Stem. Morphology and function of the stem.The stem is an elongated shoot of higher plants, which serves as a mechanical axis, also serves as a conducting and supporting base for leaves, buds, flowers. Stems perform two main functions - the carrier and the leading. Stems connect all parts of the shoot into a single system. Thanks to the stem, an increase in the surface of the plant is ensured by its branching, formation, and the arrangement of leaves and generative organs is more advantageous. The stem provides an upward and downward movement of water and substances dissolved in it. Young shoots perform the function of photosynthesis. Modified stems also perform additional functions, for example, the accumulation and storage of nutrients in perennial stems, as well as storing water in specialized stems, hooks of other plants, protection, and ensuring the vegetative propagation of plants.
The stems of plants are very diverse and differ in many ways. According to the consistency, the stems are: grassy or woody, hollow (straw) or filled (with a core). According to the cross-sectional shape, the stems are round or cylindrical (in most plant species), tetrahedral (species of the family Gluchokropivovi, norinaceous), trihedral (species of the sedge family ), multifaceted (species of quinoa, sorrel), flattened (for example, bluegrass squinted, segmented arbor). According to the nature of growth and spatial distribution, the following types of stems are distinguished: erect (strong, mainly orthotropic stems with well-defined internodes, most species of higher plants) creeping (mainly plagiotropic stems are rooted in nodes with well-defined internodes; for example, tenacious creeping, strawberries, creeping buttercup, goose cinquefoil) curly (twist around the support clockwise or vice versa; for example, ordinary hop, field bindweed, dodger, ordinary beans) climbing or thick ones (stems with poorly developed mechanical tissue are caught with the help of antennae; for example, cucumbers, common squash, sowing peas) creeping (stems with rather poorly developed mechanical tissue, creeping along the surface of the soil: for example, hernius naked, bryozoans lying, St. John's wort, insole ) ascending (the lower part of the stem spreads along the surface of the soil, the upper one is directed vertically; for example, thyme, asterisk, Veronica officinalis) shortened (the stem is poorly expressed, with very close nodes, short internodes and Turning to rosette leaves, flower forms an arrow; e.g. plantain, dandelion, daylilies, snowdrop). Modification of the stems.
A modified stem is an organ designed to store the reserve nutrients that a plant needs to survive a dormant period. In addition, modified stems are often involved in the reproduction and distribution of plants. To be assigned to this group, a modified stalk must have all the features of an ordinary stalk, that is, have a characteristic structure for it, which means the presence of a stalk proper with an apical growth point and leaves arranged in a spiral (alternately or opposite) and bearing buds in their sinuses . The difference between the altered stalk and the normal one is that it does not necessarily grow above the ground, its growth is often more or less horizontal and usually it is a container of reserve nutrients.
There are six main types of modified stems: tuber rhizome, corm, bulb, onion, offspring
The emergence of primary tissues and the primary anatomical structure.The anatomical structure of the plant stem is due to its main functions. It is characterized by the development of mechanical and conductive plant tissues. In addition, the stem is characterized by a complex system of meristems - apical, lateral and insertive, which determine its growth over a long time and the emergence of new organs.
The plant stem arises from the apical meristem, from which three layers of tissues are differentiated: integumentary, conductive, main. First, these tissues are represented by the so-called primary meristem (meristem), consisting of the protoderm, procambium and the main meristem. The primary conductive tissues develop from procambium. The first elements of the phloem are differentiated from external, located to the periphery of the cells of Procambium. The primary phloem is represented by thin-walled, short-lived elongated cells and is called protofloem, and its external cells can be represented by mechanical fibers.The primary elements of the xylem - tracheids, less often vessels with annular and spiral thickening of the walls - arise later from the prokambium internal cells and are generally defined as protoxylem. In addition to the conductive elements, it consists of parenchymal cells. Later, a structure more or less typical of the phloem is differentiated from the protofloem. Outside of protoxylem, a metaxylem is formed, consisting of tracheids or tracheas with more thickened lignified walls.
