23. The anatomical structure of the stems of herbaceous and woody monocotyledonous plants.
24. The anatomical structure of the stems of herbaceous dicotyledonous plants.
25. The anatomical structure of the stems of woody dicotyledonous plants.
26. The anatomical structure of coniferous stems.
27. The anatomical structure of the rhizomes of monocotyledonous and dicotyledonous plants.
31. The concept of the stem, escape. Functions and metamorphoses of the stem. Morphological characteristics of the stem.
A stem is an aerial, vegetative, axial organ of a plant that has radial symmetry and has apical growth.
Stem with buds and leaves - the escape. Stem sections carrying a leaf - stem nodes: between nodes - internode. Depending on the length of the internodes, the shoots can be elongated (cereals), shortened (dandelion).
An escape develops from a kidney.
Apical bud - the top of the stem, covered with young buds of leaves: a cone of growth, at the base of it there are primary tubercles - primordial leaves are formed from them. In the sinuses of the primary tubercles are secondary tubercles that give side or axillary the kidneys. Lateral branches also grow with their tops and each branch ends with an apical bud.
With the onset of autumn, all buds enter seasonal dormancy. They are called wintering. They can be vegetative (forming leaves or shoots) and vegetatively generative or flowering (spring blossoming into flowers or inflorescences). Protected by hard cuticle coated scales. They have scleroids, sometimes a cork. Blossom due to reserves deposited in the root, stem (or in the hiding scales themselves).
Many axillary buds remain at rest for a long time and grow only with their axis on the stem in accordance with its thickening - sleeping the kidneys. They start to grow if you remove the apical kidney. Sometimes they can be immersed in wood and wake up after a break or a log house.
Subordinate clauses buds are laid in nodes and on internodes, roots, rhizomes, leaves - a reserve of vegetative propagation. Origin: from cambium, pellogen, pericycle, bark. These are rejuvenation buds - they give shoots similar to young seedlings.
Morphological signs of the stem.
I. By position in space stems are: erect, rising, creeping, climbing, climbing, etc.
II. Branching shoots.
1. Dichotomous - the growth point is forked into 2 new ones (lower, fern, bryophytes).
2. Monopodial - the main axis grows at its apex, the lateral branches develop from the lateral buds, which also branch monopodially (conifers - trunks in forestry and shipbuilding are very much valued).
3. Sympodial - the apical bud dies, a lateral bud develops instead, grows often in the vertical direction, as if continuing the growth of the main stem, etc. (birch, willow, apple tree, tomato, potato).
Is known mixed branching: first mono-, then sympodial (in tree fruit, cotton).
Intensive blooming of a large number of buds makes the shoot shortened, which ensures the development of a huge number of leaves on the crown, i.e. large photosynthetic surface.
4. False dichotomous branching - two oppositely located axillary kidneys form under the apical kidney; starting to grow, they form a fork (mistletoe, dope, cloves, lilac, horse chestnut).
The study of branching systems is of great practical importance. You can adjust the crop.
III. Location: the leaves are mounted on stem nodes.
1. Another - the node has one leaf - an apple tree, a birch, a maple.
2. The opposite - two leaves per node (labioecum, valerian, hydrangea).
3. Whorled - three or more leaves - an oleander.
4. A basal rosette is a primrose.
Regular leaves are arranged on the stem in a spiral. Following its turns, you can find two sheets located vertically one above the other. The line connecting the leaves in the longitudinal rows on the stem - orthostic. Between the ends of the orthostomy, the remaining leaves are arranged in a spiral, the number of turns of the spiral between two leaves located on the same orthostich are called - leafy cycle. The next leaf arrangement is made in the form of a fraction, where the number of turns of the spiral in the leaf cycle becomes the numerator, and the number of leaves in the cycle is the denominator.
IV. The shape of the stem in cross section.
The shape of the stem in the cross section is the most diverse: round, flat, ribbed, multifaceted, etc .; can be hollow and made, naked and pubescent.
Life expectancy and biomorphological groups of plants. Cypress - 3000 years, chestnut - 2000 years, oak - 1200 years, linden - 1000 years, poplar - 300-600 years, apple tree - 200 years, sweet cherry - 40-100 years, pine - 50 years. Depending on the life expectancy and type of shoots, the plants are divided into:
the trees - Large plants with highly developed perennial stems. There is a secondary increase.
shrubs - the main trunk is absent or weakly expressed, branching begins almost at the soil surface (4-6). Barberry, irga, wild rose. There are groups of shrubs no more than 1 m - thyme, saxaul.
shrubs - shoots in the lower part remain perennial, in the upper part - annual shoots freeze or dry out - wormwood, astragalus, St. John's wort;
grassy- the death of aboveground stems is observed at the end of the growing season. Most field plants are cereals, peas, beans, flax, hemp;
biennial grassy - in the first year, only a rosette of root leaves develops, and flowers, fruits and an elongated stem - in the second (chicory, belena, marshmallow, melilot)
perennial - durable underground parts and the annual death of aboveground parts (valerian, lily of the valley, nettle).
Among herbaceous plants, there are winter and spring forms:
a) if the seeds germinate in the spring - spring;
b) winter - seed germination in the fall (0-5 degrees).
Stem functions:
1. The skeleton connecting the leaves and roots together gives the habit to the plant;
2. Conducting;
3. Stocking;
4. Synthetic;
5. The organ of vegetative propagation.
Metamorphoses:
Underground: 1. Rhizome - an underground shoot, leaves in the form of scales, the size of the rhizomes is determined by the functions: if for the stock - thick (calamus, horsetail, bought), for reproduction - with renewal buds (wheat grass, killer whale).
2. Tubers - are formed at the ends of underground stems - stalls, have kidneys - eyes (3 or more each).
3. Bulbs - a shortened stem - the bottom, buds and leaves.
Ground: 4. Spines - hawthorn, turn.
5. Mustache - pumpkin, cucumber, melon.
6. Phyllocladius - the stem in the form of a leaf - needle.
Stem - aerial, vegetative, axial organ of the plant; has a long growth in length using the apical meristem in the growth cone or insertion meristem; bears leaves that are laid in a certain order on a growth cone in the form of tubercles; can branch due to buds embedded in axillary leaves; has a radial structure and several planes of symmetry.
