23. Анатомическое строение стеблей травянистых и древесных однодольных растений.
24. Анатомическое строение стеблей травянистых двудольных растений.
25. Анатомическое строение стеблей древесных двудольных растений.
26. Анатомическое строение стеблей хвойных.
27. Анатомическое строение корневищ одно- и двудольных растений.
31. Понятие стебля, побега. Функции и метаморфозы стебля. Морфологическая характеристика стебля.
Стебель - надземный, вегетативный, осевой орган растения, имеющий радиальную симметрию и обладающий верхушечным ростом.
Стебель с почками и листьями - побег . Участки стебля, несущие лист -стеблевые узлы: между узлами - междоузлия . В зависимости от длины междоузлии побеги могут быть удлиненные (злаки), укороченные (одуванчик).
Побег развивается из почки.
Верхушечная почка - верхушка стебля, прикрытая молодыми зачатками листьев: конус нарастания, в основании его находятся первичные бугорки - из них формируются примордиальные листья. В пазухах первичных бугорков - вторичные бугорки, которые дают боковые или пазушные почки. Боковые ветви тоже растут своими верхушками и каждая ветвь кончается верхушечной почкой.
С наступлением осени все почки вступают в сезонный покой. Их называют зимующими. Они могут быть вегетативными (образующими листья или побеги) и вегетативно - генеративными или цветковыми (весной распускающиеся в цветки или соцветия). Защищены твердыми чешуями, покрытыми кутикулой. Имеют склереиды, иногда пробку. Распускаются за счет запасов, отложенных в корне, стебле (или в самих кроющих чешуях).
Многие пазушные почки остаются в покое долгое время и подрастают лишь своей осью на стебле в соответствии с его утолщением - спящие почки. Они трогаются в рост, если удалить верхушечную почку. Иногда могут быть погружены в древесину и после поломки или сруба ствола просыпаются.
Придаточные почки закладываются в узлах и на междоузлиях, корнях, корневищах, листьях - резерв вегетативного размножения. Происхождение: из камбия, феллогена, перицикла, коры. Это почки омоложения - дают побеги, похожие на молодые сеянцы.
Морфологические признаки стебля.
I. По положению в пространстве стебли бывают: прямостоячие, приподнимающиеся, ползучие, лазающие, вьющиеся и т.д.
II. Ветвление побегов .
1. Дихотомическое - точка роста вильчато разделяется на 2 новые (низшие, папоротникообразные, мохообразные).
2. Моноподиальное - главная ось растет своей верхушкой, боковые ветви развиваются из боковых почек, которые тоже ветвятся моноподиально (хвойные - очень ценятся стволы в лесотехническом деле, кораблестроении).
3. Симподиальное - верхушечная почка отмирает, вместо нее развивается боковая почка, растет часто в вертикальном направлении, как бы продолжая рост главного стебля и т.д. (береза, ива, яблоня, томат, картофель).
Известно смешанное ветвление: сначала моно-, затем симподиальное (у древесных плодовых, хлопчатника).
Интенсивное распускание большого числа почек делает побег укороченным, что обеспечивает развитие на кроне огромного количества листьев, т.е. большой фотосинтезирующей поверхности.
4. Ложнодихотомическое ветвление - под верхушечной почкой образуется две супротивно расположенные пазушные почки; трогаясь в рост, они образуют развилку (омела, дурман, гвоздика, сирень, конский каштан).
Изучение систем ветвления имеет огромное практическое значение. Можно регулировать урожай.
III. Листорасположение: листья укреплены на стеблевых узлах.
1. Очередное - узел имеет один лист - яблоня, береза, клен.
2. Супротивное - два листа в узле (губоцветные, валериана, гортензия).
3. Мутовчатое - три и более листьев - олеандр.
4. Прикорневая розетка - первоцвет.
Очередные листья располагаются на стебле по спирали. Следя за ее оборотами, можно обнаружить два листа, расположенных по вертикали один над другим. Линия, соединяющая листья в продольных рядах на стебле – ортостиха . Между концами ортостихи остальные листья располагаются по спирали, число оборотов спирали между двумя листьями, расположенными на одной ортостихе называются - листовым циклом. Составляется формула очередного листорасположения в виде дроби, где числителем становится число оборотов спирали в листовом цикле, а знаменателем - число листьев в цикле.
IV. Форма стебля в поперечном сечении .
Форма стебля в поперечном сечении бывает самой разнообразной: округлые, плоские, ребристые, многогранные и т.д.; могут быть полые и выполненные, голые и опушенные.
Продолжительность жизни и биоморфологические группы растений. Кипарис - 3000 лет, каштан - 2000 лет, дуб - 1200 лет, липа - 1000 лет, тополь - 300-600 лет, яблоня -200 лет, черешня – 40-100 лет, сосна - 50 лет. В зависимости от продолжительности жизни и типа побегов растения делят на:
деревья - крупные растения с сильно развитыми многолетними стеблями. Имеется вторичный прирост.
кустарники - главный ствол отсутствует или слабо выражен, ветвление начинается почти у поверхности почвы (4-6). Барбарис, ирга, шиповник. Выделяются группы кустарников не более 1м - чабрец, саксаул.
полукустарники - побеги в нижней части остаются многолетними, в верхней части - однолетние побеги отмерзают или отсыхают - полынь, астрагал, зверобой;
травянистые - отмирание надземных стеблей наблюдается в конце вегетационного периода. Большинство полевых растений - хлебные злаки, горох, фасоль, лен, конопля;
двулетние травянистые - на первый год развивается только розетка прикорневых листьев, а цветы, плоды и удлиненный стебель - на второй (цикорий, белена, алтей, донник);
многолетние - долговечные подземные части и ежегодное отмирание надземных частей (валериана, ландыш, крапива).
Среди травянистых растений существуют озимые и яровые формы:
а) если семена прорастают весной - яровые;
б) озимые - прорастание семян осенью (0-5 град.).
Функции стебля :
1. Остов, соединяющий листья и корни воедино, придает габитус растению;
2. Проводящая;
3. Запасающая;
4.Синтетическая;
5. Орган вегетативного размножения.
Метаморфозы:
Подземные : 1. Корневище - подземный побег, листья в виде чешуек, размеры корневищ определяются функциями: если для запаса - толстые (аир, хвощ, купена), для размножения - с почками возобновления (пырей, касатик).
2. Клубни - образуются на концах подземных стеблей - столлонов, имеют почки - глазки (по 3 и больше).
3. Луковицы - укороченный стебель - донце, почки и листья.
Наземные: 4. Колючки - боярышник, терн.
5. Усы - тыква, огурец, дыня.
6. Филлокладии - стебель в виде листа - иглица.
Стебель - надземный, вегетативный, осевой орган растения; обладает длительным ростом в длину с помощью верхушечной меристемы в конусе нарастания или вставочной меристемы; несет на себе листья, залагающиеся в определенном порядке на конусе нарастания в виде бугорков; может ветвиться за счет почек, залагающихся в пазушных листья; имеет радиальное строение и несколько плоскостей симметрии.
На растущем конце стебель несет верхушечную почку - конус нарастания, одетый молодыми, видоизмененными листочками. Заканчивается точкой роста, за которой располагаются первичные бугорки - зачатки листьев. Еще ниже в пазухах их возникают вторичные бугорки - зачатки боковых побегов.
