Стъбло - това е аксиалната структура на издънката. А анатомичната му структура следва от основните й функции. Първо, стъблото се характеризира с развитието на механични и проводими тъкани. На второ място, стъблото има сложна системамеристеми: апикални, странични и интеркаларни, които обуславят растежа му за дълго време, както и появата на нови органи. Стъблото възниква от апикалната меристема, от която се диференцират три слоя тъкани: основен, покривен и проводящ.
Структурата на едносемеделните
Целият прокамбий или първична меристема едносемеделнисе диференцира в елементи на първични проводящи тъкани. Техните стъбла, особено ако вземем предвид тревисти (житни), имат по-проста структура в сравнение със стъблата двусемеделни растения. Те също се характеризират главно първична структура. Едносемеделните имат съдово-влакнести съдови снопове затворен (без камбий), се състоят само от първични тъкани и са произволно разположени в основния паренхим на стъблото.

Устройството на двусемеделните растения
В средната част на прокамбиалната връв при двусемеделните растения се образува камбий и започва образуването на вторични проводими тъкани (метафлоем и метаксилем). Обемът на тези вторични проводящи тъкани се увеличава поради деленето на камбиевите клетки, което в крайна сметка води до силно удебеляване на стъблото.
Двусемеделните се характеризират с отворени туфи с камбий. Проводими тъкани в стъблата на двусемеделните растения подредени в пръстеноколо сърцевината. Сноповете са разделени от медуларни лъчи, които се състоят от паренхим и свързват медулата или с перицикъла, или с първичната кора. По периферията в стъблата на двусемеделните растения има механични тъкани, докато склеренхиме част от перицикъла, коленхим, заедно с основния паренхим, е част от първичната кора.
Структурата на централния цилиндър на двусемеделните растения може да бъде лъчИ нелъчеви.
По своята анатомична структура едногодишните стъбла на дървесните растения са подобни на стъблата на тревните растения: техните проводящи системи имат негредова конструкция. Тяхната отличителна чертае активна работакамбий и ранно образуване на вторична покривна тъкан - задръствания.
Благодарение на дейността на камбия в стъблото се образуват различни елементи от вторичен произход, докато първичните елементи постепенно изчезват. Камбият образува ксилемни и флоемни елементи с различна скорост: за една флоемна клетка камбият отделя няколко ксилемни клетки. В резултат на това ксилемата (или дървесината) расте много по-бързо от флоема (лика); съответно почти цялата маса на ствола и клоните на дървото пада върху ксилемата. Флоемата, напротив, съставлява сравнително тънък слой от вторичната кора.

Непрекъснат камбиален пръстен в повечето дървесни растения (като пример може да се вземе липата) се образува в самото начало на образуването на стъблото.
Делението на клетките на камбия става тангенциално, което определя подреждането на клетките в правилни редове по радиуса. С нарастването на ксилема камбият се приближава към периферията и обиколката му се увеличава поради радиалното делене на клетките му.
С вторично удебеляване вторичен ксилем или дървос паренхимни лъчи е всичко, което се отлага вътре в ствола, и всичко, което се отлага навън, т.е. към периферията на багажника, възлиза на вторичен флоем или флоемсъс сърцевинни лъчи.

Ксилемна проводяща системасе състои от трахеиди и съдове. При иглолистните растения само трахеидите изпълняват проводяща функция, а при широколистните дървесни растения - както трахеидите, така и съдовете.
Външен вид дървесни пръстенив ксилем (дървесина) възниква в резултат на периодична активност на камбия. През пролетта, като правило, има много вода и хранителни вещества, така че се образува камбият големи дървени елементис голям просвет и тънки стени. Към края на вегетационния период дейността на камбия замира и дървесината започва да преобладава. механични елементии тесни съдове.
Ядрени лъчисе състои от големи паренхимни клетки, които имат правоъгълна форма. Според произхода си тези лъчи биват първични и вторични. Първичните лъчи се простират от първичната кора до сърцевината и са по-дълги от вторичните. Основната функция на лъчите е да провеждат водата и органичните вещества в хоризонтална посока.
Механичен платв ксилема (дървесината) се състои от дебелостенни, тесни и вече лигнифицирани клетки.
Включени вторична флоемасъщо така включва 3 вида тъкани: основни, механични и проводими. Медуларните лъчи на флоема се простират от камбия навън и техните клетъчни стени не стават лигнифицирани. В периферната си част лъчите се разширяват силно. Механичната тъкан на вторичната флоема се нарича вторичен склеренхим; тя е представена от ликови влакна ( твърдо лико). Проводимата тъкан се състои от ситовидни тръби с придружаващи клетки ( меко лико).

В ранните етапи на развитие стъблото на двусемеделните има първична структура. В резултат на дейността на първичните меристеми на растежния конус се образуват епидермисът, първичната кора, централният цилиндър и сърцевината.
ЕпидермисСтъблото има малък брой устица, обикновено покрити с кутикула.
Под епидермиса има тъкани първичен кортекс.Външният му слой често се формира от коленхим, който е разположен или в непрекъснат пръстен (слънчоглед), или в отделни участъци. Основната част от първичната кора е изградена от хлоренхим и паренхим. Тук могат да се развият въздушни кухини и контейнери за секрети. Клетките на вътрешния слой на първичната кора обикновено са пълни с големи нишестени зърна (нишестена обвивка).
