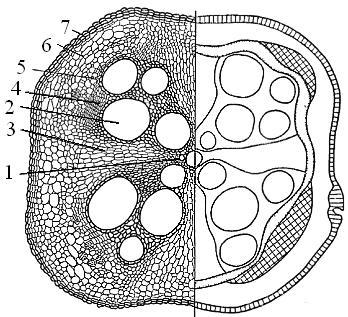
първична структура на корена на напречната секция на корена на ириса. На среза, дори при слабо увеличение, ясно се разграничават малката вътрешна част - централният цилиндър и външната първична кора, покрита с един слой клетки с коренови косми - ризодермата (епиблема). Тъй като кореновите косми умират, стените на външните клетки на първичната кора са покрити от вътрешната страна с тънък слой суберин. Суберинизацията определя непроницаемостта на клетките нито за вой, нито газове. Тези няколко слоя на кортикалните клетки се наричат \u200b\u200bекзодерма, тя изпълнява f-тата тъкан на обвивката. (Външният слой на първичния кортекс, екзодермата, се състои от плътно затворени полигонални клетки, стените на които впоследствие са запушени и изпълняват защитна функция). Под екзодермата се намират живи паренхимни клетки (паренхим) на първичната кора, разположени свободно и образуващи междуклетъчни пространства.
Вътрешният слой на първичната кора - ендодермата се състои от един ред клетки, с удебелени радиални и вътрешни стени. Стените на ендодермалните клетки повече или по-малко корк, а в някои растения (ирис) лигнифицират. Но не всички клетки на ендодермата имат непроницаема стена; сред дебелостенните клетки на места има тънкостенни живи клетки, наречени проходни клетки, защото именно чрез тях водата влиза в централния цилиндър. Външният слой на централния цилиндър - на велосипеда - се състои от един ред живи паренхимни клетки. Централната част на централния цилиндър е заета от радиален проводим лъч. Ксилемата е разположена в центъра и образува серия от остри издатини под формата на лъчи. Между издатините на ксилемата са разположени участъци от флоемата. 72. Напречен разрез на корена на ириса (Iris germanica), 1 - ризодерма (епиблема), 2 - екзодерма, 3 - основен паренхим (мезодерма), 4 - ендодерма, 5 - проходна клетка на ендодерма, 6 - цикъл, 7 - първичен лъч на ксилема, 8 - секция на първичната флоема (2-5 - първична кора, 6-8 - централен цилиндър). ,
Вторична анатомична структура на корена. Коренови култури. 1. При завършената подготовка на напречно сечение на тиквен корен се изследва вторичната анатомична структура на корена. При ниско увеличение в центъра на цилиндъра се намира четирилъчен ксилем. От лъчите на първичната ксилема започват радиални (ядрени) лъчеви места на тънкостенния жив паренхим. Широките участъци на вторичния ксилем с големи съдове и дребноклетъчен паренхим от дърво се редуват с радиални лъчи. На границата на вторичния ксилем се вижда ясно камбият. Извън камбиума, срещу всяко място на вторичния ксилем, има вторична флоема. Отвън коренът е покрит със сравнително тънък слой корк. Тъканите, разположени извън камбия (флоема, първичен паренхим, фелодерма и корк камбий) се наричат \u200b\u200bвторична кора.
 Фиг. 73. Вторичната структура на тиквения корен (Cucurbita pepo):
Фиг. 73. Вторичната структура на тиквения корен (Cucurbita pepo):
A е диаграма на напречно сечение (отляво е подробен чертеж, отдясно е схематично); Б - фрагмент от картината.
1 - първичен ксилем, 2 - вторичен ксилем, 3 - радиален лъч, 4 - камбий, 5 - първична и вторична флоема, 6 - първичен паренхим на вторичната кора, 7 - перидерма (1-3 - ксилем, 5-7 - вторичен кортекс) ,
2. На микропрепарати и живи кореноплодни моркови, репички, цвекло, проучете естеството на анатомичната структура и установете вида на кореноплодите. В напречното сечение на корена на моркова може да се разграничи лек пръстен, който очертава по-широката част от по-малката вътрешна. Това е камбият. Отвън е вторичната кора, а навътре е ксилемата. Две радиални паренхимни лъчи се простират от лъчите на първичния ксилем (диархален сноп), а между тях са поставени две ветрилообразни секции на вторичния ксилем. Вторичният кортекс се състои от паренхим и малък брой ситови тръби с придружаващи клетки. Следователно, резервните продукти в корена на морковите се отлагат във вторичната кора.
Когато разглеждат напречен участък от кореновата култура, репичките намират в центъра два-лъчева първична ксилема. Две радиални паренхимни лъчи се отдалечават от нейните краища, след това две участъци от вторичния ксилем, камбиевия слой и накрая вторичната кора. Отбелязва се, че най-голямата площ в напречното сечение е заета от нелигинизиран паренхим, разположен навътре от камбиалния пръстен, т.е. ксилемовия паренхим и радиалните лъчи. В този паренхим има и резервни продукти. Съдовете, заобиколени от пръстен на лигнифициран паренхим, са разположени в разпръснати групи по радиусите сред складиращата тъкан.
Когато се разглежда напречното сечение на корена на цвеклото в самия център, се вижда двулъчева първична ксилема, към която прилежават два участъка от вторичния ксилем, разделени от радиални паренхимни лъчи. Следва камбият със съседни участъци от вторичната флоема. След това идва слой от допълнителни камбиални пръстени. те се образуват благодарение на клетъчното делене на велосипеда, в който се образува слой от паренхимни клетки. В този слой един ред клетки започва да се дели от тангенциална септа и се превръща в нов слой камбий. Камбиалният слой поставя ксилемата навътре и флоемата навън, които образуват малки колатерални проводящи снопове, разделени един от друг чрез слой от тънкостенни паренхими. В същото време в периферния слой на паренхима се образува нов пръстен от камбиални клетки и др.
по този начин твърди слоеве върху напречната секция на кореновата култура са ксилемата на концентричните редове от проводими снопове, а сочните широки слоеве се състоят от камбий, флоема и паренхим за съхранение. Зеленчуковият корен от зеленчуци е многокамбиален.

Фиг. 16. Вторичната структура на корена на морковите (Daucus carota): 1 - перидерма; 2 - паренхим на вторичната кора; 3 - камбий; 4 - вторичен ксилем; 5 - първичен ксилем; 6 - радиален лъч; 7 - първична и вторична флоема.

Фиг. 17. Вторичната структура на корена на репичката (Raphanus sativus): 1 - перидерма; 2 - паренхим на вторичната кора; 3 - вторичен ксилем; 4 - първичен и вторичен флоем; 5 - камбий; 6 - радиален лъч; 7 - първичен ксилем

Фиг. 18. Структурата на поликамбиалния цвеклов корен (Beta vulgaris): 1-перидерма; 2 - допълнителни слоеве камбий; 3 - вторичен ксилем; 4 - радиален лъч; 5 - първичен ксилем; 6 - камбий; 7 - първична и вторична флоема; 8 - обезпечителни проводящи снопове
Такава структура, когато има централен лъч в централния цилиндър, изградена от ксилем и флоема, се нарича първичен.
При едносемеделни растения тази първична структура на корена се запазва през целия живот. Тъй като коренът се развива и расте, те притежават тъканта на ендодермата и клетките на велосипеда. Корените на еднодолни не са способни да се сгъстяват, тъй като съдовите им снопове са затворени - нямат камбий.
При двуцветни цъфтящи растения, както и при иглолистни растения и други гимнастици корените са способни да се сгъстяват. В снопа им се образува камбик, отлагащ елементи от ксилем и флоема. Отлагането на нови вторични елементи води до промяна в състава и анатомичната структура на други участъци от корена. В резултат на дейността на камбия се нарича коренна структура вторичен, Камбият се среща в паренхимната тъкан под сноповете на флоемата и над лъчите на ксилема ( "Камбиални храмове") или по-скоро между ксилемата и флоемата се образува непрекъсната лента от камбиални клетки. Паренхимните клетки придобиват меристематичен характер - те се разширяват и стават таблични. След известно време се образува непрекъснат пръстен камбий, първоначално имащи неравномерни усукани контури (фиг. 63, 2). Камбиевите клетки започват да лежат вътре в елементите на вторичния ксилем, отвън - в елементите на вторичната флоема. Отлагането на ксилема е по-бързо, участъци от първичната флоема често са изтласкани към периферията, поради което пръстенът е подравнен. В лигнатите растения по-нататък се отлагат плътни пръстени от ксилем и флоема и кореновата структура става напълно подобна на структурата на стъблото. Въпреки това, винаги в корена, в централната му част, са запазени останките на първичната ксилема - радиален лъч под формата на кръстовидни лъчи. На тази основа е лесно да се разграничи коренът от стъблото под микроскоп. Първична флоемаестествено се изтласква към самата периферна част и поради по-малкия си обем е неразличим в корен за възрастни.
С нарастването на корена, в неговата сърцевина настъпва промяна. Епидермисът се нулира и заменя periderm (корк), произтичащи от велосипеда. Първичната кора също се отделя. На много стари корени на дърветата се образува кора. По това време ендодермата, както и периклимът, умират и образуването на нови слоеве на перидермата става в по-дълбоките слоеве на корена - във вторичната флоема. По този начин във вторичната структура коренът няма кора, ендодерма или колело и е външно покрит с перидерма или кора (фиг. 63, 3 ). Материал от сайта
На тази страница материал по темите:
първичен коренова структура характеризиращ се с наличието на три тъканни системи - поглъщащо покритие епидермис с коренови косми (епиблема), първична кора и централен цилиндър rhizodermisТой изпълнява смукателна и защитна функция, състои се от един слой от тънкостенни, плътно затворени клетки с коренови косми, няма стомаси, дебели кутикули и трихоми. Първична кораЛежейки под елибелата, тя се разделя на екзодерма, мезодерма и ендодерма. exodermСъстои се от 3-4 слоя големи, плътно затворени клетки с частично запушени мембрани. Изпълнява защитни и пропускащи функции. mesoderma- част за многослойно съхранение, представлява основната част от първичната кора. Мезодермата изпълнява също транспортни и въздушни функции. ендодерма- най-вътрешният, обикновено едноредов слой на първичния кортекс, граничещ с централния цилиндър, изпълняващ функцията за поддържащо преминаване. Централна,или Аксиален цилиндърТой заема средната част на корена, заобиколен е от пръстен на колелото, от който се образуват страничните корени.В корена няма истинско ядро. В центъра могат да бъдат съдове, склеренхим, понякога - паренхим за съхранение, образуване Грешно ядро Вторична коренова структура Образува се поради появата и функционирането на вторичните странични меристеми - камбий и пелоген.Камбият се образува от перикарда над лъчите на първичния ксилем и от прокамбиума или паренхима под флоемата. Външно деленият камбий поставя вторична флоема, а към центъра - вторичен ксилем. Първичната ксилема се изтласква към оста на корена, а първичната флоема към периферията на централния цилиндър. Вторичната структура на корена може да бъде от типа на лъча или без кости Тип лъчТой се формира, ако междулъчевият камбий, образуван над лъчите на първичния ксилем, произвежда паренхима на основните лъчи, а лъчевият камбий, лежащ под флоемата, произвежда вторичната флоема и ксилемата на колатералния или двуколтерален проводящ лъч. при Тип нокаутиранеПръстеновата структура на камбия образува непрекъснати пръстени от ксилем и флоема. Вторичните трансформации в първичната коренна кора са свързани с активността на фалогена. Ако възниква от перикела, тогава коркът, образуван отвън, изолира първичния кортекс и той се отхвърля, обезмаслява се. Ако фалогенът се образува от паренхима на мезодермата, което е рядко, тогава частта от първичната кора, лежаща под фалогена, се запазва. При дървесните растения коренът обикновено е от безкръвен тип и в дървесината се образуват едногодишни растежни пръстени годишен пръстен- Това е колекция от тъкани, образувани от камбия през един вегетационен период. Покривната тъкан на дървесните корени може да бъде перидермата или кората. Наличието на радиален проводящ сноп, склеренхим или съдове от първичния ксилем в центъра на органа разграничава корена от стъблото и коренището, при което централната част е заета от истинско ядро.
Външният слой на първичната кора, граничеща с епилемата, наречена екзодерма, се състои от плътно затворени клетки. Повечето надлъжни радиални прегради между тях не представляват разширение на радиалните дялове на епиблемата, а се редуват с тях (фиг. 161, 2 , 162).
При много растения клетъчните стени на екзодермата обикновено се подлагат на вземане на проби по време на смъртта на епиблемата и тогава екзодермата се превръща в покриващ, защитен слой на корена, подобен в по-голяма или по-малка степен на кожата на стъблата и листата.
аз - космат слой (епиблема) с коренови косми; II - първична кора с екзодерма ( 2 ) и ендодерма ( 3 ), 4 - колан Caspari; III - аксиален цилиндър в него: 5 - велосипед, 6 - първичен ксилем, 7 - сърцевина 8 - камбий, 9 - първична флоема (според Муромцев).
В зависимост от средата, в която се намира коренът, и от продължителността на функционирането на екзодермата, тя достига различна степен на специализация.
Така че, в корена на някои водни едносемеделни растения (например воден цвят), екзодермата не се диференцира, тъй като перидермата се образува много рано.
В някои едносемейни с мощни корени (в фурмата с палто, в канната), напротив, клетките на екзодерма в ранен етап се делят тангенциално
чрез дялове: образува се екзодерма, състояща се от 2-20 слоя клетки. Многослойната екзодерма прилича на коркова тъкан, но се различава от нея не само по начина, по който се образува, но и по подредбата на клетките: в надлъжни радиални участъци на корена те не представляват редовни редове.
Екзодермата е характерна главно за едносемеделни растения, чиито корени нямат вторично удебеляване; първичната кора трае много дълго време и не се образува перидермис.
Под екзодермата се намира останалата част от първичната кора; най-вътрешният слой, граничещ с аксиалния цилиндър, се нарича ендодерма.
Обикновено първичната кора се състои от живи паренхимни клетки с тънки целулозни стени. Във външната област клетките му са многостранни, плътно затворени, докато клетките във вътрешната кора са закръглени до една или друга степен и между тях се образуват междуклетъчни пространства, пренасяни с въздух - много малки (в корените на много китове) или по-големи. Размерът на клетките обикновено се увеличава в посока от екзодермата към средната част на първичната кора и след това намалява в посока към ендодермата (фиг. 162). При някои растения (в ириса), други 2-3 външни слоя от първичната кора са подложени на коркиране, в допълнение към екзодермата. В много големи зърнени култури клетъчните мембрани на слоя, съседен на екзодермата, особено външните тангенциални стени, са силно уплътнени, свински и дървесни.
В корените, развиващи се във въздуха или във водата, клетките на първичната кора съдържат пластиди с хлорофил.
В паренхимните клетки на първичния кортекс се натрупват големи запаси от хранителни вещества не само в монокотиледони, но и в някои

Фиг. 162. Част от напречното сечение на корена на ириса ( Iris germanica):
ePB - епиблема; инд - трислойна екзодерма; пУР - паренхим на съхранение на първичната кора; край - ендодерма; стр. CL, - пропускателна клетка; ми - велосипед; p.ks, - първичен ксилем; п. ет, - първична флоема; т. МК, - механична тъкан.
двудолни, при които първичната кора се запазва за дълго време, например в пеперудата Ficaria ranunculoides), борецът ( Aconitum napellus). В тези случаи първичната кора е много мощна.
В областта на първичния кортекс се образува папилимулна амигдала - сегментирана (на звънци) или несегментирана (в гръцкия paulochemus) Periploca graeca), групи от каменисти клетки (в артикула в Йерусалим), склеренхимни шнурове (в палмова фурна). В много зърнени култури (включително суданска трева) два или повече слоя от външната първична кора се диференцират в склеренхим, понякога много дебелостенни.
Много растения, особено водни (каламус) и тези, които живеят на блатисти почви (някои сортове ориз, отглеждани на места, наводнени с вода), образуват големи въздушни канали в първичната кора. Експериментите показаха, че силата на въздушната система е обратно свързана с аерирането на субстрата. И така, при експерименти с царевица Бял зъб и пшеница маркиза във водните култури, двете растения са имали мощни дихателни пътища. В културите в пясък и почва проходимите от въздуха въобще не се образуват в пшеница, но в царевицата те се развиват, но слабо. При отглеждане на царевица във водни култури се оказа, че аерацията значително отслабва процеса на образуване на въздушни проходи.
Вътрешният слой на първичната кора, наречен ендодерма, се състои от плътно затворени паренхимни клетки с характерна структура. В младо състояние ендодермата е в напречно сечение пръстен от правоъгълни клетки с тънки целулозни стени, с кухини, пълни с живи съдържания; надлъжните разрези показват, че ендодермата се състои от дълги и къси клетки; къси или разпръснати между дълги или подредени в надлъжни редове. По-късно ендодермата преминава през един, два или три етапа на диференциация.
На първия етап настъпват биохимични промени в напречните и надлъжните радиални стени на клетките на ендодермата, покриващи средната лента на стената; в този раздел стената, подлагайки се на вземане на проби и лигнификация, се диференцира в рамка или пояс (Caspari пояс), който протича непрекъснато от четири страни на клетката. Каспари поясите на съседни ендодермални клетки са в непосредствена близост една до друга (фиг. 163).
В ендодермата, която навлиза във втората фаза на диференциация, върху обвивката от вътрешната страна се отлага непрекъснат сгъстяващ слой, състоящ се от целулоза и суберин. Въпреки това, някои клетки срещу групите ксилеми на аксиалния цилиндър (проходните клетки) остават на първия етап (фиг. 162).
Ендодермата на не всички растения навлиза във втория етап: в хвощ и в

Фиг. 163. Част от напречното сечение на корена на каустичния пеперуда ( Ranunculus acer):
добре - ендодерма с колани от каспари ( pC); ми - велосипед; е - флоема; к - ксилем.
някои папрати, неговото развитие е ограничено до първия етап. При teretanosperms и двудолни, ендодермата обикновено преминава през втория етап. При някои двудолни и в повечето монокотиледони той навлиза в третия етап. На този етап, в клетките на ендодермата, лигнифициран целулозен слой се отлага навътре от суберинната плоча. Отлагането обикновено е неравномерно: слабо е на външната тангенциална стена, усилено на радиалната и напречната стени в посока към центъра на корена; вътрешната допирателна стена се сгъстява в най-голяма степен. Описаното сгъстяване обикновено е ясно слоесто (фиг. 162).
В ендодермата на първия етап проходните клетки на някои растения обикновено остават тънкостенни и живи известно време и преминават вода с разтворени вещества в нея по посока на кореновия радиус (фиг. 162), но след това преминават към втория и третия етап. При някои зърнени култури клетките на един или два слоя на първичния кортеж в съседство с ендодермата стават много подобни на клетките от третата степенна ендодерма (фиг. 164). При някои растения (например зърнени култури) цялата първична кора на някои корени се подлага на склерификация с течение на времето - сгъстяване и лигнификация на клетъчните стени. След като ендодермата преминава в третия етап, първичната кора понякога умира и ексфолира. При дикотиледоните смъртта и ексфолиацията на кората настъпва в резултат на силно вторично удебеляване и образуване в перикела (виж по-долу) на перидермата и с ендодерма във втория или дори в първия етап.