщению, ветвится. Проводящие пучки, имеющие камбий, расположены одним большим массивом в центре стебля или имеют вид кольца.
15 Функции цветка.
Цветок - это видоизмененный укороченный побег, приспособленный для размножения покрытосеменных (цветковых) растений.
Исключительная роль цветка связана с тем, что в нем совмещены все процессы бесполого и полового размножения, в то время как у низших и многих высших растений они разобщены. В обоеполом цветке осуществляются микро- и мегаспорогенез, микро- и мегагаметогенез, опыление, оплодотворение, образование семян и плодов. Особенности строения цветка позволяют осуществлять перечисленные функции с минимальными затратами пластических веществ и энергии.
Строение цветка
Цветок состоит из стеблевой части (цветоножка и цветоложе), листовой части (чашелистики, лепестки) и генеративной части (тычинки, пестик или пестики). Цветок прикрепляется к стеблю посредством цветоножки. Если цветоножка сильно укорочена или отсутствует, цветок называется сидячим (подорожник, вербена, клевер). На цветоножке располагаются также два (у двудольных) и один (у однодольных) маленьких предлиста - прицветника, которые часто могут отсутствовать. Верхняя расширенная часть цветоножки называется цветоложем, на котором располагаются все органы цветка. Цветоложе может иметь различные размеры и форму - плоскую (пион), выпуклую (земляника, малина), вогнутую (миндаль), удлинённую (магнолия). У некоторых растений в результате срастания цветоложа, нижних частей покрова и андроцея образуется особая структура - гипантий. Форма гипантия может быть разнообразной и иногда участвовать в образовании плода (цинарродий - плод шиповника, яблоко). Гипантий характерен для представителей семейств розовых, крыжовниковых, камнеломковых, бобовых.
Части цветка делят на фертильные, или репродуктивные (тычинки, пестик или пестики), и стерильные (околоцветник).
функции цветка
Именно благодаря цветкам растения живут и размножаются. Красивая форма и окраска цветка, прежде всего, выполняет важную для их размножения функцию - привлекают насекомых или птиц-опылетелей. После опыления в завязи цветка образуются семена, которые дадут жизнь новым растениям. Не все растения образуют цветки и далеко не все цветки могут похвастаться привлекательными формами и расцветкой. Однако те из них, которые отличаются ярким видом, привлекают потенциальных опылителей. Нередко конкретное растение опыляется насекомыми определенного вида, у которого есть органы, необходимые для их опыления. Самые известные насекомые-опылители - это пчелы и мухи. Некоторые цветки опыляют птицы, например, колибри. У ветроопыляемых растений цветки мелкие и невзрачные на вид, поскольку их форма и расцветка не могут оказать никакого влияния на опылительный процесс. Некоторые цветки имеют, по мнению человека, неприятный запах, например, стапелия, запах которой напоминает запах разлагающегося мяса.
16 вопрос)))
Ветроопыляемые растения имеют, как правило, ряд характерных особенностей: очень мелкие и многочисленные цветки, вырабатывают много пыльцы. Одно растение способно вырабатывать миллионы пыльцевых зерен. У многих ветроопыляемых растений (лещина, осина, ольха, шелковица) цветки появляются ещё до распускания листьев.
Пчёлы предпочитают собирать пыльцу с насекомоопыляемых растений. Но если в природе мало цветущих энтомофильных растений, а пчёлы нуждаются в пыльце, они собирают её и с ветроопыляемых растений.