At the growing end, the stem carries an apical bud - a growth cone, dressed in young, mutated leaves. It ends with a growth point, behind which the primary tubercles are located - the rudiments of the leaves. Even lower in their sinuses secondary tubercles arise - the rudiments of the lateral shoots.
Hapstein's histogen theory (1818): dermatogen, perible, pleroma - is not true for the stem. At the beginning of the 20th century, Schmidt formulated the theory of tunic and body, according to which the growth cone in angiosperms and part of gymnosperms consists of two histologically different parts with different meristemic activity. The outer part is the tunic, its cells divide perpendicular to the surface of the stem and form the epidermis, sometimes the epidermis and several layers or the entire primary cortex; the inner part - the body - cells divide in all directions and form the remaining layers of the primary cortex and the center.
The primary structure of the stem develops during the differentiation of apical meristem cells.
At the level of the first leaf primordia, tunic and hull cells cease to divide; their peripheral part goes to the formation of the primary cortex, the inner part to the formation of the core. But between them there are several rows of active meristematic cells located in a ring - educational ring. Its cells at the base of young leaf primordia give rise to the primary lateral meristem - prokambia.
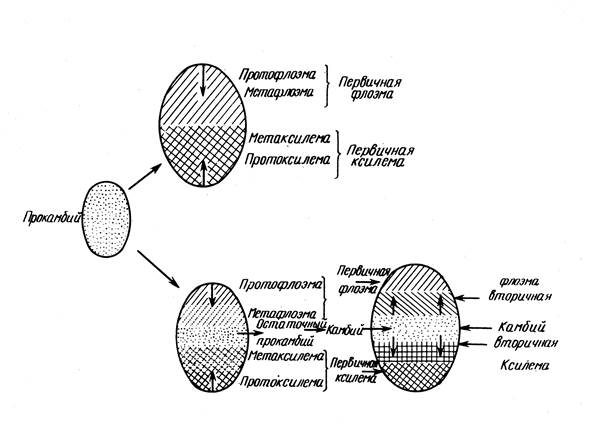
Procambius represents the embryonic phase in the development of the primary conducting meristem, therefore, its location largely determines the subsequent location of the conducting system. If it is laid with a continuous layer (ring), then continuous layers of phloem and xylem arise, if procambium is laid in the form of bundles (strands), then the primary conductive tissues are arranged in the form of bundles. Cells of the peripheral layers of the educational ring that are not involved in the formation of procambium form a pericycle.
Prokambium arises at the base of the leaf embryo and from here its development extends in two directions - acropetally, i.e. to the apex of the leaf primordium, and basipetally, i.e. down the stem, where it is attached to other bundles that arose earlier.
The phloem is formed earlier than the xylem and develops centripetally, first protofloem, then protoxylem (annular and spiral vessels), then metaxylem. In this way, the center of primary structure is formed.
Three groups of tissues are distinguished on the transverse section of the stem: integumentary, primary cortex and CTC.
The integumentary tissue of the stem herbaceous dicotyledonous plant - epidermis. The cells are slightly sinuous, elongated in the direction of the stem, stomata are few or not.
The primary cortex consists of collenchyma in the form of strands in the corners of the stem in the labiate or in its ribs in the umbellate, less often in the form of a ring in pumpkin, nightshade. At the periphery, the primary cortex consists of alternating strips of chlorenchyma and mechanical tissue. In the inner part is a colorless parenchyma. The innermost layer - the endoderm in some cases - is a starchy vagina containing small grains of protected starch.
The peripheral part of the central cylinder is the pericycle. Conductive tissues are arranged in the form of bundles or a continuous ring (depending on the location of the procambium). The transition to the secondary structure is associated with the laying of cambium, depending on which the following types of structure of the stems are distinguished:
1. Procambius is laid in a continuous ring, the primary structure is non-bundled; cambium is formed from procambium; the secondary is non-bunching.
2. Procambius is laid with strands - the primary structure is beam; cambium is formed from procambium in bundles and from the main tissue of the CTC between the bundles. A continuous cambial ring forms: the secondary structure is non-bundled.
3. The primary and secondary structure are beam-like, because inter-beam cambium does not form.
4. The primary and secondary structure are beam-like, because procambium is laid in separate cords, and the resulting inter-beam cambium forms the main tissue of C.O.C., and not the phloem and xylem.
The stem of monocotyledonous herbaceous plants.The stem of monocotyledonous herbaceous plants is characterized by a bundle structure, the bundles are randomly located, and the absence of secondary growth. The main features of the anatomical structure of the stems are determined by the system of leaf traces: from the base of the leaf into the stem are bundles located around the entire circumference, so all vascular bundles of monocotyledons are leaf traces (palm type). SVPs of monocotyledons have a characteristic shape: two large point vessels (symmetrically) and 1-2 narrower with annular and spiral-annular thickenings. An air cavity adjacent to the extreme of them formed at the site of destruction of the protoxylem elements. The size of the beams increases from the periphery to the center.
There are differences in the structure of the cortex:
1. Monocotyledons with a pronounced primary cortex - (asparagus): consists of several layers of chlorophyll-bearing cells; collenchyma is absent.
2. In other monocotyledons (palm, cereals), the primary cortex is not expressed; immediately under the epidermis there are areas of chlorenchyma surrounded by sclerenchyma, which merges with the sclerenchyma of SVP.
A large central cavity (cereal straw) is formed in the internodes of many monocotyledons, in these cases the bundles are shifted to the periphery, but are also scattered: smaller ones to the periphery, large ones to the cavity.
The structure of the stems of wood dicotyledons.The primary structure is non-powdery. The secondary is associated with the laying of the pellogen and cambium. Cambium forms a secondary phloem and xylem. Phloem - bast - hard and soft is located in the form of trapezoid, separated by the tips of the primary core rays; The sapwood function is 1 mm in cambium. Wood in the form of tree rings; sound wood is a mechanical function.
The core is friable, first living tissue with a supply of nutrients. Connected to the forehead and bark by the core rays (primary and secondary)
Fellogen is formed due to the epidermis and layers of the primary cortex, forms the periderm and crust. Under it, the primary cortex is preserved - collenchyma, chlorenchyma, storage tissue - the endoderm is not expressed (sometimes differentiated into a starchy vagina).
