Stem - this is the axial structure of the shoot. And its anatomical structure follows from its main functions. Firstly, the stem is characterized by the development of mechanical and conductive tissues. Secondly, the stem has complex system meristems: apical, lateral and intercalary, which determine its growth over a long time, as well as the emergence of new organs. The stem arises from the apical meristem, from which three layers of tissues differentiate: the main, integumentary, and conductive.
The structure of monocots
The entire procambium or primary meristem monocots differentiates into elements of primary conducting tissues. Their stems, especially if we consider herbaceous (cereals), have a simpler structure compared to stems dicotyledonous plants. They are also mainly characterized primary structure. Monocots have vascular-fibrous vascular bundles closed (without cambium), consist only of primary tissues and are randomly located in the main parenchyma of the stem.

The structure of dicotyledonous plants
In the middle part of the procambial cord in dicotyledonous plants, the formation of cambium occurs and the formation of secondary conductive tissues (metaphloem and metaxylem) begins. The volume of these secondary conducting tissues increases due to the division of cambium cells, which ultimately leads to a strong thickening of the stem.
Dicotyledons are characterized by open tufts with a cambium. Conductive tissues in the stems of dicotyledonous plants arranged in a ring around the core. The bundles are separated by medullary rays, which consist of parenchyma and connect the medulla with either the pericycle or the primary cortex. Along the periphery in the stems of dicotyledonous plants there are mechanical fabrics, wherein sclerenchyma is part of the pericycle, collenchyma, together with the main parenchyma, is part of the primary cortex.
The structure of the central cylinder of dicotyledons can be beam And non-beam.
In their anatomical structure, annual stems of woody plants are similar to the stems of herbaceous plants: their conducting systems have non-beam structure. Their distinctive feature is active work cambium and early formation of secondary integumentary tissue - traffic jams.
Thanks to the activity of the cambium, various elements of secondary origin are formed in the stem, while the primary elements gradually disappear. The cambium forms xylem and phloem elements at different rates: for one phloem cell, the cambium separates several xylem cells. As a result of this, xylem (or wood) grows much faster than phloem (bast); accordingly, almost the entire mass of the trunk and branches of the tree falls on xylem. Phloem, on the contrary, makes up a relatively thin layer of the secondary cortex.

A continuous cambial ring in most woody plants (linden can be taken as an example) is formed at the very beginning of stem formation.
Cambium cell division occurs tangentially, which determines the arrangement of cells in regular rows along the radius. As xylem grows, the cambium moves closer to the periphery and its circumference increases due to the radial division of its cells.
With secondary thickening secondary xylem or wood with parenchyma rays is everything that is deposited inside the trunk, and everything that is deposited outside, i.e. to the periphery of the trunk, amounts to secondary phloem or phloem with core rays.

Xylem conducting system consists of tracheids and vessels. In coniferous plants, only tracheids perform the conducting function, and in deciduous woody plants, both tracheids and vessels.
Appearance tree rings in xylem (wood) occurs as a result of periodic activity of the cambium. In spring, as a rule, there is a lot of water and nutrients, so the cambium forms large wood elements with large clearance and thin walls. Towards the end of the growing season, the activity of the cambium dies out and the wood begins to predominate. mechanical elements and narrow vessels.
Core rays consist of large parenchyma cells that have a rectangular shape. According to their origin, these rays can be primary and secondary. The primary rays extend from the primary cortex to the pith, and they are longer than the secondary ones. The main function of the rays is to conduct water and organic substances in a horizontal direction.
Mechanical fabric in xylem (wood) it consists of thick-walled, narrow and already lignified cells.
Part secondary phloem also includes 3 types of fabrics: basic, mechanical and conductive. Phloem medullary rays extend from the cambium outward, and their cell walls do not become lignified. In their peripheral part, the rays expand greatly. The mechanical tissue of the secondary phloem is called secondary sclerenchyma; it is represented by bast fibers ( hard bast). Conductive tissue consists of sieve tubes with companion cells ( soft bast).

At the early stages of development, the stem of dicotyledons has a primary structure. As a result of the activity of the primary meristems of the growth cone, the epidermis, primary cortex, central cylinder and core are formed.
Epidermis The stem has a small number of stomata, usually covered with a cuticle.
Below the epidermis are tissues primary cortex. Its outer layer is often formed by collenchyma, which is located either in a continuous ring (sunflower) or in separate sections. The main part of the primary cortex is composed of chlorenchyma and parenchyma. Air cavities and secretion containers can develop here. The cells of the inner layer of the primary cortex are usually filled with large starch grains (starch sheath).