Опыление цветковых растений осуществляется двумя основными способами - ветром и животными, чаще всего насекомыми. Как в том, так и в другом случае у растений вырабатываются специфические приспособления. Для насекомоопыляемых растений характерны крупные, ярко окрашенные одиночные цветки, а также соцветия, состоящие из ярких цветков различной формы. Как правило, они имеют сильный запах. В них развиты особые железы - нектарники, вырабатывающие сладкий жидкий секрет - нектар. Цветки насекомоопыляемых растений богаты пыльцой. Пыльцевые зёрна, как правило крупные и клейкие, а их оболочка нередко имеет разнообразные выросты. У ветроопыляемых растений околоцветник частично или полностью редуцируется, а их мелкие и невзрачные цветки, как правило, собраны в соцветия. пыльцы у ветроопыляемых растений, по сравнению с насекомоопыляемыми, образуется значительно больше. Однако пыльцевые зёрна у них мелкие и сухие, хорошо переносимые ветром. Они образуются в крупных пыльниках, очень часто свисающих на длинных тычиночных нитях. Рыльце пестика у многих из них раздвоено и усажено многочисленными волосками и щетинками, что позволяет улавливать из воздуха значительно больше пыльцы. Очень многие ветроопыляемые растения цветут ранней весной ещё до распускания листьев. Цветущие летом ветроопыляемые злаки распускаются строго по часам, экономя тем самым пыльцу. Повышению вероятности опыления способствует и то, что все ветроопыляемые растения произрастают тесными группами или большими скоплениями.
17. Плоды и семена. Классификация, способы распространения
Плод . Это один из самых характерных органов покрытосеменных растений. Он состоит из околоплодника и семян. Околоплодник, представляющий собой разросшуюся и сильно видоизмененную стенку завязи, обеспечивает формирование семян, защиту от неблагоприятных факторов, способствует их распространению.
Сочные плоды имеют хорошо развитую мякоть, в крупных паренхимных клетках которой накапливается много воды, углеводов, витаминов, органических кислот, различных ароматических соединений.
У сухих плодов околоплодник кожистый или деревянистый. Они могут быть односемянными и многосемянными, вскрывающимися или невскрывающимися. У многих плодов вскрывание сухого околоплодника обеспечивает распространение семян. Сухие многосемянные плоды чаще вскрываются при помощи отверстий, крышечек, створок.
Соплодие - это совокупность зрелых, сросшихся между собой плодов (сухих или сочных) одного соцветия (шелковица, хлебное дерево, ананас).
У цветковых растений сформировались различные приспособления для распространения семян и плодов. У многих растений плоды распространяются ветром. Они имеют небольшие размеры, легкие, часто снабжены крыловидными придатками или летучками (одуванчик, бодяк).
Плоды некоторых растений распространяются водой (ольха, кувшинка, кокосовая пальма, многие виды осок). Сочные плоды с яркой окраской и ароматным запахом охотно поедаются многими видами животных. Семена этих плодов не перевариваются и вместе с экскрементами попадают в почву. У сухих плодов развиваются различные прицепки, крючочки, которые цепляются за шерсть животных, одежду человека и переносятся ими (лопух, липучка, череда). У некоторых растений созревшие плоды растрескиваются и раскручивающиеся створки околоплодника отбрасывают семена -- иногда на значительное расстояние (недотрога, бешеный огурец).
Семя. В типичном случае семя состоит из зародыша и эндосперма, окруженных семенной кожурой (рис. 8.22).
Семенная кожура образуется из покровов семязачатка. Она предохраняет семя от механических повреждений, проникновения патогенных микроорганизмов, излишней потери воды. Семена многих растений имеют различные приспособления для распространения - волоски (ивы, тополя), мясистые придатки (для привлечения животных).
Зародыш - это зачаток новой особи, миниатюрный спорофит. У большинства цветковых растений зародыш состоит из зародышевого корешка, стебелька и почечки. К верхней части стебелька прикрепляются семядольные листья {семядоли). При этом у двудольных растений закладываются две семядоли (они содержат запасные вещества семени), у однодольных - одна. Единственная семядоля зародыша злаков называется щитком. Она расположена на границе с эндоспермом и при прорастании зерновки способствует поступлению растворенных питательных веществ из эндосперма к зародышу.
Эндосперм - ткань, содержащая питательные вещества. Она формируется не только у почти всех однодольных растений, но и у ряда двудольных (лен, укроп, морковь и др.). В тришюидных клетках эндосперма, а также в семядолях зародыша содержится значительный запас питательных веществ (крахмал, белки, жиры и др.), необходимых для развития зародыша и формирования проростка.
