Стебель. Морфология и функции стебля. Стебель - удлинённый побег высших растений, служащий механической осью, также выполняет функцию проводящей и опорной базы для листьев, почек, цветков.Стебли выполняют две основные функции - несущую и ведущую. Стебли соединяют все части побега в единую систему. Благодаря стеблю обеспечивается увеличение поверхности растения путем его ветвления, образования и выгоднее расположение листьев и генеративных органов. Стебель обеспечивает восходящий и нисходящий движение воды и растворенных в ней веществ. Молодые побеги выполняют функцию фотосинтеза. Видоизмененные стебли выполняют еще и дополнительные функции, например, накопления и хранения питательных веществ в многолетних стеблях, а также запасания воды в специализированных стеблях, зацепки других растений, защита, обеспечение вегетативного размножения растений.
Стебли растений весьма разнообразны и отличаются по многим признакам. По консистенции стебли бывают: травянистыми или деревянистые, пустотелыми (соломинка) или наполненными (с сердцевиной).По форме поперечного сечения стебли бывают округлые или цилиндрические (в большинстве видов растений), четырехгранные (виды семей Глухокропивови, норичниковых), трехгранные (виды семейства осоки),многогранные (виды лебеды, щавеля), сплющенные (например, мятлик сжатый, ситник членистый). По характеру роста и размещением в пространстве выделяют следующие виды стеблей: прямостоячие (крепкие, преимущественно ортотропные стебли с хорошо выраженными междоузлие, большинство видов высших растений) ползучие (преимущественно плагиотропни стебли укореняются в узлах, с хорошо выраженными междоузлие; например, живучка ползучая, земляники, лютик ползучий, лапчатка гусиная) вьющиеся (закручиваются вокруг опоры по часовой стрелке или наоборот; например, хмель обыкновенный, вьюнок полевой, повилика, фасоль обыкновенная) лазящие или цепкие (стебли с плохо развитой механической тканью зацепляются с помощью усиков; например, огурцы, тыква обыкновенная, горох посевной) стелющиеся (стебли с довольно плохо развитой механически тканью, стелющиеся по поверхности почвы: например, грыжник голый, мшанки лежащая, зверобой стелющийся, стелюшок) восходящие (нижняя часть стебля стелется по поверхности почвы, верхняя - направлена вертикально; например тимьян, звездчатка, вероника лекарственная) укороченные (стебель плохо выраженное, с очень сближенными узлами, короткими междоузлие и собранными в прикорневую розетку листьями, образует цветочную стрелку; например, подорожник, одуванчик, лилейники, подснежник).Видоизменения стеблей.
Видоизмененный стебель является органом, предназначенным для накопления запасных питательных веществ, которые необходимы растению, чтобы пережить период покоя. Кроме того, видоизмененные стебли нередко участвуют в размножении и распространении растений. Чтобы быть отнесенным к данной группе, видоизмененный стебель должен обладать всеми признаками обычного стебля, то есть иметь характерное для него строение, что означает наличие собственно стебля с верхушечной точкой роста и листьями, располагающимися по спирали (очередно или супротивно) и несущими в своих пазухах почки. Отличие видоизмененного стебля отнормального состоит в том, что он не обязательно растет над землей, рост его часто бывает более или менее горизонтальным и обычно он является вместилищем запасных питательных веществ.
Существует шесть основных типов видоизмененных стеблей: клубень корневище клубнелуковица луковица ус отпрыск
Возникновение первичных тканей и первичное анатомическое строение. Анатомическое строение стебля растений обусловлено его главными функциями. Для него характерно развитие механической и проводящей тканей растений. Кроме того, для стебля характерна сложная система меристем - верхушечные, боковые и вставочные, определяющие нарастание его в течение длительного времени и возникновение новых органов.