Central cylinder consists of a pericycle and a conducting system. Core - from parenchyma, which is often destroyed, and in its place an air cavity is formed. The pericycle is represented by one or several rows of sclerenchyma or its individual strands. The conducting system has the form of a hollow cylinder that separates the primary cortex and the core. The conducting bundles that make up this cylinder are separated by layers of interfascicular parenchyma - medullary rays that connect the core and the primary cortex. Most dicotyledons are characterized by a special type of central cylinder, the cross section of which the bundles are arranged in a ring (Fig.).
Drawing. Stem of dicotyledonous plants.
In dicotyledonous plants, the layer of procambium cells between the primary phloem and xylem retains the ability to divide and turns into secondary educational fabric - cambium, which divides, depositing elements of secondary xylem inside and secondary phloem to the periphery. The bunches of dicotyledonous plants are open; thanks to the work of the cambium, the bunch grows and its diameter increases. The activity of the cambium ensures the emergence of secondary elements in the stem, i.e., the transition from the primary to the secondary anatomical structure.
The secondary structure is characteristic of all dicotyledonous and coniferous plants - grasses and trees. The variety of structure types is due, first of all, to the arrangement of conducting tissues, which is determined by the formation of the procambium and the activity of the cambium.
During the process of differentiation of the growth cone, procambium is formed in the form of strands. If the strands are separated by sufficiently wide rows of parenchyma, then bundled or transitional stem structure; if the procambium strands are brought so close together that they merge into a cylinder, then non-beam (solid) structure.
Bundle structure of the stem found in some herbaceous plants (dill, clover, buttercup, peas, alfalfa). The procambial cords embedded in the growth cone are located in one circle around the periphery of the central cylinder. Each procambial cord turns into an open collateral bundle consisting of primary xylem, primary phloem and a strip of cambium between them. Cambium cells, dividing, give new (secondary) elements of the vascular bundle: inward - xylem to the periphery - phloem. The conducting bundles are separated by wide interbundle zones and surround a powerful core.
In most plants, the cambium works actively and the diameter of the bundles increases significantly. The activity of the bundle cambium stimulates the parenchyma that separates the bundles, which begins to divide, giving secondary educational tissue. The fascicular and interfascicular meristems close together and form a continuous cambial ring., whose activity ensures uniform thickening of the stem. If the fascicular cambium produces elements of secondary xylem and phloem, then the interfascicular cambium produces parenchyma (in dill, chickweed, etc.). In clover, alfalfa and other plants, the interfascicular cambium deposits predominantly sclerenchyma-like cells towards the xylem. These cells are elongated, thick-walled, lignified. Therefore, delay in harvesting clover reduces the quality of hay. The distinct fascicle structure is maintained in these plants throughout the life of the stem.
The bunched structure of the stem can be replaced with age by a non-tufted solid one. Transitional structure of the stem typical for sunflower. At first, the stem is formed in the same way as in clover, but the interfascicular cambium deposits not parenchyma, but xylem and phloem. Additional conducting bundles are formed. Gradually, all bundles can merge into a continuous three-layer (xylem, cambium, phloem) cylinder. In the upper (young) part the stem has a bunched structure, at the base (in the old part) it has a non-beamed structure. .
Non-bundle structure characteristic of the stems of many grasses and trees. In the growth cone of these plants, the procambial strands are so close together that they form an almost continuous cylinder (in a cross section it looks like a ring). The procambial cylinder (ring), differentiating, gives a cylinder of primary xylem to the center, a cylinder of primary phloem to the periphery, and a cambial cylinder is formed between them.
Secondary changes are associated with the work of the cambium. Cambium cells divide parallel to the surface of the stem, while 10...20 times are deposited inside more cells than out. The cambium forms elements of secondary xylem (wood) inside, and secondary phloem (bast) is deposited outside. In places, the cambium deposits parenchyma cells of the medullary rays on both sides.
Features of the non-tufted structure of grass stems can be considered using the example of a stem flax. The stem is covered with epidermis, under which there is loose chlorenchyma of the primary cortex. A large number of stomata in the epidermis and the absence of collenchyma in the primary cortex have a significant practical significance, since during the technical soaking of flax, water does not encounter obstacles and easily penetrates inside the stem. The cells of the starch sheath (endoderm) are larger and contain clearly visible starch grains.
