Sidekudosta kutsutaan myös sisäiseksi kudokseksi. Se on osa kutakin elintä ja muodostaa välikerrokset elinten välille ikään kuin yhdistäisivät ne. Sidekudos pukeuttaa verisuonia ja hermoja, osallistuu ihmisen luurankon ja sen yksittäisten elinten luurankoon, veren ja imusolmukkeiden muodostumiseen.
Sidekudos suorittaa seuraavat toiminnot: troofiset, suojaavat, tukevat (mekaaniset) ja muovit.
trofiatai ravitsevaa, toiminto Se koostuu siitä, että sidekudokseen kuuluva veri kuljettaa ravintoaineita kehossa. Lisäksi sidosastiat ja sidekudokset tunkeutuvat kaikkiin kudoksiin ja elimiin.
Suojaava toiminta sidekudos liittyy paitsi sen mekaanisiin ominaisuuksiin (luut - tiheät muodostelmat - suojaavat elimiä), mutta myös siihen, että sen soluilla on kyky fagosytoosiin: ne imevät ja sulavat haitallisia aineita. Sidekudos osallistuu myös suojaavien kappaleiden muodostumiseen, jotka luovat immuniteettia (immuniteettia sairauksille).
Viitetoiminto sidekudoksen määrää pääasiassa solujen välinen aine.
Muovitoiminto sidekudos ilmaistaan \u200b\u200bsen suuressa kyvyssä uudistua ja sopeutua ympäristöolosuhteisiin. Tämä kudos muodostuu mesodermin keskimmäisestä sukuslehdestä, ns. Sukusoluista (mesenkyymi).
Sidekudos koostuu soluista ja solujenvälisestä aineesta, joissa pääaine ja kuidut erittyvät. Toisin kuin muun tyyppiset kudokset, solujenvälinen aine on pääosin siinä, kun taas soluja on vähän. Eri tyyppisissä sidekudoksissa solujen välisten aineiden ja solujen kvantitatiivinen suhde on erilainen.
Sidekudoksen pääaine sisältää monia kuituja. Jotkut niistä, jotka sijaitsevat paksuina suorina tai useina puristenauhoina, eivät haaroitu, koostuvat erityisestä liima-aineesta ja niitä kutsutaan kollageeni- tai liimakuiduiksi. Ne ovat huonosti vetolujuisia, erittäin kestäviä. Toinen kuitutyyppi on elastinen. Ne ovat hienovaraisempia, haarautuvia. Nämä kuidut ovat vähemmän kestäviä kuin kollageenikuidut, mutta niillä on suurempi joustavuus ja kimmoisuus (ne, kuten kumi, kykenevät venymään ja saamaan sitten alkuperäisen muodonsa).
Pääkudossolut ovat fibroblasteja, fibrosyyttejä, makrofageja, syöttösoluja ja plasmasoluja. Se voi sisältää rasvasoluja, pigmenttisoluja ja jopa valkosoluja.
fibroblasteja - sidekudossolujen päätyyppi. Niillä on epäsäännöllinen tai karan muotoinen (pitkänomainen) muoto. Ydin on melko suuri, muodoltaan soikea. Fibroblastit osallistuvat solujen välisen aineen ja kuitujen muodostumiseen, haavan paranemiseen ja arvakudoksen kehitykseen. Elämänjaksonsa loppuun saaneita fibroblasteja kutsutaan fibrosyyteiksi.
makrofagit voi olla erimuotoista: pyöreä, pitkänomainen, epäsäännöllinen. Heidän kuori on taitettu, ja siinä on suuri määrä mikroviiluja, joiden avulla ne vangitsevat vieraita aineita. Näillä soluilla on yleensä yksi ydin, jonka koko on pieni, soikea tai papun muotoinen. Makrofaagit ovat ihmiskehon tärkeimpiä puolustajia. Niissä tuhoutuvat mikrobit, myrkylliset (myrkylliset) aineet neutraloidaan.
Mastosolut on epäsäännöllinen muoto, lyhyet laajat prosessit, pieni ydin. Sytoplasmassa on paljon rakeisuutta. Mastosoluilla on hyvin kehittynyt kyky ameebamaisiin liikkeisiin. He osallistuvat solujen välisen aineen muodostumiseen ja sen koostumuksen säätelyyn, tuottavat aineita, jotka estävät veren hyytymistä ja suolojen laskeutumista verisuonten seinämiin.
Plasmasolut soikea tai pyöreä muoto osallistuu suojakappaleiden muodostumiseen, etenkin kun ne vievät vieraita proteiineja kehoon.
Rasvasolut sisältävät rasvaa sytoplasmassa, työntäen ytimen reuna-alueelle. Niiden lukumäärä löysässä sidekudoksessa on epäjohdonmukainen. Parannetulla ravinnolla rasvasolujen määrä kasvaa dramaattisesti.
Pigmenttisolut - nämä ovat samoja fibroblasteja tai fibrosyyttejä, joiden sytoplasmassa on paljon väriaineita - pigmenttiä.
Suoritettujen toimintojen mukaan, jotka suuressa määrin määrittelevät solujenvälisen aineen fysikaalis-kemialliset ominaisuudet (se voi olla nestemäinen, tiheä ja erittäin kiinteä), sidekudos jaetaan suoja-troofisiin ja tukeviin. Suojaavaan troofiseen sidekudokseen kuuluvat: veri, imusolmukkeet, retikulaariset tai reticular-kudokset, kudokset, löysät kuitukudokset ja endoteelit. Tukiosana oleva sidekudos sisältää: tiheän kuitukudoksen, ruston ja luukudoksen. Kun solujen välinen aine tiivistyy, kudoksen trofinen toiminta heikkenee ja tukitoiminto kasvaa.
veri on eräänlainen sidekudos, jossa on nestemäinen solujen välinen aine ja spesifiset solut. Veren solujen välinen aine on sen nestemäinen osa - plasma, jossa veren muodostetut elementit (solut) sijaitsevat. Tilavuuden mukaan plasma on 55-60% ja muotoillut elementit 40-45% kaikesta verestä. Aikuisen 4,5-5 litraa verta.
Plasma. Veriplasma koostuu epäorgaanisista ja orgaanisista aineista. Epäorgaanisia aineita on noin 91% (90% on vettä ja 1% mineraaleja) ja orgaanisia on noin 9%. Pääosa orgaanisista aineista on proteiineja - 7%. Niitä on 3 tyyppiä - fibrinogeeni, albumiini ja globuliinit. Fibrinogeeni osallistuu veren hyytymiseen, veteen liukenemattomat albumiinin kuljetusaineet (mukaan lukien lääkkeet) ja globuliinit tarjoavat suojaavien kappaleiden muodostumisen. Globuliinien määrä kasvaa jyrkästi tartuntatauteissa. Veriplasmaa, jossa ei ole fibrinogeenia, kutsutaan veriseerumiksi. Sitä käytetään terapeuttisiin tai ennalta ehkäiseviin tarkoituksiin immuniteetin luomiseksi (passiivinen immunisointi), ja terapeuttiset seerumit valmistetaan. Luonnonmukaisia \u200b\u200borgaanisia aineita (urea, rasva, aminohapot jne.) Sisältyy myös veriplasmaan, tosin hyvin pieninä määrinä.
Verisolut. Niitä on kolme tyyppiä: punasolut - punasolut, valkosolut - valkosolut ja verihiutaleet - verihiutaleet.
Punasolut (erytros on punainen, sytos on solu) ovat spesifisiä voimakkaasti erilaistuneita soluja, jotka menettivät ytimen, mitokondriat, retikulaarisen laitteen ja solukeskuksen kehityksen aikana (kuva 6). Sammakoissa, kaloissa, linnuissa punasolut sisältävät ytimiä (kuva 7). Punasolujen sytoplasmassa on monimutkainen proteiini - hemoglobiini, jonka avulla kehossa tapahtuu kaasunvaihto: happi siirtyy keuhkoista kudoksiin ja hiilidioksidi siirretään kudoksista keuhkoihin. Punasolujen kalvo on erittäin ohut, ja sen kautta tapahtuu kaasujen vaihto. Punasolussa on kaksoismurtaisen levyn muoto, joka lisää sen pintaa lisääen hemoglobiinin parempaa kosketusta kuljetettujen kaasujen kanssa. Ytimen puuttuminen solusta, ilmeisesti, myötävaikuttaa myös suurempaan hapenottoon.
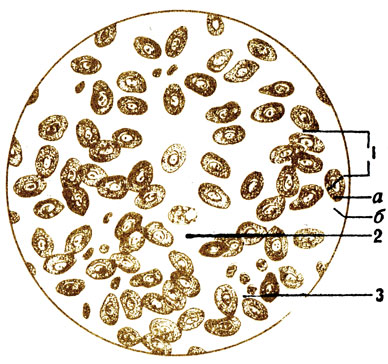
Punasolujen koko on pieni, vain 7-8 mikronia, joten ne kulkevat melko helposti ohuimmissa verisuonissa - kapillaareissa. 1 millimetrissä 3 verta sisältää 4,5-5,0 miljoonaa, ja kaikessa veressä - 25 - 28 biljoonaa punasolua. Jos olisi mahdollista laajentaa niitä vierekkäin, niin saisimme ketjun, joka riittää ympäröimään maapallon 3 kertaa päiväntasaajan ympäri. Veressä kiertävien punasolujen kokonaispinta-ala on yli 1/4 hehtaaria. Miehillä punasolujen määrä on hiukan suurempi kuin naisilla; lapsilla - enemmän kuin aikuisilla; korkeiden vuorten asukkaat, joissa ilmassa on vähemmän happea, enemmän kuin tasangon asukkaat. Jopa lyhytaikaisen (1–2 kuukauden) oleskelun aikana vuorilla punasolujen määrä kasvaa, mikä on tärkeää urheiluharjoituksen suorittamiselle siellä. Lihasaktiivisuuden lisääntyessä ne kasvavat myös lisääntyneen hapenkulutuksen vuoksi. Punasoluilla ei ole kykyä liikkua itsenäisesti, ne liikkuvat verisuonissa verivirtauksella. Ne ovat kuitenkin erittäin joustavia, kun niitä liikutetaan kapillaareissa, on selvästi nähtävissä, kuinka ne pitkittyvät, litistyvät ja muuttavat muotoaan. Punasolujen elinkaari on 80 - 120 päivää. Pernan punasolut hajoavat ja muodostuvat punasoluun. Raporttien mukaan 1/100 punasoluista tuhoutuu päivittäin, ts. Hieman yli 3 kuukautta myöhemmin, kaikki punasolut päivitetään.