Стебель растения возникает из апикальной меристемы, из которой дифференцируются три слоя тканей: покровная, проводящая, основная. Сначала эти ткани представлены так называемой первичной меристемой (промеристемой), состоящей из протодермы, прокамбия и основной меристемы.Первичные проводящие ткани развиваются из прокамбия. Первые элементы флоэмы дифференцируются из наружных, расположенных к периферии клеток прокамбия. Первичная флоэма представлена тонкостенными недолговечными удлиненными клетками и носит название протофлоэмы, а наружные клетки ее могут быть представлены механическими волокнами.Первичные элементы ксилемы - трахеиды, реже сосуды с кольчатыми и спиральными утолщениями стенок - возникают позже из внутренних клеток прокамбия и определяются в целом как протоксилема. В ее состав помимо проводящих элементов входят паренхимные клетки.Позже внутрь от протофлоэмы дифференцируется имеющая более или менее типичное для флоэмы строение. Наружу от протоксилемы формируется метаксилема, состоящая из трахеид или трахей с более утолщенными одревесневшими стенками.
Таким образом, за счет деятельности прокамбия и остальной меристемы апекса возникает первичное строение стебля растения. У однодольных растений весь прокамбий дифференцируется в элементы первичных проводящих тканей. Стебли однодольных, особенно травянистых (злаков), по сравнению со стеблями двудольных растений имеют более простое строение, для них характерно в основном первичное строение.У двудольных растений в средней части прокамбиального тяжа происходит образование камбия и начинается образование вторичных проводящих тканей (метафлоэмы и метаксилемы), объем которых увеличивается за счет деления клеток камбия.Пучки, состоящие только из первичных тканей, как у однодольных, закрытые-, пучки с камбием, характерные для двудольных, - открытые.Для травянистых однодольных растений характерно диффузное распределение проводящих пучков. Проводящие пучки закрытые, коллатеральные, реже концентрические. Из механических тканей наиболее развита склеренхима, колленхима встречается у немногих растений. Вторичного утолщения у травянистых однодольных нет.
Округлые (например: подсолнечник)
Четырехгранные
Трехгранные
Многогранные (ребристые)(например: тыква)
Крылатые (имеют кожистые выросты)
Формирование стебля
Стебель, как и корень, формируется из первичной верхушечной меристемы. В отличие от корня конус нарастания защищен от окружающей среды молодыми развивающимися листочками.Как и в корне клетки меристемы делятся митозом и специализируются в первичные постоянные ткани, но в отличие от корня в стебле нет четко выраженных зон роста. Специализация клеток происходит очень быстро, и так же как и в корне эти клетки специализируются в трех таких же участках.
С периферии 1 слой клеток превращается
в первичную покровную ткань; в первичную
кору; в центре возникает центральный
осевой цилиндр. В стебле, в отличие от
корня центральный цилиндр во много раз
занимает больше места, чем первичная
кора.
Первичная покровная ткань стебля эпидермис, 1 слой, с устьицами, с волосками. Его функция –– защита нижележащих структур.
Первичная кора стебля отвечает за фотосинтез и придает механическую устойчивость (сопротивляется от изгиба). Определяет стебель в пространстве. Она состоит из:
1) хлоренхима (от 1 до множества слоев этих клеток)
2) колленхима (от 1 и более слоев). У однодольных встречается редко.
3) эндодерма
Эндодерма стебля представляет собой 1 слой живых клеток, с целлюлозными оболочками, содержащие крахмальные зерна. Они не являются запасающим веществом, они помогают ориентироваться в пространстве, и поэтому эндодерма стебля называется крахмалоносным влагалищем. Кроме того, у различных растений в первичной коре могут формироваться вместилища выделений, могут встречаться кристаллоносные клетки, это пластическая часть стебля.
Центральный осевой цилиндр он начинается с перециклической склеренхимы. Перицикл в стебле перерождается в склеренхиму и частично в паренхиму. Перициклическая склеренхима может быть на поперечном разрезе в виде сплошного кольца или в виде отдельных тяжей.
За перициклом расположены проводящие пучки, обычно коллотыральные; редко биколлотыральные окруженные клетками основной паренхимы.
Расположение проводящих пучков различно у однодольных и двудольных растений.