Thus, due to the activities of procambium and the rest of the apex meristem, the primary structure of the plant stem arises. In monocotyledonous plants, the entire procambium differentiates into elements of primary conducting tissues. The stems of monocotyledonous, especially herbaceous (cereals), in comparison with the stems of dicotyledonous plants have a simpler structure, they are mostly primary in structure. In dicotyledonous plants, cambium forms in the middle part of the pro-cambial cord and the formation of secondary conductive tissues (metaphloem and metaxilema) begins , the volume of which increases due to the division of cambium cells. Beams consisting of only primary tissues, as in monocotyledons, closed-, bundles with cambium, characteristic of dicotyledons, are open. For herbs grained monocots characterized by diffuse distribution of vascular bundles. Conducting bundles are closed, collateral, less often concentric. Of the mechanical tissues, sclerenchyma is the most developed; collenchyma is found in few plants. There is no secondary thickening in grassy monocotyledons.
Rounded (e.g. sunflower)
Tetrahedral
Trihedral
Polyhedral (ribbed) (e.g. pumpkin)
Winged (have leathery outgrowths)
Stem formation
The stem, like the root, is formed from the primary apical meristem. Unlike the root, the growth cone is protected from the environment by young developing leaves. Like in the root, the meristem cells divide by mitosis and specialize in primary permanent tissues, but unlike the root, the stem does not have distinct growth zones. Specialization of cells occurs very quickly, and as well as in the root, these cells specialize in three of these sites.
From the periphery, 1 layer of cells turns into primary integumentary tissue; in the primary cortex; a central axial cylinder arises in the center. In the stem, unlike the root, the central cylinder many times takes up more space than the primary cortex. 
Primary integumentary tissueepidermal stem, 1 layer, with stomata, with hairs. Its function is to protect the underlying structures.
Primary barkthe stem is responsible for photosynthesis and gives mechanical stability (resists bending). Defines a stalk in space. It consists of:
1) chlorenchyma (from 1 to many layers of these cells)
2) collenchyma (from 1 or more layers). In monocotyledons is rare.
3) endoderm
The endoderm of the stem is 1 layer of living cells, with cellulosic membranes containing starch grains. They are not a storage substance, they help to navigate in space, and therefore the endoderm of the stem is called starchythe vagina.In addition, various plants in the primary cortex can form containers of secretions, crystalline cells can occur, this is the plastic part of the stem.
Central axial cylinderit begins with recycle sclerenchyma. The pericycle in the stem is degenerated into sclerenchyma and partially into the parenchyma. Pericyclic sclerenchyma can be in a transverse section in the form of a continuous ring or in the form of individual cords.
Behind the pericycle are conducting bundles, usually colloral; rarely bicollothral surrounded by cells of the main parenchyma.
The arrangement of conducting bundles is different in monocotyledonous and dicotyledonous plants.
–– in monocotyledons it is located diffusely (distractedly)
–– in dicotyledons in a circle
Such a different arrangement of the conducting bundles is due to the nature of the combination of the conducting bundles of the leaf and stem
In dicotyledons, the conducting bundles of the leaf are connected to the stem in their node. ![]()
In monocotyledonous leaf bundles, they enter the central cylinder to bend and connect with stem bundles passing through 1-2 nodes. 
In dicotyledons, 2-3 leaf bundles enter the stem from a leaf, and in monocotyledons, in connection with parallel venation, a multitude.
In monocotyledonous plants, all conductive bundles closed, i.e. do not have cambium, but in dicotyledons openbeams, i.e. there is a cambium.
The thickness of the stem of monocotyledonous plants along the entire length remains the same, while in dicotyledons the upper part of the stem is slightly less than in the lower part.
The formation of tissue stem secondary structure
The secondary structure of the stem is characteristic of annual and perennial herbaceous, woody dicotyledonous, and gymnosperms. In dicotyledonous plants, the primary structure is very short-lived, and with the onset of cambium activity, a secondary structure is formed. Depending on the tab of the procambium, several types of the secondary structure of the stem are formed. If the procambium cords are separated by wide rows of parenchyma, then the beam structure is formed, if they are brought together so that they merge into a cylinder, a non-beam structure is formed.