Централен цилиндърсе състои от перицикъл и проводяща система. Ядро -от паренхим, който често се разрушава и на негово място се образува въздушна кухина. Перициклът е представен от един или няколко реда склеренхим или негови отделни нишки. Провеждащата система има формата на кух цилиндър, който разделя първичната кора и ядрото. Проводящите снопове, които изграждат този цилиндър, са разделени от слоеве междуфасцикуларен паренхим - медуларни лъчи, които свързват сърцевината и първичната кора. Повечето двусемеделни се характеризират със специален тип централен цилиндър, в напречното сечение на който снопчетата са подредени в пръстен (фиг.).
рисуване. Стъбло на двусемеделни растения.
При двусемеделните растения слоят прокамбиеви клетки между първичната флоема и ксилемата запазва способността си да се дели и се превръща във вторична образователна тъкан - камбий, който се разделя, отлагайки елементи от вторична ксилема вътре и вторична флоема към периферията. Гроздовете на двусемеделните растения са отворени, благодарение на работата на камбия гроздът расте и диаметърът му се увеличава. Дейността на камбия осигурява появата на вторични елементи в стъблото, т.е. прехода от първичната към вторичната анатомична структура.
Вторичната структура е характерна за всички двусемеделни и иглолистни растения - треви и дървета.Разнообразието от типове структура се дължи преди всичко на подреждането на проводящите тъкани, което се определя от образуването на прокамбия и активността на камбия.
По време на процеса на диференциация на растежния конус се образува прокамбиум под формата на нишки. Ако нишките са разделени от достатъчно широки редове паренхим, тогава снопова или преходна структура на стъблото; ако нишките на прокамбия са толкова близо една до друга, че се сливат в цилиндър, тогава негредова (твърда) конструкция.
Снопова структура на стъблотосреща се в някои тревисти растения (копър, детелина, лютиче, грах, люцерна). Разположени са прокамбиалните връзки, вградени в растежния конус в един кръг по периферията на централния цилиндър. Всяка прокамбиална връв се превръща в отворен колатерален сноп, състоящ се от първичен ксилем, първичен флоем и ивица камбий между тях. Камбиевите клетки, разделяйки се, дават нови (вторични) елементи на съдовия сноп: навътре - ксилема към периферията - флоема. Проводящите снопове са разделени от широки междинни зони и обграждат мощно ядро.
В повечето растения камбият работи активно и диаметърът на снопчетата се увеличава значително, стимулира паренхима, който разделя снопчетата, който започва да се дели, давайки вторична образователна тъкан. Фасцикуларната и интерфасцикуларната меристема се затварят и образуват непрекъснат камбиален пръстен., чиято дейност осигурява равномерно удебеляване на стъблото. Ако фасцикуларният камбий произвежда елементи от вторична ксилема и флоема, тогава интерфасцикуларният камбий произвежда паренхим (при копър, мацка и др.). В детелина, люцерна и други растения интерфасцикуларният камбий отлага предимно клетки, подобни на склеренхим, към ксилемата. Тези клетки са удължени, дебелостенни, лигнифицирани. Следователно забавянето на прибирането на реколтата от детелина намалява качеството на сеното. Различната структура на пучка се поддържа в тези растения през целия живот на стъблото.
Гроздовата структура на стъблото може да се замени с напредване на възрастта с нетуфирана плътна. Преходен строеж на стъблототипичен за слънчоглед. Отначало стъблото се образува по същия начин, както при детелината, но интерфасцикуларният камбий отлага не паренхим, а ксилема и флоема. Образуват се допълнителни проводящи снопове. Постепенно всички снопове могат да се слеят в непрекъснат трислоен (ксилема, камбий, флоем) цилиндър. В горната (младата) част стъблото има снопов строеж, в основата (в старата част) е с нелъчева структура. .
Структура без снопхарактерни за стъблата на много треви и дървета. В растежния конус на тези растения прокамбиалните нишки са толкова близо една до друга, че образуват почти непрекъснат цилиндър (в напречно сечение изглежда като пръстен). Прокамбиалният цилиндър (пръстен), диференцирайки се, дава цилиндър от първична ксилема към центъра, цилиндър от първична флоема към периферията и между тях се образува камбиален цилиндър.
Вторичните промени са свързани с работата на камбия. Камбиевите клетки се делят успоредно на повърхността на стъблото, докато 10...20 пъти се отлагат вътре повече клеткиотколкото навън. Камбият образува елементи от вторична ксилема (дървесина) вътре, а вторичната флоема (лико) се отлага отвън. На някои места камбият отлага от двете страни паренхимни клетки на медуларните лъчи.
Характеристиките на нетуфираната структура на стъблата на тревата могат да се разгледат на примера на стъбло лен. Стъблото е покрито с епидермис, под който има рехав хлоренхим на първичната кора. Голямо количествоустицата в епидермиса и липсата на коленхим в първичната кора имат значителна практическо значение, тъй като по време на техническото накисване на лен водата не среща препятствия и лесно прониква вътре в стъблото. Клетките на нишестената обвивка (ендодермата) са по-големи и съдържат ясно видими нишестени зърна.