–– у однодольных расположено диффузно (рассеянно)
–– у двудольных по кругу
Такое различное расположение проводящих пучков связано с характером объединения проводящих пучков листа и стебля
У двудольных проводящие пучки листа
соединяются со стеблем в своем узле.![]()
У однодольных листовые пучки заходят
в центральный цилиндр изгибаются и
соединяются со стеблевыми пучками
проходя через 1-2 узла.
У двудольных из листа в стебель заходит 2-3 листовых пучка, а у однодольных в связи с параллельным жилкованием - множество.
У однодольных растений все проводящие пучки закрытые , т.е. не имеют камбия, а у двудольныхоткрытые пучки, т.е. имеется камбий.
Толщина стебля однодольных растений по всей длине остается одинаковой, а у двудольных верхняя часть стебля немного меньше, чем в нижней части.
Формирование тканей стебля вторичного строения
Вторичное строение стебля характерно для однолетних и многолетних травянистых, древесных двудольных, а также голосеменных растений. У двудольных растений первичное строение очень недолговечно, и с началом деятельности камбия образуется вторичная структура. В зависимости от закладки прокамбия формируется несколько типов вторичного строения стебля. Если тяжи прокамбия разделены широкими рядами паренхимы, то формируется пучковое строение, если они сближены так, что сливаются в цилиндр, - формируется непучковое строение.
Рис. 3.24. Пучковый тип строения стебля двудольного растения: А - клевер: 1 - эпидерма; 2 - хлоренхима; 3 - склеренхима перициклического происхождения; 4 - флоэма; 5 - пучковый камбий; 6 - ксилема; 7 - межпучковый камбий
Пучковое строение стебля встречается у таких растений, как клевер, горох, лютик, укроп (рис. 3.24). У них прокамбиальные тяжи закладываются в один круг по периферии центрального цилиндра. Каждый прокамбиальный тяж превращается в коллатеральный пучок, состоящий из первичной флоэмы и первичной ксилемы. В дальнейшем между флоэмой и ксилемой из прокамбия закладывается камбий, формирующий элементы вторичной флоэмы и вторичной ксилемы. К периферии органа откладывается флоэма, а к центру - ксилема, причем ксилемы откладывается больше. Первичные флоэма и ксилема остаются на периферии пучка, а вторичные элементы примыкают к камбию. Для стеблей двудольных растений характерно формирование открытых колла- теральных или биколлатеральных пучков (рис. 17, см. цв. вкл.).
Также для стеблей двудольных растений характерна дифференциация первичной коры, в состав которой входят: колленхима (уголковая (рис. 18, см. цв. вкл.) или пластинчатая), хлорофиллоносная паренхима и внутренний слой - эндодерма. В эндодерме накапливается крахмал; такое крахмалоносное влагалище играет важную роль в геотропической реакции стеблей. На границе первичной коры в центральном осевом цилиндре располагается перициклическая склеренхима - сплошным кольцом или участками в виде полудуг над флоэмой. Сердцевина стебля выражена и представлена паренхимой. Иногда часть сердцевины разрушается с образованием полости (см. рис. 3.24).
Непучковое строение характерно для древесных растений (липа) (рис. 19, см. цв. вкл.) и многих трав (лен). В конусе нарастания прокамбиальные тяжи сливаются и образуют сплошной цилиндр, видимый на поперечном срезе в виде кольца. Кольцо прокамбия кнаружи формирует кольцо первичной флоэмы, а внутрь - кольцо первичной ксилемы, между которыми закладывается кольцо камбия. Клетки камбия делятся (параллельно поверхности органа) и наружу откладывают кольцо вторичной флоэмы, а внутрь - кольцо вторичной ксилемы в соотношении 1:20. Непучковое строение рассмотрим на примере многолетнего древесного стебля липы (рис. 3.25).