Теория гистогенов Гапштейна (1818 г.): дерматоген, периблема, плерома - неверна для стебля. В начале 20 годов нашего столетия Шмидт сформулировал теорию туники и корпуса, согласно которой конус нарастания у покрытосеменных и части голосеменных состоит из двух гистологически разных частей, обладающих разной меристемной активностью. Наружная часть - туника, клетки ее делятся перпедикулярно поверхности стебля и образуют эпидерму, иногда эпидерму и несколько слоев или всю первичную кору; внутренняя часть - корпус - клетки делятся во всех направлениях и образуют остальные слои первичной коры и ЦОЦ.
Первичная структура стебля складывается при дифференциации клеток верхушечной меристемы.
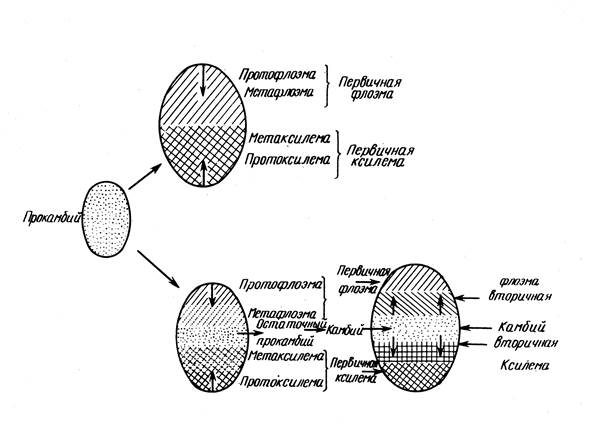
На уровне первых листовых зачатков клетки туники и корпуса перестают делиться; периферическая их часть идет на формирование первичной коры, внутренняя часть на формирование сердцевины. Но между ними сохраняется несколько рядов активных меристематических клеток, располагающихся кольцом - образовательное кольцо . Его клетки в основании молодых зачатков листьев дают начало первичной боковой меристеме - прокамбию.
Прокамбий представляет собой эмбриональную фазу в развитии первичной проводящей меристемы, поэтому его расположение в значительной степени предопределяет последующее расположение проводящей системы. Если он закладывается сплошным слоем (кольцом), то возникают сплошные слои флоэмы и ксилемы, если же прокамбий закладывается в виде пучков (тяжей), то и первичные проводящие ткани располагаются в виде пучков. Клетки периферических слоев образовательного кольца, не участвующие в формировании прокамбия, образуют перицикл.
Прокамбий возникает у основания листового зачатка и отсюда его развитие распространяется в двух направлениях - акропетально, т.е. к верхушке листового зачатка, и базипетально, т.е. вниз по стеблю, где он причленяется к другим пучкам, возникшим ранее.
Флоэма образуется раньше ксилемы и развивается центростремительно, сначала протофлоэма, затем протоксилема (кольчатые и спиральные сосуды), затем метаксилема. Таким образом формируется ЦОЦ первичного строения.
На поперечном срезе стебля различают три группы тканей: покровную, первичную кору и ЦОЦ.
Покровная ткань стебля травянистого двудольного растения – эпидерма. Клетки мало извилистые, вытянутые в направлении стебля, устьиц мало или нет.
Первичная кора состоит из колленхимы в виде тяжей в углах стебля у губоцветных или в его ребрах у зонтичных, реже в виде кольца - у тыквенных, пасленовых. На периферии первичная кора состоит из чередующихся полосок хлоренхимы и механической ткани. Во внутренней части бесцветная паренхима. Самый внутренний слой - эндодерма в ряде случаев - крахмалоносное влагалище, содержащее мелкие зерна оберегаемого крахмала.
Периферическая часть центрального цилиндра - перицикл. Проводящие ткани располагаются в виде пучков или сплошным кольцом (в зависимости от заложения прокамбия). Переход ко вторичному строению связан с заложением камбия, в зависимости от чего различают следующие типы строения стеблей:
1. Прокамбий закладывается сплошным кольцом, первичное строение непучковое; камбий образуется из прокамбия; вторичное - непучковое.
2. Прокамбий закладывается тяжами - первичное строение пучковое; камбий формируется из прокамбия в пучках и из основной ткани ЦОЦ между пучками. Образуется сплошное камбиальное кольцо: вторичное строение непучковое.
3. Первичное и вторичное строение пучковое, т.к. межпучковый камбий не образуется.
4. Первичное и вторичное строение пучковое, т.к. прокамбий закладывается отдельными тяжами, а образующийся межпучковый камбий образует основную ткань Ц.О.Ц., а не флоэму и ксилему.
Стебель однодольных травянистых растений. Для стебля однодольных травянистых растений характерно пучковое строение, пучки расположенны беспорядочно, отсутствие вторичного прироста. Основные черты анатомического строения стеблей определяются системой листовых следов: из основания листа в стебель входят пучки, располагающиеся по всей окружности, таким образом все сосудистые пучки однодольных представляют собой листовые следы (пальмовый тип). СВП однодольных имеют характерную форму: два крупных точечных сосуда (симметрично) и 1-2 более узких с кольчатыми и спирально- кольчатыми утолщениями. К крайнему из них примыкает воздушная полость, образовавшаяся на месте разрушения элементов протоксилемы. Величина пучков возрастает от периферии к центру.
В строении коры имеются различия:
1. Однодольные с выраженной первичной корой - (спаржа): состоит из нескольких слоев хлорофиллоносных клеток; колленхима отсутствует.
2. У других однодольных (пальма, злаки) первичная кора не выражена; сразу под эпидермой имеются участки хлоренхимы, окруженные склеренхимой, которая сливается со склеренхимой СВП.
В междоузлиях многих однодольных образуется крупная центральная полость (соломина злаков), в этих случаях пучки оказываются сдвинутыми к периферии, но располагаются также разбросанно: более мелкие - к периферии, крупные - к полости.
Строение стеблей древесных двудольных. Первичное строение - непучковое. Вторичное связано с заложением феллогена и камбия. Камбий образует вторичную флоэму и ксилему. Флоэма - луб - твердый и мягкий расположена в виде трапеций, разделенных верхушками первичных сердцевинных лучей; функцию проведения выполняет заболонь - 1 мм у камбия. Древесина в виде годичных колец; ядровая древесина - механическую функцию.
Сердцевина рыхлая, сначала живая ткань с запасом питательных веществ. Соединена с лубом и корой сердцевинными лучами (первичными и вторичными)
Феллоген образуется за счет эпидермы и слоев первичной коры, формирует перидерму и корку. Под ней сохраняется первичная кора - колленхима, хлоренхима, запасающая ткань, - эндодерма не выражена (иногда дифференцирована в крахмалоносное влагалище).
Согласно ГФ Х1 (1987), кора в фармацевтической практике – это наружная часть стволов, ветвей и корней деревьев и кустарников, расположенная к периферии от камбия. По внешним признакам цельная кора имеет вид трубчатых, желобоватых или плоских кусков различных размеров. Наружная поверхность коры с бурой или серой пробков обычно гладкая или с продольными (или с поперечными) морщинками, иногда с трещинками. Кора ветвей и стволов имеет округлые или продолговатые чечевички, иногда на ней могут быть листовые лишайники (при заготовке должны удаляться). Внутрення поверхность коры обычно более светлая, гладкая или ребоистая. Поперечный излом обычно неровный: занозистый, волокнистый или зернистый.