Fig. 3.24.The beam type of the structure of the stem of a dicotyledonous plant: A - clover: 1 - epidermis; 2 - chlorenchyma; 3 - sclerenchyma of pericyclic origin; 4 - phloem; 5 - bundle cambium; 6 - xylem; 7 - interbeam cambium
Beam structure of the stemfound in plants such as clover, peas, buttercup, dill (Fig. 3.24). They have prokambial cords laid in one circle around the periphery of the central cylinder. Each prokambial cord turns into a collateral bundle consisting of a primary phloem and primary xylem. Subsequently, cambium is laid between the phloem and xylem from Procambius, which forms the elements of the secondary phloem and secondary xylem. A phloem is deposited to the periphery of the organ, and a xylem to the center, with more xylem being deposited. The primary phloem and xylem remain on the periphery of the beam, and the secondary elements are adjacent to the cambium. The stems of dicotyledonous plants are characterized by the formation of open collateral or bicollateral bundles (Fig. 17, see color incl.).
Also differentiation is characteristic for the stems of dicotyledonous plants primary cortexwhich includes: collenchyma (angular (Fig. 18, see color. incl.) or lamellar), chlorophyll-bearing parenchyma and the inner layer - the endoderm. Starch accumulates in the endoderm; such starchy vaginaplays an important role in the geotropic reaction of the stems. On the border of the primary crust in the central axial cylinder is located pericyclic sclerenchyma- a continuous ring or sections in the form of half arcs above the phloem. The stem core is expressed and represented by parenchyma. Sometimes part of the core is destroyed with the formation of a cavity (see Fig. 3.24).
Non-beam structurecharacteristic of woody plants (linden) (Fig. 19, see color incl.) and many herbs (flax). In the growth cone, the pro-cambial cords merge and form a continuous cylinder, visible on the transverse section in the form of a ring. The ring of the procambium outward forms the ring of the primary phloem, and inward the ring of the primary xylem, between which the cambium ring is laid. Cambium cells divide (parallel to the surface of the organ) and lay the ring of the secondary phloem outward, and inward - the ring of the secondary xylem in a ratio of 1:20. We consider a non-bundle structure using the example of a perennial linden tree stem (Fig. 3.25).
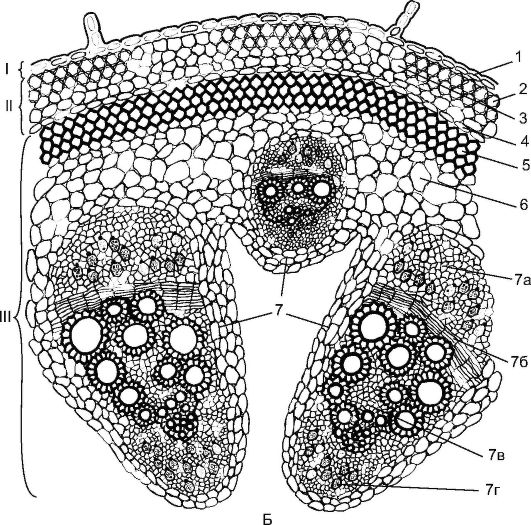
Fig. 3.24.(continued) B - pumpkin: I - integumentary tissue; II - primary bark; III - the central axial cylinder; 1 - epidermis; 2 - angular collenchyma; 3 - chlorenchyma; 4 - endoderm; 5 - sclerenchyma; 6 - the main parenchyma; 7 - bicollateral vascular-fibrous bundle: 7a - phloem; 7b - cambium; 7c - xylem; 7g - internal phloem
A young linden shoot, formed in the spring from a kidney, is covered by an epidermis. All tissues lying before the cambium are called bark. The bark is primary and secondary. Primary barkrepresented by lamellar collenchyma, located immediately below the epidermis by a continuous ring, chlorophyll-bearing parenchyma and single-row starch-bearing vagina. This layer contains grains of “protected” starch, which the plant does not consume. It is believed that this starch is involved in maintaining the plant's balance.
The central axial cylinder in linden begins with pericyclic sclerenchyma above the phloem. As a result of cambium activity, secondary bark(from cambium to periderm), represented by the secondary phloem, core rays and parenchyma of the secondary cortex. Harvest bark from linden, removing to cambium, it is especially easy to do this in spring, when cambium cells actively divide. Previously, linden bark (bast) was used for weaving bast shoes, making baskets, washcloths, etc.