Valkosolut - nämä ovat soluja, joissa on ydin. Ne ovat suurempia kuin erytrosyytit (jopa 10 mikronia), kykenevät itsenäiseen amoeboidiseen liikkeeseen, voivat jättää kapillaareja alla olevaan kudokseen.
Sytoplasman luonteesta riippuen, sulkeumien esiintyminen siinä proteiinin jyvien muodossa, pigmentti, ja myös ytimen muodosta riippuen, valkosolut jaetaan rakeisiksi ja ei-rakeisiksi. Edellisillä on rakeisuus sytoplasmassa ja segmentoitunut, jaettu erillisiin osiin, ydin. Rakeistyypistä riippuen erotetaan sen suhde väriaineisiin, neutrofiileihin, basofiileihin ja eosinofiileihin, joilla on tietyissä määrällisissä suhteissa veressä. Muuttamalla tätä suhdetta eri sairauksiin, ei vain luonne, vaan myös lopputulos määritetään.
Sytoplasman inkluusioiden leukosyyttien ei-rakeiset muodot eivät sisällä, niiden ydin ei ole jaettu osiin, sillä on pyöristetty muoto ja se sijaitsee useimmiten solun keskustassa. Ei-rakeiset valkosolut sisältävät lymfosyytit ja monosyytit.
1 mm 3: n veressä on 6-8 tuhat leukosyyttiä. Niiden lukumäärä voi kasvaa syömisen jälkeen tartuntatauteilla ja etenkin lisääntyneen lihaksen toiminnan jälkeen. Leukosyyttien elinajanodote vaihtelee: useista päivistä 2 - 3 kuukauteen. Kun infektio pääsee kehoon, he kuolevat huomattavan määrän taistelussa sitä vastaan. Valkosolujen rakeiset muodot tuotetaan punasoluissa ja lymfosyytit pernassa ja imusolmukkeissa.
Valkosolujen päätehtävä on suojaava. Ne valvovat kehon terveyttä ja auttavat sitä torjumaan erilaisia \u200b\u200bsairauksia. Ne suorittavat suojaavan toiminnan osallistumalla fagosytoosiin ja suojaavien kappaleiden muodostumiseen. Lisäksi valkosolut tuottavat entsyymejä, jotka säätelevät veren hyytymistä ja verisuonien läpäisevyyttä. Lopuksi, lymfosyyttien yksittäiset muodot voivat muodostaa erityyppisten sidekudosten soluja (fibroblasteja, makrofageja, sileälihassoluja), mikä on tärkeää palautumisprosessissa.
verihiutaleettai verilevyt, on pyöristetty tai soikea muoto, vain kooltaan 1-2 mikronia. Ne eivät sisällä ydintä. Niitä on 200-300 tuhatta 1 mm3: n veressä. Verihiutaleiden elinajanodote on 5-8 päivää. Verilevyt osallistuvat veren hyytymiseen.
imuneste, kuten veri, koostuu nestemäisestä osasta - lympoplasmasta - ja yhtenäisistä elementeistä. Toisin kuin veriplasmassa, siinä on vähemmän proteiineja, mutta enemmän aineenvaihduntatuotteita. Muodostuneista alkuaineista lymfosyytit vallitsevat siinä, punasolut puuttuvat.
Verisuonikudos koostuu epäsäännöllisen muotoisista soluista. Yhteydessä toisiinsa he muodostavat eräänlaisen verkon. Tämän verkon silmukoissa on solujen välinen aine, joka sisältää suuren määrän retikuliinikuituja, jotka punostavat solujen pintaa. Verisolut (luuydin, perna, imusolmukkeet) rakennetaan retikulaarisesta kudoksesta.
Löysä kuituinen sidekudos - tämä on kudos, jossa kaikki sidekudoksen rakenteelliset elementit ilmenevät selkeimmin: solujen välinen aine, kuidut ja solut (kuva 8). Se peittää suonet ja hermot, muodostaa ihonalaisen kudoksen, osallistuu melkein kaikkien elinten rakenteeseen.
Endoteelikudos (endoteeli) tarkoittaa vain alkuperästä sidekudosta, kun taas rakenteeltaan se muistuttaa epiteelikudosta. Sen solut ovat litteitä, sijaitsevat kellarimembraanilla. Tämän kudoksen solunsisäinen aine on pieni. Endoteeli linjaa verisuonten sisäpintaa antaen sille sileän, kiiltävän ulkonäön; kapillaarien metabolian endoteelisolujen kautta; ne suorittavat suojaavan toiminnan.
Tiheä kuituinen sidekudos sillä on ominaispiirre - siinä on pääosin kollageenikuituja, jotka kerätään kimppuina, jotka on suunnattu vetovoimien suunnasta riippuen. Soluja (lähinnä fibroblasteja) on vähän, ja ne sijaitsevat kuitukimppujen välissä. Tästä kudoksesta rakennettiin ligaatioita, jänteitä, fasciaeja, intermuskulaarisia septoja, periosteumia, perichondriumia ja muita (kuva 9).

Jänteissä ja nivelsiteissä kollageenikuitukimput sijaitsevat yhdensuuntaisesti, fasciassa, aponeuroosissa, intermuskulaarisissa septoissa - kerroksissa toistensa yläpuolella (mitä paksumpi fascia, sitä enemmän kerroksia), ja kuitujen suunta eri kerroksissa on erilainen: joissakin - suorassa kulmassa, toisissa - terävä, mikä antaa näille muodostelmille erityisen lujuuden. Jos elastiset kuidut ovat vallitsevia tiheässä kuituisessa sidekudoksessa, niin sitä kutsutaan elastiseksi sidekudokseksi. Joustavien kuitujen läsnäolo auttaa elintä tai kehon osaa palaamaan alkuperäiseen asentoonsa muodonmuutoksen jälkeen.
rusto (rusto) fysikaalis-kemiallisissa ominaisuuksissa ja toiminnallisissa piirteissä on hyvin erilainen kuin muun tyyppiset sidekudokset. Sen solujen välinen aine on melko tiheä, minkä vuoksi se suorittaa pääasiassa tuki- ja suojaustoiminnot (mekaaniset). Rustoa on kolme tyyppiä: hyaliini tai lasimainen, kollageenikuituinen ja elastinen. Rustikudoksessa ei ole verisuonia. Aineenvaihdunta (ravitsemus ja rappeutumistuotteiden poistaminen) suoritetaan sidekudoksen kalvon suonien läpi, jotka peittävät ruston ulkopuolella (perikondrium). Perikondriumin suonista peräisin olevat ravinteet tunkeutuvat ruston solujen väliseen aineeseen. Rustossa, joka kattaa luiden nivelpinnat, ravintoaineet tulevat nivelonteloa täyttävästä nivelnesteestä tai lähellä olevista luuastioista. Perikondriumin takia tapahtuu myös rustojen kasvua.
Hyaliinirusto on suurin levinneisyys ihmiskehossa. Solunvälinen aine on läpikuultava, sinertävänvalkoinen. Rustosolut sijaitsevat erityisissä onteloissa, joita ympäröi kapseli, joka on tiheämpi kuin solujen välinen aine. Hyaliinirusto muodostaa kylkiluiden etupäät, henkitorven rustot, keuhkoputket, suurimman osan kurkunpään rustasta ja peittää luiden nivelpinnat. Alkionjaksolla merkittävä osa luurankoa koostuu hyaliinirustosta. Vanhuudessa kalkki voi kerääntyä hyaliinirustoon (kuva 10).

Kollageeni kuitu rusto vähemmän joustava, mutta kestävämpi. Sen solujen välinen aine sisältää suuren määrän kollageenikuitujen kimppuja, jotka sijaitsevat suunnilleen samansuuntaisesti. Solut sijaitsevat kuitukimppujen välissä. Tästä rustosta rakennetaan nikamalevyt, rusto, joka yhdistää häpyluut (kuva 11).
Joustava rusto vähemmän kestävä, mutta erittäin joustava, kalkkipitoisuus ei tapahdu siinä koskaan. Ruston solujenvälisessä aineessa on paljon joustavia kuituja, jotka kietoutuvat keskenään muodostaen tiheän verkon. Sen solut muistuttavat kynttilän liekin muotoa ja sijaitsevat 2-3 kapselissa kuitujen välissä. Elastinen rusto sijaitsee siellä, missä ei tarvita suurta vastustusta aktiivisille voimille. Äänenkorkeus, epiglottis, ulkoisen kuulokanavan seinä ja kuuloputki on rakennettu siitä (kuva 12).

Luukudos on tihein kaikista sidekudostyypeistä. Sen solujen välinen aine koostuu kuiduista, jotka liittyvät usein kimppuihin, ja pääaineesta, jossa suuri osa epäorgaanisista yhdisteistä, pääasiassa kalsiumsuoloista, luun tukitoiminto on siten selvin. Tiheydestä huolimatta luukudos on elävä järjestelmä, mutta se muuttuu koko ihmisen elämän ajan, ja siihen liittyy päivittämällä sen rakenneosat, mikä varmistaa sen mukautuvuuden ympäristöolosuhteisiin (kuva 13).

Luukudoksen uusinta riippuu iästä, ravinnosta, sisäisen erityksen elinten toiminnasta ja muista tekijöistä. Luukudoksen muutokset ilmeisimmin tapahtuvat lihaksen toiminnan aikana: Luukudoksen sisäinen rakenne ei muutu, vaan myös elinten muoto - ne muodostavat luut.