According to the Global Fund X1 (1987), bark in pharmaceutical practice - This is the outer part of the trunks, branches and roots of trees and shrubs, located to the periphery of the cambium. By external signs the whole bark has the appearance of tubular, grooved or flat pieces of various sizes. The outer surface of the cortex with brown or gray corks is usually smooth or with longitudinal (or transverse) wrinkles, sometimes with cracks. The bark of branches and trunks has rounded or oblong lentils, sometimes leaf lichens can be on it (they must be removed during harvesting). The inner surface of the cortex is usually lighter, smoother, or ribbed. Transverse kink is usually uneven: splintering, fibrous or granular.
On a micropreparationpay attention to the outer cortex, located to the periphery from the end of the core rays and consisting of the periderm and the remnants of the primary cortex, and the inner, consisting of the phloem. The thickness, color and nature of the cork, the presence of collenchyma, the ratio of the thickness of the primary and secondary cortex, the width of the core rays, the number, location and structure of bast fibers, stony cells, as well as the inclusion of calcium oxalate, lactic cells, and cells with essential oil are of diagnostic value.
The structure of coniferous stems.The primary is bundle, then a continuous cambial ring and phloem and xylem rings are formed. Pellogen is laid.
In the cow: there are no collenchyma, there are tar passages.
2. In the phloem there are no companion cells, bast fibers, bast parenchyma.
3. The cambium is multirow.
Wood consists of tracheids, mechanical fibers and wood parenchyma are absent.
The primary structure is beam.
The structure of the stems of woody monocotyledonous plants.Peculiar to tree-like representatives of the lily family (dracaena, yucca, aloe). A young stalk near the growth cone is built as in all monocotyledons: integumentary tissue - the epidermis; beneath it is the primary cortex, represented by several layers of lamellar collenchyma; A center begins with a pericycle and contains closed collateral SVPs located randomly. Thickening occurs due to the formation of a thickening ring formed from the pericycle or from the inner layer of the primary cortex. It forms a centrifugal centroxile concentric SVP. The integumentary tissue is cork.
The structure of rhizomes of monocotyledonous plants.Similar to a tree stem. The integumentary tissue is a lignified exoderm; primary cortex is always developed, represented by storage tissue; endoderm typical with Caspari spots and horseshoe-shaped thickenings. In the CSC - collateral bundles (V) and concentric centrofloem. Formed from a thickening ring.
The structure of the rhizomes of dicotyledons.The integumentary tissue is a cork, the periderm is formed from the phallogen primary bark - storage parenchyma; endoderm with Caspari spots or horseshoe-shaped; in the center, the location of the tissues is similar to the ground stem; in the beam type - beam; in transitional and nonpunchy - nonpunchy.
The evolution of the stele.In the process of evolution, a change was observed in the structure and relative position of various plant tissues, especially those conducting. Several evolutionary types of stem structure have been established relating to the center or stela (Van Tihem - French botanist). The stellar theory later received brilliant confirmation in the study of extinct plants (riniophytes). The most ancient type is protostela. The xylem stratum, covered with a layer of phloem, actinostela - xylem has the shape of a star (pluno, horsetail), surrounded by a phloem
Siphon wall - the core appears.
Diktiostela - siphonostela is cut into separate bundles (ferns).
Eustela - open collateral bundles bundles (dicotyledonous).
Atactostela - in monocotyledons closed collateral bundles located randomly.
ConclusionA variety of structural types of the stem and rhizomes must be known for macro- and microscopic diagnosis of medicinal plant materials.
The stem is part of the medicinal plant material - grass (Herba). With macroscopic diagnosis herbs in accordance with the State Pharmacopoeia X1 (1987) note the following features of the stem: type of branching, cross-sectional shape, pubescence, dimensions (length and diameter at the base), leaf arrangement. On micropreparations the cross section of the stem is noted by the structural features of the epidermal cells, the location of the conducting bundles, the presence and nature of the location of mechanical tissues, crystalline inclusions, receptacles, secretory channels, lactis, and other diagnostic features.
Rhizomes (Rhizomata) in the analysis by external signs they differ in shape (simple or branched, cylindrical or oval, distinct, straight, curved or twisted, etc.), features of the outer surface (flat or wrinkled with traces of removed roots), and the nature of the fracture (flat, grainy, splinous or fibrous). Sometimes, with the naked eye, the nature of the arrangement of conductive beams is considered at a break. With microscopic diagnosis note a knot-free or bundle structure, location features and types of bundles, the structure of the integumentary tissue, storage and core, the presence of a variety of containers, mechanical elements, channels, milk stores, calcium oxalate crystals, spare nutrients (starch, mucus, inulin, fatty oil, etc. P.).
In analysis bulbs (Bulbi), tubers (Tubera) and corms (Bulbotubera), shape (spherical, ovoid, oval, oblong, flattened, spindle-shaped, etc.), size, nature of the surface, structural features of the scales (dry or juicy) matter. With microscopic diagnostics note the features of storage tissue and the location of the bundles.
The structure of the stem of dicotyledonous herbaceous plants
In the early stages of development, the dicotyledonous stem has a primary structure. As a result of the activity of the primary meristems of the growth cone, the epidermis, primary cortex, central cylinder and core are formed.
Epidermisthe stem has a small number of stomata, usually covered with cuticles. Under the epidermis are tissues primary cortex.Its outer layer (exoderm) is often formed by the collenchyma, which is located either in a continuous ring (sunflower), or in separate sections. In plants with ribbed stems (pumpkin), the collenchyma fills the protrusions of the stem. In thin stalks of the collenchyma, it is usually angular, in stems with active secondary thickening - lamellar. The main part of the primary cortex (mesoderm) is composed of chlorenchyma and parenchyma. Here, airways and containers of secretions can develop. Cells of the inner layer of the primary cortex (endoderm) are usually filled with large starch grains (starchy vagina).
Central cylinder(stela) consists of a pericycle and a conductive system. The core isfrom the parenchyma, which is often destroyed, and in its place an air cavity is formed. The pericycle is represented by one or more rows of sclerenchyma or its individual cords. The conductive system has the form of a hollow cylinder that separates the primary cortex and core. The conductive bundles that make up this cylinder are separated by inter-bundle parenchyma, the core rays that connect the core and primary cortex. Most dicotyledons are characterized by a special type of central cylinder - the eustellus, on the cross section of which the beams are arranged in a ring (Fig. 4).
Figure 4. Eustel dicotyledons.
In dicotyledonous plants, the layer of procambium cells between the primary phloem and xylem retains the ability to divide and turns into a secondary educational tissue - cambium, which is divided by putting inward elements of the secondary xylem, and to the periphery - the secondary phloem. Bunches of dicotyledonous plants are open, thanks to the work of the cambium, the bunch grows, its diameter increases. The activity of the cambium ensures the emergence of secondary elements in the stem, i.e., the transition from the primary to the secondary anatomical structure.
The secondary structure is characteristic of all dicotyledonous and coniferous plants - grasses and trees. A variety of types of structure is due, first of all, to the location of conductive tissues, which is determined by the laying of Procambius and the activity of cambium.
Procambium in the process of differentiation of the growth cone is laid in the form of cords. If the cords are separated by wide enough rows of parenchyma, then a bundle or transitional structure of the stem is formed; if the prokambium cords are so close that they merge into a cylinder, a non-bundle (solid) structure is formed.
1. The beam structure of the stem found in some herbaceous plants (dill, clover, buttercup, peas, alfalfa). Procambial cords laid in a cone of growth in one circle around the periphery of the central cylinder. Each prokambial cord turns into an open collateral bundle consisting of a primary xylem, a primary phloem and a strip of cambium between them. Cambial cells, dividing, give new (secondary) elements of the conducting beam: inward - xylem to the periphery - phloem. In this case, the youngest sections of the phloem and xylem adjoin the cambium, while the older ones move to the periphery of the beam. The most extreme positions will be occupied by the primary phloem and xylem. Conducting beams are separated by wide inter-beam zones and surround a powerful core.
In most plants, cambium works actively and the diameter of the bundles increases significantly. The activity of beam cambium stimulates the parenchyma, which separates the bundles, which begins to divide, giving a secondary educational tissue. The bundle and inter-bundle meristems close together and form a continuous cambial ring, the activity of which ensures uniform thickening of the stem. If the bundle cambium gives elements of secondary xylem and phloem, then the inter-beam cambium produces a parenchyma (in dill, cereal stellate, etc.). In clover, alfalfa and other plants, the inter-beam cambium lays predominantly sclerenchim-like cells in the direction of xylem. These cells are elongated, thick-walled lignified. Therefore, the delay in harvesting clover reduces the quality of hay. A distinct beam structure is preserved in these plants throughout the life of the stem.
The beam structure has a stalk clover. Under the epidermis is the primary cortex, represented by chlorophyll-bearing parenchyma. The outer row of its cells is the lamellar internal collenchyma - the cells of the starchy vagina (endoderm). The boundary between the primary cortex and the central cylinder is clearly visible due to the sclerichym of pericyclic origin.
The sclerenchyma forms half arcs above open collateral bundles arranged in a circle. The bundle contains a primary phloem adjacent to the sclerenchyma, secondary phloem, cambium, secondary xylem and primary xylem. The beam cambium merges with the inter-beam, forming a solid meristematic ring. The inter-beam cambium lays a lignified parenchyma inside, which connects the conducting bundles. There is a ring (cylinder) of lignified tissues, providing high strength of the stem. However, the conductive tissues remained fragmented in the form of bundles. The stem core is represented by parenchyma.
The bundle structure of the stem can be replaced by a non-bundle solid with age. 2. Transitional structure of the stemcharacteristic of sunflower. Initially, the stem is formed in the same way as in clover, but the inter-beam cambium does not lay the parenchyma, but the xylem and phloem. Additional conductive beams are formed. Gradually, all the bundles can merge into a continuous three-layer (xylem, cambium, phloem) cylinder. In the upper (young) part, the stem has a beam structure, at the base (in the old part) it is non-bunch.
3. Non-beam structure characteristic of the stems of many herbs and trees. In the growth cone of these plants, the procambial cords are so close that they form an almost continuous cylinder (in the cross section it looks like a ring). Differentiating, the pro-cambial cylinder (ring) gives the primary xylem cylinder to the center, the primary phloem cylinder to the periphery, and a cambial cylinder is formed between them.
Secondary changes are associated with the work of the cambium. Cambium cells divide parallel to the surface of the stem, while 10 ... 20 times more cells are deposited inward than outward. Inside the cambium forms the elements of the secondary xylem (wood), outwardly the secondary phloem (bast) is deposited. In some places, the cambium lays on both sides the parenchymal cells of the core rays.
Peculiarities of the non-bunching structure of grass stems can be considered on the example of a stem flax. The stem is covered with an epidermis, under which there is a loose chlorenchyma of the primary cortex. A large number of stomata in the epidermis and the absence of collenchyma in the primary cortex are of significant practical importance, since during the technical flax wash water does not encounter obstacles and easily penetrates the stem. The cells of the starchy vagina (endoderm) are larger and contain well-marked starchy grains.
The central cylinder adjacent to the endoderm begins with groups of sclerenchymal cells - bast fibers, between which parenchyma cells are located. Bast fibers occur in the pericycle and in the primary phloem. They differ in very thick, usually non-lignified walls, consisting of 75 ... 90% of cellulose. The length of the bast fibers is 4 ... 60 mm, the diameter is 12 ... 37 microns. During the growth of a sclerenchymal cell, its nucleus divides, but cytokinesis does not occur, and the protoplast is multinucleated. The cell wall is very thickened, the cavity of the cell is similar to a narrow channel. The cell wall consists of many layers of cellulose and is almost devoid of pores. A mature bast fiber cell is dead. Protoplast after thickening of the cell wall is destroyed. Bast fibers have an unusually high strength. The tensile strength in them corresponds to the resistance of steel, and the elasticity is higher. Bast fiber cells are very closely interconnected. Flax fiber is an excellent spinning material used in the textile industry.
Then in a sequential order are the cylinders (rings): the secondary phloem, cambium, secondary xylem with clearly visible core rays. The primary xylem is located in areas adjacent to the core. The parenchyma of the core is rapidly destroyed (in the process of lengthening the stem), forming a central cavity.
The stems of dicotyledonous herbaceous plants are characterized by:
1. secondary structure, very early on following the primary;
2. the presence of primary integumentary tissue - the epidermis;
3. developed primary cortex, divided into collenchyma (exoderm), parenchyma (mesoderm) and starchy vagina (endoderm);
4. beam or non-beam (solid) structure;
5. the correct arrangement of collateral or bicollateral conducting bundles in one circle around the periphery of the central cylinder;
6. the presence of cambium (open bunches),
7. The core is well developed.
The primary anatomical structure of the stem.
Definition and function of the stem.
The stem is an axial, usually radially symmetrical organ with a long apical (apical) growth.
Stem Functions:
1. carries out two-way movement of substances between roots and leaves,
2. supports the crown of the plant,
3. contributes to an increase in the total assimilation surface of the plant due to branching,
4. participates in the storage of spare substances,
5. at a young age - also carries out photosynthesis.
These functions determine the presence of integumentary, well-developed conductive, mechanical tissues and a functionally developed parenchyma in the stem.
The structure of the growth cone.
In the cone of growth of the stem of angiosperms, two zones are expressed: outer - tunic, and inner - case which differ in planes of cell divisions (theory of A. Schmidt and J. Buder) (Fig. 1).
A tunic may consist of one or more layers of cells dividing perpendicular surface, in most dicotyledonous plants it is bilayer. The outer layer of the tunic gives rise to protodermfrom which the epidermis covering the leaves and stem is then formed. If the tunic is multi-layered, then the inner layer forms the tissues of the primary cortex.
In the presence of a single-layer tunic, the body forms the entire primary bark and stela. Bodyformed by a mass of cells which are divided in all planes.
The rudiments of the leaves most often occur in the second layer of the tunic, and the axillary buds are laid in the body.
Figure 1. b) apex of angiosperms.
As a result of the activity of the primary meristems of the apex, the primary anatomical structure of the stem is formed: the epidermis, primary cortex, central cylinder and core.
Secondary thickening occurs due to the activity of cambium.
The primary anatomical structure of the stem.
The structure of the stem formed as a result of the activity of the primary meristems of the apex is called primary. In the stem, with the primary structure, three anatomical and topographic zones can be distinguished:
1. integumentary tissue,
2. primary cortex
3. The central cylinder.
The epidermis protects the stem from evaporation.
Under the epidermis is the outer layer of the primary cortex - exoderm -represented by chlorenchyma and (in dicotyledons) collenchyma. In the inner boundary layer with the central cylinder, endoderm -starch grains usually accumulate, and then it turns into the so-called starchy vagina (starchy layer).Starch grains under the influence of gravity can settle in the cells. Due to this, the endoderm plays an important role in the geotropic (from the Greek trpos - rotation, direction), growth movements of the plant stem due to the directed action of the Earth's gravity)
On the periphery of the central cylinder in pericyclelocated sclerenchyma. Sclerenchymal fibers form a continuous layer or are collected near conductive bundles, outside of the primary phloem. The main element of the central cylinder (stele) is conductive bundles. They are formed from pro-cambial cords and consist of primary xylem and phloem. Differentiation of the procambium is centripetal: the phloem is formed from the outer side of the beam to its center, and the xylem - towards it (Fig. 2). Procambium can completely turn into primary conducting complexes, forming a closed collateral bundle (in monocotyledons), or preserved cells in the center of the bundle form cambium. Such open bunches are characteristic of dicotyledonous plants. The work of the cambium provides the formation of secondary xylem and phloem.
Figure 2
The laying of a procambium determines the structure of the conducting system:
1. If the procambium forms a continuous hollow cylinder, then xylem and phloem cylinders (in some coniferous and dicotyledonous) are formed from it; non-beam structure.
2. If the procumbia consists of individual strands (for most monocotyledonous, many dicotyledonous and coniferous), then the resulting primary conductive tissues retain the same beam structure.
Prokambium occurs at the base of the leaf anlage, and its development proceeds in two directions: to the top of the leaf anlage and down the stem, where it connects with the bands that arose earlier. In the same sequence is the differentiation of phloem and xylem. Usually phloem elements are formed earlier, then xylem. Conductive beams are located in the parenchyma of the central cylinder.
A clearly visible core develops in the center of the stem. Parenchymal thin-walled core cells are often loosened, die off and filled with air, then all the tissue appears white (in sunflower, corn, elderberry). Sometimes the core dies very early, when the extension of the internodes has not yet ended. This entails a rupture of the core and the formation of a hollow stem.
The primary structure of the stem is preserved in monocotyledonous plants throughout life, and in dicotyledons and gymnosperms, only in the early phases of its development.
The anatomical structure of the stem of a dicotyledonous plant beam structure
The stem is the axis of the shoot. Along with the leaf, it is the main structural part of the shoot. The main functions are support and conductive. Through the stem, a connection is made between the roots and leaves and the exchange of products of air and mineral nutrition. In addition, reserve nutrients are often deposited in the stem. The stem, like the entire shoot as a whole, is an “open” growth system, i.e. it grows for a long time, and new leaves appear on it.
The stem as part of the shoot has a system of meristems supporting the growth of tissues in length and thickness. Growth in length is due to the apical and intercalary meristems, and in thickness in dicotyledons due to lateral secondary meristems - cambium and pellogen.
The primary structure of the stem develops as the cells of the apical shoot meristem differentiate. The apical meristem of dicotyledonous shoot rather early differentiates into several groups of cells that differ in the features of division and the degree of meristematic activity. Its outer layers are transformed into a protoderm, the cells of which later form the primary integumentary tissue - the epidermis.
Fig. 1. Beam type with interbeam cambium in the stem of kirkazona (Aristolochia clematitis) in a cross section (I) and a diagram of the structure of the stem at different levels (II)
A is a slice at the level of occurrence of procambium; B - at the level of occurrence of cambium; B - at the level of the formed structure. 1 - procambium, 2 - epidermis, 3 - collenchyma, 4 - cortical parenchyma, 5 - endoderm (3-5 - primary cortex), 6 - pericycle sclerenchyma, 7 - phloem, 8 - xylem, 9 - bundle cambium (7-9 - open collateral bundle), 10 - inter-beam cambium,11 - core beam, 12 - parenchyma of the core (6-12 - central cylinder)
At the base level of the first leaf primordia, apical meristem cells located to the periphery and in the center of the apex cease to actively divide, increase in size and vacuolize. From these cells, the primary cortex and core are formed. Many dicotyledons in the initial ring differentiate the circle of isolated pro-cambial cords. The cells of the initial ring located between these strands later differentiate into parenchymal elements. On transverse sections, these rays have the form of radial bands connecting the core to the primary cortex.
Procambium in a number of dicotyledons can also be laid in the form of a continuous ring. It can develop throughout the entire thickness of the initial ring or form from its part. Procambius is the precursor of primary conductive tissues: the primary phloem and primary xylem. The phloem begins to form earlier and is laid in the outer parts of the pro-cambial cords or pro-cambial rings. The phloem develops centripetally, i.e. the very first elements occupy the outer position, and the latest - the internal. Xylem is laid in the interior of the Procambius and develops centrifugally. Thus, the phloem and xylem are formed towards each other. The first elements of xylem are proxylem, narrow, relatively thin-walled vessels or tracheids with spiral or annular secondary thickenings. Metaxylem is formed somewhat later than protoxylem and consists of staircase and porous vessels. The main factor controlling the differentiation of both the phloem and xylem is the auxin phytohormone produced by leaf primordia and moving along the procambium from the apex to the base. The primary phloem and xylem formed from Procambius constitute the basis of the axial cylinder, or stela.
The stela, which occupies the central part of the stem, consists of conductive tissues, a core, a pericycle, and those permanent tissues that arise from it. To the outside of the recycle is the primary cortex, which consists of a parenchyma, often collenchyma and sometimes secretory elements. The core usually consists of relatively thin-walled parenchymal cells. Spare nutrients are often deposited in the core. Idioblasts are often found here, i.e. individual cells filled with tannids, crystals, mucus, etc. Sometimes part of the core is destroyed and a cavity forms. The peripheral part of the core adjacent to the xylem is called the perimedullary zone.
The secondary thickening of the stem in most dicotyledons occurs quite early, which leads to the formation of the secondary body of the plant. These changes are mainly associated with the activity of the lateral secondary meristem — the cambium, and partly another secondary meristem — the phallogen. Due to the appearance of secondary tissues, dicotyledonous plants grow in thickness. Secondary changes in the central cylinder begin with the laying of cambium. Cambium arises from the remains of Procambius, on the border of the primary xylem and phloem.
Cambium cells are highly vacuolated and elongated in the vertical or horizontal direction. The first - fusiform initials give conductive elements of conductive tissues, the second - ray initials - form horizontally oriented radiation cells of the core rays. Cambium initial cells are capable of two types of division - periclinal and anticlinal. In the first case, the cell plate is laid parallel to the surface of the stem, in the second - perpendicularly. The result is a continuous series of derivative cells stretching from the cambium radially outward and inward. Cells deposited towards the outer surface and the stem gradually differentiate into the secondary phloem, toward the core into the secondary xylem.
The activity of cambium is activated by the phytohormones gibberillin and auxin coming from the kidneys and young leaves. Ultimately, in the structure of the stem of the annual shoot of a dicotyledonous plant, it is possible to distinguish a modified central cylinder, including permanent tissues arising from the pericycle, the remains of the primary and secondary phloem, cambium, secondary and the remnants of the primary xylem and the core. The modified central cylinder is surrounded by the remnants of the primary cortex.
Secondary rootThe root is the main organ of a higher plant. It performs the function of mineral and water nutrition. Another important function is the fixing, "anchoring" of the plant in the soil. Through the root, plants absorb water and ions of mineral salts dissolved in it from the soil. The roots also carry out the biosynthesis of a number of secondary metabolites, in particular alkaloids. The root is capable of metamorphosis. Most often, it is a place to store spare nutrients. Sometimes it plays the role of a respiratory organ, can serve as an organ of vegetative propagation.
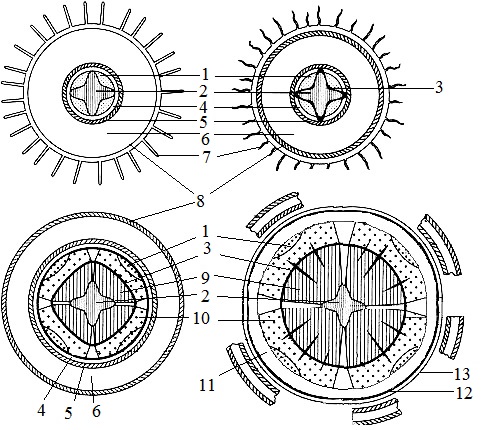
Fig. 2. The transition from the primary structure of the root to the secondary
The formation of the secondary structure of the root is primarily associated with the activity of cambium, which ensures the growth of the root in thickness. The cambium initially arises from thin-walled parenchymal cells in the form of fragmented areas on the inside of the phloem cords between the rays of the primary xylem. Some parts of the pericycle, which are located outward from the rays of the primary xylem, soon acquire cambial activity. As a result, a continuous cambial layer is formed.
Cells of the secondary xylem lay to the center of the cambium, and cells of the secondary phloem to the periphery. The cells of the cambium, which are embedded in the pericycle, form broad radial light rays of the parenchyma located between the cords of the secondary conductive tissue. These rays, sometimes called primary core rays, provide a physiological connection between the central part of the root and the primary cortex. Later, secondary core rays that "bind" the secondary xylem and phloem can also be laid.
As a result of the activity of the cambium, the primary phloem is pushed outward, and the “star” of the primary xylem remains in the center of the root. Its "rays" persist for a long time, sometimes until the end of the life of the root. In addition to the secondary changes occurring in the central cylinder, significant changes occur in the primary cortex. Due to the rapid growth of secondary tissues from within, causing a strong thickening of the root, the primary cortex is often torn. By this time, the cells of the pericycle, dividing around the entire circumference of the axial cylinder, form a wide zone of parenchymal cells, in the external part of which the phallogen is laid, which lays the cork outside and the phalloderm inside. The cork isolates the primary cortex from conductive tissues, the cortex dies and is discarded. Cells of the phalloderm and parenchyma, formed due to the pericycle, subsequently grow and form the parenchymal zone surrounding the conductive tissues. Sometimes this zone is called the secondary crust. Outside, the roots of dicotyledons, having a secondary structure, are covered by the periderm. Crust is rarely formed, only on old tree roots.
In the early stages of development, the dicotyledonous stem has a primary structure. As a result of the activity of the primary meristems of the growth cone, the epidermis, primary cortex, central cylinder and core are formed.
Epidermisthe stem has a small number of stomata, usually covered with cuticles.
Under the epidermis are tissues primary cortex.Its outer layer is often formed by a collenchyma, which is located either in a continuous ring (sunflower), or in separate sections. The main part of the primary cortex is composed of chlorenchyma and parenchyma. Here, airways and containers of secretions can develop. The cells of the inner layer of the primary cortex are usually filled with large starchy grains (starchy vagina).
Central cylinderconsists of a pericycle and a conductive system. The core isfrom the parenchyma, which is often destroyed, and in its place an air cavity is formed. The pericycle is represented by one or more rows of sclerenchyma or its individual cords. The conductive system has the form of a hollow cylinder that separates the primary cortex and core. The conductive bundles that make up this cylinder are separated by inter-beam parenchyma interlayers - the core rays that connect the core and primary cortex. Most dicotyledons are characterized by a special type of central cylinder, on the cross section of which the beams are arranged in a ring (Fig.).
Drawing. The stem of dicotyledonous plants.
In dicotyledonous plants, the layer of procambium cells between the primary phloem and xylem retains the ability to divide and turns into a secondary educational tissue - cambium, which is divided by putting inward elements of the secondary xylem, and to the periphery - the secondary phloem. Bunches of dicotyledonous plants are open, thanks to the work of the cambium, the bunch grows, its diameter increases. The activity of the cambium ensures the emergence of secondary elements in the stem, i.e., the transition from the primary to the secondary anatomical structure.
The secondary structure is characteristic of all dicotyledonous and coniferous plants - grasses and trees. A variety of types of structure is due, first of all, to the location of conductive tissues, which is determined by the laying of Procambius and the activity of cambium.
Procambium in the process of differentiation of the growth cone is laid in the form of cords. If the cords are separated by wide enough rows of parenchyma, then formed bundle or transitional structure of the stem; if the prokambium cords are so close that they merge into a cylinder, then non-bundle (solid) structure.
Beam structure of the stem found in some herbaceous plants (dill, clover, buttercup, peas, alfalfa). Procambial cords laid in a cone of growth in one circle around the periphery of the central cylinder. Each prokambial cord turns into an open collateral bundle consisting of a primary xylem, a primary phloem and a strip of cambium between them. Cambial cells, dividing, give new (secondary) elements of the conducting beam: inward - xylem to the periphery - phloem. Conducting beams are separated by wide inter-beam zones and surround a powerful core.
In most plants, cambium works actively and the diameter of the bundles increases significantly. The activity of beam cambium stimulates the parenchyma, which separates the bundles, which begins to divide, giving a secondary educational tissue. The bundle and inter-bundle meristems merge and form a continuous cambial ring, the activity of which ensures uniform thickening of the stem. If the bundle cambium gives elements of secondary xylem and phloem, then the inter-beam cambium produces a parenchyma (in dill, cereal stellate, etc.). In clover, alfalfa and other plants, the inter-beam cambium lays predominantly sclerenchim-like cells in the direction of xylem. These cells are elongated, thick-walled lignified. Therefore, the delay in harvesting clover reduces the quality of hay. A distinct beam structure is preserved in these plants throughout the life of the stem.
The bundle structure of the stem can be replaced by a non-bundle solid with age. Transitional structure of the stemcharacteristic of sunflower. Initially, the stem is formed in the same way as in clover, but the inter-beam cambium does not lay the parenchyma, but the xylem and phloem. Additional conductive beams are formed. Gradually, all the bundles can merge into a continuous three-layer (xylem, cambium, phloem) cylinder. In the upper (young) part, the stem has a beam structure, at the base (in the old part) it is non-bunch .
Non-beam structure characteristic of the stems of many herbs and trees. In the growth cone of these plants, the procambial cords are so close that they form an almost continuous cylinder (in the cross section it looks like a ring). Differentiating, the pro-cambial cylinder (ring) gives the primary xylem cylinder to the center, the primary phloem cylinder to the periphery, and a cambial cylinder is formed between them.
Secondary changes are associated with the work of the cambium. Cambium cells divide parallel to the surface of the stem, while 10 ... 20 times more cells are deposited inward than outward. Inside the cambium forms the elements of the secondary xylem (wood), outwardly the secondary phloem (bast) is deposited. In some places, the cambium lays on both sides the parenchymal cells of the core rays.
Peculiarities of the non-bunching structure of grass stems can be considered on the example of a stem flax. The stem is covered with an epidermis, under which there is a loose chlorenchyma of the primary cortex. A large number of stomata in the epidermis and the absence of collenchyma in the primary cortex are of significant practical importance, since during the technical flax wash water does not encounter obstacles and easily penetrates the stem. The cells of the starchy vagina (endoderm) are larger and contain well-marked starchy grains.
The central cylinder adjacent to the endoderm begins with groups of sclerenchymal cells - bast fibers, between which parenchyma cells are located. Bast fibers occur in the pericycle and in the primary phloem. They differ in very thick, usually non-lignified walls, consisting of 75 ... 90% of cellulose. A mature bast fiber cell is dead. Protoplast after thickening of the cell wall is destroyed. Bast fibers have an unusually high strength. The tensile strength in them corresponds to the resistance of steel, and the elasticity is higher. Bast fiber cells are very closely interconnected. Flax fiber is an excellent spinning material used in the textile industry.
Then in a sequential order are the cylinders (rings): the secondary phloem, cambium, secondary xylem with clearly visible core rays. The primary xylem is located in areas adjacent to the core. The parenchyma of the core is rapidly destroyed (in the process of lengthening the stem), forming a central cavity.
The stems of dicotyledonous herbaceous plants are characterized by:
1. secondary structure, very early on following the primary;
2. the presence of primary integumentary tissue - the epidermis;
3. developed primary cortex, divided into collenchyma (exoderm), parenchyma (mesoderm) and starchy vagina (endoderm);
4. beam or non-beam (solid) structure;
5. the correct arrangement of collateral or bicollateral conducting bundles in one circle around the periphery of the central cylinder;
6. the presence of cambium (open bunches).
Escape metamorphoses.
If additional functions begin to prevail over the main ones, or an organ acquires some new functions in the process of development, it changes very morphologically and anatomically. Such modifications of the organs are called metamorphoses(from the Greek. metamorphosis - transformation, transformation). Bodies having the same origin, but performing different functions are called homologous . Homologous organs are rhizomes, tubers, bulbs, thorns of hawthorn, cladoda, phyllocladius, tendrils of grapes and pumpkins, as all of them historically developed from a leaf-stem shoot.
Bodies that perform the same functions and are similar morphologically, but having a different origin, are called similar . Root cones and shoot tubers, shoot spikes of hawthorn and leaf spines of barberry, leaf tendrils of peas and shoot shoots of pumpkin are similar.
Modifications of underground shoots.
Underground shoots are widespread in nature: rhizomes, tubers, corms, bulbs. They participate in the supply of substances and the vegetative propagation of plants.
Rhizomeis an escape with scaly leaves, buds and adnexal roots. On the surface of the rhizome, scars from last year's bottom leaves and dead shoots are clearly visible. Rhizome has many subordinate roots. Usually underground rhizomes. Kidneys developing into aerial shoots. Rhizomes are characteristic of wheatgrass, iris, water lilies.
Stolon - elongated thin shoot with underdeveloped leaves. Unlike rhizomes, it is usually short-lived. In muscatus, sour oxides, stolons are involved in the accumulation of nutrients deposited in very thickened grassroots, but the main function of the stolon is to participate in vegetative propagation.
Tuberin contrast to the rhizome and stolon, it is greatly shortened and thickened. Spare substances in it are localized in the stem parenchyma.
Tubers can develop on rhizomes, stolons, the main shoot and other parts of plants. They can be underground and aboveground.
Powerful rhizome tubers with a lot of starch are characteristic for taro, or edible colocasia from the aroid family, which is cultivated in tropical and subtropical countries.
At potato tubers are formed at the end of the underground stolons - whitish fragile shoots with scaly leaves developing from axillary buds located at the base of leafy shoots. On the surface of the tuber are axillary kidneys - eyessitting in small pits framed on one side eyebrows -the remains of the bases of the leaves.
Tubers jerusalem artichoke, or earthen pearare also formed on the stolons. Conductive tissues in them are poorly developed. The tubers are rich in inulin, which is deposited mainly in the stem parenchyma.
Tubers that form on stolons and rhizomes are short-lived. They last only one year, between two vegetative periods, dying off after the development of foliage shoots from their buds, consuming nutrients until the roots are formed, and the plants do not switch to self-nutrition.
Bulb, like a tuber, it is a specialized, modified shortened shoot that serves not only for storing nutrients, but also for transferring adverse periods of the year, vegetative renewal and reproduction of plants. Bulbs are characteristic of monocotyledonous plants from the families of lily, amaryllis and others. As an exception, they are found in dicotyledons: some types of sour and ginger.
Stem called the bottom,in the bulb is strongly reduced and more or less flattened. It carries leaves in the form of scales, and in the lower part - accessory roots. Unlike rhizomes and tubers, fleshy scales serve as storage organs of the bulb. Bulbs are formed in a tulip, hazel grouse, daffodil, hyacinth, bluebell, snowdrop and other plants.
Depending on the development of the lateral kidneys, the bulbs are simplewhen the kidneys are small (onions, lilies, etc.), and complicated - kidneys in the sinuses of dry scales are large, highly developed (garlic).
Corm- Underground modification of the shoot, combining the signs of rhizomes and bulbs. It develops from the shoot with a strongly shortened stem, from the lower part of which the adventitious roots depart. At the top of the corm or at its base is a kidney, from which a flower-bearing shoot is formed. The stem of the corm is covered with the bases of dead last year's leaves, which look like dry membranous scales. Of the kidneys in their sinuses, new corms develop - children. Corms have saffron, gladiolus, colchicum.
Modifications of aboveground shoots
Head outcabbage is essentially a kidney of gigantic proportions: the meaty stem occupies the middle part of the head of cabbage ( stoker) along which succulent fleshy leaves are located. The smallest and youngest of them are inside the head of the head, around the growth cone, which ends with the stump. Brussels sprouts produce small heads of cabbage from the side buds.
In a head of cabbage, both escape organs are involved in the accumulation of nutrients: water-soluble sugars and other biologically valuable compounds, including vitamins, but the main role is played by leaves.
Overhead tubers, developing from the bottom of the main shoot, characteristic of kohlrabi cabbage.
Small nodules can also develop in the field of inflorescence. In the Highlander viviparous, for example, they are formed in the sinuses of the lower covering leaves of the inflorescence. Their buds begin to germinate already on the mother plant. After falling off, they take root, forming new plants.
Metamorphosed shoots can serve not only for storing nutrients, reproduction and reproduction of plants, but also perform other functions.
In some plants, shoots during the development process lose their leaves, and the stem becomes a photo-synthesizing organ, called in this case treasure . Usually it is flattened and has the ability to grow for a long time (in a tropical plant, mullenbekia, or homalocladus, from the buckwheat family, prickly pear). In the nodes, the constrictions are well pronounced, so the cladode looks articulated. In humid conditions, cladodes can be foliage, but in dry weather the leaves fall off. As an organ of photosynthesis, cladody has a well-developed chlorophyll-bearing tissue located under the epidermis.
Cladody, in appearance similar to leaves, is called phyllocladies . They develop from the lateral buds, so they are always in the bosom of a small membranous or scaly leaf. Phyllocladies are very diverse. In asparagus pinnate, or bristly, they are small, needle-shaped, in asparagus Sprenger, or densely flowered, they are flattened, linear. Leathery lanceolate or ovoid outlines leathery phallocladias are characteristic of the needle species found in the Mediterranean, Transcaucasia, and Crimea. In summer, on the underside of phyllocladium, in its middle part, an inflorescence develops, located in the bosom of the small covering leaf, in the winter and winter, the needle blooms, and in spring orange berries ripen on the phyllochladia.
A fairly common type of shoot change is thorn . This is a very ligniferous leafless shortened shoot with a sharp tip. In hawthorn, an shoot that develops in the spring in the leaf sinus first has small leaves and an apical bud. Soon the apical growth ceases, the leaves fall off, the stem is very lignified and turns into a thorn. The thorns of the turn also develop.
Clinging plants are equipped antennae , which, like thorns, are in some plants modified side shoots. Passiflora have simple, unbranched antennae, with a twisted tip. They are in the axils of the leaves, like flowers. Branched tendrils of grapes are modified inflorescences. In girlish grapes, the ends of the branched antennae are disc-shaped and play the role of suction cups to help the plant climb to the support. Antennas of shoot origin are also formed in pumpkins, watermelons, cucumbers and other representatives of the pumpkin family.