На микропрепарате обращают внимание на наружнюю кору, располагающуюся к периферии от окончания сердцевинных лучей и состоящую из перидермы и остатков первичной коры, и внутреннюю, состоящую из флоэмы. Имеет диагностическое значение толщина, окраска и характер пробки, наличие колленхимы, соотношение толщины первичной и вторичной коры, ширина сердцевинных лучей, количество, расположение и строение лубяных волокон, каменистых клеток, а также включения оксалата кальция, млечники, клетки с эфирным маслом.
Строение стеблей хвойных. Первичное - пучковое, затем образуется сплошное камбиальное кольцо и кольца флоэмы и ксилемы. Закладывается феллоген.
В коровой части: нет колленхимы, есть смоляные ходы.
2.Во флоэме нет клеток-спутниц, лубяных волокон, лубяной паренхимы.
3. Камбий многорядный.
Древесина состоит из трахеид, механические волокна и древесная паренхима отсутствуют.
Первичное строение пучковое.
Строение стеблей древесных однодольных растений. Свойственно древовидным представителям семейства лилейных (драцена, юкка, алоэ). Молодой стебель близ конуса нарастания построен как у всех однодольных: покровная ткань - эпидерма; под ней - первичная кора, представленная несколькими слоями пластинчатой колленхимы; ЦОЦ начинается перициклом и содержит закрытые коллатеральные СВП, расположенные беспорядочно. Утолщение происходит за счет формирования кольца утолщения, образующегося из перицикла или из внутреннего слоя первичной коры. Образует центробежно центроксилемные концентрические СВП. Покровная ткань - пробка.
Строение корневищ однодольных растений. Аналогично древесному стеблю. Покровная ткань - одревесневшая экзодерма; первичная кора всегда развита, представлена запасающей тканью; эндодерма типичная с пятнами Каспари и подковообразными утолщениями. В ЦОЦ - коллатеральные пучки (V) и концентрические центрофлоэмные. Образуются из кольца утолщения.
Строение корневищ двудольных. Покровная ткань - пробка, перидерма образуется из феллогена; первичная кора - запасающая паренхима; эндодерма с пятнами Каспари или подковообразная; в ЦОЦ расположение тканей аналогично наземному стеблю; у пучкового типа - пучковое; у переходного и непучкового - непучковое.
Эволюция стелы. В процессе эволюции наблюдалось изменение в строении и взаимном расположении различных тканей растения, особенно проводящих. Установлено несколько эволюционных типов строения стебля, касающихся ЦОЦ или стелы (Ван Тигем - французский ботаник). Стеллярная теория позднее получила блестящее подтверждение при изучении вымерших растений (риниофитов). Наиболее древний тип - протостела. Тяж ксилемы, покрытый слоем флоэмы, актиностела - ксилема имеет форму звезды (плауны, хвощи), окруженной флоэмой
Сифоностела - появляется сердцевина.
Диктиостела - сифоностела рассекается на отдельные пучки (папоротники).
Эустела - открытые коллатеральные пучки пучки (двудольные).
Атактостела - у однодольных закрытые коллатеральные пучки, расположенные беспорядочно.
Заключение. Разнообразие типов строения стебля и корневищ необходимо знать при макро- и микроскопической диагностике лекарственного растительного сырья.
Стебель является частью лекарственного растительного сырья – трава (Herba). При макроскопической диагностике травы в соответствии с Государственной Фармакопеей Х1 (1987) отмечают следующие особенности стебля: тип ветвления, форму поперечного сечения, опушение, размеры (длину и диаметр у основания), листорасположение. На микропрепаратах поперечного среза стебля отмечают особенности строения клеток эпидермиса, расположение проводящих пучков, наличие и характер расположения механических тканей, кристаллических включений, вместилищ, секреторных каналов, млечников и других особенностей диагностического плана.
Корневища (Rhizomata) при анализе по внешним признакам отличаются формой (простые или разветвленные, цилиндрические или овальные, четковидные, прямые, изогнутые или перекрученные и т.д.), особенностями наружной поверхности (ровная или морщинистая со следами удаленных корней), характером излома (ровный, зернистый, занозистый или волокнистый). Иногда невооруженным глазом рассматривают на изломе характер расположения проводящих пучков. При микроскопической диагностике отмечают беспучковое или пучковое строение, особенности расположения и типы пучков, строение покровной ткани, запасающей и сердцевины, наличие разнообразных вместилищ, механических элементов, каналов, млечников, кристаллов оксалата кальция, запасных питательных веществ (крахмал, слизь, инулин, жирное масло и т.п.).
При анализе луковиц (Bulbi), клубней (Tubera) и клубнелуковиц (Bulbotubera) имеет значение форма (шаровидные, яйцевидные, овальные, продолговатые, сплющенные, веретеновидные и т.д.), размеры, характер поверхности, особенности строения чешуй (сухие или сочные). При микроскопической диагностике отмечают особенности запасающей ткани и расположения пучков.
Строение стебля двудольных травянистых растений
На ранних этапах развития стебель двудольных имеет первичное строение. В результате деятельности первичных меристем конуса нарастания формируются эпидерма, первичная кора, центральный цилиндр и сердцевина.
Эпидерма стебля имеет небольшое число устьиц, обычно покрыта кутикулой. Под эпидермой находятся ткани первичной коры. Наружный слой ее (экзодерма) часто образован колленхимой, которая располагается либо сплошным кольцом (подсолнечник), либо отдельными участками. У растений с ребристыми стеблями (тыква) колленхима заполняет выступы стебля. В тонких стеблях колленхима обычно уголковая, в стеблях с активным вторичным утолщением - пластинчатая. Основная часть первичной коры (мезодерма) сложена хлоренхимой и паренхимой. Здесь могут развиваться воздухоносные полости и вместилища выделений. Клетки внутреннего слоя первичной коры (эндодермы) обычно заполнены крупными крахмальными зернами (крахмалоносное влагалище).
Центральный цилиндр (стела) состоит из перицикла и проводящей системы. Сердцевина - из паренхимы, которая часто разрушается, а на ее месте образуется воздухоносная полость. Перицикл представлен одним или несколькими рядами склеренхимы или отдельными ее тяжами. Проводящая система имеет вид полого цилиндра, который разделяет первичную кору и сердцевину. Проводящие пучки, составляющие этот цилиндр, разделены прослойками межпучковой паренхимы – сердцевинными лучами, которые связывают сердцевину и первичную кору. Большинству двудольных свойственен особый тип центрального цилиндра – эустель, на поперечном срезе которой пучки расположены кольцом (рис. 4) .
Рисунок 4. Эустель двудольных.
У двудольных растений слой клеток прокамбия между первичными флоэмой и ксилемой сохраняет способность к делению и превращается во вторичную образовательную ткань - камбий , который делится, откладывая внутрь элементы вторичной ксилемы, а к периферии - вторичной флоэмы. Пучки двудольных растений открытые, благодаря работе камбия пучок растет, диаметр его увеличивается. Деятельность камбия обеспечивает возникновение вторичных элементов в стебле, т. е. переход от первичного к вторичному анатомическому строению.