Классификация семян проста. Различают пять типов семян в зависимости от того, где накапливаются запасные продукты: в эндосперме, нуцеллусе, зародыше, в эндосперме и нуцеллусе в эндосперме и зародыше.
Развитие семязачатка начинается с появлением срединного бугорка - нуцеллуса, внутри которого на ранних стадиях развития выделяется одна крупная спорогенная клетка. Позднее она подвергается редукционному (мейотическому) делению и образует 4 неравноценные клетки -мегаспоры. Поэтому нуцеллус следует рассматривать как мегаспорангий. Нуцелус составляет центральную часть семязачатка
Семя имеет очень важную особенность: в условиях, неблагоприятных для прорастания, оно может значительное время пребывать в состоянии покоя. С наступлением благоприятных условий температуры и влажности семя начинает поглощать воду и при достаточном доступе воздуха прорастает. Семена распространяются водой, ветром, животными, случайное распределение(бешеный огурец..одногрупники внимание: когда семена бешеного огурца достигают полной зрелости, окружающие их мясистые ткани превращаются в слизистую массу.
При отделении огурца от плодоножки создается давление на содержимое плода по принципу реактивной тяги, которая двигает плод в противоположную полету семян сторону, что способствует разбрасыванию семян на более значительной площади.
18.Лишайники. Особенности строения, распространение, цикл развития, значение в природе и жизни человека. Представители
Лишайники – это пример облигатного симбиоза грибов с водоpослями. По хаpактеpу полового споpоношения лишайники относят к двум классам: сумчатые (pазмножаются споpами, созpевающими в сумках), к котоpым относятся почти все pазновидности лишайников, и базидиальные (споpы созpевают в базидиях), насчитывающие всего несколько десятков видов. По строению тела (слоевища ) различают накипные (коpковые), листоватые и кустистые лишайники. Вегетативное тело накипных лишайников наиболее пpимитивно, оно бывает зеpнистым, пористым или в виде коpочек. Более pазвиты листоватые лишайники, имеющие вид более или менее pассечённых пластинок. Высоко оpганизованы кустистые лишайники, имеющие вид кустиков, свисающих нитей или пpямостоящих выpостов.
Распространение. Лишайники растут на почве (эпигейные ), камнях (эпилитные ) или древесных стволах (эпифитные ), получая необходимую для жизни влагу из атмосферы. Некоторые виды обитают на морской литорали. Впеpвые поселяясь на бесплодных местах, лишайники обpазуют пpи отмиpании пеpегной, на котоpом потом могут поселиться дpугие pастения. Лишайники обнаружены даже в бесплодных арктических пустынях и внутри антарктических горных пород. Лишайники распространены по всему миру, но особенно разнообразны в тропиках, высокогорьях и в тундре. А вот в лабораториях лишайники достаточно быстро погибают. И только в 1980 году американские ученые сумели «соединить» водоросль и гриб, выращенный из споры. Лишайники – это организмы-биоиндикаторы; они растут только в экологически чистых местах, поэтому их не встретишь в больших городах и промышленных зонах.
Размножение. Размножаются лишайники главным образом вегетативным путем - частями слоевища, а также особыми специализированными образованиями - соредиями и изидиями (рис. 6.4).
Соредии формируются под верхней корой в фотосинтезирующем слое и состоят из одной или нескольких клеток фикобион-та, оплетенных гифами гриба. Под давлением разросшейся массы многочисленных соредий корковый слой таллома разрывается, и соредии выходят на поверхность, откуда разносятся ветром, водой и в благоприятных условиях прорастают в новые талломы лишайников.
Изидии представляют собой мелкие выросты слоевища в виде палочек, бугорков, покрытых снаружи корой. Состоят они из нескольких клеток фикобионта, оплетенных гифами гриба. Изидии отламываются и формируют новые слоевища.