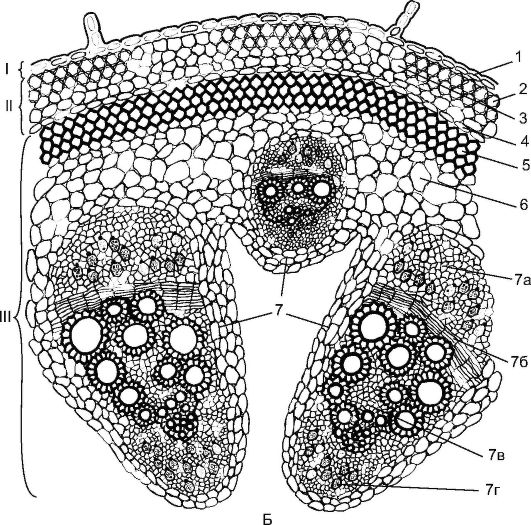
Рис. 3.24. (продолжение) Б - тыква: I - покровная ткань; II - первичная кора; III - центральный осевой цилиндр; 1 - эпидерма; 2 - уголковая колленхима; 3 - хлоренхима; 4 - эндодерма; 5 - склеренхима; 6 - основная паренхима; 7 - биколлатеральный сосудисто-волокнистый пучок: 7а - флоэма; 7б - камбий; 7в - ксилема; 7г - внутренняя флоэма
Молодой побег липы, образовавшийся весной из почки, покрыт эпидермой. Все ткани, лежащие до камбия, называют корой. Кора бывает первичная и вторичная. Первичная кора представлена пластинчатой колленхимой, располагающейся сразу под эпидермой сплошным кольцом, хлорофиллоносной паренхимой и однорядным крахмалоносным влагалищем. В этом слое находятся зерна «оберегаемого» крахмала, которые растение не расходует. Считается, что этот крахмал участвует в поддержании растением равновесия.
Центральный осевой цилиндр у липы начинается с перициклической склеренхимы над участками флоэмы. В результате деятельности камбия возникает вторичная кора (от камбия до перидермы), пред- ставленная вторичной флоэмой, сердцевинными лучами и паренхимой вторичной коры. Кору с липы заготавливают, снимая до камбия, особенно легко это делать весной, когда клетки камбия активно делятся. Раньше кору липы (лыко) использовали для плетения лаптей, изготовления коробов, мочалок и др.
Трапециевидная флоэма разделена треугольными первичными сердцевинными лучами, пронизывающими древесину до сердцевины. Состав флоэмы у липы неоднороден. В ней имеются составляющие твердый луб одревесневшие лубяные волокна, а мягкий луб представлен ситовидными трубками с клетками-спутницами и лубяной паренхимой. Луб теряет способность проводить органические вещества обычно через год и обновляется новыми слоями за счет деятельности камбия.
Камбий образует и вторичные сердцевинные лучи, но они не доходят до сердцевины, теряясь во вторичной древесине. Сердцевинные лучи служат для продвижения воды и органических веществ в ра- диальном направлении. В паренхимных клетках сердцевинных лучей к осени откладываются запасные питательные вещества (крахмал, масла), расходуемые весной на рост молодых побегов.
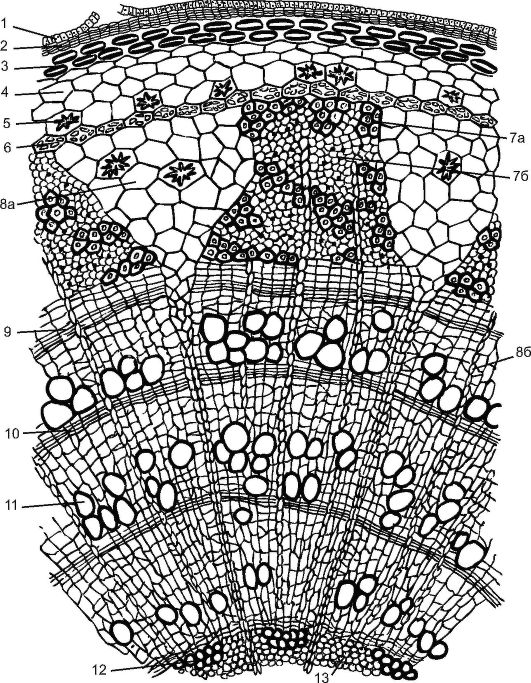
Рис. 3.25. Поперечный разрез трехлетней ветви липы: 1 - остатки эпидермы; 2 - пробка; 3 - пластинчатая колленхима; 4 - хлоренхима; 5 - друзы; 6 - эндодерма; 7 - флоэма: 7а - твердый луб, (лубяные волокна); 7б - мягкий луб - (ситовидные трубки с клетками-спутницами и лубяная паренхима); 8а - первичный сердцевинный луч; 8б - вторичный сердцевинный луч; 9 - камбий; 10 - осенняя древесина; 11 - весенняя древесина; 12 - первичная ксилема; 13 - паренхима сердцевины
Уже летом под эпидермой закладывается феллоген и формируется вторичная покровная ткань - перидерма. К осени, с образованием перидермы, клетки эпидермы отмирают, но их остатки сохраняются в течение 2-3 лет. Наслоение многолетних перидерм формирует корку.