The central cylinder adjacent to the endodermis begins with groups of sclerenchyma cells - bast fibers, between which parenchyma cells are located. Bast fibers arise in the pericycle and in the primary phloem. They are distinguished by very thick, usually non-lignified walls, consisting of 75...90% cellulose. A mature bast fiber cell is dead. Protoplast after thickening cell wall is destroyed. Bast fibers have unusually high strength. Their tensile strength corresponds to that of steel, and their elasticity is higher. The cells of bast fiber are very closely connected to each other. Flax fiber is an excellent spinning raw material used in the textile industry.
Next, cylinders (rings) are located in sequential order: secondary phloem, cambium, secondary xylem with clearly visible medullary rays. Primary xylem is located in areas adjacent to the pith. The parenchyma of the pith is quickly destroyed (during the process of stem elongation), forming a central cavity.
The stems of dicotyledonous herbaceous plants are characterized by:
1. secondary structure, arising very early after the primary;
2. the presence of primary integumentary tissue - the epidermis;
3. developed primary cortex, divided into collenchyma (exoderm), parenchyma (mesoderm) and starch sheath (endoderm);
4. beam or non-beam (solid) structure;
5. correct arrangement of collateral or bicollateral vascular bundles in one circle along the periphery of the central cylinder;
6. presence of cambium (bundles open).
Metamorphoses of the shoot.
If additional functions begin to prevail over the main ones, or an organ acquires some new functions in the process of development, it changes greatly both morphologically and anatomically. Such modifications of organs are called metamorphoses(from the Greek metamorphosis - transformation, transformation). Organs that have the same origin but perform different functions called homologous . Homologous organs are rhizomes, tubers, bulbs, hawthorn spines, cladodes, phyllocladies, tendrils of grapes and pumpkins, because all of them historically developed from leafy shoots.
Organs that perform the same functions and are similar morphologically, but have different origins are called similar . Similar are root cones and shoot tubers, hawthorn shoot spines and barberry leaf spines, pea leaf tendrils and pumpkin shoot tendrils.
Modifications of underground shoots.
Underground shoots are widespread in nature: rhizomes, tubers, corms, bulbs. They are involved in the supply of substances and vegetative propagation of plants.
Rhizome It is a shoot with scale-like leaves, buds and adventitious roots. Scars from last year's lower leaves and dead shoots are clearly visible on the surface of the rhizome. The rhizome has many adventitious roots. Usually the rhizomes are underground. Buds developing into above-ground shoots. Rhizomes are characteristic of wheatgrass, iris, and water lilies.
Stolon - an elongated thin shoot with underdeveloped leaves. Unlike the rhizome, it is usually short-lived. In musk and wood sorrel, stolons participate in the accumulation of nutrients deposited in the strongly thickening lower leaves, but the main function of the stolon is to participate in vegetative propagation.
Tuber in contrast to the rhizome and stolon, it is greatly shortened and thickened. The reserve substances in it are localized in the stem parenchyma.
Tubers can develop on rhizomes, stolons, main shoots and other parts of plants. They can be underground or aboveground.
Powerful rhizomatous tubers with a large amount of starch are characteristic of taro, or colocasia edible from the aroid family, which is cultivated in tropical and subtropical countries.
U potatoes tubers are formed at the end of underground stolons - whitish fragile shoots with scale-like leaves, developing from axillary buds located at the base of leafy shoots. On the surface of the tuber there are axillary buds - eyes sitting in small pits framed on one side eyebrows - remnants of leaf bases.
Tubers Jerusalem artichoke, or earthen pear, are also formed on stolons. The conductive tissues in them are poorly developed. Tubers are rich in inulin, which is deposited mainly in the stem parenchyma.
Tubers formed on stolons and rhizomes are short-lived. They last only one year, between two growing seasons, dying off after their buds develop into leafy shoots that consume nutrients until roots form and the plants become self-feeding.
Bulb, like a tuber, it is a specialized, modified shortened shoot that serves not only for storing nutrients, but also for enduring unfavorable periods of the year, vegetative regeneration and plant reproduction. Bulbs are characteristic of monocotyledonous plants from the families Liliaceae, Amaryllis and others. As an exception, they are found in dicotyledons: some species of wood sorrel and butterwort.
The stem called bottom, in the bulb it is greatly reduced and more or less flattened. It bears leaves in the form of scales, and in the lower part there are adventitious roots. Unlike the rhizome and tuber, the storage organs of the bulb are fleshy scales. Bulbs are formed in tulips, hazel grouse, narcissus, hyacinth, scilla, snowdrop and other plants.
Depending on the development of the lateral buds, the bulbs are simple when the buds are small (onions, lilies, etc.), and complex– the buds in the axils of the dry scales are large, highly developed (garlic).