Значение лишайников в биосфере и народном хозяйстве. Известно около 26 тыс. видов лишайников. Они широко распространены в природе за исключением мест, где воздух насыщен вредными газами. Лишайники весьма чувствительны к загрязнению воздуха и поэтому большинство из них в крупных городах, а также вблизи заводов и фабрик быстро погибает. По этой причине они могут служить индикаторами загрязненности воздуха вредными веществами.
Будучи автогетеротрофными организмами, лишайники аккумулируют солнечную энергию и создают органические вещества в местах, недоступных другам организмам, а также разлагают органику, участвуя в общем круговороте веществ в биосфере. Лишайники играют существенную роль в почвообразовательном процессе, так как они постепенно растворяют и разрушают горные породы, на которых поселяются, а за счет разложения их слоевищ происходит формирование почвенного гумуса. Таким образом, лишайники вместе с бактериями, цианобактериями, грибами и некоторыми водорослями создают условия для других, более совершенных организмов, в том числе для высших растений и животных.
В хозяйственной деятельности человека важную роль играют прежде всего кормовые лишайники, такие как олений мох, или ягель, исландский мох и другие, которые поедаются не только северными оленями, но и маралами, кабаргой, косулями, лосями. Некоторые виды лишайников (лишайниковая манна, гигрофора) используют в пищу, также они нашли применение в парфюмерной промышленности -для получения ароматических веществ, в фармацевтической - для изготовления препаратов против туберкулеза, фурункулеза, кишечных заболеваний, эпилепсии и др. Из лишайников получают лишайниковые кислоты (известно около 250), обладающие антибиотическими свойствами.
Представители: Олений мох, кустистый лишайник «борода».
У двудольных растений стебель, до начала деятельности камбия, производящего коренную перестройку его анатомии, имеет следующее строение: снаружи он покрыт эпидермисом , далее идет первичная кора , составленная несколькими рядами преимуще-ственно паренхиматических клеток; далее к центру располагается цен-тральный цилиндр , составленный сосудисто-волокнистыми пуч-ками и сердцевиной. Эндодерма , отделяющая кору от центрального цилиндра, имеется лишь у немногих растений.
Эпидермис составлен однородными, плотно придвинутыми друг к другу клетками обычного для эпидермиса строения. В первичной коре обычно различают два слоя. Наружный составлен из клеток, плотно сое-диненных друг с другом. Клетки эти имеют утолщения в углах оболочек и представляют собой колленхиму. Внутренний слой коры составлен бо-лее крупными, рыхло соединенными между собой клетками, имеющими тонкие оболочки. Эндодерма (при наличии ее) представлена одним слоем крупных клеток, содержащих зерна крахмала («крахмальное влагалище» ).
Центральный цилиндр начинается перициклом , который яв-ляется местом образования придаточных корней (возникающих эндогенно). Иногда перицикл развивается в многоклетную механическую ткань, оде-вающую снаружи сосудистый пучок. Такие перицикловые во-локна имеют утолщенные, часто одревесневшие стенки, или же они сох-раняют клетчатковый состав (рис. 105). Основной, наиболее важной частью центрального цилиндра является проводящая ткань в виде открытых сосудисто-волокнистых пучков, расположенных правильным кольцом. Они коллатерального типа, т. е. ксилема их обращена внутрь, флоэма — кнаружи, а между ними проходит узкая прослойка камбия (рис. 106). Пучки разделены группами паренхимных клеток, соединяющих кору с сердцевиной. Это будут первичные сердцевинные лучи . Иногда сосудистые пучки расположены не одним, а двумя слоями. В этом случае внутренние пучки состоят только из флоэмы. При тесном примы-кании их к наружным пучкам возникают биколлатеральные пучки (у тыквы ).
Сердцевина, занимающая центральную часть стебля, составлена круп-ными, плотно соединенными паренхимными клетками. Иногда сердцевина исчезает, а вместо нее в центре стебля образуется воздушная полость (у зонтичных ). Материал с сайта