Luukudoksessa erotellaan kolmen tyyppiset soluelementit: osteosyytit, osteoblastit ja osteoklastit.
osteosyyttien (oss - luu, sytos - solu) - luukudoksen pääsolu - on epäsäännöllinen muoto, suuri joukko pitkiä prosesseja, joiden avulla se on kosketuksessa naapurisoluihin. Nämä luusolut sijaitsevat erityisissä onteloissa.
osteoblastit - luojat, luukudoksen luojat. Ne sijaitsevat luunmuodostumisprosessissa. Niiden muoto voi olla kuutio, pyramidi tai kulma. Luukudoksen muodostuessa osteoblastit muuttuvat osteosyyteiksi.
osteoklastien - monisydämelliset solut. Ne ovat suurempia kuin osteosyytit ja osteoblastit. Jokaisella osteoklastilla voi olla jopa 50 ydintä. Paikkaan, jossa osteoklasti joutuu kosketuksiin luuaineen kanssa, muodostuu pieni vaikutelma. Tällaisissa vaikutelmissa lahtia ja osteoklastit valehtelevat. Nämä solut tuhoavat luukudoksen, jonka tilalle muodostuu uusi. Luukudoksessa tapahtuu jatkuvasti molempia prosesseja - sekä tuhoamisprosessia että luomisprosessia, jotka varmistavat luun jälleenrakentamisen.
Luukudoksia on kahta tyyppiä: karkea kuitu ja hieno kuitu tai lamellikuitu.
Karkea luu Se löytyy suuremmassa määrin alkiosta, aikuisilla sitä löytyy vain lihaksen jänteiden kiinnityspaikoista luihin, kallon luiden välisissä nivelissä. Karkean kuituisen luukudoksen solujenvälisessä aineessa kuituniput ovat paksuja, järjestettyinä yhdensuuntaisesti, kulmassa tai verkon muodossa. Osteosyyteillä on litistynyt muoto.
Hieno kuitutai lamellinen, luukudos kaikkein voimakkaasti eriytetty. Sen rakenteellinen ja toiminnallinen yksikkö on luulevy. Solunvälisessä aineessa kuitulevyt ovat ohuita, suuntautuneita tiettyihin suuntiin toistensa suuntaisesti. Osteosyytit sijaitsevat levyjen välissä tai levyjen sisällä. Levyt on sijoitettu siten, että kahden vierekkäisen levyn kuidut kulkevat melkein suorassa kulmassa, mikä antaa luukudokselle erityisen lujuuden ja joustavuuden. Lähes kaikki aikuisen luurangan luut on rakennettu hienokuituisesta luukudoksesta.
Olemme jo kirjoittaneet ST: n perustermeistä ja yleisistä komponenteista edellisessä artikkelissa sidekudoksen ominaisuuksista. Kuvailkaamme nyt yksilöä sidekudosryhmät (PT).
Loose ST - Tämä on pää- ja pääkudos, kun kyse on sidekudoksesta (kuva 10). Elastiset (1), kollageeni- (2) kuidut sekä jotkut solut sisältyvät sen amorfiseen komponenttiin. Tyypillisin solu on fibroblast (lat. Fibrakuitu, kreikkalainen blastosolu tai itu). Fibroblast pystyy syntetisoimaan amorfisen komponentin aineosat ja muodostamaan kuituja. Toisin sanoen solun - fibroblastin - todellinen tehtävä on kyky syntetisoida solujen välistä ainetta. Fibroblastit (3), joissa on suuri ydin (a) endoplasmassa (b) ja ektoplasma (c), sisältävät melko vaikuttavan endoplasmisen retikulumin, ne syntetisoivat proteiineja, kuten kollageenia ja elastiinia. Nämä proteiinit ovat vastaavien kuitujen rakentajat. Toinen tärkeä löysän CT: n solu on histiosyytti (4). Mikro-organismien tulisi olla varovaisia \u200b\u200bnäiden solujen suhteen, koska solujenväliseen aineeseen pääsy fagosoi ne tai yksinkertaisesti syö niitä. Lopuksi värikuvassa I voidaan nähdä vielä yksi tärkeä löysän CT: n solu - se on syöttösolu, joka varastoi kaksi biologisesti aktiivista yhdistettä: hepariinia ja histamiinia. Hepariini on aine, joka estää veren hyytymistä. Histamiini on aine, joka osallistuu erilaisiin allergisiin reaktioihin ja tulehduksellisiin prosesseihin. Histamiinin vapautumisen vuoksi syöttösoluista havaitaan oireita, kuten ihon punoitus, urtikaria, kutina, rakkuloiden muodostuminen, palaminen ja anafylaktisen sokin kehittyminen. 
Kuva I. Löysä sidekudos
Löysä CT tapahtuu kaikkien alusten mukana. Aorttaa ympäröi kokonainen tyyny - adventitia, ja pienimpiä kapillaareja ympäröi erittäin ohut kuitu- ja soluverkko. Alukset ovat suojattuja, vahvistettuja ja tukeutuvat tämän tyyppisiin ST: iin. Ja tämä tarkoittaa, että löysä CT sijaitsee missä tahansa suonessa. Tästä syystä on syytä tuoda esiin pää- ja pää sidekudoksena.
Käytännöllinen lääkäri kohtaa päivittäisessä työssään hyvin usein yhden löysän sidekudoksen oireen - turvotuksen. Amorfisen komponentin muodostavat glykosaminoglykaanit kykenevät pitämään vettä itsessään, mitä he tekevät milloin tahansa. Ja tämä mahdollisuus ilmenee joissain patologisissa prosesseissa: sydämen vajaatoiminta, imusolmukkeet, munuaissairaudet, tulehdukset ja niin edelleen. Tässä tapauksessa neste kerääntyy sidekudokseen, joka turpoaa, jolloin iho turpoaa. Joskus silmien alla oleva turvotus voi olla alkuperäinen oire sairaudelle, kuten glomerulonefriitille - munuaisen immuunitulehdukselle. ![]()
Tiheä ST Se sisältää hyvin pienen määrän solukomponentteja ja solujen välisen aineen amorfisen komponentin, suurin osa tiheästä sidekudoksesta on kuitua. Kaksi tiheän ST: n muotoa erotetaan toisistaan. Tiukasti muotoilematon ST (Kuvio 11) on täydellinen kuituhäiriö (4). Sen kuidut on kudottu niin kuin he haluavat; fibroblastit (5) voidaan suunnata mihin tahansa suuntaan. Tämäntyyppinen CT osallistuu ihon muodostumiseen, se sijaitsee orvaskeden (1) ja löysän CT-kerroksen (2) alla, joka ympäröi suonia (3), ja antaa iholle tietyn lujuuden. Mutta tässä hän ei vertaa kestävyyteen tiheä koristeltu st(Kuva 12), joka koostuu tiukasti järjestetyistä kimpuista (5), joissa puolestaan \u200b\u200bon tietty kollageeni- (3) ja / tai elastisten (4) kuitujen suunta. Suunniteltu sidekudos on osa jänteistä, nivelsiteistä, silmämunan valkomisesta kalvosta, fasciasta, dura materista, aponeuroosista ja joistain muista anatomisista muodostelmista. Kuidut kääritään (1) ja "kerrostetaan" (7) löysällä CT: llä, joka sisältää astiat (2) ja muut elementit (6). Jännekuitujen samansuuntaisuuden vuoksi ne saavat myös niiden suuren lujuuden ja jäykkyyden. 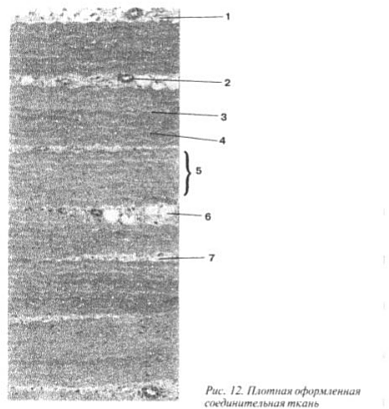