Слой ксилемы, образуемый камбием, у древесных растений значительно шире, чем слой флоэмы. Древесина функционирует в тече- ние нескольких лет. Отмершие же клетки древесины не участвуют в проведении веществ, но способны поддерживать колоссальную тяжесть кроны растения.
Состав древесины неоднороден, в нее входят: трахеиды (рис. 20, см. цв. вкл.), трахеи, древесинная паренхима и либриформ. Древесина характеризуется наличием годичных колец. Ранней весной, когда в расте- нии возникает активное сокодвижение, камбий в ксилеме формирует широкопросветные и тонкостенные проводящие элементы - сосуды и трахеиды, а с приближением осени, когда эти процессы замирают и деятельность камбия ослабевает, возникают узкопросветные толстостенные сосуды, трахеиды и древесинные волокна. Таким образом, образуется годичный прирост, или годичное кольцо (от одной весны до другой), хорошо различимое на поперечном срезе. По годичным кольцам можно определить возраст растения (см. рис. 3.25).
Особенности строения стебля двудольных:
1) рост стебля в толщину (за счет деятельности камбия);
2) хорошо дифференцированная первичная кора (колленхима, хлорофиллоносная паренхима, крахмалоносная эндодерма);
3) биколлатеральные и коллатеральные пучки только открытого типа (с камбием);
4) сосудисто-волокнистые пучки расположены по кольцу или сливаются (непучковое строение);
5) наличие сердцевины;
6) для древесных растений характерно наличие в ксилеме годичных колец.
Особенности строения корневищ двудольных. Покровной тканью корневищ двудольных может быть эпидерма, а у многолетних корневищ эпидерма сменяется перидермой. Первичная кора представлена запасающей паренхимой и эндодермой с пятнами Каспари. Причем ширина первичной коры приближается к ширине центрального цилиндра. Строение центрального осевого цилиндра, сосудисто-волокнистых пучков и их расположение в нем имеют те же особенности, что и для надземных стеблей.
В зависимости от экологических условий, в которых формируется тот или другой вид растения, создаются различные особенности структуры его органов, в том числе и стебля.
Кроме того, стебель в ряде случаев выполняет функции других органов. В этом случае изменения структуры стебля особенно существенны и обычно причисляются к разряду аномалий. Своеобразным строением стебля характеризуются лианы. Особенно богато представлены лианы в тропиках, где они наиболее разнообразны и мощно развиты. Но и в умеренных широтах произрастает достаточно лиан. Хмель, ломонос, плющ, виноградная лоза, горох, фасоль, глициния, вьюнок и др. - настоящие лианы, не столь роскошные, как в тропиках, но все же имеющие в структуре стебля черты, присущие всем лианам.