Централният цилиндър, съседен на ендодермиса, започва с групи от склеренхимни клетки - ликови влакна, между които са разположени паренхимни клетки. Ликовите влакна възникват в перицикъла и в първичната флоема. Те се отличават с много дебели, обикновено нелигнифицирани стени, състоящи се от 75...90% целулоза. Зряла клетка от ликово влакно е мъртва. Протопласт след удебеляване клетъчна стенае унищожена. Ликовите влакна имат необичайно висока якост. Якостта им на опън съответства на тази на стоманата, а еластичността им е по-висока. Клетките на ликовото влакно са много тясно свързани помежду си. Ленените влакна са отлична суровина за предене, използвана в текстилната промишленост.
След това цилиндрите (пръстените) са разположени в последователен ред: вторичен флоем, камбий, вторичен ксилем с ясно видими медуларни лъчи. Първичната ксилема е разположена в области, съседни на сърцевината. Паренхимът на сърцевината бързо се разрушава (в процеса на удължаване на стъблото), образувайки централна кухина.
Стъблата на двусемеделните тревисти растения се характеризират с:
1. вторична структура, възникващи много рано след първичната;
2. наличие на първична покривна тъкан - епидермис;
3. развита първична кора, разделена на коленхим (екзодерма), паренхим (мезодерма) и нишестена обвивка (ендодерма);
4. гредова или негредова (плътна) конструкция;
5. правилно подреждане на колатерални или биколатерални съдови снопове в един кръг по периферията на централния цилиндър;
6. наличие на камбий (снопове отворени).
Метаморфози на издънката.
Ако допълнителните функции започнат да преобладават над основните или органът придобива нови функции в процеса на развитие, той се променя значително както морфологично, така и анатомично. Такива модификации на органи се наричат метаморфози(от гръцката metamorphosis - трансформация, трансформация). Органи, които имат същия произход, но изпълняват различни функциинаречен хомоложни . Хомоложни органи са коренища, грудки, луковици, шипове на глог, кладодии, филокладии, жилки на грозде и тиква, т.к. всички те исторически са се развили от листни издънки.
Наричат се органи, които изпълняват еднакви функции и са сходни морфологично, но имат различен произход подобни . Подобни са кореновите шишарки и грудките на леторастите, шиповете на леторастите на глога и шиповете на листата на берберис, листата на граха и листата на тиквата.
Модификации на подземни издънки.
Подземните издънки са широко разпространени в природата: коренища, грудки, луковици. Те участват в доставката на вещества и вегетативното размножаване на растенията.
КоренищеТова е издънка с люспести листа, пъпки и допълнителни корени. На повърхността на коренището ясно се виждат белези от миналогодишните долни листа и мъртви издънки. Коренището има много допълнителни корени. Обикновено коренищата са подземни. Пъпките се развиват в надземни издънки. Коренищата са характерни за житната трева, ириса и водните лилии.
Столон - удължен тънък издънка с недоразвити листа. За разлика от коренището, обикновено е краткотрайно. При мускус и киселец столоните участват в натрупването на хранителни вещества, отложени в силно удебелените долни листа, но основната функция на столона е да участва във вегетативното размножаване.
Грудказа разлика от коренището и столона, той е силно скъсен и удебелен. Резервните вещества в него са локализирани в паренхима на стъблото.
Грудките могат да се развиват върху коренища, столони, главни леторасти и други части на растенията. Те могат да бъдат подземни и надземни.
Характерни са мощни коренищни грудки с голямо количество нишесте таро, или колоказия годни за консумацияот семейство ароидни, което се култивира в тропическите и субтропичните страни.
U картофигрудките се образуват в края на подземните столони - белезникави крехки издънки с люспести листа, развиващи се от аксиларни пъпки, разположени в основата на листните издънки. На повърхността на грудката има аксиларни пъпки - очиседят в малки ями, рамкирани от едната страна вежди -остатъци от основата на листата.
Грудки Ерусалимски артишок, или земна круша, също се образуват върху столони. Проводимите тъкани в тях са слабо развити. Клубените са богати на инулин, който се отлага главно в паренхима на стъблото.
Клубените, образувани върху столони и коренища, са краткотрайни. Те издържат само една година, между два вегетационни сезона, умирайки, след като пъпките им се развият в листни издънки, които консумират хранителни вещества, докато се образуват корени и растенията започнат да се хранят сами.
Крушка, подобно на грудка, това е специализирана, модифицирана съкратена издънка, която служи не само за съхраняване на хранителни вещества, но и за издържане на неблагоприятни периоди от годината, вегетативно възобновяване и възпроизводство на растенията. Луковиците са характерни за едносемеделните растения от семейства Лилиеви, Амарилисови и други. По изключение се срещат при двусемеделни растения: някои видове горски киселец и жълтурчета.
Стъблото се обади дъно,в луковицата е силно намалена и повече или по-малко сплесната. Носи листа под формата на люспи, а в долната част има адвентивни корени. За разлика от коренището и грудката, органите за съхранение на луковицата са месести люспи. Луковици се образуват при лалета, лешник, нарцис, зюмбюл, сцила, кокиче и други растения.
В зависимост от развитието на страничните пъпки, луковиците са простокогато пъпките са малки (лук, лилии и др.) и комплекс– пъпките в пазвите на сухите люспи са едри, силно развити (чесън).
Корм- подземна модификация на издънка, съчетаваща характеристиките на коренище и луковица. Развива се от издънка със силно скъсено стъбло, от долната част на което излизат допълнителни корени. На върха на луковицата или в основата й има пъпка, от която се образува цветонос. Стъблото на луковицата е покрито с основата на мъртви миналогодишни листа, които приличат на сухи филмови люспи. От пъпките, разположени в техните синуси, се развиват нови луковици - деца. Луковидните луковици включват шафран, гладиолус и безцветие.
Модификации на надземни издънки
Кочанзелето е по същество гигантска пъпка: средната част на главата на зелето е заета от месесто стъбло ( покер), по които има сочни, месести листа. Най-малките и най-младите от тях са разположени вътре в главата, около растежния конус, с който завършва стъблото. Брюкселското зеле оформя малки кочани от страничните пъпки.
В главата на зелето и двата органа на издънката участват в натрупването на хранителни вещества: водоразтворими захари и други биологично ценни съединения, включително витамини, но основната роля принадлежи на листата.
Надземни грудки, развиващи се от долната част на главния издънка, са характерни за зелето колраби.
В областта на съцветието също могат да се развият малки възли. При живородния възел например те се образуват в пазвите на долните покривни листа на съцветието. Техните пъпки започват да поникват върху майчиното растение. След падане те се вкореняват, образувайки нови растения.
Метаморфозираните издънки могат да служат не само за съхранение на хранителни вещества, размножаване и размножаване на растения, но и да изпълняват други функции.
При някои растения издънките губят листа по време на развитие и стъблото се превръща в фотосинтетичен орган, наречен в този случай кладодиум . Обикновено е сплескан и има способността да расте дълго време (в тропическото растение Mühlenbeckia или Homalocladus от семейството на елдата, бодлива круша). Възлите имат добре дефинирани стеснения, така че кладодиумът изглежда сегментиран. При влажни условия кладодиите могат да бъдат листопадни, но при сухо време листата падат. Като орган на фотосинтезата кладодиите имат добре развита тъкан, носеща хлорофил, разположена под епидермиса.
Cladodia, според външен видподобни на листа се наричат филокладии . Те се развиват от страничните пъпки, така че винаги се намират в пазвите на малък филмов или люспест лист. Филокладиите са много разнообразни. При перестите или настръхналите аспержи те са дребни, игловидни, при аспержите на Sprenger или гъсто нацъфтели са сплескани и линейни. Широко ланцетни или яйцевидни очертания, кожените филокладии са характерни за видовете месарска метла, срещащи се в Средиземноморието, Закавказието и Крим. През лятото от долната страна на филокладия, в средната му част, се развива съцветие, разположено в пазвата на малкия покривен лист, през есента и зимата цъфти метлата, а през пролетта узряват оранжеви плодове; филокладията.
Доста често срещан тип промяна на издънките е трън . Това е силно дървесен, безлистен, скъсен издънка с остър връх. При глога издънката, която се развива в пазвата на листата през пролетта, първоначално има малки листа и връхна пъпка. Скоро апикалният растеж спира, листата падат, стъблото силно лигнифицира и се превръща в трън. Шиповете се развиват по същия начин.
Прилепналите растения са оборудвани антени , които, подобно на шипове, са модифицирани странични издънки в някои растения. Пасифлората има прости, неразклонени пипала с извит връх. Те се намират в пазвите на листата, както и цветовете. Разклонените пипала на гроздето са модифицирани съцветия. При момичешкото грозде краищата на разклонените пипала са дисковидни и играят ролята на вендузи, помагайки на растението да се изкачи върху опора. В тиквата, динята, краставицата и други представители на семейството на тиквите също се образуват пипала с произход от издънки.
Първична анатомична структура на стъблото.
Дефиниция и функции на стъблото.
Стъблото е аксиален, обикновено радиално симетричен орган с продължителен апикален (апикален) растеж.
Функции на стъблото:
1. осъществява двупосочно движение на веществата между корените и листата,
2. поддържа короната на растението,
3. спомага за увеличаване на общата асимилационна повърхност на растението поради разклоняване,
4. участва в съхраняването на резервни вещества,
5. в млада възраст – извършва и фотосинтеза.
Тези функции определят наличието в стъблото на покривни, добре развити проводящи, механични тъкани и функционално развит паренхим.
Структура на растежния конус.
В конуса на растеж на стъблото на покритосеменните се изразяват две зони: външна - туника, а вътрешните – рамка,които се различават в равнините на клетъчно делене (теория на А. Шмид и Й. Будер) (фиг. 1).
Туниката може да се състои от един или повече слоеве клетки, които се делят перпендикуляренповърхност, при повечето двусемеделни растения е двуслоен. Външният слой на туниката поражда протодермис,от който след това се образува епидермисът, покриващ листата и стъблото. Ако туниката е многопластова, тогава вътрешният слой образува тъканта на първичната кора.
При наличие на еднослойна туника цялата първична кора и стела се формират от корпуса. Рамкаобразувани от маса клетки които са разделени във всички равнини.
Листните примордии най-често се появяват във втория слой на туниката, а в тялото се образуват аксиларни пъпки.
Фигура 1. б) връх на покритосеменни растения.
В резултат на дейността на първичните меристеми на върха се формира първичната анатомична структура на стъблото: епидермис, първична кора, централен цилиндър и сърцевина.
Вторичното удебеляване възниква поради активността на камбия.
Първична анатомична структура на стъблото.
Структурата на стъблото, образувана в резултат на дейността на първичните меристеми на върха, се нарича първичен. В стъблото, с първичната структура, могат да се разграничат три анатомични и топографски зони:
1. покривна тъкан,
2. първична кора
3. централен цилиндър.
Епидермисът предпазва стъблото от изпаряване.
Под епидермиса е външният слой на първичната кора - екзодерма -представени от хлоренхим и (при двусемеделните) коленхим. Във вътрешния слой, граничещ с централния цилиндър - ендодермис -обикновено се натрупват нишестени зърна, а след това се превръща в т.нар вагина, носеща нишесте (слой, носещ нишесте).Зърната нишесте могат да се утаят в клетките под въздействието на гравитацията. Поради това ендодермисът играе важна роля в геотропичната ((от гръцки trpos - завой, посока), растежни движения на стъблата на растенията, причинени от насоченото действие на гравитацията) реакция на стъблата
По периферията на централния цилиндър в перициклетсе намира склеренхим. Склеренхимните влакна образуват непрекъснат слой или се събират близо до васкуларни снопове, навън от първичния флоем. Основен елементцентрален цилиндър (стела) - проводими снопове. Те се образуват от прокамбиални нишки и се състоят от първична ксилема и флоема. Диференциацията на прокамбия се извършва центростремително: флоема се образува от външната страна на снопа към центъра му, а ксилема към него (фиг. 2). Прокамбият може да бъде напълно превърнат в първични проводящи комплекси, образувайки затворен колатерален сноп (при едносемеделните) или останалите клетки в центъра на снопа образуват камбий. Такива отворени гроздове са характерни за двусемеделните растения. Работата на камбия осигурява образуването на вторична ксилема и флоема.
Фигура 2.
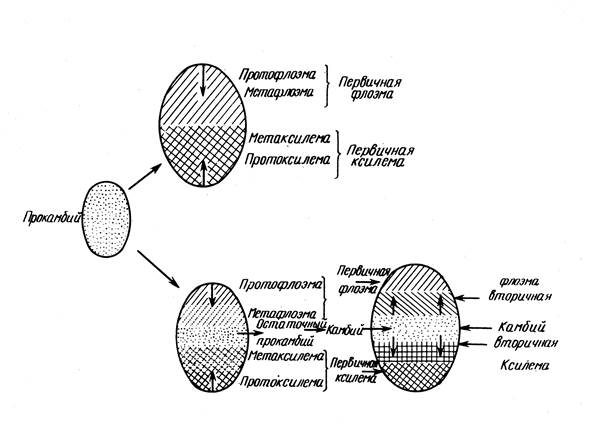
Образуването на прокамбия определя структурата на проводящата система:
1. Ако прокамбиумът образува твърд кух цилиндър, тогава от него се образуват вложени цилиндри от ксилема и флоема (в някои иглолистни и двусемеделни) - негредова конструкция.
2. Ако прокамбиумът се състои от отделни нишки (при повечето едносемеделни, много двусемеделни и иглолистни), тогава възникващите първични проводящи тъкани запазват същите пакетна структура.
Прокамбиумът възниква в основата на листния примордиум и развитието му протича в две посоки: към върха на листния примордиум и надолу по стъблото, където се свързва с нишките, които са възникнали по-рано. Диференциацията на флоема и ксилемата се извършва в същата последователност. Обикновено първо се образуват флоемни елементи, след това ксилема. Проводимите снопове са разположени в паренхима на централния цилиндър.
В центъра на стъблото се развива ясно видима сърцевина. Паренхимните тънкостенни клетки на сърцевината често се разхлабват, умират и се изпълват с въздух, след което цялата тъкан изглежда бяла (при слънчоглед, царевица, бъз). Понякога сърцевината умира много рано, когато удължаването на междувъзлията все още не е приключило. Това води до разкъсване на сърцевината и образуване на кухо стъбло.
Първичният строеж на стъблото се запазва при едносемеделните през целия живот, а при двусемеделните и голосеменните - само в ранните фази от развитието му.
Структурата на стъблото.
Стъблото е оста на издънката, носеща листа и пъпки. Основните функции на стъблото са опорни и проводящи. Стъблото осигурява връзката между корените и листата. Освен това в стъблото често се отлагат резервни хранителни вещества. Понякога стъблото е асимилиращ орган.
В резултат на дейността на прокамбия и останалите първична меристемаКонусът на растежа образува първичната структура на стъблото. В първичното стъбло обикновено се разграничават първична кора и стела (централен цилиндър). За разлика от корена, първичната кора е покрита отвън с епидермис.
При голосеменните и повечето двусемеделни покритосеменни растежът на стъблото в дебелина се осъществява от камбия, който образува вторични тъкани. Появява се под формата на цилиндър между първичната ксилема и първичната флоема и остава в относително същата позиция за неопределено време, отлагайки вторична ксилема към центъра на оста и вторична флоема навън.
Вторично удебеляване възниква и в резултат на активността на фелогена (корковия камбий).
При двусемеделните дървета и храсти, както и при иглолистните дървета, вторичните удебелявания могат да продължат много години. В резултат на това стъблото има три основни части: кора, дърво и сърцевина.
Разлики в структурата на стъблото на едносемеделните и двусемеделните растения.
При едносемеделните стъблото е тревисто, неспособно на вторично удебеляване и рядко се разклонява. Съдови снопчета без камбий са разпръснати по цялото стебло.
При двусемеделните стъблото е тревисто или вдървесинено, способно на вторично удебеляване и разклонения. Съдовите снопове, които имат камбий, са разположени в една голяма маса в центъра на стъблото или имат вид на пръстен.
15 Функции на цветето.
Цветето е модифициран скъсен издънка, пригоден за възпроизвеждане на покритосеменни (цъфтящи) растения.
Изключителната роля на цветето се дължи на факта, че то съчетава всички процеси на безполово и половото размножаване, докато в по-ниските и мн. висши растенияте са разединени. В двуполов цвят се извършват микро- и мегаспорогенеза, микро- и мегагаметогенеза, опрашване, оплождане и образуване на семена и плодове. Структурните особености на цветето позволяват изброени функциис минимален разход на пластични вещества и енергия.
Структура на цветето
Цветето се състои от стволова част (дръжка и вместилище), листна част (чашелистчета, венчелистчета) и генеративна част (тичинки, плодник или плодници). Цветът е прикрепен към стъблото с помощта на дръжка. Ако дръжката е силно скъсена или липсва, цветето се нарича приседнало (живовляк, върбинка, детелина). Педицелът също така съдържа два (при двусемеделните) и един (при едносемеделните) малки предлистчета - прицветници, които често могат да липсват. Горната разширена част на дръжката се нарича вместилище, върху което са разположени всички органи на цветето. Съдът може да има различни размери и форми - плосък (божур), изпъкнал (ягода, малина), вдлъбнат (бадем), издължен (магнолия). При някои растения, в резултат на сливането на съда, долните части на обвивката и андроцея, специална структура- хипантиум. Формата на хипантия може да бъде разнообразна и понякога да участва в образуването на плода (цинародий - шипка, ябълка). Hypanthium е характерен за представители на семействата роза, цариградско грозде, саксифраж и бобови растения.
Частите на цветето се делят на плодовити или репродуктивни (тичинки, плодник или плодници) и стерилни (околоцветник).
функции на цветя
Именно благодарение на цветята растенията живеят и се размножават. Красивата форма и цвят на цветето, на първо място, изпълнява важна функция за тяхното размножаване - те привличат насекоми или опрашващи птици. След опрашването в яйчника на цветето се образуват семена, които ще дадат живот на нови растения. Не всички растения дават цветя и не всички цветя могат да се похвалят с привлекателни форми и цветове. Тези с цветен външен вид обаче привличат потенциални опрашители. Не е необичайно определено растение да бъде опрашено от определен вид насекоми, които имат органите, необходими за опрашването им. Най-известните насекоми опрашители са пчелите и мухите. Някои цветя се опрашват от птици, като например колибри. При растенията, опрашвани от вятъра, цветята са малки и незабележими на вид, тъй като тяхната форма и цвят не могат да окажат влияние върху процеса на опрашване. Някои цветя, според хората, имат неприятна миризма, например стапелия, чиято миризма напомня миризмата на разлагащо се месо.
16 въпрос)))
Опрашваните от вятъра растения обикновено имат редица характерни особености: много малки и многобройни цветове, произвеждат много прашец. Едно растение може да произведе милиони поленови зърна. При много растения, опрашвани от вятъра (лешник, трепетлика, елша, черница), цветята се появяват дори преди листата да цъфтят.
Пчелите предпочитат да събират прашец от опрашвани от насекоми растения. Но ако в природата има малко цъфтящи ентомофилни растения и пчелите се нуждаят от цветен прашец, те също го събират от опрашвани от вятъра растения.
Опрашването на цъфтящите растения се извършва по два основни начина - чрез вятър и животни, най-често насекоми. И в двата случая растенията развиват специфични адаптации. Опрашваните от насекоми растения се характеризират с големи, ярко оцветени единични цветя, както и съцветия, състоящи се от ярки цветя различни форми. Като правило те имат силна миризма. Имат развити специални жлези – нектарници, които произвеждат сладък течен секрет – нектар. Цветовете на растенията, опрашвани от насекоми, са богати на прашец. Поленовите зърна обикновено са големи и лепкави, а обвивката им често има различни издатини. При ветроопрашваните растения околоцветникът е частично или напълно редуциран, а дребните им и незабележими цветове обикновено са събрани в съцветия. Опрашваните от вятъра растения произвеждат много повече прашец от растенията, опрашвани от насекоми. Въпреки това поленовите им зърна са малки и сухи, добре транспортирани от вятъра. Те се образуват в големи прашници, често висящи на дълги нишки. Стигмата на много от тях е раздвоена и покрита с множество косми и четинки, което им позволява да улавят много повече прашец от въздуха. Много ветроопрашвани растения цъфтят в началото на пролетта, дори преди листата да цъфтят. Опрашваните от вятъра треви, които цъфтят през лятото, цъфтят строго според часовника, като по този начин спестяват цветен прашец. Вероятността за опрашване също се увеличава от факта, че всички опрашвани от вятъра растения растат в близки групи или големи клъстери.
17. Плодове и семена. Класификация, методи на разпространение
Плодът. Той е един от най-характерните органи на покритосеменните растения. Състои се от перикарп и семена. Перикарпът, който е обрасла и силно модифицирана стена на яйчника, осигурява образуването на семена, защита от неблагоприятни фактори и насърчава тяхното разпространение.
Сочни плодовеимат добре развита пулпа, в големите паренхимни клетки на която се натрупват много вода, въглехидрати, витамини, органични киселини и различни ароматни съединения.
В сухите плодовеперикарпа жилав или дървесен. Те могат да бъдат едносеменни или многосеменни, дехисцентни или недехисцентни. При много плодове отделянето на сухия перикарп позволява разпръскване на семената. Сухите многосеменни плодове често се отварят с помощта на дупки, капачки и капаци.
Безплодие- това е колекция от зрели, слети плодове (сухи или сочни) от едно съцветие (черница, хлебно дърво, ананас).
Цъфтящите растения са развили различни адаптации за разпръскване на семена и плодове. Много растения имат плодове, които се разпространяват от вятъра.Те са малки, леки и често снабдени с крилати израстъци или мухи (глухарче, трън).
Плодовете на някои растения се разпространяват вода(елша, водна лилия, кокосова палма, много видове острица). Сочните плодове с ярък цвят и ароматна миризма се ядат с готовност от много видове животни. Семената на тези плодове не се усвояват и попадат в почвата заедно с екскрементите. Сухите плодове развиват различни приставки, кукички, които се захващат за животински косми и човешко облекло и се носят от тях (репей, велкро, връв). При някои растения зрелите плодове се напукват и развиващите се клапи на перикарпа изхвърлят семена - понякога на значително разстояние (импатиенс, луда краставица).
семена.Обикновено семето се състои от ембрион и ендосперм, заобиколен от семенна обвивка (фиг. 8.22).
Тестаобразувани от обвивката на яйцеклетката. Тя защитава семето отмеханични повреди, проникване на патогенни микроорганизми, прекомерна загуба на вода. Семената на много растения имат различни приспособления за разпръскване - власинки (върба, топола), месести придатъци (за привличане на животни).
Зародиш - това е зачатъкът на нов индивид, миниатюрен спорофит.При повечето цъфтящи растения зародишът се състои от зародишно коренче, дръжка и пъпка. Котиледонните листа са прикрепени към върха на стъблото (котиледони).В този случай при двусемеделните растения се образуват два котиледона (те съдържат резервни вещества на семето), при едносемеделните растения - един. Единичният котиледон на житния зародиш се нарича щит.Намира се на границата с ендосперма и по време на покълването на зърното подпомага притока на разтворени хранителни вещества от ендосперма към ембриона.
Ендосперм- тъкан, съдържаща хранителни вещества. Образува се не само в почти всички едносемеделни растения, но и в редица двусемеделни растения (лен, копър, моркови и др.). Трихуидните клетки на ендосперма, както и котиледоните на ембриона, съдържат значителен запас от хранителни вещества (нишесте, протеини, мазнини и др.), Необходими за развитието на ембриона и образуването на разсада.
Класификация на семенатапросто. Има пет вида семена в зависимост от това къде се натрупват резервните продукти: в ендосперма, нуцелуса, ембриона, в ендосперма и нуцелуса в ендосперма и ембриона.
Развитието на яйцеклетката започва с появата на среден туберкул - нуцелус, вътре в който в ранните етапи на развитие се откроява една голяма спорогенна клетка. По-късно претърпява редукционно (мейотично) делене и образува 4 неравни клетки - мегаспори. Следователно нуцелусът трябва да се счита за мегаспорангий. Ядрото образува централната част на яйцеклетката
Семената имат една много важна характеристика: при условия, неблагоприятни за покълване, те могат да останат в латентно състояние за значителен период от време. С настъпването на благоприятни условия на температура и влажност семето започва да абсорбира вода и при достатъчен достъп до въздух покълва. Семената се разпространяват от вода, вятър, животни, случайно разпределение (луда краставица..внимание на съучениците: когато семената на лудата краставица достигнат пълна зрялост, околните месести тъкани се превръщат в лигава маса.
При отделянето на краставица от дръжката се създава натиск върху съдържанието на плода според принципа реактивна тяга, който движи плода в посока, обратна на полета на семената, което допринася за разпръскването на семената върху по-голяма площ.
18. Лишеи. Структурни особености, разпространение, цикъл на развитие, значение в природата и живота на човека. Представители
Лишеи- Това е пример за облигатна симбиоза на гъби и водорасли. Според естеството на сексуалното спороношение лишеите се класифицират в два класа: торбести (размножават се чрез спори, които узряват в торбички), който включва почти всички разновидности на лишеи, и базидиални (спорите узряват в базидии), наброяващи само няколко десетки вида. Според структурата на тялото ( тали) има корови (корови), фолиозни и плодови лишеи. Вегетативното тяло на коровите лишеи е най-примитивно, то може да бъде гранулирано, поресто или под формата на корички. По-развити са листните лишеи, които имат вид на повече или по-малко разчленени плочи. Фрутикозните лишеи са силно организирани, имат вид на храсти, висящи нишки или изправени израстъци.