Corm- an underground modification of a shoot, combining the characteristics of a rhizome and a bulb. It develops from a shoot with a greatly shortened stem, from the lower part of which adventitious roots extend. At the top of the corm or at its base there is a bud from which a flowering shoot is formed. The stem of the corm is covered with the bases of dead last year's leaves, which look like dry filmy scales. From the buds located in their sinuses, new corms develop - children. Corms include saffron, gladiolus, and colchicum.
Modifications of aboveground shoots
Kochan cabbage is essentially a gigantic bud: the middle part of the head of cabbage is occupied by a fleshy stem ( poker), along which there are juicy, fleshy leaves. The smallest and youngest of them are located inside the head of cabbage, around the growth cone with which the stalk ends. Brussels sprouts form small heads of cabbage from the side buds.
In the head of cabbage, both organs of the shoot are involved in the accumulation of nutrients: water-soluble sugars and other biologically valuable compounds, including vitamins, but the main role belongs to the leaves.
Aboveground tubers, developing from the lower part of the main shoot, are characteristic of kohlrabi cabbage.
Small nodules can also develop in the inflorescence area. In the knotweed viviparous, for example, they are formed in the axils of the lower covering leaves of the inflorescence. Their buds begin to sprout on the mother plant. After falling, they take root, forming new plants.
Metamorphosed shoots can serve not only for storing nutrients, reproduction and propagation of plants, but also perform other functions.
In some plants, shoots lose leaves during development, and the stem becomes a photosynthetic organ, called in this case cladodium . It is usually flattened and has the ability to grow for a long time (in the tropical plant Mühlenbeckia, or Homalocladus, from the buckwheat family, prickly pear). The nodes have well-defined constrictions, so the cladodium looks segmented. In humid conditions, cladodes can be deciduous, but in dry weather the leaves fall off. As an organ of photosynthesis, cladodes have well-developed chlorophyll-bearing tissue located under the epidermis.
Cladodia, according to appearance similar to leaves are called phyllocladies . They develop from lateral buds, so they are always found in the axil of a small filmy or scale-like leaf. Phyllocladies are very diverse. In pinnate or bristly asparagus, they are small, needle-shaped, in Sprenger's asparagus, or densely flowered, they are flattened and linear. Broadly lanceolate or ovoid in outline, leathery phyllocladia are characteristic of butcher's broom species found in the Mediterranean, Transcaucasia, and Crimea. In the summer, on the underside of the phyllocladium, in its middle part, an inflorescence develops, located in the axil of the small covering leaf; in the fall and winter, the butcher's broom blooms, and in the spring, orange berries ripen on the phyllocladia.
A fairly common type of shoot change is thorn . This is a highly woody, leafless, shortened shoot with a sharp tip. In hawthorn, the shoot that develops in the leaf axil in spring initially has small leaves and an apical bud. Soon the apical growth stops, the leaves fall off, the stem becomes very lignified and turns into a thorn. Thorns develop in the same way.
Clinging plants are equipped antennae , which, like spines, are modified lateral shoots in some plants. Passionflower has simple, unbranched tendrils with a curled apex. They are found in the axils of the leaves, just like the flowers. The branched tendrils of grapes are modified inflorescences. In girlish grapes, the ends of the branched tendrils are disc-shaped and play the role of suction cups, helping the plant to climb onto a support. Tendrils of shoot origin are also formed in pumpkin, watermelon, cucumber and other members of the pumpkin family.
Primary anatomical structure of the stem.
Definition and functions of the stem.
The stem is an axial, usually radially symmetrical organ with long-term apical (apical) growth.
Functions of the stem:
1. carries out two-way movement of substances between roots and leaves,
2. supports the crown of the plant,
3. helps to increase the total assimilation surface of the plant due to branching,
4. participates in the storage of reserve substances,
5. at a young age – also carries out photosynthesis.
These functions determine the presence in the stem of integumentary, well-developed conductive, mechanical tissues and functionally developed parenchyma.
Structure of the growth cone.
In the cone of growth of the stem of angiosperms, two zones are expressed: outer - tunic, and internal – frame, which differ in the planes of cell division (theory of A. Schmidt and J. Buder) (Fig. 1).
The tunica may consist of one or more layers of cells dividing perpendicular surface, in most dicotyledonous plants it is two-layered. The outer layer of the tunic gives rise to protodermis, from which the epidermis is then formed, covering the leaves and stem. If the tunic is multilayered, then the inner layer forms the tissue of the primary cortex.
In the presence of a single-layer tunic, the entire primary cortex and stele are formed by the corpus. Frame formed by a mass of cells which are divided in all planes.
Leaf primordia most often appear in the second layer of the tunic, and axillary buds are formed in the body.