Вторичное строение характерно для всех двудольных и хвойных растений - трав и деревьев. Разнообразие типов строения обусловлено, прежде всего, расположением проводящих тканей, которое определяется заложением прокамбия и деятельностью камбия.
Прокамбий в процессе дифференциации конуса нарастания закладывается в виде тяжей. Если тяжи разделены достаточно широкими рядами паренхимы, то формируется пучковое или переходное строение стебля; если тяжи прокамбия сближены настолько, что сливаются в цилиндр, то формируется непучковое (сплошное) строение.
1. Пучковое строение стебля встречается у некоторых травянистых растений (укроп, клевер, лютик, горох, люцерна). Заложенные в конусе нарастания прокамбиальные тяжи располагаются в один круг по периферии центрального цилиндра . Каждый прокамбиальный тяж превращается в открытый коллатеральный пучок, состоящий из первичной ксилемы, первичной флоэмы и полоски камбия между ними. Клетки камбия, делясь, дают новые (вторичные) элементы проводящего пучка: внутрь - ксилему к периферии - флоэму. При этом наиболее молодые участки флоэмы и ксилемы примыкают к камбию, а более старые отодвигаются к периферии пучка. Самые крайние положения займут первичные флоэма и ксилема. Проводящие пучки разделены широкими межпучковыми зонами и окружают мощную сердцевину.
У большинства растений камбий работает активно и диаметр пучков значительно увеличивается Деятельность пучкового камбия стимулирует паренхиму, разделяющую пучки, которая начинает делиться, давая вторичную образовательную ткань. Пучковая и межпучковая меристемы смыкаются и образуют сплошное камбиальное кольцо, деятельность которого обеспечивает равномерное утолщение стебля. Если пучковый камбий дает элементы вторичных ксилемы и флоэмы, то межпучковый камбий производит паренхиму (у укропа, звездчатки злаковидной и др.). У клевера, люцерны и других растений межпучковый камбий откладывает преимущественно склеренхимоподобные клетки в сторону ксилемы. Эти клетки удлиненные, толстостенные одревесневающие. Поэтому задержка с уборкой клевера снижает качество сена. Отчетливое пучковое строение сохраняется у этих растений в течение всей жизни стебля.
Пучковое строение имеет стебель клевера . Под эпидермой расположена первичная кора, представленная хлорофиллоносной паренхимой. Наружный ряд ее клеток - пластинчатая колленхима внутренний - клетки крахмалоносного влагалища (эндодермы). Граница между первичной корой и центральным цилиндром хорошо заметна благодаря склеренхиме перициклического происхождения.
Склеренхима образует полудуги над открытыми коллатеральными пучками, расположенными по кругу. В составе пучка - первичная флоэма, примыкающая к склеренхиме, вторичная флоэма, камбий, вторичная ксилема и первичная ксилема. Пучковый камбий смыкается с межпучковым, образуя цельное меристематическое кольцо. Межпучковый камбий откладывает внутрь одревесневающую паренхиму, которая соединяет проводящие пучки. Возникает кольцо (цилиндр) лигнифицированных тканей, обеспечивающее высокую прочность стебля. Однако проводящие ткани остались разобщенными в виде пучков. Сердцевина стебля представлена паренхимой.
Пучковое строение стебля может с возрастом смениться непучковым сплошным. 2. Переходное строение стебля характерно для подсолнечника . Вначале стебель формируется так же, как у клевера, но межпучковый камбий откладывает не паренхиму, а ксилему и флоэму. Формируются добавочные проводящие пучки. Постепенно все пучки могут слиться в сплошной трехслойный (ксилема, камбий, флоэма) цилиндр. В верхней (молодой) части стебель имеет пучковое строение, у основания (в старой части) - непучковое .
3. Непучковое строение свойственно стеблям многих трав и деревьев. В конусе нарастания этих растений прокамбиальные тяжи настолько сближены, что образуют почти сплошной цилиндр (на поперечном срезе он имеет вид кольца). Прокамбиальный цилиндр (кольцо), дифференцируясь, дает к центру цилиндр первичной ксилемы, к периферии - цилиндр первичной флоэмы, а между ними образуется камбиальный цилиндр.
Вторичные изменения связаны с работой камбия. Клетки камбия делятся параллельно поверхности стебля, при этом внутрь откладывается в 10...20 раз больше клеток, чем наружу. Внутрь камбий формирует элементы вторичной ксилемы (древесины), наружу откладывается вторичная флоэма (луб). Местами камбий откладывает в обе стороны паренхимные клетки сердцевинных лучей.
Особенности непучкового строения стеблей трав можно рассмотреть на примере стебля льна . Стебель покрыт эпидермой, под которой располагается рыхлая хлоренхима первичной коры. Большое количество устьиц в эпидерме и отсутствие колленхимы в первичной коре имеют существенное практическое значение, так как во время технической мочки льна вода не встречает препятствий и легко проникает внутрь стебля. Клетки крахмалоносного влагалища (эндодермы) более крупные, содержат хорошо заметные крахмальные зерна.
Примыкающий к эндодерме центральный цилиндр начинается группами склеренхимных клеток – лубяных волокон, между которыми расположены клетки паренхимы. Лубяные волокна возникают в перицикле и в первичной флоэме. Они отличаются очень толстыми, обычно неодревесневшими стенками, состоящими на 75...90 % из целлюлозы. Длина лубяных волокон 4...60 мм, диаметр 12...37 мкм. Во время роста склеренхимной клетки ее ядро делится, но цитокинеза не происходит, и протопласт оказывается многоядерным. Клеточная стенка сильно утолщена, полость клетки похожа на узкий канал. Клеточная стенка состоит из многих слоев целлюлозы и почти лишена пор. Созревшая клетка лубяного волокна мертвая. Протопласт после утолщения клеточной стенки разрушается. Лубяные волокна обладают необыкновенно высокой прочностью. Сопротивление разрыву у них соответствует сопротивлению стали, а упругость выше. Клетки лубяного волокна очень тесно соединены между собой. Льняное волокно - великолепное прядильное сырье, используемое в текстильной промышленности.
Далее в последовательном порядке располагаются цилиндры (кольца): вторичной флоэмы, камбия, вторичной ксилемы с хорошо заметными сердцевинными лучами. Первичная ксилема расположена участками, примыкающими к сердцевине. Паренхима сердцевины быстро разрушается (в процессе удлинения стебля), образуя центральную полость.
Стебли двудольных травянистых растений характеризуются:
1. вторичным строением, очень рано возникающим вслед за первичным;
2. наличием первичной покровной ткани - эпидермы;
3. развитой первичной корой, делящейся на колленхиму (экзодерма), паренхиму (мезодерма) и крахмалоносное влагалище (эндодерма);
4. пучковым или непучковым (сплошным) строением;
5. правильным расположением коллатеральных или биколлатеральных проводящих пучков в один круг по периферии центрального цилиндра;
6. наличием камбия (пучки открытые),
7. сердцевина хорошо развита.
Первичное анатомическое строение стебля.
Определение и функции стебля.
Стебель – осевой, обычно радиально симметричный орган, обладающий длительным верхушечным (апикальным) ростом.