The trapezoid phloem is divided by triangular primary core rays penetrating the wood to the core. The composition of the phloem in linden is heterogeneous. It contains hardwood bast fibers that make up a solid bast, and soft bast is represented by sieve tubes with companion cells and bast parenchyma. The bast loses its ability to conduct organic matter usually after a year and is updated with new layers due to the activity of cambium.
The cambium also forms secondary core rays, but they do not reach the core, being lost in the secondary wood. Core rays are used to move water and organic matter in a radial direction. In the parenchymal cells of the core rays, reserve nutrients (starch, oils) are deposited by autumn, spent in the spring on the growth of young shoots.
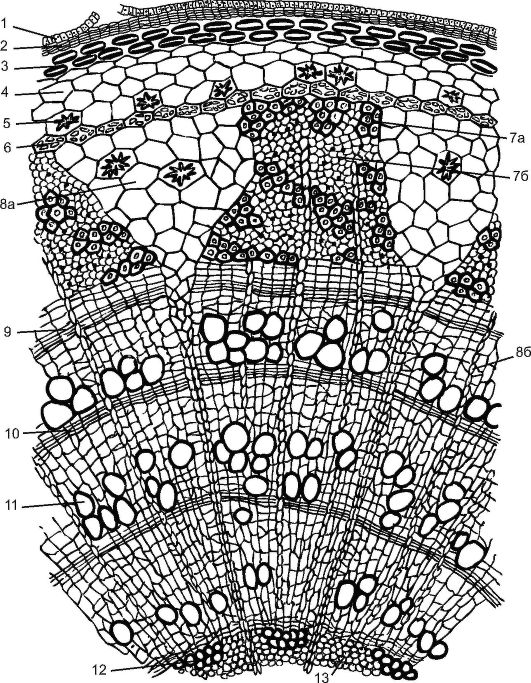
Fig. 3.25.Cross section of a three-year-old branch of linden: 1 - the remains of the epidermis; 2 - cork; 3 - lamellar collenchyma; 4 - chlorenchyma; 5 - Druze; 6 - endoderm; 7 - phloem: 7a - hard bast, (bast fibers); 7b - soft bast - (sieve tubes with companion cells and bast parenchyma); 8a - primary core ray; 8b - secondary core beam; 9 - cambium; 10 - autumn wood; 11 - spring wood; 12 - primary xylem; 13 - core parenchyma
In the summer, a pellogen is laid under the epidermis and a secondary integumentary tissue is formed - the periderm. By the fall, with the formation of the periderm, epidermal cells die off, but their remains remain for 2-3 years. Layering of perennial periderm forms a crust.
The xylem layer formed by cambium in woody plants is much wider than the phloem layer. Wood has been functioning for several years. Dead wood cells do not participate in the conduct of substances, but are able to maintain the colossal severity of the crown of the plant.
The composition of wood is heterogeneous, it includes: tracheids(Fig. 20, see color incl.), trachea, wood parenchymaand libriform.Wood is characterized by the presence of tree rings.In early spring, when active sap flow occurs in the plant, cambium in xylem forms broad-lumen and thin-walled conductive elements - vessels and tracheids, and with the approach of autumn, when these processes freeze and cambium activity weakens, narrow-lumen thick-walled vessels, tracheids and wood fibers arise. Thus, a one-year growth is formed, or a one-year ring (from one spring to another), clearly distinguishable in a cross section. According to the annual rings, you can determine the age of the plant (see Fig. 3.25).
Structural features of the dicotyledon stem:
1) the growth of the stem in thickness (due to the activity of the cambium);
2) a well differentiated primary cortex (collenchyma, chlorophyll-bearing parenchyma, starchy endoderm);
3) bicollateral and collateral bundles of only open type (with cambium);
4) the vascular-fibrous bundles are arranged in a ring or merge (non-beam structure);
5) the presence of a core;
6) woody plants are characterized by the presence in the xylem of tree rings.
Structural features of the rhizomes of dicotyledons.The epidermis may be the integumentary tissue of the dicotyledon rhizomes, and in perennial rhizomes the epidermis is replaced by the epidermis. The primary cortex is represented by storage parenchyma and endoderm with Caspari spots. Moreover, the width of the primary crust approaches the width of the central cylinder. The structure of the central axial cylinder, vascular-fibrous bundles and their location in it have the same features as for aboveground stems.
Depending on the environmental conditions in which one or another type of plant is formed, various structural features of its organs are created, including the stem.
In addition, the stem in some cases performs the functions of other organs. In this case, changes in the structure of the stem are especially significant and are usually classified as anomalies. The peculiar structure of the stem is characterized by vines. Vines are especially richly represented in the tropics, where they are the most diverse and powerfully developed. But even in temperate latitudes enough lianas grow. Hops, clematis, ivy, vine, peas, beans, wisteria, bindweed, etc. are real vines, not as luxurious as in the tropics, but still have features in the structure of the stem that are inherent in all vines.
The most characteristic feature of the structure of the stem of the vines is a rather pronounced dissection of the conducting system and the strong development of parenchymal core rays. Even in vines with a well-defined tree stem (vine), the core rays consist of a parenchyma that differs little from the parenchyma of the cortex and core, and these core rays are very high, stretch at a great distance along the length of the stem, resembling long ribbons set vertically along the stem, dissecting the array of the central cylinder into separate sectors. In typical woody plants, core rays, due to the insignificance of their height on tangential sections of wood, look like short spindle-shaped clusters of cells. An even sharper dismemberment of the central cylinder of the stem is revealed in tropical vines. In some tropical vines, parenchymal cells scattered in various places in the wood of the central cylinder begin to divide and grow intensely, pushing apart individual sections of the vascular system. As a result, the old stems of such vines are distinguished by a very irregular and whimsical structure. In general, the structure of the stem of woody vines resembles the structure of the stem of some herbaceous plants.
A number of plants growing in the desert regions of Africa and America, where it rains very rarely, and then for a short time of the year, special specific features are inherent in the shape and structure of the stem. In such plants, the stems are fleshy, green, take a different shape - a ball, plates or are equipped with wide lamellar ribs. Such stems act as leaves. The leaves are often very small and sometimes look like dry spines. Plants with fleshy stems and reduced leaves are called stem succulents. Among them are various types of prickly pears, cacti, and euphorbia.
The reduction of leaf blades to dry spines leads to a significant reduction in the evaporating surface of the entire plant, consisting entirely of a peculiarly modified stem in its aerial part. The stalk itself turns into an organ adapted to accumulate water reserves. This makes it possible for the plant to store moisture in rainy periods and slowly spend it in a time of rainless weather.
In the structure of the integumentary tissues of most types of succulent stems, there are adaptations to a possible reduction in the loss of water through transpiration by the plant. In this case, the system of conductive tissues should be greatly reduced. Mechanical tissues inside the succulent stems become unnecessary and almost absent.
Cellular juice of tissue cells of succulent stems consists of solutions of various acids, sugars and mucous substances. These substances help to reduce the yield of water by the plant. The tissue of the succulent stems is very homogeneous and consists mainly of a thin-walled parenchyma penetrated by underdeveloped vascular bundles: there are few vessels in each bundle, all of them are narrow-lobed and have a primitive structure. Weak development of vascular bundles is consistent with a slight development of leaves, since the leaf in its development stimulates the formation of vascular-fibrous bundles of the stem. In typical, well-defined succulent stems of secondary xylem, little is formed due to weak cambial activity, the vascular system consists mainly of primary elements that arose in the vascular meristem. Consequently, both physiologically and from the point of view of the history of development, the formation of a weak conducting system in the stems of succulent plants is understandable.
Naturally, among the succulents there are no tree forms; all of them are grassy plants in their structure.
In plants of the so-called sclerenchymal type, also growing in arid places, normal leaves and stems are developed, the conductive system is composed of powerfully developed vascular-fibrous bundles or their fusion products. Vascular elements in such bundles are wide-ranging, numerous and more highly organized than those of succulents. Secondary elements in woody plants of this group are clearly expressed, and in the stems of herbaceous plants, the lignification of the cell membranes of the main parenchyma is sharply expressed. The mechanical system is highly developed, its anatomical elements are thick-walled.