Figure 1. b) apex of angiosperms.
As a result of the activity of the primary meristems of the apex, the primary anatomical structure of the stem is formed: epidermis, primary cortex, central cylinder and core.
Secondary thickening occurs due to the activity of the cambium.
Primary anatomical structure of the stem.
The structure of the stem formed as a result of the activity of the primary meristems of the apex is called primary. In the stem, with the primary structure, three anatomical and topographic zones can be distinguished:
1. cover tissue,
2. primary cortex
3. central cylinder.
Epidermis protects the stem from evaporation.
Beneath the epidermis is the outer layer of the primary cortex - exodermis - represented by chlorenchyma and (in dicotyledons) collenchyma. In the inner layer bordering the central cylinder - endodermis - usually starch grains accumulate, and then it turns into the so-called starch-bearing vagina (starch-bearing layer). Starch grains can settle in cells under the influence of gravity. Due to this, the endodermis plays an important role in the geotropical ((from the Greek trpos - turn, direction), growth movements of plant stems caused by the directional action of gravity) reaction of the stems
Along the periphery of the central cylinder in pericycle sclerenchyma is located. Sclerenchyma fibers form a continuous layer or are collected near vascular bundles, outward from the primary phloem. Main element central cylinder (stele) - conductive bundles. They are formed from procambial strands and consist of primary xylem and phloem. Procambium differentiation occurs centripetally: phloem is formed from the outer side of the bundle towards its center, and xylem is formed towards it (Fig. 2). The procambium can be completely converted into primary conducting complexes, forming a closed collateral bundle (in monocots), or the remaining cells in the center of the bundle form the cambium. Such open bunches are characteristic of dicotyledonous plants. The work of the cambium ensures the formation of secondary xylem and phloem.
Figure 2.
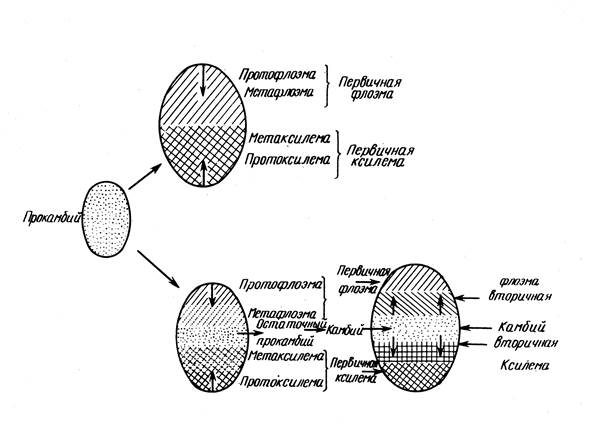
The formation of the procambium determines the structure of the conducting system:
1. If the procambium forms a solid hollow cylinder, then nested cylinders of xylem and phloem are formed from it (in some conifers and dicotyledons) - non-beam structure.
2. If the procambium consists of separate strands (in most monocots, many dicotyledons and conifers), then the emerging primary conducting tissues retain the same bundle structure.
The procambium arises at the base of the leaf primordium, and its development proceeds in two directions: to the top of the leaf primordium and down the stem, where it connects with the cords that arose earlier. The differentiation of phloem and xylem occurs in the same sequence. Typically, phloem elements are formed first, then xylem. Conductive bundles are located in the parenchyma of the central cylinder.
A clearly visible pith develops in the center of the stem. The parenchymal thin-walled cells of the core often loosen, die and fill with air, then the entire tissue appears white (in sunflower, corn, elderberry). Sometimes the core dies very early, when the elongation of the internodes has not yet finished. This entails rupture of the core and the formation of a hollow stem.
The primary structure of the stem is preserved in monocots throughout life, and in dicotyledons and gymnosperms - only in the early phases of its development.
The structure of the stem.
The stem is the axis of the shoot, bearing leaves and buds. The main functions of the stem are supporting and conducting. The stem provides a connection between the roots and leaves. In addition, reserve nutrients are often deposited in the stem. Sometimes the stem is an assimilating organ.
As a result of the activity of procambium and the rest primary meristem The growth cone forms the primary structure of the stem. In the primary stem, a primary cortex and a stele (central cylinder) are usually distinguished. Unlike the root, the primary cortex is covered on the outside with epidermis.
In gymnosperms and most dicotyledonous angiosperms, the growth of the stem in thickness is carried out by the cambium, which forms secondary fabrics. It appears in the form of a cylinder between the primary xylem and the primary phloem and remains in relatively the same position indefinitely, depositing secondary xylem towards the center of the axis, and secondary phloem outwards.