Наиболее характерной чертой структуры стебля лиан являются довольно резко выраженная расчлененность проводящей системы и сильное развитие паренхимных сердцевинных лучей. Даже у лиан с прекрасно выраженным древесным стеблем (виноградная лоза) сердцевинные лучи состоят из паренхимы, мало отличающейся от паренхимы коры и сердцевины, причем эти сердцевинные лучи очень высоки, тянутся на большом расстоянии вдоль длины стебля, напоминая длинные, вертикально поставленные вдоль стебля ленты, рассекающие массив центрального цилиндра на отдельные секторы. У типичных древесных растений сердцевинные лучи вследствие незначительности их высоты на тангентальных разрезах древесины имеют вид коротких веретеноподобных скоплений клеток. Еще резче расчленение центрального цилиндра стебля выявляется у тропических лиан. У некоторых тропических лиан разбросанные в различных местах древесины центрального цилиндра паренхимные клетки начинают усиленно делиться и разрастаться, раздвигая отдельные участки сосудистой системы. Вследствие этого старые стебли таких лиан отличаются очень неправильным и прихотливым строением. В общем же строение стебля древесных лиан напоминает строение стебля некоторых травянистых растений.
У ряда растений, произрастающих в пустынных местностях Африки и Америки, где вообще дожди выпадают очень редко и то в течение короткого времени года, форме и структуре стебля присущи особые специфические черты. У таких растений стебли мясистые, зеленые, принимают различную форму - шара, пластинки или снабжены широкими пластинчатыми ребрами. Такие стебли выполняют функцию листьев. Листья часто очень мелкие и иногда имеют вид сухих колючек. Растения с мясистыми стеблями и редуцированными листьями называют стеблевыми суккулентами . К числу их относятся различные виды опунций, кактусов, молочаев.
Редукция листовых пластинок до сухих колючек ведет к значительному уменьшению испаряющей поверхности всего растения, состоящего в своей надземной части целиком из своеобразно видоизмененного стебля. Самый стебель превращается в орган, приспособленный для накопления запасов воды. Это дает возможность растению запасать влагу в дождливые периоды и медленно расходовать ее в пору бездождья.
В строении покровных тканей большинства типов суккулентных стеблей существуют приспособления к возможному понижению потери растением воды через транспирацию. При этом должна быть сильно редуцирована и система проводящих тканей. Механические ткани внутри суккулентных стеблей становятся ненужными и почти отсутствуют.
Клеточный сок клеток тканей суккулентных стеблей состоит из растворов различных кислот, сахаров и слизистых веществ. Эти вещества способствуют понижению отдачи воды растением. Ткань суккулентных стеблей очень однородна и состоит преимущественно из тонкостенной паренхимы, пронизанной слаборазвитыми сосудистыми пучками: сосудов в каждом пучке немного, все они узкополостные и примитивной структуры. Слабое развитие сосудистых пучков вполне соответствует незначительному развитию листьев, так как лист в своем развитии стимулирует образование сосудисто-волокнистых пучков стебля. В типичных, хорошо выраженных суккулентных стеблях вторичной ксилемы образуется мало вследствие слабой камбиальной деятельности, сосудистая система состоит преимущественно из первичных элементов, возникших в васкулярной меристеме. Следовательно, и физиологически, и с точки зрения истории развития формирование слабой проводящей системы в стеблях суккулентных растений вполне объяснимо.
Естественно, что среди суккулентов нет древесных форм, все они по своей структуре - травянистые растения.
У растений так называемого склеренхимного типа , тоже произрастающих в засушливых местах, развиты нормальные листья и стебли, проводящая система слагается из мощно развитых сосудисто-волокнистых пучков или продуктов их слияния. Сосудистые элементы в таких пучках широкополоетны, многочисленны и более высоко организованы, чем у суккулентов. Вторичные элементы у древесных растений этой группы выражены отчетливо, а в стеблях травянистых растений резко выражено одревеснение оболочек клеток основной паренхимы. Механическая система развита сильно, анатомические элементы ее толстостенны.
Стебли растений, произрастающих в воде, - так называемых водных растений - при относительно богатом разнообразии форм обладают одним общим структурным признаком: у них хорошо развита система межклетных ходов и межклетников. Межклетники в виде более или менее больших полостей образуют систему воздушных ходов, пронизывающих как.сердцевину, так и кору стебля. При этом особенно сильно развита сердцевина, занимающая значительную долю общего поперечника стебля. У водных растений, особенно среди представителей класса однодольных, механическая система иногда преобладает над проводящей системой. Группы сосудистых анатомических элементов, собранных в пучки, обычно бывают сравнительно незначительных размеров, массивы же, состоящие из механических элементов, сосредоточенные преимущественно по периферии стебля, достигают по сравнению с сосудистыми пучками большой мощности.