Разпръскване.Лишеите растат на почвата ( епигейски), камъни ( епилитен) или стволове на дървета ( епифитен), получавайки необходимата за живота влага от атмосферата. Някои видове живеят в морската крайбрежна зона. Когато за първи път се заселят на безплодни места, лишеите образуват хумус, когато умрат, върху който след това могат да се заселят други растения. Лишеи дори са открити в безплодни арктически пустини и вътре в антарктически скали. Лишеите са разпространени по целия свят, но са особено разнообразни в тропиците, планините и тундрата. Но в лабораториите лишеите умират доста бързо. И едва през 1980 г. американски учени успяха да „комбинират“ водорасло и гъба, отгледана от спора. Лишеите са биоиндикаторни организми; растат само на екологично чисти места, така че няма да ги намерите в големите градове и индустриални зони.
Възпроизвеждане.Лишеите се размножават основно по вегетативен път - чрез части от талуса, както и чрез специални специализирано образование- соредия и изидия (фиг. 6.4).
Соредиясе образуват под горната кора във фотосинтетичния слой и се състоят от една или няколко фикобионтни клетки, преплетени с гъбични хифи. Под натиска на обраслата маса от многобройни соредии кортикалния слой на талуса се счупва и соредиите излизат на повърхността, откъдето се носят от вятъра, водата и при благоприятни условия прерастват в нови тали лишеи.
ИзидияТе са малки израстъци на талуса под формата на пръчки, туберкули, покрити с кора отвън. Те се състоят от няколко фикобионтни клетки, преплетени с гъбични хифи. Изидията се откъсва и образува нови тали.
Значението на лишеите в биосферата и националната икономика.Известни са около 26 хиляди вида лишеи. Те са широко разпространени в природата, с изключение на местата, където въздухът е наситен с вредни газове. Лишеите са много чувствителни към замърсяването на въздуха и затова повечето от тях са големи градове, както и в близост до заводи и фабрики, бързо умира. Поради тази причина те могат да служат като индикатори за замърсяване на въздуха с вредни вещества.
Като автохетеротрофни организми, лишеите акумулират слънчева енергия и създават органична материяна места, недостъпни за други организми, а също така разграждат органичната материя, участвайки в общия кръговрат на веществата в биосферата. Лишеите играят важна роля в почвообразуващия процес, тъй като постепенно се разтварят и унищожават скали, върху които се заселват и поради разлагането на техните тали се образува почвен хумус. По този начин лишеите, заедно с бактериите, цианобактериите, гъбите и някои водорасли, създават условия за други, по-напреднали организми, включително висши растения и животни.
IN стопанска дейностЗа хората важна роля играят предимно хранителни лишеи, като мъх от северен елен или мъх, исландски мъх и други, които се ядат не само от северни елени, но и от елени, мускусни елени, сърни и лосове. Някои видове лишеи (лишайник, хигрофора) намират приложение и в парфюмерийната индустрия за производство на ароматни вещества, във фармацевтичната индустрия за производство на лекарства срещу туберкулоза, фурункулоза, чревни заболявания, епилепсия, Лишеевите киселини се получават от лишеи (известни са около 250) с антибиотични свойства.
представители:Еленски мъх, храстовиден лишей на брадата.
U двусемеделни растениястъблото, преди началото на активността на камбия, което води до радикално преструктуриране на неговата анатомия, има следната структура: отвън е покрито епидермис, следва първичен кортекс, съставен от няколко реда предимно паренхимни клетки; по към центъра се намира централен цилиндър, съставен от съдово-фиброзни снопчета и ядро. Ендодерма, отделящ кората от централния цилиндър, присъства само в няколко растения.
Епидермиссъставен от хомогенни клетки с обичайната за епидермиса структура, плътно притиснати една към друга. IN първичен кортексобикновено се разграничават два слоя. Външният се състои от плътно свързани една с друга клетки. Тези клетки имат удебеления в ъглите на мембраните и представляват коленхим. Вътрешният слой на кората е съставен от по-големи, хлабаво свързани помежду си клетки с тънки черупки. Ендодермата (ако има такава) е представена от един слой големи клетки, съдържащи зърна нишесте („нишестена вагина“).
Централен цилиндързапочва перициклет, което е мястото на образуване на адвентивни корени (възникващи ендогенно). Понякога перицикълът се развива в многоклетъчна механична тъкан, която покрива външната страна на съдовия сноп. Такива перициклични влакнаимат удебелени, често лигнифицирани стени или запазват влакнестия си състав (фиг. 105). Основната, най-важна част от централния цилиндър е проводима тъкан под формата на отворени съдово-фиброзни снопове, подредени в правилен пръстен. Те са от колатерален тип, тоест ксилемата им е обърната навътре, флоемата е обърната навън, а между тях има тесен слой камбий (фиг. 106). Сноповете са разделени от групи паренхимни клетки, свързващи кората със сърцевината. Това ще бъдат първични медуларни лъчи. Понякога съдовите снопове са разположени не в един, а в два слоя. В този случай вътрешните снопове се състоят само от флоема. Когато са плътно долепени до външните снопове, възникват двустранни снопове (в тикви).
Сърцевината, която заема централната част на стъблото, е изградена от големи, плътно свързани паренхимни клетки. Понякога сърцевината изчезва и вместо нея в центъра на стъблото се образува въздушна кухина (в чадър).Материал от сайта