Rasvakudos (Kuva 13) on levinnyt melkein kaikkialle ihoon, retroperitoneaaliseen tilaan, varteen ja mesenteriaan. Rasvakudossoluja kutsutaan liposyyteiksi (1 ja kuva II). Ne sijaitsevat erittäin tiheästi, kulkevat vain pienet verisuonet, kuten kapillaarit (2), ja niiden mukana kaikkialla olevat fibroblastit, joilla on yksittäiset kuidut (3). Liposyyteistä puuttuu melkein kokonaan sytoplasmaa ja ne täytetään suurilla jatkuvilla rasvapisareilla. Ydin siirtyi sivulle, huolimatta siitä, että se on solun säätelijä. 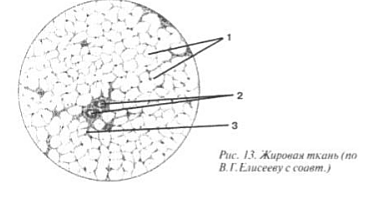

Kuva II. Rasvakudos
Rasvakudos on tärkeä kehon energialähde. Loppujen lopuksi, kun se hajottaa rasvaa, se vapautuu paljon enemmän kuin käytettäessä hiilihydraatteja ja proteiineja. Lisäksi tässä tapauksessa muodostuu huomattava määrä vettä, siksi rasvakudos osoittautuu samanaikaisesti sitoutuneen veden varastosäiliöksi (ei ole varmaa, että tämä ST-variantti sijaitsee kamelinkohoissa, jotka hajoavat rasvaa hitaasti ylittäessään kuumien aavikkojen läpi). On vielä yksi toiminto. Vastasyntyneillä ihossa havaittiin erityinen alalaji - ruskea rasvakudos. Se sisältää valtavan määrän mitokondrioita ja tästä syystä se on syntyneen vauvan tärkein lämmönlähde. 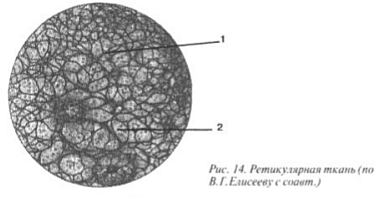
Verisuonikudos, sijaitsevat imusysteemin elimissä: punasoluissa, imusolmukkeissa, kateenkorva (kateenkorva), perna, koostuu moniprosessisoluista, joita kutsutaan retikulosyyteiksi. Latinalainen sana reticulum tarkoittaa "verkkoa", joka sopii täydellisesti tähän kudokseen (kuva 14). Retikulosyytit, kuten fibroblastit, syntetisoivat kuituja (1), joita kutsutaan retikulaarisiksi kuiduiksi (variantti kollageenista). Tämäntyyppinen CT tarjoaa hematopoieesin, ts. Melkein kaikki verisolut (2) kehittyvät eräänlaisena riippumatossa, joka koostuu verkkokudos (kuva III). 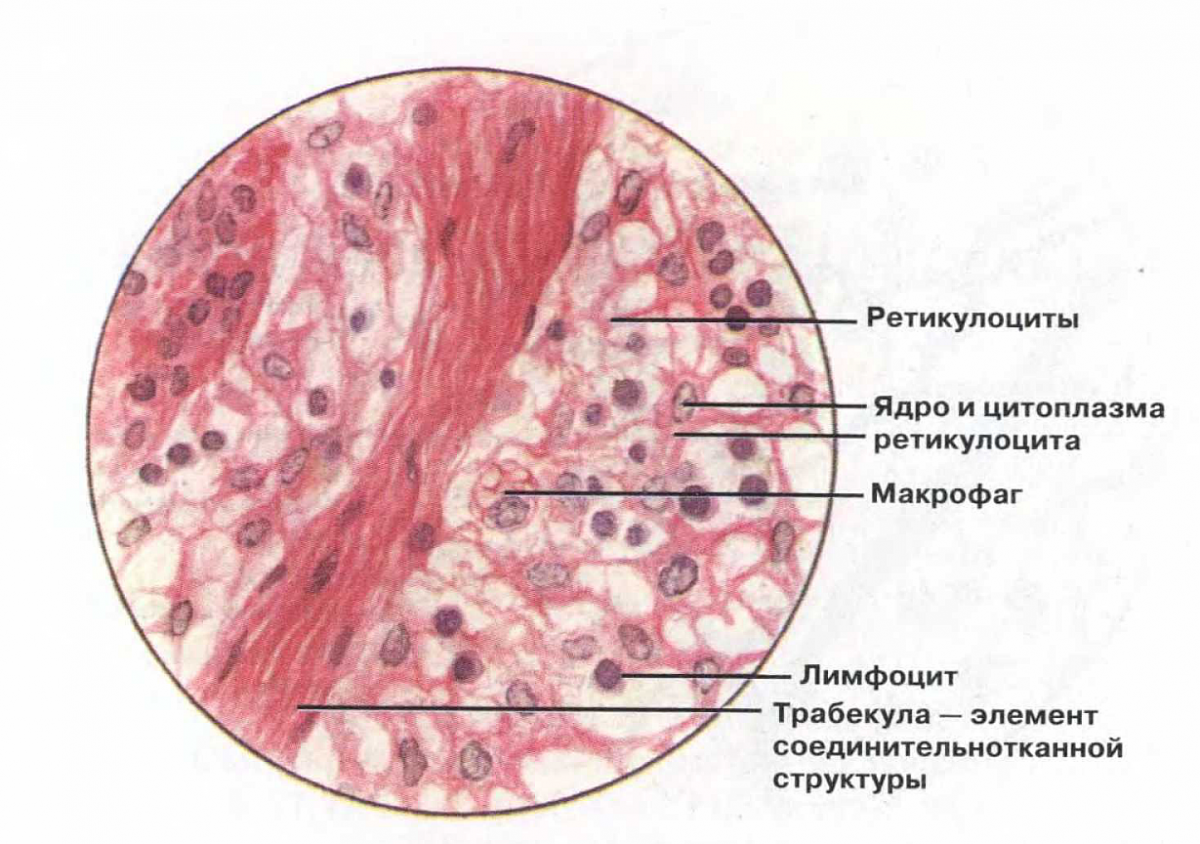
Kuva III. Verisuonikudos
ST: n viimeinen alalaji on pigmenttikudos (Kuva 15) löytyy melkein kaikesta, mikä on voimakkaasti värjätty. Esimerkkejä ovat hiukset, silmämunan verkkokalvo ja ruskettunut iho. Pigmentti kangas edustavat melanosyytit, solut, jotka on täytetty tärkeimmän eläinpigmentin - melaniinin rakeilla (1). Ne ovat tähtimuotoisia: keskellä sijaitsevasta ytimestä sytoplasma poikkeaa terälehdistä (2). 
Nämä solut voivat aiheuttaa pahanlaatuisen kasvaimen - melanooman. Taudista on viime aikoina tullut paljon yleisempää kuin ennen. Viime vuosikymmenen aikana ihosyövän esiintyvyys on lisääntynyt dramaattisesti. Uskotaan, että tämä johtuu otsonikerroksen paksuuden muutoksesta, joka suojaa planeettamme voimakkaalla kerroksella ultraviolettisäteilyn tappavalta vaikutukselta. Napojen yläpuolella se laski 40–60%, tutkijat puhuvat jopa otsonireikistä. Seurauksena ihmisissä, jotka paistavat auringon alla, syntymämerkkien melanosyytit ovat ensimmäiset, jotka reagoivat ultraviolettisäteiden mutageeniseen vaikutukseen. Jakaumatta keskeytyksettä, ne aiheuttavat kasvaimen kasvua. Valitettavasti melanooma etenee nopeasti ja etäpesäkkeet tulevat yleensä varhaisessa vaiheessa.
rusto (Kuva 16) - kudos, jonka solujenvälisessä aineessa on erittäin ”kiinteä”, väkevä, amorfinen komponentti. Glykosamiini ja proteoglykaanit tekevät siitä tiheän, joustavan, kuten hyytelö. Tällä kertaa sekä solujen välisen aineen amorfisia että kuitukomponentteja ei syntetisoida fibroblasteilla, vaan rustokudoksen nuorilla soluilla, joita kutsutaan kondroblasteiksi (2). Rustolla ei ole verisuonia. Sen ravitsemus tulee pintakerroksen kapillaareista - perikondriumista (1), jossa kondroblastit tosiasiallisesti sijaitsevat. Vain "kypsyneet" ne peitetään erityisellä kapselilla (5) ja kulkeutuvat itse ruston (3) amorfiseen aineeseen, jonka jälkeen niitä kutsutaan kondrosyyteiksi (4). Lisäksi solujenvälinen aine on niin tiheä, että jakautuessaan konrosyyttiä (6) sen tytärsolut eivät voi dispergoitua, pysyen yhdessä pienissä onteloissa (7). 
Rustokudos muodostaa kolmen tyyppisiä rustoja. Ensimmäisessä, hyaliinirustossa, on hyvin vähän kuituja siinä, ja se löytyy kylkiluiden ja rintalastan liitoskohdasta, henkitorven, keuhkoputkien ja kurkunpään luiden nivelpinnoilla. Toinen tyyppi rusto on elastinen (kuva IV), joka sisältää monia kimmoisia kuituja ja sijaitsee keuhkossa ja kurkunpään alueella. Kuitumainen rusto, jossa kollageenikuitut sijaitsevat pääasiassa, muodostaa häpyluun ja nikamalevyt. 
Kuva IV. Joustava rusto
Luukudos kantaa kolmen tyyppisiä soluja. Nuoret osteoblastit ovat toiminnaltaan samanlaisia \u200b\u200bkuin fibro- ja kondroblastit. Ne muodostavat luun solujen välisen aineen, joka sijaitsee pintakerroksessa, jossa on runsaasti verisuonia - periosteum. Ikääntyvät, osteoblastit sisältyvät itse luun koostumukseen, josta tulee osteosyyttejä. Alkionjakson aikana ihmiskeholla ei ole luita sellaisenaan. Alkiossa on sellaisenaan rustolevyjä, tulevien luiden malleja. Mutta vähitellen luutuminen alkaa, mikä vaatii ruston tuhoamista ja todellisen luukudoksen muodostumista. Tuhoajia tässä ovat solut - osteoklastit. Ne murskavat rustoa, mikä tilaa tilaa osteoblasteille ja heidän työlleen. Muuten, ikääntyvä luu korvataan jatkuvasti uudella, ja jälleen osteoklastit osallistuvat vanhan luun tuhoamiseen.
Luukudoksen solujen välinen aine sisältää pienen määrän orgaanisia aineita (30%), erityisesti kollageenikuituja, jotka ovat tiukasti suuntautuneita luun tiiviiseen aineeseen (kuva V) ja ovat sotkuisia. Amorfinen komponentti, joka "ymmärtää" olevan "tarpeeton tässä elämän juhlassa", puuttuu käytännössä. Sen sijaan täällä sijaitsee erilaisia \u200b\u200bepäorgaanisia suoloja, sitraatteja, hydroksiapatiittikiteitä, yli 30 hivenainetta. Jos poltat luun tulessa, niin kaikki kollageeni palaa; muoto kuitenkin tallennetaan, mutta kosketa vain sormea \u200b\u200bja luu murenee. Yön jälkeen jonkin hapon liuoksessa, jossa kaikki epäorgaaniset suolat liukenevat, luu voidaan leikata veitsellä kuin voita, ts. Se menettää lujuutensa, mutta se sidotaan kaulan ympärille (jäljellä olevien kuitujen ansiosta) edelläkävijänä. 
Kuva V. Luukudos
Viimeisenä mutta ei vähäisimpänä sidekudosryhmäon verta. Sen tutkiminen vaatii valtavan määrän tietoa. Siksi emme vähennä veren merkitystä kuvauksella tässä, vaan jätä tämä aihe erilliseen tarkasteluun.
Sidekudokset ovat laajalle levinneitä kudoksia, joilla on mesenkymaalinen alkuperä. Ne varmistavat muiden kudosten eheyden ylläpitämisen, muodostavat elinten stroman, sisältävät veri- ja imusolmukkeet ja osallistuvat kaikkien kudosten ja elinten troofiseen tukeen. Sidekudoksista erotetaan kuitukudokset (löysät ja tiheät) ja kudokset, joilla on erityisiä ominaisuuksia (esimerkiksi rasvaiset, retikulaariset). Eri sidekudokset eroavat fysikaalisista ominaisuuksistaan. Suurimmassa osassa elimiä ja niiden välillä löysä sidekudos toimii parenhyymin pakkauksena, järjestäen sängyn sen histologisille elementeille (elin stroma). Monilla solutyypeillä, elimillä ja kudoksilla sidekudosympäristö ei ole pelkästään passiivisesti tukeva rooli, mutta sillä on myös organisoiva ja informatiivinen merkitys. Tiheät sidekudokset antavat voimaa iholle, muodostavat elinkapseleita ja kestävät merkittäviä mekaanisia rasituksia (esimerkiksi nivelsiteet ja jänteet). Sidekudoksella on tärkeä metabolinen arvo paitsi siksi, että se sisältää verisuonia. Joten valkoinen rasvakudos varastoi rasvaa energianlähteenä ja ruskea rasvakudos osallistuu vastasyntyneiden ruumiinlämmön säätelyyn. Sarja sidekudossoluja
kuuluu suojaamiseksi patogeenisiä mikro-organismeja vastaan. Sidekudos on ponnahduslauta tulehduksellisille ja immuunivasteille.
ULKOISMATERIAALI
Kaikkien kuituisten sidekudosten koostumus sisältää merkittävän määrän solujen välistä ainetta. Ekstrasellulaarisen matriisin kemiallinen koostumus, rakenne ja tilavuus määrittävät kunkin sidekudostyypin fysikaaliset ominaisuudet. Solunulkoinen matriisi koostuu pääaineesta ja erityyppisistä kuiduista, jotka on upotettu siihen.
Pääaine
Pääaine on amorfinen materiaali, jolla on geeliominaisuudet. Kudosneste sitoutuu pääaineen komponentteihin muodostaen väliaineen molekyylien kulkemiseen sidekudoksen läpi ja veren aineenvaihdunnalle. Pääaine sisältää glykosaminoglykaaneja, proteoglykaaneja ja glykoproteiineja. Perusaineen molekyylit ovat sitoutuneet tiukasti solunulkoisiin matriisikuituihin ja ovat vuorovaikutuksessa eri solujen kanssa.
glykosaminoglykaanit
Glykosaminoglykaanit ovat polysakkaridit, jotka on rakennettu toistuvista disakkaridiyksiköistä, joista yksi on yleensä uronihappo ja toinen on aminosokeri (N-asetyyliglukosamiini, N-asetyyligalaktoosamiini). Glykosaminoglykaanit ovat happamia (negatiivisesti varautuneita) yhdisteitä niiden disakkaridiyksiköissä on monia hydroksyyli-, karboksyyli- ja sulfaattiryhmiä. Glykosaminoglykaanimolekyylit ovat hydrofiilisiä, sitovat suuren määrän vesimolekyylejä ja ioneja (erityisesti Na +), jotka tulevat solunulkoiseen nesteeseen. Glykosaminoglykaanien suuret molekyylit, jotka ovat sitoutuneet toisiinsa, muodostavat geelin, jonka kautta metaboliitit diffundoituvat. Seuraavat glykosaminoglykaanivalikoimat tunnetaan: hyaluronihappo, kondroitiinisulfaatti, dermataanisulfaatti, kerataanisulfaatti, heparaanisulfaatti ja hepariini (heparinisulfaatti).
proteoglykaanien
Proteoglykaani on kuituinen keskusproteiini, johon glykosaminoglykaanit kiinnittyvät kovalenttisesti. Siten muodostetaan suuria makromolekyylejä, jotka sisältävät 90-95% hiilihydraatteja. Proteoglykaanit voivat sitoutua ei-kovalenttisesti hyaluronihapon ketjuihin muodostaen vielä suurempia komplekseja (esimerkiksi rustoproteoglykaanit).
glykoproteiinit
Glykoproteiinit koostuvat polypeptidiketjuista, jotka on kytketty haarautuneisiin polysakkaridiin ja sitoutuvat solut solunulkoiseen matriisiin. On glykoproteiineja, jotka muodostavat kuiturakenteita (fibronektiini ja fibrilliini), samoin kuin useita ei-kuituproteiineja (laminiini, tenastsiini ja entaktiini).
fibronektiinilöytyy kellarimembraanista, kollageenikuitujen ja solujen ympärillä. Veriplasmasta löytyy suuri määrä fibronektiiniä. Fibronektiini osallistuu solujen tarttumiseen sitoutumalla ne solunulkoisen matriisin komponentteihin, erityisesti kollageeniin ja glykosaminoglykaaniin. Alkiogeneesissä ja haavojen paranemisessa fibronektiini muodostaa polut solujen kulkeutumiseen. Fibronektiinireseptori on integriini, joka on rakennettu solukalvoon. Solun sisällä integriini on vuorovaikutuksessa membraanin lähellä olevan sytoskeletonin aktiinimikrokuitujen kanssa ja yhdistyy ulkoisesti fibronektiinin kanssa. Fibronektiini puolestaan \u200b\u200bmuodostaa sidoksia kollageenin ja glykosaminoglykaanien kanssa. Tämä vahvistaa rakenteellisen jatkuvuuden sytoskeleton ja solunulkoisen matriksin välillä (kuva. 6-18 ).
fibrilliini(М r ≈350kD) muodostaa mikrofibrillejä, joiden halkaisija on 8–12 μm ja jotka vahvistavat soluja solunulkoisten komponenttien välillä; Se on osa joustavia kuituja, mikä tarjoaa niiden järjestetyn rakenteen. Fibrilliinigeenimutaatiot johtavat oireyhtymän kehittymiseen Marfan.
Erityyppiset kuidut upotetaan solunulkoiseen matriisiin: kollageeni- ja retikuliinikuitut sekä kollageenista rakennetut elastiset kuidut (koostuvat elastiinista ja fibrilliinistä).
Kollageenikuitu
Kollageenikuidut ovat useimpien sidekudosten pääkomponentti, ja kollageeni on runsasproteiini.
Kollageeneja.Kollageenimolekyyli - spiraali, jossa on kolme pro-a-ketjua. Tällaisen spiraalin pituus on 300 nm ja halkaisija on 1,5 nm. Kaikentyyppiset kollageenit sisältävät alueita, joissa toistuva sekvenssi on kolme aminohappoa glysiinin ollessa kolmannessa asemassa. Ensimmäinen aminohappo tässä sekvenssissä voi olla mikä tahansa, toinen - proliini, hydroksiproliini tai lysiini. Lysiinitähteiden välisten ristisidosten runsauden vuoksi kollageenikuiduilla on korkea lujuus. Ainakin 13 tyyppiä kollageenia tunnetaan (välilehti. 6-2 ).
Kuva 6-18.Fibronektiinin välittämä solujen assosiaatio kollageenikuitujen kanssa. Integriinin sytoplasminen domeeni on vuorovaikutuksessa lähellä kalvoa olevien mikrohiutaleiden (aktiini) kanssa ja solunulkoinen domeeni on vuorovaikutuksessa fibronektiinin kanssa, joka puolestaan \u200b\u200bliittyy kollageenikuituihin. Fibronektiinimolekyylissä on myös domeeneja sitoutumiseksi hepariiniin ja fibriiniin.
Ensimmäisten viiden tyypin kollageeni on yleisintä, loput ovat suhteellisen harvinaisia \u200b\u200bja pieninä määrinä. Kollageeni tyypit I, II ja III muodostavat fibrillejä jaksollisuudella 65 nm. Tyypin I kollageeni on yksi yleisimmistä, sitä syntetisoivat fibroblastit, osteoblastit ja GMC: t. Tyypin II kollageenia syntetisoivat kondroblastit, kondrosyytit, verkkokalvon hermosolut, ja sitä on läsnä lasiassa. Tyypin III kollageeni on osa retikuliinikuituja, jotka muodostavat herkän tukiverkoston joissakin elimissä ja kudoksissa (esimerkiksi maksassa, luuytimessä, imusoluelimissä), ja sitä syntetisoivat fibroblastit ja MMC: t. Tyypin IV kollageeni muodostaa kellarimembraanien retikulaarisen perustan ja syntetisoidaan useimmissa epiteelisoluissa. Tyypin V kollageenia on verisuonissa, GMC: n ympärillä ja luumatriksissa; sitä syntetisoivat MMC ja osteoblastit. Kollageenigeenimutaatiot johtavat useiden sairauksien (epätäydellinen osteogeneesi, akondrogeneesi ja akondroplasia, oireyhtymät) kehitykseen Ehlers - Danlo).
Kollageenikuitumuodostus(Fig. 6-19 ). Rakeisessa endoplasmisessa retikulumissa solunsisäisessä vaiheessa tapahtuu pro-a-ketjujen synteesi ja kokoonpano. Edelleen kompleksin säiliöissä Golginpolypeptidien hydroksylointi ja glykosylaatio suoritetaan, minkä seurauksena spiraalit muodostuvat kolmesta pro-a-ketjusta (prokollageeni). Jokainen pro-a-ketju sisältää terminaaliset alueet - rekisteröintipeptidit. Nämä peptidit estävät kollageenikollageenin solunsisäisen sitoutumisen kollageenifibrilliin. Kollageenimolekyylit kerääntyvät erittäviin rakeisiin ja tunkeutuvat solunulkoiseen tilaan. Solujen ulkopuolella prokollageenimolekyylin rekisteröintipeptidit pilkotaan käyttämällä prokollageenipeptidaasia (syntynyttä molekyyliä kutsutaan joskus tropokollageeniksi). Tätä pilkkoutumista ei tapahdu tyypin IV ja V-kollageenissa, mikä selittää tällaisten kollageenista rakennettujen fibrillien puuttumisen. Seuraavaksi kollageenifibrillit kootaan (kuva 5). 6-20 ). Troppokollageenimolekyyleillä on hännän ja pään päät ja ne yhdistyvät spontaanisti rinnakkaisiin ketjuihin, linjautuen peräkkäin: "päästä päähän" ja "sivulta toiselle". Samanaikaisesti pään ja hännän päiden väliin jää rakoja, joiden leveys on 35 nm.
Taulukko 6-2. Kollageenin jakautuminen kudoksiin ja elimiin
| tyyppi | Kudokset ja elimet |
| minä | Iho, jänteet, luut, sarveiskalvon, istukan, valtimoiden, maksan, dentiinin, kasvaimet |
| II | Rusto, nikamalevyt, lasimainen huumori, sarveiskalvo |
| III | Keuhkot, valtimoiden, kohtu, maksan retikuliinikuidut ja veren muodostavat elimet |
| IV | Kellarikerrokset |
| V | Istukka, iho, verisuonet, sileälihaskudos, rabdomyosarkooma |
| VI | Verisuonet, nivelsiteet, iho, kohtu, keuhkot, munuaiset |
| VII | Amnion, iho, ruokatorvi, sarveiskalvo |
| IX | |
| X | rusto |
| XI | Rusto, nikamalevyt, lasimainen |
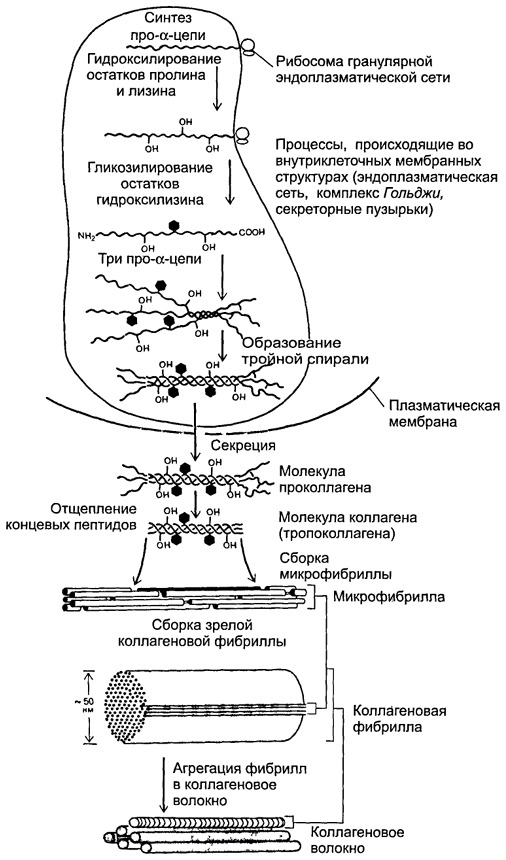
Kuva 6-19.Kollageenikuidun muodostuminen. Polypeptidi-a-ketjujen synteesi, niiden hydroksylaatio ja glykosylaatio, samoin kuin kolmen a-ketjun heeliksin muodostuminen tapahtuvat rakeisessa endoplasmisessa retikulumissa ja kompleksissa Golgin.Kollageenimolekyylit kerääntyvät eritysrakuloihin ja erittyvät solunulkoiseen tilaan. Solujen ulkopuolella terminaaliset peptidit pilkotaan polypeptideistä ja muodostuu kollageenifibrillejä.
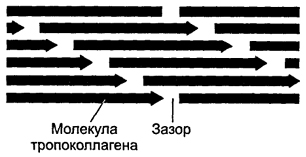
Kuva 6-20.Polarisoituneiden tropokolgeenimolekyylien pakkaaminen kollageenikuitun muodostumisen aikana. Yhdistettäessä peräkkäin molekyylejä ketjussa vierekkäisten molekyylien häntä ja päät välillä, 35 nm: n leveysrako säilyy. Yhden ketjun jokainen molekyyli siirtyy suhteessa viereisen ketjun molekyyliin neljänneksellä sen pituudesta. Tämä muodostaa kollageenifibrillin, jolla on poikittaisjuokaisu jaksolla 65 nm.
Jokainen ketjun tropokollageenimolekyyli on siirtynyt viereisen ketjun molekyyliin suhteessa neljäsosaa sen pituudesta, mikä on syynä tummien ja vaaleiden vyöhykkeiden vuorotteluun jaksolla 65 nm. Paksummat fibrillit muodostuvat lisäämällä uusia tropokollageenimolekyylejä. Kuparipitoinen entsyymi lysyylioksidaasi osallistuu ristisidosten muodostumiseen tropokollageenimolekyylien välille. Kollageenin vuorovaikutus glykosaminoglykaanien ja fibronektiinin kanssa säätelee fibrillien kokoamista. Kollageenikuitu muodostuu monista kollageenifibrilleistä, jotka on kytketty rinnakkain glykoproteiineilla. Kollageenikuitukimput - kokoelma kollageenikuituja upotettuna solujen väliseen matriisiin.
- C-vitamiini- yksi kofaktorista, joka tarvitaan ristisidosten muodostumiseen polypeptidiketjujen välille. C-vitamiinin puute johtaa heikosti hydroksyloituneiden pro-α-ketjujen muodostumiseen, jotka eivät pysty muodostamaan kolmoisheliilejä ja kollageenifibrillejä. Seurauksena on luiden, hampaiden ja verisuonten seinämien vikoja.
Kollageeni-soluvuorovaikutus(Taulukko. 6-3 ). Kollageenikuitut ympäröivät monia soluja. Joillakin soluilla (esimerkiksi hepatosyytit ja MMC) on kollageenireseptoreita ja ne sitoutuvat suoraan siihen. Muut solut sitoutuvat kollageeniin erityisillä välittäjänä sitovilla proteiineilla (useammin fibronektiini, harvemmin chondronektiini ja laminiini).
Joustavat kuidut
Joustavat rakenteet(kuiduilla ja kalvoilla) on selkeät elastiset ominaisuudet. Joustavia kuituja on joustavassa rustossa, iholla, keuhkoissa ja verisuonissa. Päättyneet joustavat kalvot ovat ominaisia \u200b\u200bsuurille valtimoille.
Taulukko 6-3. Sitoutuvat molekyylit kollageenin ja erityyppisten solujen välillä
Joustava kuitu(haarautuneen langan halkaisija 0,2–1,0 μm) koostuu fibrilliinistä ja amorfisesta elastiinista (kuva. 6-21 ). Glykoproteiinielastiini, kuten kollageeni, sisältää paljon glysiiniä ja proliinia. Fibroblastit ja MMC: t syntetisoivat elastiinia ja fibrilliiniä rakeisen endoplasmisen retikulumin säiliöissä. Monimutkaisessa Golginpolypeptidit pakataan
 |
| Kuva 6-21. Joustava kuitu. . Molekyylien välisten sidosten muodostumisen takia elastiinipolypeptidien välillä muodostuu elastinen molekyylien verkosto, joka palauttaa muodon muodonmuutoksen jälkeen. B. Fibrilliinimikrofibrillit, jotka organisoivat elastiinia, sijaitsevat sekä elastisen kuidun ulkopuolella että sen sisällä. |
Retikuliinikuidut
Retikuliinikuitut ovat ohuita filamentteja, joiden halkaisija on 0,5–2,0 μm ja jotka koostuvat tyypin III kollageenista, joka on sitoutunut glykoproteiineihin ja proteoglykaaniin. Tyypin III kollageenin α1-ketjua koodaavan geenin puutteiden kanssa retikuliinikuitujen muodostuminen on häiriintynyt, mikä johtaa repeämien esiintymiseen suoliston seinämässä ja / tai suurissa valtimoissa, joissa retikuliinikuidut muodostavat membraanin GMC: n ympärille (oireyhtymä) Elersa - Danlotyyppi IV).
Kellarikerros
Pohjakalvo on erityinen solunulkoisen matriisin tyyppi, sen levyn tai levyn muoto on 20–200 nm paksu (ks. Kuva. 16-2 ) ja koostuu erityisistä proteiineista, jotka toimivat erityyppisten solujen yhdistämiseksi
(epiteeli, lihakset, schwannin)ympäröivän sidekudoksen kanssa. Kellarikerros koostuu kahdesta kerroksesta - lamina lucida(vaalea kerros) 10-50 nm paksu plasmamembraanin vieressä, ja lamina densa(elektronitiheä kerros, paksuus vaihtelee). perusta lamina lucidaja lamina densa -kolmiulotteinen verkko, joka koostuu tyypin IV kollageenikuiduista, joihin laminiini, entaktiini ja heparaanisulfaatti ovat sitoutuneet.
Ankkurikuitut
Suurin osa kellarikalvoista on tiukasti kytketty löysään sidekudokseen. Ihon, ruokatorven, sarveiskalvon ja amnion monikerroksisen epiteelin kellarimembraanit on kytketty alla olevassa sidekudoksessa oleviin ankkurilevyihin lyhyillä ankkurikuiduilla. Ankkurikuitut koostuvat pääasiassa tyypin VII kollageenista, ankkurilevyt sisältävät tyypin IV kollageenia. Ankkurikuitut yhdistävät myös ankkurointilevyt stroomaan. Seurauksena muodostuu tiheä kudottujen kuitujen verkko, joka stabiloi pohjakalvon kudoksissa, joilla on merkittäviä vaikutuksia leikkaukseen.
Kellarimembraani yhdistää solut edistämällä niiden organisointia säiliössä; Se tukee solujen fenotyyppiä ja vaikuttaa erityyppisiin soluaktiivisuuksiin: tarttuvuus, erilaistuminen, polarisaatio, migraatio. Pohjakalvo estää epiteelisolujen tunkeutumisen alla olevaan sidekudokseen. Pahanlaatuisissa kasvaimissa tämän toiminnan rikkominen johtaa kasvainsolujen itämiseen ja etäpesäkkeiden muodostumiseen. Pohjakalvo suodattaa makromolekyylejä (esim. Munuaisissa) lamina densaviivyttää molekyylejä, joiden M r on suurempi kuin 50 kD).
Sidekudossolut ovat erilaisia. Näitä ovat fibroblastit, fibrosyytit, kondroblastit, kondroosyytit, osteoblastit, osteosyytit, makrofagit, syöttösolut, valkosolut, plasmosyytit, perisyytit, adiposyytit. Sidekudossolut jaetaan kahteen ryhmään - asukkaat ja maahanmuuttajat. Asukkaat - fibroblastit ja fibrosyytit (kuituinen sidekudos), kondroblastit ja kondrosyytit (rusto), osteoblastit ja osteosyytit (luukudos), syöttösolut, adiposyytit, makrofagit. Maahanmuuttajat - valkosolut (neutrofiilit, eosinofiilit, basofiilit, monosyytit, lymfosyytit). Kun tulehduspisteitä esiintyy, nämä solut poistuvat verenkierrosta sidekudokseen. Immunologisen seurannan lymfosyytit kiertävät jatkuvasti veren, sidekudosten ja imusolmukkeiden välillä.
Toiminnalliset ryhmät.Suoritetusta toiminnosta riippuen sidekudoksen solut voidaan jakaa kolmeen pääryhmään. Solut, jotka vastaavat solunulkoisen aineen molekyylien synteesistä ja ylläpitävät kudoksen rakenteellista eheyttä, ovat fibroblastit ja fibrosyytit, kondroblastit ja kondrosyytit, osteoblastit ja osteosyytit, odontoblastit, retikulaariset solut.
Rasvojen kertymisestä ja aineenvaihdunnasta vastaavat solut ovat rasvasolut; nämä solut muodostavat rasvakudoksen. Solut, joilla on suojaavia toimintoja (mukaan lukien immunologiset), ovat syöttösolut, makrofagit ja kaikenlaiset valkosolut.
Fibroblastit ja fibrosyytit
fibroblasti-
Fibroblast (kuva. 6-22 ks. myös kuva 3 insertissä) - yleisin sidekudossolujen tyyppi; salaa solunulkoisen matriisin komponentit, osallistuu haavan paranemiseen, kykenee lisääntymään ja muuttumaan. Fibroblast on litistynyt solu, jonka muoto on tähti, muodostaa laajat sphenoidiprosessit; sisältää suuren soikean ytimen, jossa on useita nukleoleja. Solun koko on muuttuva. Fibroblast syntetisoi intensiivisesti proteiinia, joten sen sytoplasma sisältää suuren määrän rakeisen endoplasmisen retikulumin säiliöitä, hyvin määritellyn kompleksin Golginmonia mitokondrioita. Siellä on lysosomeja ja eritysrakeita, glykogeenia, lukuisia mikrofilamentteja ja mikrotubuluksia.
Toiminto.Fibroblastit syntetisoivat kollageenit, elastiinin, fibronektiinin, glykosaminoglykaanit, proteoglykaanit ja muut solunulkoisen matriisin komponentit. Fibroblastit tuottavat erilaisia \u200b\u200bsytokiinejä - granulosyyttien ja makrofaagien siirtomaa stimuloiva tekijä (GM-CSF), granulosyyttien pesäkkeitä stimuloiva tekijä (G-CSF) ja makrofagien pesäkkeitä stimuloiva tekijä (M-CSF). Luuytimen fibroblastit erittävät ILS: n ja IL7: n. Tulehduksen ja haavojen parantumisen myötä kasvutekijät aktivoivat fibroblastit.
Kuva 6-22.Fibroblast ( ) ja fibrosyytit ( B). Fibroblast (aktiivinen solumuoto) sisältää tarkasti määritellyt organelit: rakeinen endoplasmainen reticulum, kompleksi Golginmitokondrioita. Fibroblast muodostaa suuria pitkänomaisia \u200b\u200bprosesseja. Fibrosyytissä organelit ovat paljon pienempiä, solulla ei ole prosesseja ja se on fusiform-muotoinen.
bFGF ja PDGF lisääntyvät ja muuttuvat vauriokohtaan. Fibroblastit sisältävät erilaisia \u200b\u200bentsyymejä, mukaan lukien kollagenaasit, jotka tuhoavat kollageenia. Tuhoamalla vaurioituneita ja syntetisoimalla uusia solunulkoisen matriisin makromolekyylejä fibroblasti edistää sen uudelleenjärjestelyä ja arpia vauriokohdassa (tulehdus).
fibroblasti-
Fibrosyytit (kuva. 6-22 ) On kypsä fibroblastimuoto, joka on tiheässä muodostuneessa sidekudoksessa. Fibrosyytti on fusiform-muotoinen. Tiivistetty ydin on pitkänomainen ja sijaitsee solua pitkin. Rakeisessa endoplasmisessa retikulumissa on hajallaan olevia säiliöitä, pieni määrä mitokondrioita. monimutkainen Golginhuonosti kehittynyt. Solu sisältää suhteellisen vähän eritysrakeita. Fibrosyytin tehtävänä on ylläpitää kudosrakennetta päivittämällä jatkuvasti (vaikkakin hitaasti) solunulkoisen matriisin komponentteja. Haavan paranemisella fibrosyytit voidaan stimuloida synteettiseen aktiivisuuteen. Aktivoituneella fibrosyytillä on fibroblastin piirteet: ydin on pyöristetty, endoplasmisen retikuluksen säiliöiden, mitokondrioiden lukumäärä kasvaa; monimutkainen Golgintulee entistä voimakkaammaksi.
Perikryptaaliset fibroblastit
Suoliston limakalvossa on perikriptaalisia fibroblasteja - stroomasoluja, joilla on MMC: n morfologisia piirteitä. Niiden uskotaan säätelevän epiteelisolujen kasvua ja erilaistumista. Nämä fibroblastit ilmentävät sileän lihaksen aktiinia.
myofibroblasteja
Myofibroblastit ovat supistuvia soluja, joilla on yhteisiä piirteitä GMC: n kanssa (ks. Luku 7 ).
makrofagit
Makrofagi on monosyyttien erilaistunut muoto. Makrofagit ovat ammatillisia fagosyyttejä, niitä on kaikissa kudoksissa ja elimissä. Tämä on erittäin liikkuva solupopulaatio, joka voi liikkua nopeasti. Elinajanodote on kuukautta. Kudosmakrofagit säilyttävät jonkin verran kykyä jakaa (esimerkiksi alveolaariset makrofagit kroonisissa tulehduksellisissa prosesseissa). Useiden makrofagien fuusion seurauksena tulehduksen painopisteessä muodostuu vieraiden kappaleiden monisydämen jättiläissoluja. Makrofaage jaetaan asuin- ja liikkuviin. Residentejä makrofageja on kudoksissa normaalisti ilman tulehdusta. Niistä erotellaan vapaiksi, pyöristetyiksi ja kiinteinä makrofageina - tähdenmuotoisiksi soluiksi, jotka kiinnittävät prosessinsa solunulkoiseen matriisiin tai muihin soluihin. Liikkuvat makrofagit ovat siirtävien (nimeltään) makrofagien populaatio.
Mononukleaaristen fagosyyttien järjestelmä.Sidekudoksen makrofagit ovat osa mononukleaarista fagosyyttijärjestelmää. Mononukleaarisen fagosyyttijärjestelmän solut eroavat muista fagosyyttisistä soluista kolmella tavalla:
| Kuva 6-23.Makrofagi. Ydin on muodoltaan epäsäännöllinen ja siinä on lovia. Sytoplasma sisältää ribosomeja, mitokondrioita, monisoluisia kappaleita ja hyvin kehittynyt kompleksi Golginja rakeinen endoplasminen reticulum. Lysosomeja on lukuisia. Fagosomien, fagolysosomien ja jäännöskappaleiden läsnäolo on ominaista. Solu muodostaa sytoplasmiset prosessit, jotka liittyvät muuttoliikkeeseen ja fagosytoosiin. |  |
joilla on makrofaagien morfologia, ovat peräisin luuytimestä, Ig: stä ja komplementtikomponentit moduloivat fagosyyttistä aktiivisuutta. Mononukleaarinen fagosyyttijärjestelmä sisältää histiosyytit (kudoksen makrofagit), alveolaariset makrofagit, osteoklastit, solut von Kupffer,solu Langerhansinsolu Hofbauer,vieraiden kappaleiden jättiläissolut ja todennäköisesti CNS-mikroglia-solut.
morfologia
Rakenne (kuva. 6-23 ) makrofagi riippuu sen aktiivisuudesta ja lokalisoinnista. Solun halkaisija on noin 20 mikronia. Ytimen muoto on epäsäännöllinen, ja siinä on syvennyksiä. Sytoplasmassa on mitokondrioita, vapaita ribosomeja, hyvin määritelty kompleksi Golginmonisoluiset elimet, rakeinen endoplasmainen retikulumi, lysosomit, fagolysosomit ja jäännöskappaleet, joiden aine voi vapautua makrofaagista eksosytoosin avulla. Bysterisidisiä aineita on läsnä lysosomeissa: myeloperoksidaasi, lysotsyymi, proteinaasit, happohydrolaasit, kationiset proteiinit, laktoferriini, superoksididismutaasi - entsyymi, joka edistää H 2 O 2: n, OH -, O - 2: n muodostumista. Plasmakalvon alla on suuria määriä aktiinimikrofilamentteja, mikrotubuluksia, välifilamentteja, jotka ovat välttämättömiä siirtymiselle ja fagosytoosille. Makrofaagit kulkeutuvat useiden aineiden pitoisuusgradienttia pitkin eri lähteistä. Aktivoidut makrofagit muodostavat epäsäännöllisen sytoplasmisen pseudopodian, joka liittyy amoeboidiliikkeeseen ja fagosytoosiin.