The stems of plants growing in water - the so-called aquatic plants - with a relatively rich variety of forms have one common structural feature: they have a well-developed system of intercellular passages and intercellular spaces. The intercellular spaces in the form of more or less large cavities form a system of air passages that penetrate both the core and the bark of the stem. In this case, the core is especially strongly developed, occupying a significant share of the total diameter of the stem. In aquatic plants, especially among representatives of the monocotyledonous class, the mechanical system sometimes prevails over the conducting system. Groups of vascular anatomical elements collected in bundles are usually of relatively small size, while arrays consisting of mechanical elements concentrated mainly along the periphery of the stem reach greater power in comparison with vascular bundles.
The structure of underground stems that perform the function of reservoirs of reserves — rhizomes and tubers — deviates even more sharply from the normal type. Rhizome is a perennial shoot and usually reaches a much greater thickness than overground shoots, wherever they grow, in water or another medium. In connection with the main purpose of the rhizome - to serve as a storage of spare substances - parenchymalization of its tissues is the most characteristic feature of the structure. The core tissue, consisting of parenchymal rounded cells, thin-walled, with small intercellular spaces between them, serves as reserve tissue in typical rhizomes. By its massiveness, the core in the rhizomes prevails over other tissues. Mechanical and conductive tissues, inferior in power to the core and cortex, are less pronounced in the rhizome than in overground shoots.
The structure of the rhizome as a perennial shoot is composed under the influence of various formative environmental factors. The influence of these factors can be either direct or acting through a correlation in the development of plant organs: for example, the degree of development of leaves, the formation of flower shoots, and the deposition of reserve substances affect the structure of the rhizome. The dependence of the structure of the rhizome on the degree of development of leaves on them is especially noticeable.
During underground growth, when only scaly-like small leaves are on the rhizome, a very weak conducting system is formed, in the xylem part consisting of rare narrow-cavity vessels of a primitive structure. After the aboveground fruit-bearing shoot begins to develop with normal large leaves, the cambium activity intensifies and, as a result, the wood develops, and the number and lumen of the vessels, which have a more perfect structure than those formed before the start of the growth of the aboveground shoot, increase.
A peculiar morphological feature of the rhizome is very short internodes. Apparently, the insignificance of the length of the internodes is in close correlation with the weak differentiation of leaves on the rhizome.
According to its morphology, in some respects, a rhizome is similar to another modification of the underground shoot - a tuber. The main difference between the rhizome and the tuber is the duration of existence: the rhizome is a perennial shoot, but the tuber usually lasts one to two years. Therefore, only those structural features that rhizomes have at the first stage of the formation of their tissues are revealed in the tuber. The predominance of parenchyma in the tuber is even more distinct than in the rhizome.
The degree of parenchymalization of the tuber tissue resembles that of the stem of succulent plants. Just as in the stems of succulent plants, the entire mass of tuber tissue, with the exception of the integumentary (periderm), is a storage tissue penetrated by underdeveloped and primitively constructed bundles of the conducting system. Along with the reduction of conductive tissue in a normal tuber, usually no mechanical tissue is formed. The tissues that make up the tuber are uniformly constructed and adapted to perform only one function - the deposition of stocks.

A typical example of a tuber structure may be potato tuber. The figure shows a fragment of a transverse section of a tuber of one of the varieties of potatoes (Voltman), from its central part. For convenience, the demonstration of the tissue structure of starch, abundantly filling all cells, is not shown. The tuber parenchyma is cut only in one place by a narrow strip of smaller cells, consisting of four groups. Each group contains 1-3 narrow-lumen vessels, the rest of the tissue consists of meristem-like cells, among which there are several sieve tubes. Vessels with a poorly developed structure are primitive, some of which are in a state of obliteration and destruction.
Such a tissue, consisting of primitive and underdeveloped vascular elements, is located in a ring throughout the tuber, representing a central cylinder. Outside there is the primary crust, inside - the core. In tubers having only scaly similarities of leaves, secondary tissues are not formed as a result of cambium activity.
If you find an error, please select a piece of text and press Ctrl + Enter.