Secondary thickening also occurs as a result of the activity of phellogen (cork cambium).
In tree and shrub dicotyledons, as well as in conifers, secondary thickenings can continue for many years. As a result, the stem has three main parts: bark, wood and pith.
Differences in the structure of the stem of monocotyledonous and dicotyledonous plants.
In monocots, the stem is herbaceous, incapable of secondary thickening, and rarely branches. Vascular bundles without cambium are scattered throughout the stem.
In dicotyledons, the stem is herbaceous or woody, capable of secondary thickening, and branches. The vascular bundles that have a cambium are located in one large mass in the center of the stem or have the appearance of a ring.
15 Functions of a flower.
A flower is a modified shortened shoot adapted for the reproduction of angiosperms (flowering) plants.
The exclusive role of the flower is due to the fact that it combines all the processes of asexual and sexual reproduction, while in lower and many higher plants they are disunited. In a bisexual flower, micro- and megasporogenesis, micro- and megagametogenesis, pollination, fertilization, and the formation of seeds and fruits take place. The structural features of the flower allow listed functions with minimal consumption of plastic substances and energy.
Flower structure
A flower consists of a stem part (pedicel and receptacle), a leaf part (sepals, petals) and a generative part (stamens, pistil or pistils). The flower is attached to the stem by a pedicel. If the peduncle is greatly shortened or absent, the flower is called sessile (plantain, verbena, clover). The peduncle also contains two (in dicotyledons) and one (in monocotyledons) small preleaves - bracts, which may often be absent. The upper expanded part of the peduncle is called the receptacle, on which all the organs of the flower are located. The receptacle can have different sizes and shapes - flat (peony), convex (strawberry, raspberry), concave (almond), elongated (magnolia). In some plants, as a result of the fusion of the receptacle, the lower parts of the integument and the androecium, special structure- hypanthium. The shape of the hypanthium can be varied and sometimes participate in the formation of the fruit (cynarrhodium - rose hip, apple). Hypanthium is characteristic of representatives of the rose, gooseberry, saxifrage, and legume families.
The parts of a flower are divided into fertile or reproductive (stamens, pistil or pistils) and sterile (perianth).
flower functions
It is thanks to flowers that plants live and reproduce. The beautiful shape and color of the flower, first of all, performs an important function for their reproduction - they attract insects or pollinating birds. After pollination, seeds are formed in the ovary of the flower, which will give life to new plants. Not all plants produce flowers, and not all flowers can boast attractive shapes and colors. However, those that are colorful in appearance attract potential pollinators. It is not uncommon for a particular plant to be pollinated by a specific species of insect that has the organs needed to pollinate them. The most famous insect pollinators are bees and flies. Some flowers are pollinated by birds, such as hummingbirds. In wind-pollinated plants, the flowers are small and inconspicuous in appearance, since their shape and color cannot have any effect on the pollination process. Some flowers have, in the opinion of people, an unpleasant odor, for example, stapelia, the smell of which is reminiscent of the smell of decaying meat.
16 question)))
Wind-pollinated plants usually have a number of characteristic features: very small and numerous flowers, produce a lot of pollen. One plant can produce millions of pollen grains. In many wind-pollinated plants (hazel, aspen, alder, mulberry), flowers appear even before the leaves bloom.
Bees prefer to collect pollen from insect-pollinated plants. But if there are few flowering entomophilous plants in nature, and bees need pollen, they also collect it from wind-pollinated plants.
Pollination of flowering plants is carried out in two main ways - by wind and animals, most often insects. In both cases, plants develop specific adaptations. Insect-pollinated plants are characterized by large, brightly colored single flowers, as well as inflorescences consisting of bright flowers various shapes. As a rule, they have a strong odor. They have developed special glands - nectaries, which produce a sweet liquid secretion - nectar. The flowers of insect-pollinated plants are rich in pollen. Pollen grains are usually large and sticky, and their shell often has various projections. In wind-pollinated plants, the perianth is partially or completely reduced, and their small and inconspicuous flowers are usually collected in inflorescences. Wind-pollinated plants produce much more pollen than insect-pollinated plants. However, their pollen grains are small and dry, well transported by the wind. They are formed in large anthers, often hanging on long filaments. The stigma of many of them is forked and lined with numerous hairs and bristles, which allows them to capture much more pollen from the air. Many wind-pollinated plants bloom in early spring even before the leaves bloom. Wind-pollinated grasses that bloom in summer bloom strictly according to the clock, thereby saving pollen. The likelihood of pollination is also increased by the fact that all wind-pollinated plants grow in close groups or large clusters.