Еще резче отклоняется от нормального типа строение подземных стеблей, выполняющих функцию вместилищ запасов, - корневищ и клубней. Корневище является многолетним побегом и обычно достигает значительно большей толщины, чем надземные побеги, где бы они ни произрастали, - в воде или иной среде. В связи с основным назначением корневища - служить хранилищем запасных веществ - паренхиматизация его тканей представляет собой наиболее характерную черту структуры. Запасающей тканью в типичных корневищах служит сердцевина, состоящая из паренхимных округлых клеток, тонкостенных, с небольшими межклетниками между ними. По своей массивности сердцевина в корневищах преобладает над прочими тканями. Механические и проводящие ткани, уступая по мощности сердцевине и коре, в корневище выражены слабее, чем в надземных побегах.
Структура корневища как многолетнего побега слагается под воздействием разнообразных формирующих факторов внешней среды. Влияние этих факторов может быть как непосредственным, так и действующим через корреляцию в развитии органов растения: так, степень развития листьев, образование цветочных побегов, откладывание запасных веществ влияют на структуру корневища. Особенно заметна зависимость строения корневища от степени развития на них листьев.
Во время подземного роста, когда на корневище находятся лишь чешуеподобные мелкие листочки, соответственно формируется очень слабая проводящая система, в ксилемной части состоящая из редких узкополостных сосудов примитивной структуры. После того как начнет развиваться надземный плодоносящий побег с нормальными большими листьями, усиливается активность камбия и вследствие этого развивается древесина, а также увеличивается число и просвет сосудов, которые имеют более совершенную структуру по сравнению с образовавшимися до начала роста надземного побега.
Своеобразной морфологической особенностью корневища являются очень короткие междоузлия. По-видимому, незначительность длины междоузлий находится в тесной корреляции со слабой дифференциацией листьев на корневище.
На корневище по своей морфологии в некоторых отношениях похоже другое видоизменение подземного побега - клубень. Основное различие между корневищем и клубнем заключается в продолжительности существования: корневище - многолетний побег, клубень же обычно существует один-два года. Поэтому в клубне выявлены только те структурные черты, которые корневища имеют на первой стадии формирования своих тканей. Преобладание паренхимы в клубне еще более отчетливо, чем в корневище.
Степенью паренхиматизации ткань клубня напоминает ткань стебля суккулентных растений. Так же, как в стеблях суккулентных растений, вся масса ткани клубня за исключением покровной (перидермы) представляет собой запасающую ткань, пронизанную слаборазвитыми и примитивно построенными пучками проводящей системы. Наряду с редукцией проводящей ткани в нормальном клубне обычно совершенно не образуется механической ткани. Ткани, слагающие клубень, однообразно построены и приспособлены к выполнению только одной функции - отложению запасов.

Типичным примером структуры клубня может быть клубень картофеля. На рисунке изображен фрагмент поперечного разреза клубня одного из сортов картофеля («Вольтман»), из центральной части его. Для удобства демонстрации тканевой структуры крахмал, в изобилии заполняющий все клетки, не изображен. Паренхима клубня лишь в одном месте рассекается узкой полоской более мелких клеток, состоящей из четырех групп. В каждой группе находится 1-3 узкополостных сосуда, остальная ткань состоит из меристемоподобных клеток, среди которых находится несколько ситовидных трубок. Сосуды, отличающиеся слаборазвитой структурой, примитивны, причем некоторые из них находятся в состоянии облитерации и разрушения.
Такая ткань, состоящая из примитивных и слаборазвитых сосудистых элементов, расположена кольцом по всему клубню, представляя собой центральный цилиндр. Кнаружи расположена первичная кора, ковнутри - сердцевина. В клубнях, имеющих только чешуевидные подобия листьев, вторичных тканей в результате деятельности камбия не образуется.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter .