Connective tissue also called the tissue of the internal environment. It is part of every organ and forms layers between the organs, as if connecting them. Connective tissue covers blood vessels and nerves, participates in the formation of the human skeleton and the skeleton of its individual organs, and in the formation of blood and lymph.
Connective tissue performs the following functions: trophic, protective, supporting (mechanical) and plastic.
Trophic, or nutritious, function is that blood, which belongs to connective tissue, carries nutrients throughout the body. In addition, by dressing the vessels, the connective tissue along with them penetrates into all tissues and organs.
Protective function connective tissue is associated not only with its mechanical properties (bones - dense formations - protect organs), but also with the fact that its cells have the ability to phagocytosis: they absorb and digest harmful substances. Connective tissue is also involved in the formation of protective bodies that create immunity (immunity to diseases).
Support function Connective tissue is determined mainly by the intercellular substance.
Plastic function connective tissue is expressed in its high ability to regenerate and adapt to environmental conditions. This tissue is formed from the middle germ layer of the mesoderm, from the so-called germinal connective tissue (mesenchyme).
Connective tissue consists of cells and intercellular substance, in which the ground substance and fibers are secreted. Unlike other types of tissue, intercellular substance predominates in it, while there are few cells. In different types of connective tissue, the quantitative ratio of intercellular substance and cells is different.
The ground substance of connective tissue contains many fibers. Some of them, located in the form of thick straight or somewhat convoluted ribbons, do not branch, consist of a special adhesive substance and are called collagen or adhesive fibers. They are poorly stretchable, very durable. Another type of fiber is Elastic. They are thinner and branching. These fibers are less strong than collagen, but have greater elasticity and elasticity (like rubber, they can stretch and then return to their original shape).
The main cells of the tissue are fibroblasts, fibrocytes, macrophages, mast cells and plasma cells. It may contain fat cells, pigment cells and even white blood cells.
Fibroblasts- the main type of connective tissue cells. They have an irregular or spindle-shaped (elongated) shape. Their core is quite large and oval in shape. Fibroblasts participate in the formation of intercellular substance and fibers, in wound healing, and the development of scar tissue. Fibroblasts that have completed their life cycle, are called fibrocytes.
Macrophages can be various shapes: round, elongated, irregular. Their shell is folded, with a large number of microvilli, with the help of which they capture foreign substances. These cells usually have one nucleus, small in size, oval or bean-shaped. Macrophages are the main defenders of the human body. They destroy microbes and neutralize toxic (poisonous) substances.
Mast cells They have an irregular shape, short wide processes, and a small core. There are many grains in the cytoplasm. Mast cells have a well-developed ability for amoeboid movements. They participate in the formation of intercellular substance and regulation of its composition, produce substances that prevent blood clotting and the deposition of salts in the walls of blood vessels.
Plasma cells oval or round in shape participate in the formation of protective bodies, especially react when a foreign protein is introduced into the body.
Fat cells contain fat in the cytoplasm, pushing the nucleus to the periphery. Their number in loose connective tissue is variable. With increased nutrition, the number of fat cells increases sharply.
Pigment cells- these are the same fibroblasts or fibrocytes, in the cytoplasm of which there is a lot of coloring matter - pigment.
According to the functions performed, which are determined to a large extent by the physicochemical characteristics of the intercellular substance (it can be liquid, dense and very hard), connective tissue is divided into protective-trophic and supporting. Protective-trophic connective tissue includes: blood, lymph, reticular, or reticular, tissue, loose fibrous tissue and endothelium. Supporting connective tissue includes: dense fibrous tissue, cartilage and bone tissue. As the intercellular substance becomes denser, the trophic function of the tissue decreases and the supporting function increases.
Blood- This is a type of connective tissue with a liquid intercellular substance and specific cells. The intercellular substance of blood is its liquid part - plasma, which contains the formed elements (cells) of blood. By volume, plasma makes up 55-60%, and formed elements 40-45% of all blood. In the adult body there are 4.5-5 liters of blood.
Plasma. Blood plasma consists of inorganic and organic substances. Inorganic substances it contains about 91% (90% is water and 1% minerals), and organic about 9%. The main part of organic matter is proteins - 7%. There are 3 types of them - fibrinogen, albumin and globulins. Fibrinogen takes part in blood clotting, albumins transport poorly water-soluble substances (including drugs), and globulins ensure the formation of protective bodies. The amount of globulins increases sharply during infectious diseases. Blood plasma devoid of fibrinogen is called blood serum. It is used for therapeutic or prophylactic purposes to create immunity (passive immunization), and therapeutic serums are prepared. Blood plasma also contains organic substances of a non-protein nature (urea, fat, amino acids, etc.), although in very small quantities.
Formed elements of blood. There are three types of them: red blood cells - erythrocytes, white blood cells - leukocytes and platelets - platelets.
Red blood cells(erythros - red, cytos - cell) are specific highly differentiated cells that, during development, have lost their nucleus, mitochondria, reticular apparatus and cell center (Fig. 6). In frogs, fish, and birds, red blood cells contain nuclei (Fig. 7). In the cytoplasm of erythrocytes there is complex protein- hemoglobin, with the help of which gas exchange is carried out in the body: oxygen is transferred from the lungs to the tissues, and from the tissues to the lungs - carbon dioxide. The membrane of red blood cells is very thin, and gases are exchanged through it. The red blood cell has the shape of a biconcave disk, which increases its surface, facilitating better contact of hemoglobin with the transported gases. The absence of a nucleus in the cell also appears to allow for greater oxygen uptake.
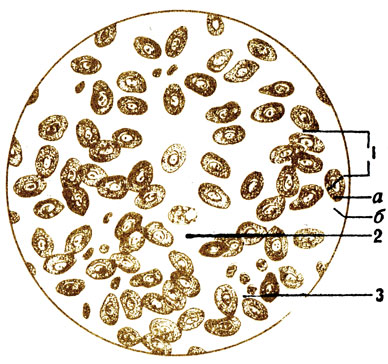
The size of red blood cells is small, only 7-8 microns, so they pass quite easily through the thinnest blood vessels - capillaries. 1 mm 3 of blood contains 4.5-5.0 million, and in total there are 25-28 trillion red blood cells. If it were possible to place them side by side, it would create a chain that would be enough to encircle the globe 3 times along the equator. Total surface red blood cells circulating in the blood, more than 1/4 hectare. In men, the number of red blood cells is slightly higher than in women; in children - more than in adults; Residents of high mountain areas, where there is less oxygen in the air, have more than residents of the plains. Even with a short-term (1-2 months) stay in mountainous areas, the number of red blood cells increases, which is important for conducting sports training there. With increased muscle activity, they also become larger due to the increased demand for oxygen. Red blood cells do not have the ability to independent movements, they move through the blood vessels with the blood flow. However, they are very elastic; when they move in the capillaries, you can clearly see how they lengthen, flatten, changing their shape. The lifespan of red blood cells is 80-120 days. Red blood cells break down in the spleen and are formed in the red bone marrow. According to available data, 1/100 of the red blood cells are destroyed every day, i.e., in a little more than 3 months, all red blood cells are renewed.
Leukocytes- these are cells with a nucleus. They are larger than red blood cells (up to 10 microns), capable of independent amoeboid movement, and can exit the capillaries into the underlying tissue.
Depending on the nature of the cytoplasm, the presence of inclusions in it in the form of grains of protein, pigment, and also depending on the shape of the nucleus, leukocytes are divided into granular and non-granular. The former have granularity in the cytoplasm and a segmented nucleus, divided into separate parts. Depending on the type of granularity and its relationship to dyes, neutrophils, basophils and eosinophils are distinguished, which are found in certain quantitative ratios in the blood. The change in this ratio in various diseases determines not only their nature, but also their outcome.
Non-granular forms of leukocytes do not contain inclusions in the cytoplasm; their nucleus is not divided into parts, has a round shape and is most often located in the center of the cell. Non-granular leukocytes include lymphocytes and monocytes.
1 mm 3 of blood contains 6 - 8 thousand leukocytes. Their number may increase after eating, during infectious diseases, and especially after intense muscle activity. The lifespan of leukocytes varies: from several days to 2 - 3 months. When an infection enters the body, they die in significant numbers in the fight against it. Granular forms of leukocytes are produced in the red bone marrow, and lymphocytes are produced in the spleen and lymph nodes.
The main function of leukocytes is protective. They guard the health of the body, helping it fight various diseases. They perform a protective function by participating in phagocytosis and in the formation of protective bodies. In addition, leukocytes produce enzymes that regulate blood clotting and vascular permeability. Finally, separate forms lymphocytes can form cells various types connective tissue (fibroblasts, macrophages, smooth muscle cells), which is important in recovery processes.
Platelets, or blood platelets, are round or oval-shaped bodies measuring only 1 - 2 microns in size. They do not contain a kernel. There are 200 - 300 thousand of them in 1 mm 3 of blood. The lifespan of platelets is 5 - 8 days. Blood platelets take part in blood clotting.
Lymph, like blood, consists of a liquid part - lymphoplasm - and formed elements. Unlike blood plasma, it contains less protein, but more metabolic products. Of the formed elements, lymphocytes predominate in it, erythrocytes are absent.
Reticular tissue consists of irregularly shaped cells. In contact with each other, they form a kind of network. In the loops of this network there is an intercellular substance containing a large number of reticulin fibers entwining the surface of the cells. Hematopoietic organs (bone marrow, spleen, lymph nodes) are built from reticular tissue.
Loose fibrous connective tissue- this is the tissue in which all the structural elements of connective tissue are most clearly expressed: intercellular substance, fibers and cells (Fig. 8). It covers blood vessels and nerves, forms subcutaneous tissue, and participates in the structure of almost all organs.
Endothelial tissue(endothelium) is related to connective tissue only in origin, while in structure it resembles epithelial tissue. Its cells are flat and located on the basement membrane. There is little intercellular substance in this tissue. Endothelium lines the inner surface of blood vessels, giving it a smooth, shiny appearance; Metabolism occurs through the endothelial cells of the capillaries; They also perform a protective function.
Dense fibrous connective tissue has characteristic feature- it is dominated by collagen fibers, which are collected in bundles oriented depending on the direction of traction forces. There are few cells here (mainly fibroblasts), and they are located between the fiber bundles. Ligaments, tendons, fascia, intermuscular septa, periosteum, perichondrium, etc. are built from this tissue (Fig. 9).

In tendons and ligaments, bundles of collagen fibers are located in parallel, in fascia, aponeuroses, intermuscular septa - in layers above each other (the thicker the fascia, the more layers), and the direction of the fibers in different layers is different: in some - at a right angle, in others - under sharp, which gives these formations special strength. If the dense fibrous connective tissue is dominated by elastic fibers, it is called elastic connective tissue. The presence of elastic fibers helps an organ or part of the body return to its original position after changing shape.
Cartilage tissue(cartilage) by physical and chemical properties and functional features differs sharply from other types of connective tissue. Intercellular substance It is quite dense, and therefore it mainly performs supporting and protective (mechanical) functions. There are three types of cartilage: hyaline, or vitreous, collagen-fibrous and elastic. Cartilage tissue does not have blood vessels. Metabolism (nutrition and removal of waste products) is carried out through the vessels of the connective tissue membrane covering the outside of the cartilage (perichondrium). Nutrients from the vessels of the perichondrium penetrate into the intercellular substance of the cartilage. The cartilage that covers the articular surfaces of bones receives nutrients from the synovial fluid that fills the joint cavity or from nearby bone vessels. The growth of cartilage occurs due to the perichondrium.
Hyaline cartilage has the greatest distribution in the human body. Its intercellular substance is translucent, bluish-white. Cartilage cells are located in special cavities surrounded by a capsule, which is denser than the intercellular substance. Hyaline cartilage forms the anterior ends of the ribs, the cartilage of the trachea, bronchi, most of the cartilage of the larynx and covers the articular surfaces of bones. In the embryonic period, a significant part of the skeleton consists of hyaline cartilage. In old age, lime may be deposited in hyaline cartilage (Fig. 10).

Collagen fibrocartilage less elastic, but more durable. Its intercellular substance contains a large number of bundles of collagen fibers located more or less parallel. The cells are located between the fiber bundles. The intervertebral discs, the cartilage connecting the pubic bones, are built from this cartilage (Fig. 11).
Elastic cartilage less durable, but very elastic, calcification never occurs in it. In the intercellular substance of cartilage there are many elastic fibers that intertwine with each other, forming a dense network. Its cells resemble the shape of a candle flame and are located 2-3 in capsules between the fibers. Elastic cartilage is located where great resistance to acting forces is not required. The auricle, epiglottis, wall of the external auditory canal and auditory tube are built from it (Fig. 12).

Bone is the densest of all types of connective tissue. Its intercellular substance consists of fibers, which are often connected into bundles, and the ground substance, in which a large percentage inorganic compounds, predominantly calcium salts, so the supporting function of the bone is most pronounced. However, despite the density bone- a living system, it undergoes changes throughout a person’s life, accompanied by the renewal of its constituent elements, which ensures its adaptability to conditions environment(Fig. 13).

Restructuring of bone tissue depends on age, nutrition, function of the internal secretion organs and other factors. The most pronounced changes in bone tissue occur during muscle activity: not only the internal structure of bone tissue changes, but also the shape of the organs - the bones that it forms.
There are three types of cellular elements in bone tissue: osteocytes, osteoblasts and osteoclasts.
Osteocyte(oss - bone, cytos - cell) - the main cell of bone tissue - has an irregular shape, a large number of long processes with which it contacts neighboring cells. These bone cells lie in special cavities.
Osteoblasts- creators, creators of bone tissue. They are located where the process of bone formation occurs. Their shape can be cubic, pyramidal or angular. As bone tissue forms, osteoblasts transform into osteocytes.
Osteoclasts- multinucleate cells. They are larger than osteocytes and osteoblasts. Each osteoclast can have up to 50 nuclei. A small depression is formed at the point of contact of the osteoclast with the bone substance. Osteoclasts lie in such depressions, coves. These cells destroy bone tissue, in its place new one is formed. Both processes continuously occur in bone tissue - both the process of destruction and the process of creation, ensuring the reconstruction of the bone.
There are two types of bone tissue: coarse-fiber and fine-fiber, or lamellar.
Rough fibrous bone tissue It is found to a greater extent in the fetus; in adults it is found only in the places where muscle tendons attach to the bones, in the sutures between the bones of the skull. In the intercellular substance of coarse-fibrous bone tissue, the bundles of fibers are thick, located parallel, at an angle, or in the form of a network. Osteocytes have a flattened shape.
Fine fiber, or lamellar, bone the most highly differentiated. Its structural and functional unit is the bone plate. In the intercellular substance of the plate, the fibers are thin and oriented in certain directions parallel to each other. Osteocytes lie between the laminae or within the laminae. The plates are arranged so that the fibers in two adjacent plates run almost at right angles, which ensures special strength and elasticity of bone tissue. Almost all the bones of the adult human skeleton are built from fine-fiber bone tissue.
We have already written about the basic terms and general components of CT in a previous article on the characteristics of connective tissue. Now let’s characterize some connective tissue groups(ST).
Loose ST- this is the main and main tissue when it comes to connective tissue (Fig. 10). Elastic (1), collagen (2) fibers, as well as some cells are included in its amorphous component. The most basic cell is the fibroblast (Latin fibra - fiber, Greek blastos - sprout or germ). Fibroblast is capable of synthesizing the constituent elements of the amorphous component and forming fibers. That is, the actual function of a fibroblast cell is the ability to synthesize intercellular substance. Fibroblasts (3) with a large nucleus (a) in their endoplasm (b) and ectoplasm (c) contain a rather impressive endoplasmic reticulum, in which proteins such as collagen and elastin are synthesized. These proteins are the builders of the corresponding fibers. Another important cell of the loose CT is the histiocyte (4). Microorganisms should be wary of these cells, because when they get into the intercellular substance, they phagocytose them or, simply put, eat them. Finally, in color picture I you can see another important cell of the loose CT - this is a mast cell; it stores two biologically active compounds: heparin and histamine. Heparin is a substance that prevents blood clotting. Histamine is a substance that takes part in various allergic reactions and inflammatory processes. Due to the release of histamine from mast cells, symptoms such as redness of the skin, hives, itching, blistering, burning and anaphylactic shock are observed. 
Picture I. Loose connective tissue
Loose CT accompanies all vessels. The aorta is covered with a whole cushion - adventitia, and the smallest capillaries are surrounded by a very thin web of fibers and cells. The vessels are protected, strengthened and, as it were, based on this type of ST. This means that loose CT is located wherever vessels are present. It is for this reason that it is worth highlighting as the main and main connective tissue.
A practical doctor in his daily work very often encounters one manifestation of loose connective tissue - edema. Glycosaminoglycans, which form an amorphous component, are able to retain water, which they do whenever possible. And this opportunity appears in certain pathological processes: heart failure, lymph stagnation, kidney disease, inflammation, and so on. In this case, fluid accumulates in the connective tissue, which swells, making the skin swollen. Sometimes swelling under the eyes can be the initial symptom of a disease such as glomerulonephritis - an immune inflammation of the kidney. ![]()
Dense ST contains a very small number of cellular components and an amorphous component of the intercellular substance; most of the dense connective tissue is made up of fibers. There are two forms of dense CT. Dense unformed ST(Fig. 11) has a complete disorder of fibers (4). Its fibers intertwine as they please; fibroblasts (5) can be oriented in any direction. This type of ST is involved in the formation of the skin; it is located under the epidermis (1) and the layer of loose ST (2) surrounding the vessels (3), and gives the dermis a certain strength. But in this she cannot compare with strength densely decorated ST(Fig. 12), which consists of strictly ordered bundles (5), which in turn have a certain direction of collagen (3) and/or elastic (4) fibers. Formed connective tissue is part of tendons, ligaments, the tunica albuginea of the eyeball, fascia, dura mater, aponeuroses and some other anatomical formations. The fibers are wrapped (1) and “layered” (7) with loose CT containing vessels (2) and other elements (6). Thanks to the parallelism of the tendon fibers, they get their high strength and rigidity. 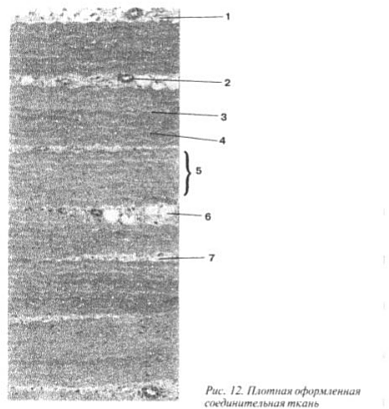
Adipose tissue(Fig. 13) is distributed almost everywhere in the skin, retroperitoneum, omentum, and mesentery. The cells of adipose tissue are called lipocytes (1 and picture II). They are very densely located, allowing only small vessels such as capillaries (2) to pass between them, and with them the ubiquitous fibroblasts with individual fibers (3). Lipocytes are almost completely devoid of cytoplasm and filled with large solid droplets of fat. The nucleus turns out to be displaced to the side, despite the fact that it is a regulator of the cell’s functioning. 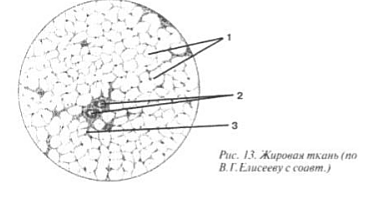

Picture II. Adipose tissue
Adipose tissue is the body's most important source of energy. After all, when fat is broken down, it is released much more than when using carbohydrates and proteins. In addition, a significant amount of water is formed in this case, so adipose tissue simultaneously turns out to be a reserve reservoir of bound water (it is not for nothing that this particular variant of ST is found in the humps of camels, which slowly break down fat when crossing hot deserts). There is one more function. In newborn children, a special subtype was found in the skin - brown adipose tissue. It contains a huge number of mitochondria and due to this it is the most important source of heat for the baby that is born. 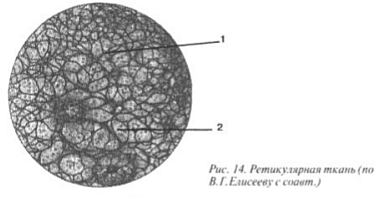
Reticular tissue, located in the organs of the lymphatic system: in the red bone marrow, lymph nodes, thymus (thymus gland), spleen, consists of multi-processed cells called reticulocytes. The Latin word reticulum means “network,” which suits this fabric perfectly (Fig. 14). Reticulocytes, like fibroblasts, synthesize fibers (1), called reticular (a variant of collagen). This type of CT ensures hematopoiesis, that is, almost all blood cells (2) undergo development, in a kind of hammock consisting of reticular tissue(picture III). 
Picture III. Reticular tissue
The last subspecies of ST itself is pigment tissue(Fig. 15) is found in almost everything that is intensely colored. Examples include hair, the retina of the eyeball, and tanned skin. Pigment fabric represented by melanocytes, cells filled with granules of the main animal pigment - melanin (1). They have a star-shaped shape: from the nucleus located in the center, the cytoplasm diverges into petals (2). 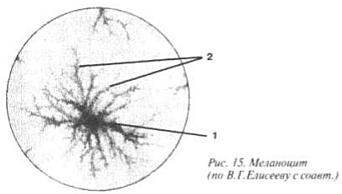
These cells can give rise to a malignant tumor - melanoma. The disease has recently become much more common than before. In the last decade, the incidence of skin cancer has increased very sharply, it is believed that this is due to changes in the thickness of the ozone layer, which protects our planet with a thick layer from the deadly effects of ultraviolet radiation. Over the poles it decreased by 40-60%; scientists even talk about “ ozone holes" As a result, in people roasting under the sun, the melanocytes of birthmarks are the first to respond to the mutagenic effect of ultraviolet rays. Dividing continuously, they give rise to tumor growth. Unfortunately, melanoma progresses rapidly and usually metastasizes early.
Cartilage tissue(Fig. 16) - tissue that has in its intercellular substance a very “high-quality”, concentrated amorphous component. Glycosamine and proteoglycans make it dense and elastic, like jelly. This time, both the amorphous and fibrous components of the intercellular substance are synthesized not by fibroblasts, but by young cells of cartilage tissue, which are called chondroblasts (2). Cartilage has no blood vessels. Its nutrition comes from the capillaries of the most superficial layer - the perichondrium (1), where the chondroblasts are actually located. Only when they “mature” do they become covered with a special capsule (5) and pass into the amorphous substance of the cartilage itself (3), after which they are called chondrocytes (4). Moreover, the intercellular substance is so dense that when a chondrocyte divides (6), its daughter cells cannot separate, remaining together in small cavities (7). 
Cartilaginous tissue forms three variants of cartilage. The first, hyaline cartilage, has very few fibers, and it is found at the junction of the ribs with the sternum, in the trachea, in the bronchi and larynx, on the articular surfaces of bones. The second type of cartilage is elastic (picture IV), containing many elastic fibers, it is located in the auricle and larynx. Fibrous cartilage, which contains mainly collagen fibers, forms the pubic symphysis and intervertebral discs. 
Picture IV. Elastic cartilage
Bone contains three types of cells. Young osteoblasts are similar in function to fibroblasts and chondroblasts. They form the intercellular substance of bone, located in the most superficial layer rich in blood vessels - the periosteum. As they age, osteoblasts are incorporated into the bone itself, becoming osteocytes. During the embryonic period, the human body does not have bones as such. The embryo has cartilaginous “blanks”, models of future bones. But gradually ossification begins, requiring the destruction of cartilage and the formation of real bone tissue. The destroyers here are cells - osteoclasts. They crush the cartilage, making room for osteoblasts and their work. By the way, aging bone is constantly replaced by new ones, and again it is osteoclasts that are responsible for destroying used bone.
The intercellular substance of bone tissue contains a small amount of organic substances (30%), in particular collagen fibers, which are strictly oriented in the compact substance of the bone (picture V) and random in the spongy bone. The amorphous component, having “realized” that it is “superfluous at this celebration of life,” is practically absent. Instead, there are various inorganic salts, citrates, hydroxyapatite crystals, and more than 30 microelements. If you heat a bone in a fire, all the collagen will burn away; the shape will be preserved, but just touch it with your finger and the bone will crumble. And after a night in a solution of some acid, in which all inorganic salts dissolve, the bone can be cut like butter with a knife, that is, it will lose strength, but on the neck (thanks to the remaining fibers) it will be tied like a pioneer tie. 
Picture V. Bone tissue
Last but not least connective tissue group, is blood. Studying it requires a huge amount of information. Therefore, we will not diminish the significance of blood by describing it here, but will leave this topic for separate consideration.
Connective tissues are widespread tissues of mesenchymal origin. They ensure the maintenance of the integrity of other tissues, form the stroma of organs, contain blood and lymphatic vessels, and participate in the trophic provision of all tissues and organs. Among the connective tissues there are fibrous fabrics(loose and dense) and tissues with special properties (for example, adipose, reticular). Different connective tissues differ in physical properties. In most organs and between them, loose connective tissue acts as a package of parenchyma, organizing a bed for its histological elements (organ stroma). For many cell types, organs and tissues, the connective tissue environment plays not only a passively supportive role, but also has organizing and informational significance. Dense connective tissues provide strength to the skin, form organ capsules, and allow them to withstand significant mechanical loads (for example, ligaments and tendons). Connective tissue is metabolically important not only because it contains blood vessels. Thus, white adipose tissue stores fat as a source of energy, and brown adipose tissue is involved in the regulation of body temperature in newborns. A number of connective tissue cells
belongs to the function of protection against pathogenic microorganisms. Connective tissue is a springboard for inflammatory and immune reactions.
EXTRACELLULAR MATRIX
All fibrous connective tissues contain a significant amount of intercellular substance. Chemical composition, the structure and volume of the extracellular matrix determine physical properties each type of connective tissue. The extracellular matrix consists of a ground substance and various types of fibers embedded in it.
Main substance
The main substance is an amorphous material with gel properties. Tissue fluid binds with the components of the main substance, forming a medium for the passage of molecules through the connective tissue and for the exchange of substances with the blood. The main substance contains glycosaminoglycans, proteoglycans and glycoproteins. The molecules of the main substance are tightly bound to the fibers of the extracellular matrix and interact with various cells.
Glycosaminoglycans
Glycosaminoglycans are polysaccharides built from repeating disaccharide units, one of which is usually uronic acid, and the other is an amino sugar (N-acetylglucosamine, N-acetylgalactosamine). Glycosaminoglycans are acidic (negatively charged) compounds, because their disaccharide units contain many hydroxyl, carboxyl and sulfate groups. Glycosaminoglycan molecules are hydrophilic and bind a large number of water molecules and ions (in particular, Na +) entering the extracellular fluid. Large glycosaminoglycan molecules bound together form a gel through which metabolites diffuse. The following types of glycosaminoglycans are known: hyaluronic acid, chondroitin sulfate, dermatan sulfate, keratan sulfate, heparan sulfate and heparin (heparin sulfate).
Proteoglycans
Proteoglycan is a fibrous central protein with glycosaminoglycans covalently attached to it. This is how large macromolecules are formed, containing 90-95% carbohydrates. Proteoglycans can bind non-covalently to hyaluronic acid chains, forming even larger complexes (for example, cartilage proteoglycans).
Glycoproteins
Glycoproteins consist of polypeptide chains connected to branched polysaccharides and connect cells to the extracellular matrix. There are glycoproteins that form fibrous structures (fibronectin and fibrillin), as well as a number of non-fibrous proteins (laminin, tenascin and entactin).
Fibronectin found in the basement membrane, around collagen fibers and cells. A large amount of fibronectin is found in blood plasma. Fibronectin is involved in cell adhesion by linking them to components of the extracellular matrix, in particular collagen and glycosaminoglycans. During embryogenesis and wound healing, fibronectin forms pathways for cell migration. The fibronectin receptor is an integrin built into cell membrane. Inside the cell, integrin interacts with actin microfilaments of the perimembrane cytoskeleton, and outside it connects with fibronectin. In turn, fibronectin forms bonds with collagen and glycosaminoglycans. This establishes structural continuity between the cytoskeleton and the extracellular matrix (Fig. 6-18 ).
Fibrillin(M r ≈350 kD) forms microfibrils with a diameter of 8–12 μm, strengthening the connections between extracellular components; is part of elastic fibers, ensuring their ordered structure. Mutations of the fibrillin gene lead to the development of the syndrome Marfana.
Different types of fibers are immersed in the extracellular matrix: collagen and reticulin fibers built from collagens and elastic fibers (they include elastin and fibrillin).
Collagen fibers
Collagen fibers are the main component of most connective tissues, and collagen is the most abundant protein.
Collagens. The collagen molecule is a helix of three pro-α chains. The length of such a spiral is 300 nm, diameter is 1.5 nm. All types of collagen contain regions with a repeating sequence of three amino acids with glycine in the third position. The first amino acid in this sequence can be any, the second can be proline, hydroxyproline or lysine. Due to the abundance of cross-links between lysine residues, collagen fibers have high strength. At least 13 types of collagen are known (Table. 6-2 ).
Rice. 6-18. Fibronectin-mediated cell communication with collagen fibers. The cytoplasmic domain of integrin interacts with near-membrane microfilaments (actin), and the extracellular domain interacts with fibronectin, which in turn is associated with collagen fibers. The fibronectin molecule also contains domains for binding to heparin and fibrin.
The most common collagens are the first five types, the rest are relatively rare and in small quantities. Types I, II and III collagens form fibrils with a periodicity of 65 nm. Type I collagen is one of the most common, synthesized by fibroblasts, osteoblasts, and SMCs. Type II collagen is synthesized by chondroblasts, chondrocytes, and retinal neurons and is present in the vitreous body. Type III collagen is part of reticulin fibers, which form a delicate supporting network in some organs and tissues (for example, in the liver, bone marrow, lymphoid organs), and is synthesized by fibroblasts and SMCs. Type IV collagen forms the reticulate basis of basement membranes and is synthesized by most epithelial cells. Type V collagen is present in blood vessels, around SMCs and in the bone matrix, and is synthesized by SMCs and osteoblasts. Mutations of collagen genes lead to the development of various diseases (osteogenesis imperfecta, achondrogenesis and achondroplasia, syndromes Ehlers–Danlos).
Formation of collagen fibers(rice. 6-19 ). At the intracellular stage, synthesis and assembly of pro-α chains occurs in the granular endoplasmic reticulum. Further in the tanks of the complex Golgi Hydroxylation and glycosylation of polypeptides occurs, resulting in the formation of helices of three pro-α chains (procollagen). Each pro-α chain contains terminal sections - registration peptides. These peptides prevent the intracellular association of procollagen helices into collagen fibrils. Procollagen molecules accumulate in secretory granules and are released into the extracellular space. Outside the cell, the registration peptides of the procollagen molecule are cleaved off using procollagen peptidase (the resulting molecule is sometimes called tropocollagen). This shedding does not occur in collagen types IV and V, which explains the absence of fibrils built from collagens of these types. Next, the assembly of collagen fibrils occurs (Fig. 6-20 ). Tropocollagen molecules have tail and head ends and spontaneously combine into parallel chains, lining up sequentially: “end to end” and “side to side.” In this case, gaps 35 nm wide remain between the head and tail ends.
Table 6-2. Distribution of collagen in tissues and organs
| Type | Tissues and organs |
| I | Skin, tendons, bones, cornea, placenta, arteries, liver, dentin, tumors |
| II | Cartilage, intervertebral discs, vitreous body, cornea |
| III | Lungs, arteries, uterus, reticulin fibers in the liver and hematopoietic organs |
| IV | Basement membranes |
| V | Placenta, skin, blood vessels, smooth muscle tissue, rhabdomyosarcoma |
| VI | Blood vessels, ligaments, skin, uterus, lungs, kidneys |
| VII | Amnion, skin, esophagus, cornea |
| IX | |
| X | Cartilage |
| XI | Cartilage, intervertebral discs, vitreous |
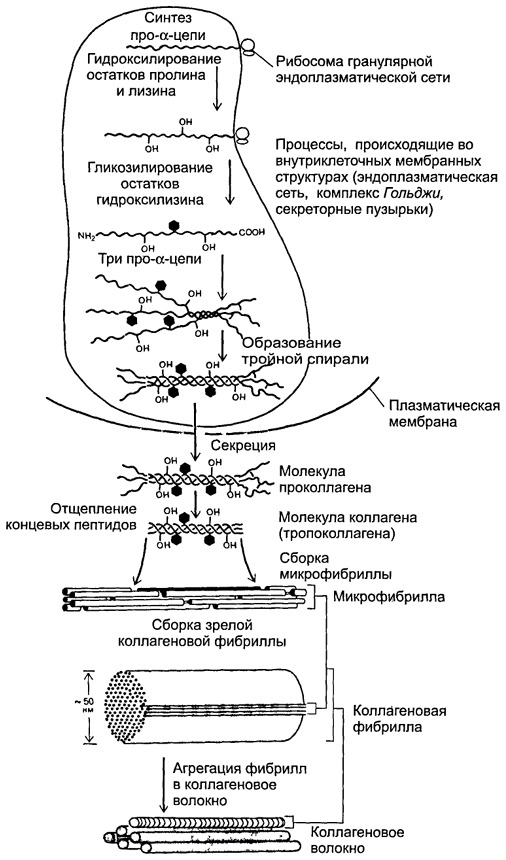
Rice. 6-19. Formation of collagen fiber. The synthesis of polypeptide α-chains, their hydroxylation and glycosylation, as well as the formation of a helix of three α-chains occur in the granular endoplasmic reticulum and complex Golgi. Procollagen molecules accumulate in secretory vesicles and are released into the extracellular space. Outside the cell, the terminal peptides are split off from the polypeptides and collagen fibrils are formed.
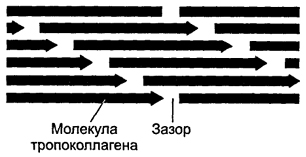
Rice. 6-20. Packing of polarized tropocollagen molecules during collagen fiber formation. When molecules are sequentially combined into a chain, a gap of 35 nm wide is maintained between the tail and head ends of neighboring molecules. Each molecule of one chain is displaced relative to the molecule of the neighboring chain by a quarter of its length. This is how a collagen fibril is formed, which has a transverse striation with a periodicity of 65 nm.
Each tropocollagen molecule in the chain is displaced relative to the molecule in the neighboring chain by a quarter of its length, which is the reason for the alternation of dark and light stripes with a periodicity of 65 nm. Thicker fibrils are formed by the attachment of new tropocollagen molecules. The copper-containing enzyme lysyl oxidase is involved in the formation of cross-links between tropocollagen molecules. The interaction of collagen with glycosaminoglycans and fibronectin regulates fibril assembly. Collagen fiber is formed from many collagen fibrils connected in parallel by glycoproteins. Collagen fiber bundles are a collection of collagen fibers immersed in the intercellular matrix.
- Vitamin C– one of the cofactors necessary for the formation of cross-links between polypeptide chains. Vitamin C deficiency leads to the formation of weakly hydroxylated pro-α chains, which are unable to form triple helices and collagen fibrils. As a result, defects in bones, teeth, and vascular walls occur.
Interaction of collagen with cells(Table 6-3 ). Collagen fibers surround many cells. Some cells (for example, hepatocytes and SMCs) have collagen receptors and directly bind to it. Other cells bind to collagen through special binding intermediary proteins (usually fibronectin, less often chondronectin and laminin).
Elastic fibers
Elastic structures(fibers and membranes) have pronounced elastic properties. Elastic fibers are present in elastic cartilage, skin, lungs, and blood vessels. Fenestrated elastic membranes are characteristic of large arteries.
Table 6-3. Linking molecules between collagen and various cell types
Elastic fiber(branching thread with a diameter of 0.2–1.0 μm) consists of fibrillin and amorphous elastin (Fig. 6-21 ). The glycoprotein elastin, like collagen, contains a lot of glycine and proline. Fibroblasts and SMCs synthesize elastin and fibrillin in the cisterns of the granular endoplasmic reticulum. In complex Golgi polypeptides are packaged into
 |
| Rice. 6-21. Elastic fiber. A. Thanks to the establishment of intermolecular bonds between elastin polypeptides, an elastic network of molecules is formed that restores its shape after deformation. B. Fibrillin microfibrils, organizing elastin, are located both outside and inside the elastic fiber. |
Reticulin fibers
Reticulin fibers are thin filaments with a diameter of 0.5–2.0 μm, consisting of type III collagen associated with glycoproteins and proteoglycans. With defects in the gene encoding the α1 chain of collagen type III, the formation of reticulin fibers is disrupted, which leads to the appearance of breaks in the intestinal wall and/or large arteries, where reticulin fibers form a sheath around the SMC (syndrome Ehlers–Danlos type IV).
basement membrane
The basement membrane is a special type of extracellular matrix; it looks like a sheet or plate 20–200 nm thick (see Fig. 16-2 ) and consists of special proteins that serve to connect cells of various types
(epithelial, muscle, Schwannian) with surrounding connective tissue. The basement membrane consists of two layers - lamina lucida(light layer) 10–50 nm thick, adjacent to the plasmalemma, and lamina densa(electron-dense layer varying in thickness). The basis lamina lucida And lamina densa – a three-dimensional network consisting of type IV collagen fibers, to which laminin, entactin and heparan sulfate are associated.
Anchor fibers
Most basement membranes are firmly attached to loose connective tissue. The basement membranes of the stratified epithelium of the skin, esophagus, cornea and amnion are connected by short anchor fibers to anchor plates in the underlying connective tissue. The anchor fibers consist mainly of type VII collagen, the anchor plates contain type IV collagen. Anchor fibers also connect anchor plates in the stroma. The result is a dense network of interwoven fibers that stabilizes the basement membrane in tissues subject to significant shear forces.
The basement membrane unites cells, facilitating their organization into a layer; maintains cell phenotype and influences different types cellular activity: adhesion, differentiation, polarization, migration. The basement membrane prevents epithelial cells from invading the underlying connective tissue. In malignant neoplasms, disruption of this function leads to the germination of tumor cells and the formation of metastases. The basement membrane filters macromolecules (for example, in the kidney lamina densa traps molecules with M r greater than 50 kDa).
Connective tissue cells are diverse. These are fibroblasts, fibrocytes, chondroblasts, chondrocytes, osteoblasts, osteocytes, macrophages, mast cells, leukocytes, plasmacytes, pericytes, adipocytes. Connective tissue cells are divided into two groups - residents and immigrants. Residents – fibroblasts and fibrocytes (fibrous connective tissue), chondroblasts and chondrocytes (cartilage tissue), osteoblasts and osteocytes (bone tissue), mast cells, adipocytes, macrophages. Immigrants are leukocytes (neutrophils, eosinophils, basophils, monocytes, lymphocytes). When foci of inflammation occur, these cells leave the bloodstream into the connective tissue. Lymphocytes that carry out immunological surveillance constantly circulate between the blood, connective tissues, and lymph.
Functional groups. Depending on the function they perform, connective tissue cells can be divided into three main groups. The cells responsible for the synthesis of extracellular substance molecules and maintaining the structural integrity of the tissue are fibroblasts and fibrocytes, chondroblasts and chondrocytes, osteoblasts and osteocytes, odontoblasts, reticular cells.
The cells responsible for the accumulation and metabolism of fat are adipocytes; these cells form adipose tissue. Cells with protective functions(including immunological) - fat, macrophages and all types of leukocytes.
Fibroblasts and fibrocytes
Fibroblast
Fibroblast (Fig. 6-22 , see also fig. 3 inset) – the most common type of connective tissue cells; secretes components of the extracellular matrix, participates in wound healing, and is capable of proliferation and migration. Fibroblast is a flattened, stellate-shaped cell that forms wide wedge-shaped processes; contains a large oval nucleus with several nucleoli. Cell size is variable. Fibroblast intensively synthesizes protein, so its cytoplasm contains large quantities of cisterns of the granular endoplasmic reticulum, a well-defined complex Golgi, many mitochondria. There are lysosomes and secretory granules, glycogen, numerous microfilaments and microtubules.
Functions. Fibroblasts synthesize collagens, elastin, fibronectin, glycosaminoglycans, proteoglycans and other components of the extracellular matrix. Fibroblasts produce various cytokines—granulocyte-macrophage colony-stimulating factor (GM-CSF), granulocyte colony-stimulating factor (G-CSF), and macrophage colony-stimulating factor (M-CSF). Bone marrow fibroblasts secrete ILZ and IL7. During inflammation and wound healing, fibroblasts are activated by growth factors
Rice. 6-22. Fibroblast ( A) and fibrocyte ( B). Fibroblast (the active form of the cell) contains well-defined organelles: granular endoplasmic reticulum, complex Golgi, mitochondria. The fibroblast forms large elongated processes. In a fibrocyte, there are significantly fewer organelles, the cell is devoid of processes and has a spindle-shaped shape.
bFGF and PDGF proliferate and migrate to the site of injury. Fibroblasts contain various enzymes, including collagenases, that break down collagen. By destroying damaged and synthesizing new macromolecules of the extracellular matrix, fibroblast promotes its restructuring and the formation of scars at the site of damage (inflammation).
fibrocyte
Fibrocyte (Fig. 6-22 ) is a mature form of fibroblast present in dense, formed connective tissue. The fibrocyte has a spindle-shaped shape. The compacted nucleus is elongated and located along the cell. There are scattered cisterns of granular endoplasmic reticulum and a small number of mitochondria. Complex Golgi poorly developed. The cell contains relatively few secretory granules. The function of the fibrocyte is to maintain tissue structure through continuous (albeit slow) renewal of extracellular matrix components. During wound healing, fibrocytes can be stimulated to synthetic activity. An activated fibrocyte acquires the features of a fibroblast: the nucleus is rounded, the number of endoplasmic reticulum cisterns and mitochondria increases; complex Golgi becomes more pronounced.
Pericryptal fibroblasts
The intestinal mucosa contains pericryptal fibroblasts, stromal cells that exhibit the morphological features of SMCs. They are believed to regulate the growth and differentiation of epithelial cells. These fibroblasts express smooth muscle actin.
Myofibroblasts
Myofibroblasts are contractile cells that have common features with MMC (see chapter 7 ).
Macrophages
Macrophage is a differentiated form of monocytes. Macrophages are professional phagocytes; they are found in all tissues and organs. This is a highly mobile population of cells that can move quickly. Life expectancy is months. Tissue macrophages retain some ability to divide (for example, alveolar macrophages in chronic inflammatory processes). In the focus of inflammation, as a result of the fusion of several macrophages, multinucleated giant cells of foreign bodies are formed. Macrophages are divided into resident and mobile. Resident macrophages are present in tissues normally, in the absence of inflammation. Among them, there are free, round-shaped, and fixed macrophages - star-shaped cells, attached with their processes to the extracellular matrix or other cells. Motile macrophages are a population of migrating (called) macrophages.
Mononuclear phagocyte system. Connective tissue macrophages are part of the mononuclear phagocyte system. Cells of the mononuclear phagocyte system differ from other phagocytic cells according to three criteria:
| Rice. 6-23. Macrophage. The kernel is irregular in shape, with notches. The cytoplasm contains ribosomes, mitochondria, multivesicular bodies Well developed complex Golgi and granular endoplasmic reticulum. There are numerous lysosomes. The presence of phagosomes, phagolysosomes, and residual bodies is characteristic. The cell forms cytoplasmic processes involved in migration and phagocytosis. | 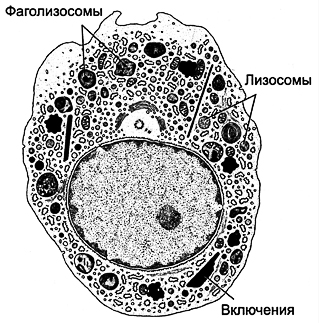 |
have the morphology of macrophages, originate from the bone marrow, phagocytic activity is modulated by Ig and complement components. The mononuclear phagocyte system includes histiocytes (tissue macrophages), alveolar macrophages, osteoclasts, cells von Kupffer, cells Langerhans, cells Hofbauer, foreign body giant cells and probably microglial cells of the central nervous system.
Morphology
Structure (Fig. 6-23 ) of a macrophage depends on its activity and location. Cell diameter is about 20 microns. The kernel is irregular in shape, with indentations. The cytoplasm contains mitochondria, free ribosomes, and a well-defined complex Golgi, multivesicular bodies, granular endoplasmic reticulum, lysosomes, phagolysosomes and residual bodies, the material of which can be released from the macrophage by exocytosis. Lysosomes contain bactericidal agents: myeloperoxidase, lysozyme, proteinases, acid hydrolases, cationic proteins, lactoferrin, superoxide dismutase - an enzyme that promotes the formation of H 2 O 2, OH -, O - 2. Under the plasmalemma, actin microfilaments, microtubules, and intermediate filaments necessary for migration and phagocytosis are present in large quantities. Macrophages migrate along a concentration gradient of many substances coming from various sources. Activated macrophages form irregularly shaped cytoplasmic pseudopodia that participate in amoeboid movement and phagocytosis.
Macrophages are professional phagocytes. They capture denatured proteins and aged red blood cells (fixed macrophages of the liver, spleen, and bone marrow) from the blood. Macrophages phagocytose cell debris and tissue matrix. Nonspecific phagocytosis is characteristic of alveolar macrophages, capturing dust particles of various natures, soot, etc. Specific phagocytosis occurs when a macrophage interacts with an opsonized bacterium. An activated macrophage secretes more than 60 factors. Macrophages exhibit antibacterial
activity, releasing lysozyme, acid hydrolases, cationic proteins, lactoferrin, H 2 O 2, OH -, O 2 -. Antitumor activity consists of the direct cytotoxic effect of H 2 O 2, arginase, cytolytic proteinase, tumor necrosis factor (TNF) of the macrophage. A macrophage is an antigen-presenting cell: it processes Ag and presents it to lymphocytes, which leads to stimulation of lymphocytes and the launch of immune reactions. IL1 from macrophages activates T lymphocytes and, to a lesser extent, B lymphocytes. The macrophage produces lipid mediators - prostaglandin PGE 2 and leukotrienes, platelet activating factor (PAF). The cell also secretes α-IFN, which blocks viral replication. An activated macrophage secretes enzymes that destroy the extracellular matrix (elastase, hyaluronidase, collagenase). On the other hand, growth factors synthesized by macrophages effectively stimulate the proliferation of epithelial cells (transforming growth factor TGFα, bFGF), proliferation and activation of fibroblasts (platelet-derived growth factor PDGF), collagen synthesis by fibroblasts (transforming growth factor TGFβ), and the formation of new blood vessels (fibroblast growth factor bFGF). Thus, the main processes underlying wound healing (reepithelialization, extracellular matrix formation, repair of damaged vessels) are mediated by growth factors produced by macrophages. By producing a number of colony-stimulating factors (macrophages - M-CSF, granulocytes - G-CSF), macrophages influence the differentiation of blood cells.
Mast cell
Mast cells are morphologically and functionally similar to blood basophils, but they are different cell types. The mast cell, like the basophil, originates from a precursor in the bone marrow, but final differentiation takes place in the connective tissue. Mast cells are resident cells of connective tissue. There are especially many of them in the skin, in the mucous membrane of the respiratory and digestive systems, and around blood vessels. Mast cell (Fig. 6-24 ) contains numerous large metachromatic granules (modified lysosomes). Various receptors are built into the cell membrane, including receptors for the Fc fragment of IgE.
Mast cells synthesize and accumulate in granules a variety of biologically active substances, mediators and enzymes: heparin (heparin sulfate), histamine, tryptase, chymase, elastase, dipeptidase, plasminogen activator, acid hydrolases, eosinophil chemotaxis factor (ECF), neutrophil chemotaxis factor (NCF). The main component of mast cell granules is the negatively charged sulfated glycosaminoglycan heparin, synthesized and stored exclusively by mast cells. Heparin secreted by the cell binds antithrombin III circulating in the blood, sharply enhancing its anticoagulant activity. Histamine causes contraction of the SMC, hypersecretion of mucus, and increased vascular permeability with the development of edema. Tryptase promotes the breakdown of fibrinogen,
conversion of C3 to anaphylatoxin C3a, activation of collagenase, degradation of fibronectin. Tryptase, chymase, carboxypeptidase B, other proteases and acid hydrolases, released from the degranulating cell, cause destruction of the tissue matrix. When mast cells are activated (along with the secretion of granule contents), arachidonic acid metabolites are formed - prostaglandins, thromboxane TXA 2 and leukotrienes. These mediators have vaso- and bronchoactive properties. Platelet activating factor (PAF), which is one of the most powerful spasmogens, is also formed from membrane phospholipids.
Mast cells are involved in inflammatory and allergic reactions. Activation and degranulation of mast cells, like basophils, is mediated by IgE (see Fig. 11-4 ). Mast cells have high-affinity surface receptors for the Fc fragments of IgE. The binding of Ag (allergen) to the IgE molecule on the surface of the mast cell is accompanied by exocytosis of the granule contents and the formation of arachidonic acid metabolites.
Plasma cell
Plasma cells (Fig. 6-25 ) differentiate from immunologically activated B lymphocytes, synthesize and secrete Ig (AT). The large round or oval nucleus is located eccentrically and contains dispersed chromatin. In the cytoplasm there is a well-developed complex Golgi and a mass of large cisterns of granular endoplasmic reticulum, concentrically located around the nucleus.
Pericytes
Pericytes are process cells adjacent externally to arterioles, venules and capillaries. Most numerous in postcapillary venules.
Morphology
Pericytes have a disc-shaped nucleus with small depressions, contain the usual set of organelles, multivesicular bodies, microtubules and glycogen. The area facing the wall of the vessel contains bubbles. Contractile proteins are present near the nucleus and in the processes, incl. actin and myosin. Pericytes are covered by a basement membrane, but are closely associated with
endothelial cell, because the basement membrane between them may be absent. Gap and adhesive contacts were identified in these places.
Pericytes synthesize components of the capillary basement membrane; control the proliferation of endothelial cells both during normal vascular growth and during their regeneration; modulate the function of endothelial cells by regulating the transport of macromolecules from capillaries to tissues. During wound healing and vascular restoration, pericytes differentiate into SMCs.
Adipocytes
Among adipocytes, white and brown fat cells are distinguished.
White fat cell
During differentiation, fat droplets appear in the cytoplasm of the mesenchymal cell, merging as their number increases. A differentiated adipocyte is a large round cell with a diameter of 120 microns, containing one large drop of fat, pushing the cytoplasm and all organelles to the periphery. In the cytoplasm, a narrow rim surrounding a drop of fat, there is a flattened nucleus, free ribosomes, smooth and granular endoplasmic reticulum, a complex Golgi and mitochondria. Adipocytes secrete the hormone leptin, the interaction of which with the receptors of nerve cells of the ventromedial and lateral nuclei of the hypothalamus leads to the activation of the central mechanisms of eating behavior; leptin counteracts the accumulation of adipose tissue.
Synthesis and storage of fat. Adipocytes synthesize lipoprotein lipase, which diffuses onto the surface of endothelial cells (mainly in the capillaries of adipose tissue). Dietary lipids are transported into adipose tissue as VLDL and in the form of chylomicrons. Lipoprotein lipase hydrolyzes these lipids to fatty acids. Free fatty acids penetrate adipocytes and are stored in fat droplets in the form of triglycerides. Fat cells are capable of synthesizing fatty acids
also from glucose and amino acids. Insulin stimulates lipoprotein lipase synthesis, fat deposition, and glucose uptake by adipocytes. At the same time, insulin slows down the mobilization of fat from adipocytes, suppressing the activity of enzymes that break down fat. Stored triglycerides are hydrolyzed by hormone-sensitive lipase activated by cAMP. The resulting free fatty acids enter the lumen of the capillaries, where they non-covalently bind to albumin and are transported to the liver.
Brown fat cell
The brown fat cell contains many small fat droplets and large mitochondria. In brown adipocytes, a natural mechanism for uncoupling oxidative phosphorylation functions, which is biologically useful as a way to generate heat. The brown color of cells and tissues as a whole is due to the presence of iron-containing pigments in mitochondria. Activated hormone-sensitive lipase hydrolyzes triglycerides into fatty acids and glycerol. The released fatty acids are metabolized to produce heat. Increased heat generation is explained by the presence of the transmembrane protein thermogenin in the inner membrane of mitochondria. Thermogenin allows the reverse flow of protons previously transported into the intermembrane space without passing through the ATP synthetase system. Thus, the energy generated by the proton current is not used for ATP synthesis, but is dissipated as heat.
TYPES OF CONNECTIVE TISSUE
There are fibrous connective tissues and connective tissues with special properties. Fibrous connective tissues (depending on the number of fibers and the main substance in the extracellular matrix) are divided into loose and dense. In turn, dense connective tissue is divided into formed and unformed, which is determined by the organization of fibers in the extracellular matrix. Connective tissues with special properties include embryonic (mesenchyme), adipose and reticular. These tissues are characterized by a predominance of cells of the same type.
Fibrous connective tissue
Loose connective tissue
Loose (unformed) connective tissue (Fig. 6-26 ) is found in all organs, forms their stroma and accompanies the vessels. This tissue contains relatively few randomly distributed collagen and elastic fibers. Between the fibers there is a large amount of ground substance with various cells immersed in it: fibroblasts, migrating and resident macrophages, mast cells, pericytes, adipocytes, plasma cells, leukocytes. Glycosaminoglycan molecules, intertwining, form a network, in the cells and channels of which a large amount of tissue fluid with dissolved substances is retained.

Figure 6-26 Loose connective tissue The intercellular substance consists for the most part from the main substance with randomly distributed fibers, collagen and elastic. Characteristic cellular diversity (fibroblasts, mast cells, various leukocytes, adipocytes, macrophages, plasma cells)
substances. Leukocytes, macrophages, mast and plasma cells take an active part in protective reactions.
Dense connective tissues
Dense connective tissue contains a large number of densely arranged fibers. The amount of the main substance is relatively small.
Dense, unformed connective tissue comprises large quantity densely but randomly arranged fibers. Fibroblasts (fibrocytes), macrophages, and mast cells are present between the fibers. Characteristic of the skin itself, the periosteum.
Dense formed connective tissue. The fibers are arranged tightly, forming parallel bundles. In the narrow spaces between the fibers, fibrocytes line up in chains. Ligaments, tendons and fibrous membranes are formed from such tissue.
- Elastic bunch(For example, ligamenta flava And ligamentum nuchae) consists of elastin, which forms thick fibers. Between them are thin collagen fibers and fibrocytes.
- Tendon(rice. 6-27 ) consists of collagen fibers that form tendon bundles of the I, II and III orders. Between the first order bundles there are rows of fibrocytes (tendon cells) with lamellar processes. Groups of first order bundles, surrounded by loose connective tissue with vessels and nerves, form second order bundles. Several bundles of the second order are united by loose connective tissue into bundles III order. When a tendon is damaged, activated fibrocytes and fibroblasts synthesize collagen for new fibers.
- Fibrous membranes- a type of dense, shaped connective tissue. Bundles of collagen fibers and fibrocytes lying between them are arranged in layers. In each layer, wavy bundles of collagen fibers run parallel in one direction, different from the directions in adjacent layers. Individual bundles of fibers pass from one layer to another, connecting them together. Fibrous membranes include fascia, aponeuroses, tendon center of the diaphragm, capsules internal organs, dura mater, sclera.
Connective tissues with special properties
Connective tissues with special properties include mesenchyme, reticular and adipose tissue.
Mesenchyme
Embryonic connective tissue - mesenchyme (Fig. 6-28 ) is the source of origin of cells of all connective tissues. Mesenchymal cells are stellate or spindle-shaped with delicate branching processes that form a network. In the oval core – dispersed

Figure 6-27 Tendon A– cross section B– longitudinal section IN And G– diagram of the structure of the tendon in transverse and longitudinal sections
chromatin and clearly visible nucleoli. The gel-like extracellular material consists almost exclusively of ground substance and a minimal amount of scattered fibers (mainly reticulin).
Reticular tissue
Reticular tissue (Fig. 6-29 ) has a network-like structure and consists of reticulin fibers and reticular cells with long processes. Reticular cells together with reticulin fibers form a loose network. Reticular tissue creates a specific microenvironment for developing blood cells; in addition, reticular cells synthesize various hematopoietic growth factors (ILZ, IL7, GM-CSF, G-CSF, M-CSF).
Adipose tissue
Adipose tissue is found in many organs. There are white and brown adipose tissue.
White fat makes up almost all of the body's adipose tissue. Participates in the absorption from the blood, synthesis, storage and mobilization of neutral lipids (triglycerides). The distribution of adipose tissue in the body is influenced by sex hormones and hormones of the adrenal cortex. Fat cells (adipocytes) form clusters (lobules), separated by partitions of loose connective tissue. In the latter, blood vessels and nerves pass into the adipose tissue. Individual fat cells are surrounded by a network of reticulin and collagen fibers. Fibroblasts and mast cells are present in connective tissue septa.
Brown fat in a newborn it is involved in thermoregulation. In adults, brown fat is found in small quantities in the mediastinum, along the aorta and under the skin between the shoulder blades. Brown adipose tissue is abundantly supplied with blood capillaries, forming a network around each adipocyte, and has pronounced sympathetic innervation.