17. Fruits and seeds. Classification, methods of distribution
Fetus. It is one of the most characteristic organs of angiosperms. It consists of pericarp and seeds. The pericarp, which is an overgrown and highly modified wall of the ovary, ensures the formation of seeds, protection from unfavorable factors, and promotes their spread.
Juicy fruits have well-developed pulp, in the large parenchyma cells of which a lot of water, carbohydrates, vitamins, organic acids, and various aromatic compounds accumulate.
In dry fruits pericarp leathery or woody. They can be single-seeded or multi-seeded, dehiscent or indehiscent. In many fruits, dehiscence of the dry pericarp allows seed dispersal. Dry multi-seeded fruits are often opened using holes, caps, and flaps.
Infertility- this is a collection of mature, fused fruits (dry or juicy) of one inflorescence (mulberry, breadfruit, pineapple).
Flowering plants have evolved various adaptations for dispersing seeds and fruits. Many plants have fruits that spread by the wind. They are small, light, and often equipped with wing-like appendages or flies (dandelion, thistle).
The fruits of some plants spread water(alder, water lily, coconut palm, many types of sedges). Juicy fruits with a bright color and aromatic smell are readily eaten by many species of animals. The seeds of these fruits are not digested and end up in the soil along with excrement. Dry fruits develop various attachments, hooks that cling to animal hair and human clothing and are carried by them (burdock, Velcro, string). In some plants, ripe fruits crack and the unwinding valves of the pericarp discard seeds - sometimes over a considerable distance (impatiens, mad cucumber).
Seed. Typically, a seed consists of an embryo and endosperm surrounded by a seed coat (Fig. 8.22).
Testa formed from the integument of the ovule. She protects the seed from mechanical damage, penetration of pathogenic microorganisms, excessive water loss. The seeds of many plants have various devices for dispersal - hairs (willow, poplar), fleshy appendages (to attract animals).
Germ - this is the rudiment of a new individual, a miniature sporophyte. In most flowering plants, the embryo consists of a germinal root, a stalk and a bud. Cotyledon leaves are attached to the top of the stem (cotyledons). In this case, in dicotyledonous plants two cotyledons are formed (they contain reserve substances of the seed), in monocotyledonous plants - one. The single cotyledon of the cereal embryo is called shield. It is located on the border with the endosperm and during germination of the grain it facilitates the flow of dissolved nutrients from the endosperm to the embryo.
Endosperm- tissue containing nutrients. It is formed not only in almost all monocotyledonous plants, but also in a number of dicotyledonous plants (flax, dill, carrots, etc.). The trichuid cells of the endosperm, as well as the cotyledons of the embryo, contain a significant supply of nutrients (starch, proteins, fats, etc.) necessary for the development of the embryo and the formation of the seedling.
Seed classification simple. There are five types of seeds depending on where the reserve products accumulate: in the endosperm, nucellus, embryo, in the endosperm and nucellus in the endosperm and embryo.
The development of the ovule begins with the appearance of a median tubercle - the nucellus, inside of which one large sporogenic cell stands out in the early stages of development. Later, it undergoes reduction (meiotic) division and forms 4 unequal cells - megaspores. Therefore, the nucellus should be considered a megasporangium. The nucelus forms the central part of the ovule
The seed has a very important feature: in conditions unfavorable for germination, it can remain dormant for a significant period of time. With the onset of favorable conditions of temperature and humidity, the seed begins to absorb water and, with sufficient access to air, germinates. Seeds are spread by water, wind, animals, random distribution (mad cucumber..classmates attention: when the seeds of the mad cucumber reach full maturity, the surrounding fleshy tissues turn into a slimy mass.
When separating a cucumber from the stalk, pressure is created on the contents of the fruit according to the principle jet thrust, which moves the fruit in the direction opposite to the flight of the seeds, which contributes to the scattering of seeds over a larger area.
18. Lichens. Structural features, distribution, development cycle, significance in nature and human life. Representatives
Lichens- This is an example of an obligate symbiosis of fungi and algae. According to the nature of sexual sporulation, lichens are classified into two classes: marsupials (reproduce by spores that ripen in pouches), which includes almost all varieties of lichens, and basidial (spores ripen in basidia), numbering only a few dozen species. According to body structure ( thalli) there are crustose (crust), foliose and fruticose lichens. The vegetative body of crustose lichens is the most primitive; it can be granular, porous or in the form of crusts. More developed are foliose lichens, which have the appearance of more or less dissected plates. Fruticose lichens are highly organized, having the appearance of bushes, hanging threads or erect outgrowths.