Функции стебля:
1. осуществляет двустороннее передвижение веществ между корнями и листьями,
2. поддерживает крону растения,
3. способствует увеличению общей ассимиляционной поверхности растения благодаря ветвлению,
4. участвует в хранении запасных веществ,
5. в молодом возрасте – осуществляет также фотосинтез.
Эти функции определяют наличие в стебле покровных, хорошо развитых проводящих, механических тканей и функционально развитой паренхимы.
Строение конуса нарастания.
В конусе нарастания стебля покрытосеменных растений выражены две зоны: наружная – туника , и внутренняя – корпус, которые различаются плоскостями клеточных делений (теория А. Шмидта и Дж. Будера) (рис. 1).
Туника может состоять из одного или нескольких слоев клеток, делящихся перпендикулярно поверхности, у большинства двудольных растений она двухслойная. Наружный слой туники дает начало протодерме, из которой затем образуется эпидерма, покрывающая листья и стебель. Если туника многослойная, то внутренний слой образует ткани первичной коры.
При наличии однослойной туники всю первичную кору и стелу образует корпус. Корпус образован массой клеток, которые делятся во всех плоскостях .
Зачатки листьев чаще всего возникают во втором слое туники, а пазушные почки закладываются в корпусе.
Рисунок 1. б) апекс покрытосеменных растений.
В результате деятельности первичных меристем апекса формируется первичное анатомическое строение стебля: эпидерма, первичная кора, центральный цилиндр и сердцевина.
Вторичное утолщение происходит за счет деятельности камбия.
Первичное анатомическое строение стебля.
Строение стебля, сформировавшегося в результате деятельности первичных меристем апекса, называется первичным . В стебле, при первичном строении можно различить три анатомо-топографические зоны:
1. покровную ткань,
2. первичную кору
3. центральный цилиндр.
Эпидерма, защищает стебель от испарения.
Под эпидермой находится наружный слой первичной коры - экзодерма - представленная хлоренхимой и (у двудольных) колленхимой. Во внутреннем, пограничном с центральным цилиндром слое - эндодерме - обычно накапливаются крахмальные зерна, и тогда он превращается в так называемое крахмалоносное влагалище (крахмалоносный слой). Крахмальные зерна под действием силы тяжести могут оседать в клетках. Благодаря этому эндодерма играет важную роль в геотропической ((от греч. trpos - поворот, направление), ростовые движения стебля растений обусловленные направленным действием силы земного тяготения) реакции стеблей
По периферии центрального цилиндра в перицикле расположена склеренхима. Склеренхимные волокна образуют сплошной слой или собраны около проводящих пучков, кнаружи от первичной флоэмы. Главный элемент центрального цилиндра (стелы) - проводящие пучки. Они образованы из прокамбиальных тяжей и состоят из первичных ксилемы и флоэмы . Дифференциация прокамбия проходит центростремительно: флоэма формируется от внешней стороны пучка к его центру, а ксилема - навстречу ей (рис. 2). Прокамбий может полностью превратиться в первичные проводящие комплексы, образовав закрытый коллатеральный пучок (у однодольных), или сохранившиеся клетки в центре пучка формируют камбий. Такие открытые пучки характерны для двудольных растений. Работа камбия обеспечивает формирование вторичных ксилемы и флоэмы.
Рисунок 2.
Заложение прокамбия предопределяет строение проводящей системы:
1. Если прокамбий образует сплошной полый цилиндр, то из него формируются вложенные друг в друга цилиндры ксилемы и флоэмы (у некоторых хвойных и двудольных) – непучковое строение .
2. Если же прокамбий состоит из отдельных тяжей (у большинства однодольных, многих двудольных и хвойных), то возникающие первичные проводящие ткани сохраняют такое же пучковое строение .
Прокамбий возникает у основания листового зачатка, и его развитие идет в двух направлениях: к верхушке листового зачатка и вниз по стеблю, где он соединяется с тяжами, возникшим ранее. В той же последовательности идет и дифференциация флоэмы и ксилемы. Обычно раньше образуются элементы флоэмы, затем ксилемы. Проводящие пучки располагаются в паренхиме центрального цилиндра.
В центре стебля развивается хорошо заметная сердцевина. Паренхимные тонкостенные клетки сердцевины нередко разрыхляются, отмирают и заполняются воздухом, тогда вся ткань кажется белой (у подсолнечника, кукурузы, бузины). Иногда сердцевина отмирает очень рано, когда удлинение междоузлий еще не закончилось. Это влечет за собой разрыв сердцевины и образование полого стебля.
Первичное строение стебля сохраняется у однодольных растений в течение всей жизни, а у двудольных и голосеменных - лишь в ранние фазы его развития.
Анатомическое строение стебля двудольного растения пучкового строения
Стебель представляет собой ось побега. Наряду с листом он является основной структурной частью побега. Основные функции – опорная и проводящая. Через стебель осуществляется связь между корнями и листьями и обмен продуктами воздушного и минерального питания. Кроме того, в стебле нередко откладываются запасные питательные вещества. Стебель, как и весь побег в целом, представляет собой "открытую" систему роста, т.е. он длительное время нарастает, и на нем возникают новые листья.
Стебель как часть побега имеет систему меристем, поддерживающих нарастание тканей в длину и толщину. Рост в длину осуществляется за счет верхушечной и вставочных меристем, а в толщину у двудольных за счет боковых вторичных меристем – камбия и феллогена.
Первичная структура стебля складывается по мере дифференциации клеток верхушечной меристемы побега. Верхушечная меристема побега двудольных довольно рано дифференцируется на несколько групп клеток, различающихся по особенностям деления и степени меристематической активности. Наружные ее слои преобразуются в протодерму, клетки которой позднее формируют первичную покровную ткань – эпидерму.
Рис. 1. Пучковый тип с межпучковым камбием в стебле кирказона (Aristolochia clematitis) в поперечном срезе (I) и схема строения стебля на разных уровнях (II)
А - срез на уровне появления прокамбия; Б - на уровне появления камбия; В - на уровне сформированной структуры. 1 - прокамбий, 2 - эпидерма, 3 - колленхима, 4 - паренхима коры, 5 - эндодерма (3-5 - первичная кора), 6 - склеренхима перицикла, 7 - флоэма, 8 - ксилема, 9 - пучковый камбий (7-9 - открытый коллатеральный пучок), 10 - межпучковый камбий,11 - сердцевинный луч, 12 - паренхима сердцевины (6-12 - центральный цилиндр )
На уровне оснований первых листовых примордиев клетки верхушечной меристемы, расположенные к периферии и в центре апекса, перестают активно делиться, увеличиваются в размерах и вакуолизируются. Из этих клеток формируются первичная кора и сердцевина. У многих двудольных в инициальном кольце дифференцируется круг изолированных друг от друга прокамбиальных тяжей. Клетки инициального кольца, расположенные между этими тяжами, дифференцируются позднее в паренхимные элементы. На поперечных срезах эти лучи имеют вид радиальных полос, соединяющих сердцевину с первичной корой.
Прокамбий у ряда двудольных может закладываться также и в виде сплошного кольца. Он может развиваться по всей толще инициального кольца или формироваться из его части. Прокамбий является предшественником первичных проводящих тканей: первичной флоэмы и первичной ксилемы. Флоэма начинает формироваться раньше и закладывается в наружных частях прокамбиальных тяжей или прокамбиального кольца. Флоэма развивается центростремительно, т.е. самые первые элементы занимают наружное положение, а самые поздние – внутреннее. Ксилема закладывается во внутренних участках прокамбия и развивается центробежно. Таким образом, флоэма и ксилема формируются навстречу друг другу. Первые элементы ксилемы – проксилемы, узкие, сравнительно тонкостенные сосуды или трахеиды со спиральными или кольчатыми вторичными утолщениями. Метаксилема образуется несколько позднее протоксилемы и состоит из лестничных и пористых сосудов. Основным фактором, контролирующим дифференцировку как флоэмы, так и ксилемы, является фитогормон ауксин, вырабатываемый листовыми примордиями и перемещающийся по прокамбию от верхушки к основанию. Образовавшиеся из прокамбия первичные флоэма и ксилема составляют основу осевого цилиндра, или стелы.
Стела, занимающая центральную часть стебля, состоит из проводящих тканей, сердцевины, перицикла и тех постоянных тканей, которые из него возникают. К наружи от перецикла располагается первичная кора, которая состоит из из паренхимы, нередко колленхимы и иногда секреторных элементов. Сердцевина обычно состоит из относительно тонкостенных паренхимных клеток. В сердцевине часто откладываются запасные питательные вещества. Здесь же нередко встречаются идиобласты, т.е. отдельные клетки, заполненные таннидами, кристаллами, слизью и др. Иногда часть сердцевины разрушается и образуется полость. Периферическая часть сердцевины, примыкающая к ксилеме, называется перимедуллярной зоной.
Вторичное утолщение стебля у большинства двудольных возникает довольно рано, что приводит к формированию вторичного тела растения. Эти изменения связаны главным образом с активностью боковой вторичной меристемы – камбия и отчасти другой вторичной меристемы – феллогеном. За счет появления вторичных тканей осуществляется рост растений двудольных в толщину. Вторичные изменения в центральном цилиндре начинаются с заложения камбия. Камбий возникает из остатков прокамбия, на границе первичных ксилемы и флоэмы.
Клетки камбия сильно вакуолизированы и удлинены в вертикальном или в горизонтальном направлении. Первые – веретеновидные инициали дают проводящие элементы проводящих тканей, вторые – лучевые инициали – образуют горизонтально ориентированные лучевые клетки сердцевинных лучей. Инициальные клетки камбия способны к двум типам деления – периклинальному и антиклинальному. В первом случае клеточная пластинка закладывается параллельно поверхности стебля, во втором – перпендикулярно. В результате возникает непрерывный ряд производных клеток, тянущихся от камбия радиально кнаружи и вовнутрь. Клетки, откладывающиеся в сторону наружной поверхности и стебля, постепенно дифференцируются во вторичную флоэму, в сторону сердцевины – во вторичную ксилему.
Деятельность камбия активизируется фитогормонами гиббериллином и ауксином, поступающими из почек и молодых листьев. В конечном итоге в структуре стебля однолетнего побега двудольного растения возможно выделить видоизмененный центральный цилиндр, включающий постоянные ткани, возникшие из перицикла, остатки первичной и вторичную флоэму, камбий, вторичную и остатки первичной ксилемы и сердцевину. Видоизмененный центральный цилиндр окружен остатками первичной коры.
Корень вторичного строенияКорень – основной орган высшего растения. Он осуществляет функцию минерального и водного питания. Другая важная функция – закрепление, "заякоривание" растения в почве. Через корень растения поглощают из почвы воду и растворенные в ней ионы минеральных солей. В корнях осуществляется также биосинтез ряда вторичных метаболитов, в частности алколоидов. Корень способен к метаморфозам. Наиболее часто он является местом хранения запасных питательных веществ. Иногда выполняет роль дыхательного органа, может служить органом вегетативного размножения.
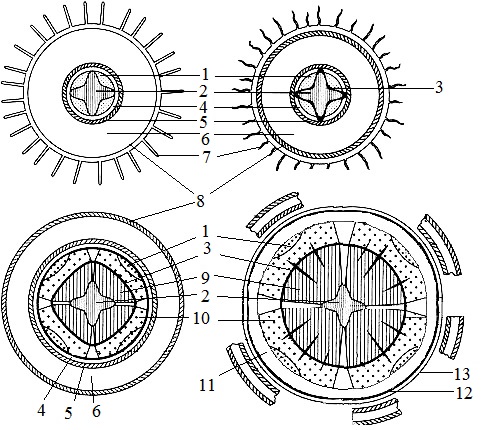
Рис. 2. Переход от первичного строения корня к вторичному
Образование вторичной структуры корня связано прежде всего с деятельностью камбия, который обеспечивает рост корня в толщину. Камбий вначале возникает из тонкостенных паренхимных клеток в виде разобщенных участков с внутренней стороны тяжей флоэмы между лучами первичной ксилемы. Камбиальную активность вскоре приобретают и некоторые участки перицикла, располагающиеся кнаружи от лучей первичной ксилемы. В результате образуется непрерывный камбиальный слой.
К центру камбий откладывает клетки вторичной ксилемы, а к периферии – клетки вторичной флоэмы. Клетки камбия, заложившегося в перицикле, образуют широкие радиальные светлые лучи паренхимы, располагающиеся между тяжами вторичной проводящей ткани. Эти лучи, иногда называемые первичными сердцевинными лучами, обеспечивают физиологическую связь центральной части корня с первичной корой. Позднее могут закладываться и вторичные сердцевинные лучи, "связывающие" вторичную ксилему и флоэму.
В результате деятельности камбия первичная флоэма оттесняется кнаружи, а "звезда" первичной ксилемы остается в центре корня. Ее "лучи" сохраняются долго, иногда до конца жизни корня. Помимо вторичных изменений, происходящих в центральном цилиндре, существенные перемены происходят и в первичной коре. Вследствие быстрого нарастания изнутри вторичных тканей, обусловливающего сильное утолщение корня, первичная кора нередко разрывается. К этому времени клетки перицикла, делясь по всей окружности осевого цилиндра, образуют широкую зону паренхимных клеток, во внешней части которой закладывается феллоген, откладывающий наружи пробку, а внутрь феллодерму. Пробка изолирует первичную кору от проводящих тканей, кора отмирает и сбрасывается. Клетки феллодермы и паренхима, сформированная за счет перицикла, в дальнейшем разрастаются и составляют паренхимную зону, окружающую проводящие ткани. Иногда эту зону называют вторичной корой. Снаружи корни двудольных, имеющие вторичное строение, покрыты перидермой. Корка образуется редко, лишь на старых корнях деревьев.
На ранних этапах развития стебель двудольных имеет первичное строение. В результате деятельности первичных меристем конуса нарастания формируются эпидерма, первичная кора, центральный цилиндр и сердцевина.
Эпидерма стебля имеет небольшое число устьиц, обычно покрыта кутикулой.
Под эпидермой находятся ткани первичной коры. Наружный слой ее часто образован колленхимой, которая располагается либо сплошным кольцом (подсолнечник), либо отдельными участками. Основная часть первичной коры сложена хлоренхимой и паренхимой. Здесь могут развиваться воздухоносные полости и вместилища выделений. Клетки внутреннего слоя первичной коры обычно заполнены крупными крахмальными зернами (крахмалоносное влагалище).
Центральный цилиндр состоит из перицикла и проводящей системы. Сердцевина - из паренхимы, которая часто разрушается, а на ее месте образуется воздухоносная полость. Перицикл представлен одним или несколькими рядами склеренхимы или отдельными ее тяжами. Проводящая система имеет вид полого цилиндра, который разделяет первичную кору и сердцевину. Проводящие пучки, составляющие этот цилиндр, разделены прослойками межпучковой паренхимы – сердцевинными лучами, которые связывают сердцевину и первичную кору. Большинству двудольных свойственен особый тип центрального цилиндра, на поперечном срезе которого пучки расположены кольцом (рис.) .
Рисунок. Стебель двудольных растений.
У двудольных растений слой клеток прокамбия между первичными флоэмой и ксилемой сохраняет способность к делению и превращается во вторичную образовательную ткань - камбий , который делится, откладывая внутрь элементы вторичной ксилемы, а к периферии - вторичной флоэмы. Пучки двудольных растений открытые, благодаря работе камбия пучок растет, диаметр его увеличивается. Деятельность камбия обеспечивает возникновение вторичных элементов в стебле, т. е. переход от первичного к вторичному анатомическому строению.
Вторичное строение характерно для всех двудольных и хвойных растений - трав и деревьев. Разнообразие типов строения обусловлено, прежде всего, расположением проводящих тканей, которое определяется заложением прокамбия и деятельностью камбия.
Прокамбий в процессе дифференциации конуса нарастания закладывается в виде тяжей. Если тяжи разделены достаточно широкими рядами паренхимы, то формируется пучковое или переходное строение стебля ; если тяжи прокамбия сближены настолько, что сливаются в цилиндр, то формируется непучковое (сплошное) строение .
Пучковое строение стебля встречается у некоторых травянистых растений (укроп, клевер, лютик, горох, люцерна). Заложенные в конусе нарастания прокамбиальные тяжи располагаются в один круг по периферии центрального цилиндра . Каждый прокамбиальный тяж превращается в открытый коллатеральный пучок, состоящий из первичной ксилемы, первичной флоэмы и полоски камбия между ними. Клетки камбия, делясь, дают новые (вторичные) элементы проводящего пучка: внутрь - ксилему к периферии - флоэму. Проводящие пучки разделены широкими межпучковыми зонами и окружают мощную сердцевину.
У большинства растений камбий работает активно и диаметр пучков значительно увеличивается Деятельность пучкового камбия стимулирует паренхиму, разделяющую пучки, которая начинает делиться, давая вторичную образовательную ткань. Пучковая и межпучковая меристемы смыкаются и образуют сплошное камбиальное кольцо , деятельность которого обеспечивает равномерное утолщение стебля. Если пучковый камбий дает элементы вторичных ксилемы и флоэмы, то межпучковый камбий производит паренхиму (у укропа, звездчатки злаковидной и др.). У клевера, люцерны и других растений межпучковый камбий откладывает преимущественно склеренхимоподобные клетки в сторону ксилемы. Эти клетки удлиненные, толстостенные одревесневающие. Поэтому задержка с уборкой клевера снижает качество сена. Отчетливое пучковое строение сохраняется у этих растений в течение всей жизни стебля.
Пучковое строение стебля может с возрастом смениться непучковым сплошным. Переходное строение стебля характерно для подсолнечника . Вначале стебель формируется так же, как у клевера, но межпучковый камбий откладывает не паренхиму, а ксилему и флоэму. Формируются добавочные проводящие пучки. Постепенно все пучки могут слиться в сплошной трехслойный (ксилема, камбий, флоэма) цилиндр. В верхней (молодой) части стебель имеет пучковое строение, у основания (в старой части) - непучковое.
Непучковое строение свойственно стеблям многих трав и деревьев. В конусе нарастания этих растений прокамбиальные тяжи настолько сближены, что образуют почти сплошной цилиндр (на поперечном срезе он имеет вид кольца). Прокамбиальный цилиндр (кольцо), дифференцируясь, дает к центру цилиндр первичной ксилемы, к периферии - цилиндр первичной флоэмы, а между ними образуется камбиальный цилиндр.
Вторичные изменения связаны с работой камбия. Клетки камбия делятся параллельно поверхности стебля, при этом внутрь откладывается в 10...20 раз больше клеток, чем наружу. Внутрь камбий формирует элементы вторичной ксилемы (древесины), наружу откладывается вторичная флоэма (луб). Местами камбий откладывает в обе стороны паренхимные клетки сердцевинных лучей.
Особенности непучкового строения стеблей трав можно рассмотреть на примере стебля льна . Стебель покрыт эпидермой, под которой располагается рыхлая хлоренхима первичной коры. Большое количество устьиц в эпидерме и отсутствие колленхимы в первичной коре имеют существенное практическое значение, так как во время технической мочки льна вода не встречает препятствий и легко проникает внутрь стебля. Клетки крахмалоносного влагалища (эндодермы) более крупные, содержат хорошо заметные крахмальные зерна.
Примыкающий к эндодерме центральный цилиндр начинается группами склеренхимных клеток – лубяных волокон, между которыми расположены клетки паренхимы. Лубяные волокна возникают в перицикле и в первичной флоэме. Они отличаются очень толстыми, обычно неодревесневшими стенками, состоящими на 75...90 % из целлюлозы. Созревшая клетка лубяного волокна мертвая. Протопласт после утолщения клеточной стенки разрушается. Лубяные волокна обладают необыкновенно высокой прочностью. Сопротивление разрыву у них соответствует сопротивлению стали, а упругость выше. Клетки лубяного волокна очень тесно соединены между собой. Льняное волокно - великолепное прядильное сырье, используемое в текстильной промышленности.
Далее в последовательном порядке располагаются цилиндры (кольца): вторичной флоэмы, камбия, вторичной ксилемы с хорошо заметными сердцевинными лучами. Первичная ксилема расположена участками, примыкающими к сердцевине. Паренхима сердцевины быстро разрушается (в процессе удлинения стебля), образуя центральную полость.
Стебли двудольных травянистых растений характеризуются:
1. вторичным строением, очень рано возникающим вслед за первичным;
2. наличием первичной покровной ткани - эпидермы;
3. развитой первичной корой, делящейся на колленхиму (экзодерма), паренхиму (мезодерма) и крахмалоносное влагалище (эндодерма);
4. пучковым или непучковым (сплошным) строением;
5. правильным расположением коллатеральных или биколлатеральных проводящих пучков в один круг по периферии центрального цилиндра;
6. наличием камбия (пучки открытые).
Метаморфозы побега.
Если дополнительные функции начинают преобладать над главными, или орган в процессе развития приобретает какие-то новые функции, он очень сильно изменяется и морфологически, и анатомически. Такие видоизменения органов называют метаморфозами (от греч. metamorphosis - превращение, преобразование). Органы, имеющие одинаковое происхождение, но выполняющие разные функции называют гомологичными . Гомологичными органами являются корневища, клубни, луковицы, колючки боярышника, кладодии, филлокладии, усики винограда и тыквы, т.к. все они исторически развились из листостебельного побега.
Органы, выполняющие одинаковые функции и сходные морфологически, но имеющие разное происхождение, называют аналогичными . Аналогичны корневые шишки и побеговые клубни, побеговые колючки боярышника и листовые колючки барбариса, листовые усики гороха и побеговые усики тыквы.
Видоизменения подземных побегов.
В природе широко распространены подземные побеги: корневища, клубни, клубнелуковицы, луковицы. Они участвуют в запасе веществ и вегетативном размножении растений.
Корневище представляет собой побег с чешуевидными листьями, почками и придаточными корнями. На поверхности корневища хорошо заметны рубцы от прошлогодних низовых листьев и отмерших побегов. Корневище имеет много придаточных корней. Обычно корневища подземные. Почки, развивающиеся в надземные побеги. Корневища характерны для пырея, ириса, кувшинки.
Столон - удлиненный тонкий побег с недоразвитыми листьями. В отличие от корневища он обычно недолговечен. У мускусницы, кислицы столоны участвуют в накоплении питательных веществ, откладывающихся в сильно утолщающихся низовых листьях, но основная функция столона - участие в вегетативном размножении.
Клубень в отличие от корневища и столона сильно укорочен и утолщен. Запасные вещества в нем локализуются в стеблевой паренхиме.
Клубни могут развиваться на корневищах, столонах, главном побеге и других частях растений. Они могут быть подземными и надземными.
Мощные корневищные клубни с большим количеством крахмала характерны для таро, или колоказии съедобной из семейства ароидных, которую культивируют в тропических и субтропических странах.
У картофеля клубни формируются на конце подземных столонах - беловатых хрупких побегах с чешуевидными листьями, развивающихся из пазушных почек, находящихся в основании олиственных побегов. На поверхности клубня располагаются пазушные почки - глазки , сидящие в небольших ямках, обрамленных с одной стороны бровками - остатками оснований листьев.
Клубни топинамбура , или земляной груши , тоже образуются на столонах. Проводящие ткани в них слабо развиты. Клубни богаты инулином, который откладывается преимущественно в стеблевой паренхиме.
Клубни, формирующиеся на столонах и корневищах, недолговечные. Они сохраняются только один год, между двумя вегетационными периодами, отмирая после развития из их почек олиственных побегов, потребляющих питательные вещества до тех пор, пока не образуются корни, и растения не перейдут к самостоятельному питанию.
Луковица , как и клубень, представляет собой специализированный, видоизмененный укороченный побег, служащий не только для хранения питательных веществ, но и для перенесения неблагоприятных периодов года, вегетативного возобновления и размножения растений. Луковицы свойственны однодольным растениям из семейств лилейных, амариллисовых и других. Как исключение, они встречаются у двудольных: некоторых видов кислицы и жирянки.
Стебель, называемый донцем, в луковице сильно редуцирован и более или менее уплощен. Он несет листья в виде чешуй, а в нижней части - придаточные корни. В отличие от корневища и клубня запасающими органами луковицы служат мясистые чешуи. Луковицы образуются у тюльпана, рябчика нарцисса, гиацинта, пролески, подснежника и других растений.
В зависимости от развития боковых почек луковицы бывают простыми , когда почки невелики (лук, лилия и др.), и сложными – почки в пазухах сухих чешуй крупные, сильно развитые (чеснок).
Клубнелуковица - подземное видоизменение побега, сочетающее признаки корневища и луковицы. Она развивается из побега с сильно укороченным стеблем, от нижней части которого отходят придаточные корни. На верхушке клубнелуковицы или в ее основании находится почка, из которой образуется цветоносный побег. Стебель клубнелуковицы покрыт основаниями отмерших прошлогодних листьев, имеющих вид сухих пленчатых чешуй. Из почек, находящихся в их пазухах, развиваются новые клубнелуковицы - детки. Клубнелуковицы имеют шафран, гладиолус, безвременник.
Видоизменения надземных побегов
Кочан капусты представляет собой, по существу, почку гигантских размеров: среднюю часть кочана занимает мясистый стебель (кочерыга ), вдоль которого расположены сочные мясистые листья. Наиболее мелкие и молодые из них находятся внутри кочана, вокруг конуса нарастания, которым заканчивается кочерыга. У брюссельской капусты образуются мелкие кочаны из боковых почек.
В кочане оба органа побега участвуют в накоплении питательных веществ: водорастворимых сахаров и других биологически ценных соединений, в том числе витаминов, но основная роль принадлежит листьям.
Надземные клубни , развивающиеся из нижней части главного побега, характерны для капусты кольраби.
Мелкие клубеньки могут развиваться и в области соцветия. У горца живородящего, например, они образуются в пазухах нижних кроющих листьев соцветия. Их почки начинают прорастать уже на материнском растении. После опадения они укореняются, образуя новые растения.
Метаморфизированные побеги могут служить не только для хранения питательных веществ, воспроизведения и размножения растений, но выполнять и другие функции.
У некоторых растений побеги в процессе развития теряют листья, и фо-тосинтезирующим органом становится стебель, называемый в этом случае кладодием . Обычно он уплощен и обладает способностью к длительному росту (у тропического растения мюленбекии, или гомалокладуса, из семейства гречишных, опунция). В узлах хорошо выражены перетяжки, поэтому кладодий выглядит членистым. Во влажных условиях кладодии могут быть олиственными, но в сухую погоду листья опадают. Как орган фотосинтеза кладодий имеет хорошо развитую хлорофиллоносную ткань, расположенную под эпидермой.
Кладодии, по внешнему виду сходные с листьями, называют филлокладиями . Они развиваются из боковых почек, поэтому всегда находятся в пазухе небольшого пленчатого или чешуевидного листа. Филлокладии очень разнообразны. У спаржи перистой, или щетинистой, они мелкие, игольчатые, у спаржи Шпренгера, или густоцветковой, - уплощенные, линейные. Широколанцетные или яйцевидные в очертаниях кожистые филлокладии характерны для видов иглицы, встречающихся в Средиземноморье, Закавказье, Крыму. Летом на нижней стороне филлокладия, в его средней части, развивается соцветие, расположенное в пазухе мелкого кроющего листа, осенью и зимой иглица цветет, а весной на филлокладиях созревают оранжевые ягоды.
Довольно распространенный тип изменения побегов - колючка . Это сильно одревесневающий безлистный укороченный побег с острой верхушкой. У боярышника побег, развивающийся весной в пазухе листа, сначала имеет мелкие листья и верхушечную почку. Вскоре верхушечный рост прекращается, листья опадают, стебель сильно одревесневает и превращается в колючку. Так же развиваются колючки у терна.
Цепляющиеся растения снабжены усиками , которые, как и колючки, представляют собой у некоторых растений видоизмененные боковые побеги. У пассифлоры усики простые, неветвистые, с закрученной верхушкой. Они находятся в пазухах листьев, как и цветки. Ветвистые усики винограда представляют собой видоизмененные соцветия. У девичьего винограда окончания разветвленных усиков дисковидно расширены и играют роль присосок, помогающих растению взбираться на опору. Усики побегового происхождения образуются также у тыквы, арбуза, огурца и других представителей семейства тыквенных.