Spreading. Lichens grow on the soil ( epigeic), stones ( epilithic) or tree trunks ( epiphytic), receiving the moisture necessary for life from the atmosphere. Some species live in the marine littoral zone. When they first settle in barren places, lichens form humus when they die, on which other plants can then settle. Lichens have even been found in barren Arctic deserts and inside Antarctic rocks. Lichens are distributed throughout the world, but are especially diverse in the tropics, highlands and tundra. But in laboratories, lichens die quite quickly. And only in 1980, American scientists were able to “combine” an algae and a mushroom grown from a spore. Lichens are bioindicator organisms; they grow only in environmentally friendly places, so you won’t find them in big cities and industrial areas.
Reproduction. Lichens reproduce mainly by vegetative means - by parts of the thallus, as well as by special specialized education- soredia and isidia (Fig. 6.4).
Soredia are formed under the upper bark in the photosynthetic layer and consist of one or several phycobiont cells entwined with fungal hyphae. Under the pressure of the overgrown mass of numerous soredia, the cortical layer of the thallus breaks, and the soredia come to the surface, from where they are carried by wind, water and, under favorable conditions, grow into new lichen thalli.
Isidia They are small outgrowths of the thallus in the form of sticks, tubercles, covered on the outside with bark. They consist of several phycobiont cells entwined with fungal hyphae. Isidia break off and form new thalli.
The importance of lichens in the biosphere and national economy. About 26 thousand species of lichens are known. They are widespread in nature, except in places where the air is saturated with harmful gases. Lichens are very sensitive to air pollution and therefore most of them are major cities, as well as near plants and factories, quickly dies. For this reason, they can serve as indicators of air pollution with harmful substances.
Being autoheterotrophic organisms, lichens accumulate solar energy and create organic matter in places inaccessible to other organisms, and also decompose organic matter, participating in the general cycle of substances in the biosphere. Lichens play a significant role in the soil-forming process, as they gradually dissolve and destroy rocks, on which they settle, and due to the decomposition of their thalli, soil humus is formed. Thus, lichens, together with bacteria, cyanobacteria, fungi and some algae, create conditions for other, more advanced organisms, including higher plants and animals.
IN economic activity For humans, an important role is played primarily by food lichens, such as reindeer moss, or moss, Icelandic moss and others, which are eaten not only by reindeer, but also by deer, musk deer, roe deer, and moose. Some types of lichens (lichen manna, hygrophora) are used for food; they have also found application in the perfume industry for the production of aromatic substances, in the pharmaceutical industry for the manufacture of drugs against tuberculosis, furunculosis, intestinal diseases, epilepsy, etc. Lichen acids are obtained from lichens ( about 250 are known) with antibiotic properties.
Representatives: Reindeer moss, bushy beard lichen.
U dicotyledonous plants the stem, before the onset of cambium activity, which produces a radical restructuring of its anatomy, has the following structure: on the outside it is covered epidermis, next comes primary cortex, composed of several rows of predominantly parenchymal cells; further towards the center is located central cylinder, composed of vascular-fibrous bundles and core. Endoderm, separating the bark from the central cylinder, is present in only a few plants.
Epidermis composed of homogeneous cells of the usual structure for the epidermis, tightly pressed together. IN primary cortex two layers are usually distinguished. The outer one is made up of cells tightly connected to each other. These cells have thickenings in the corners of the membranes and represent collenchyma. The inner layer of the cortex is composed of larger, loosely interconnected cells with thin shells. The endoderm (if present) is represented by one layer of large cells containing starch grains (“starch vagina”).
Central cylinder begins pericycle, which is the site of formation of adventitious roots (arising endogenously). Sometimes the pericycle develops into a multicellular mechanical tissue that covers the outside of the vascular bundle. Such pericyclic fibers have thickened, often lignified walls, or they retain their fiber composition (Fig. 105). The main, most important part of the central cylinder is conductive tissue in the form of open vascular-fibrous bundles arranged in a regular ring. They are of the collateral type, that is, their xylem is turned inward, the phloem is turned outward, and between them there is a narrow layer of cambium (Fig. 106). The bundles are separated by groups of parenchyma cells connecting the cortex to the pith. These will be primary medullary rays. Sometimes the vascular bundles are located not in one, but in two layers. In this case, the internal bundles consist only of phloem. When they are closely adjacent to the external bundles, bicollateral bundles arise (in pumpkins).
The core, which occupies the central part of the stem, is composed of large, tightly connected parenchyma cells. Sometimes the core disappears, and instead of it an air cavity forms in the center of the stem (in umbrella).Material from the site