Makrofaagit ovat ammatillisia fagosyyttejä. Ne vangitsevat denaturoidut proteiinit verestä, ikääntyneistä punasoluista (maksan, pernan, luuytimen kiinteät makrofagit). Makrofaagit fagosytoivat solujätteet ja kudosmatriisin. Epäspesifinen fagosytoosi on ominaista alveolaarisille makrofageille, jotka vangitsevat erityyppisiä pölyhiukkasia, nokea jne. Spesifistä fagosytoosia tapahtuu, kun makrofagi on vuorovaikutuksessa opsonisoidun bakteerin kanssa. Aktivoidut makrofagit erittävät yli 60 tekijää. Makrofaageilla on antibakteerisia vaikutuksia
aktiivisuus, erittävä lysotsyymi, happamat hydrolaasit, kationiset proteiinit, laktoferriini, H202, OH -, 02 -. Kasvainten vastainen aktiivisuus on H202: n, arginaasin, sytolyyttisen proteinaasin, tuumorinekroositekijän (TNF) makrofagin suora sytotoksinen vaikutus. Makrofagi on antigeeniä esittelevä solu: se prosessoi Ag: tä ja esittää sen lymfosyyteille, mikä johtaa lymfosyyttien stimulaatioon ja laukaisemaan immuunivasteita. Makrofagi IL1 aktivoi T-lymfosyyttejä ja vähemmässä määrin B-lymfosyyttejä. Makrofagi tuottaa lipidivälittäjiä - prostaglandiini PGE 2: ta ja leukotrieenejä, verihiutaleiden aktivaatiokerrointa (PAF). Solu erittää myös a-IFN: ää, joka estää viruksen replikaation. Aktivoidut makrofagit erittävät entsyymejä, jotka tuhoavat solunulkoisen matriisin (elastaasi, hyaluronidaasi, kollagenaasi). Toisaalta makrofagien syntetisoimat kasvutekijät stimuloivat tehokkaasti epiteelisolujen lisääntymistä (transformoivat kasvutekijä TGFa, bFGF), fibroblasteiden lisääntymistä ja aktivointia (kasvutekijä PDGF-verihiutaleista), kollageenisynteesiä fibroblasteilla (transformoivat kasvutekijä TGFβ), uusien verisuonten muodostumista (fibroblastikasvutekijä bFGF). Siten makrofagien tuottamat kasvutekijät välittävät pääasiallisia haavan paranemisen taustalla olevia prosesseja (epiteelin muutos, solunulkoisen matriisin muodostuminen, vaurioituneiden suonien palauttaminen). Tuottamalla useita pesäkkeitä stimuloivia tekijöitä (makrofaagit - M-CSF, granulosyytit - G-CSF), makrofagit vaikuttavat verisolujen erilaistumiseen.
Rasvasolu
Mastosolut ovat morfologisesti ja toiminnallisesti samanlaisia \u200b\u200bkuin veren basofiilit, mutta ne ovat erilaisia \u200b\u200bsolutyyppejä. Mastosolu, kuten basofiili, tulee edeltäjästä luuytimessä, mutta lopullinen erilaistuminen tapahtuu sidekudoksessa. Mastosolut ovat sidekudoksen pysyviä soluja. Niitä on erityisen paljon iholla, hengityselinten ja ruuansulatuksessa olevien elinten limakalvossa, verisuonten ympärillä. Mastosolu (Fig. 6-24 ) sisältää lukuisia suuria metakromaattisia rakeita (modifioituja lysosomeja). Solukalvoon upotetaan erilaisia \u200b\u200breseptoreita, mukaan lukien reseptorit IgE Fc-fragmentille.
Mastosolut syntetisoivat ja keräävät rakeeseen erilaisia \u200b\u200bbiologisesti aktiivisia aineita, välittäjiä ja entsyymejä: hepariini (hepariinisulfaatti), histamiini, tryptaasi, kimaasi, elastaasi, dipeptidaasi, plasminogeeniaktivaattori, happohydrolaasit, eosinofiilien kemotaksisifaktori (ECF), kemotrofiatekijä NC . Syöttösolurakeiden pääkomponentti on negatiivisesti varautunut sulfatoitu glykosaminoglykaanihepariini, joka syntetisoidaan ja varastoidaan yksinomaan syöttösolujen kautta. Solun erittämä hepariini sitoo veressä kiertävää antitrombiini III: ta, tehostaen voimakkaasti sen antikoagulanttivaikutusta. Histamiini aiheuttaa MMC: n vähenemistä, liman hypersekreetiota, verisuonien läpäisevyyden lisääntymistä edeeman kehittyessä. Tryptaasi edistää fibrinogeenin hajoamista,
c3: n muuttuminen anafylatoksiini C3a: ksi, kollagenaasin aktivointi, fibronektiinin hajoaminen. Tryptaasi, kimaasi, karboksipeptidaasi B, muut proteaasit ja happohydrolaasit, jotka vapautuvat degranuloivasta solusta, aiheuttavat kudosmatriisin tuhoutumisen. Kun syöttösolu aktivoidaan (yhdessä rakeiden sisällön erittymisen kanssa), muodostuu arakidonihapon metaboliitteja - prostaglandiineja, tromboksaania TXA 2 ja leukotrieenejä. Näillä välittäjillä on vaso- ja bronhoaktiiviset ominaisuudet. Verihiutaleita aktivoiva tekijä (PAF), joka on yksi voimakkaimmista spasmogeeneistä, muodostuu myös kalvojen fosfolipideistä.
Syöttösolu on mukana tulehduksellisissa ja allergisissa reaktioissa. Ikäsolujen, kuten basofiilien, aktivoitumista ja degranulointia välittää IgE (katso kuva. 11-4 ). Mastosoluilla on korkean affiniteetin pintareseptoreja IgE Fc-fragmentteja kohtaan. Ag: n (allergeenin) sitoutumiseen IgE-molekyyliin mastosolun pinnalla liittyy rakeiden sisällön eksosytoosi, arakidonihapon metaboliittien muodostuminen.
Plasmasolu
Plasmasolut (Fig. 6-25 ) erottaa immunologisesti aktivoiduista B-lymfosyyteistä, syntetisoida ja erittää Ig (AT). Suuri pyöreä tai soikea ydin on eksentrinen ja sisältää dispergoitunutta kromatiinia. Sytoplasmassa hyvin kehittynyt kompleksi Golginja suurten rakeisten endoplasmisen retikulumin säiliöiden massa, joka on samankeskisesti sijoitettu ytimen ympärille.
perisyyttejä
Perisyytit ovat prosessisoluja, jotka sijaitsevat ulkopuolelta valtimoleihin, laskimoihin ja kapillaareihin. Yleisimmin kapillaarin jälkeisissä laskimoissa.
morfologia
Perisyytteillä on diskoidinen ydin, jolla on pienet sisennykset, ja ne sisältävät tavanomaisen joukon organelleja, monisoluisia kappaleita, mikrotubuluksia ja glykogeenia. Pidä suonen seinää vasten olevalla alueella kuplia. Supistuvia proteiineja on läsnä ytimen lähellä ja prosesseissa, mukaan lukien aktiini ja myosiini. Perisyytit peitetään kellarikalvolla, mutta liittyvät läheisesti toisiinsa
endoteelisolu, kuten niiden välinen kellarikalvo voi puuttua. Näissä paikoissa on tunnistettu ura- ja liimakoskettimet.
Perisyytit syntetisoivat kapillaarin pohjakalvon komponentit; kontrolloida endoteelisolujen lisääntymistä sekä normaalin verisuonikasvun että niiden regeneraation aikana; moduloi endoteelisolujen toimintaa säätelemällä makromolekyylien kuljetusta kapillaareista kudoksiin. Haavan paranemisen ja verisuonten palautumisen aikana perisyytit erottuvat MMC: ssä.
rasvasolut
Rasvasoluissa erotetaan valkoiset ja ruskeat rasvasolut.
Valkoinen rasvasolu
Erottelun aikana rasvapisarat ilmestyvät mesenkymaalisen solun sytoplasmaan, sulautuen niiden määrän kasvaessa. Erotettu rasvasolu on iso pyöristetty solu, jonka halkaisija on 120 μm, ja se sisältää yhden suuren pisaran rasvaa, työntäen sytoplasman ja kaikki organelit reuna-alueelle. Sytoplasmassa, kapeassa kehässä, joka ympäröi tippaa rasvaa, on litistetty ydin, vapaat ribosomit, sileä ja rakeinen endoplasminen reticulum, kompleksi Golginja mitokondriat. Adiposyytit erittävät leptiinihormonia, jonka vuorovaikutus hypotalamuksen ventromediaalisten ja lateraalisten ytimien hermosolujen reseptoreiden kanssa johtaa syömiskäyttäytymisen keskusmekanismien sisällyttämiseen; leptiini estää rasvakudoksen kertymistä.
Rasvojen synteesi ja varastointi.Adiposyytit syntetisoivat lipoproteiinilipaasia, joka diffundoituu endoteelisolujen pinnalle (pääasiassa rasvakudoksen kapillaareihin). Ruoan lipidit kuljetetaan rasvakudokseen osana VLDL: tä ja kyllomikronien muodossa. Lipoproteiinilipaasi hydrolysoi nämä lipidit rasvahapoiksi. Vapaat rasvahapot tunkeutuvat rasvasoluihin ja varastoidaan rasvapisaroihin triglyseridien muodossa. Rasvasolut kykenevät syntetisoimaan rasvahappoja.
myös glukoosista ja aminohapoista. Insuliini stimuloi lipoproteiinilipaasisynteesiä, rasvan laskeutumista ja glukoosin imeytymistä adiposyyteissä. Insuliini kuitenkin hidastaa rasvan mobilisoitumista rasvasoluista, estäen rasvaa hajottavien entsyymien aktiivisuuden. Varastoidut triglyseridit hydrolysoidaan hormoniherkällä cAMP-aktivoidulla lipaasilla. Tuloksena olevat vapaat rasvahapot pääsevät kapillaarin onteloon, missä ne sitoutuvat ei-kovalenttisesti albumiiniin ja kulkeutuvat maksaan.
Ruskea rasvasolu
Ruskea rasvasolu sisältää paljon pieniä rasvapisaroita ja suuria mitokondrioita. Ruskeassa adiposyytissä on hapettavien fosforylaatiofunktioiden dissosioitumisen luonnollinen mekanismi, joka on biologisesti käyttökelpoinen tapa lämmön tuottamiseksi. Solun ja kudoksen ruskea väri johtuu rautaa sisältävien pigmenttien läsnäolosta mitokondrioissa. Aktivoitu hormoniherkkä lipaasi hydrolysoi triglyseridejä rasvahapoiksi ja glyseroliksi. Vapautuneet rasvahapot metaboloituvat lämmön muodostamiseksi. Lisääntynyt lämmöntuotto selittyy termogeeniinikalvon läpäisevän proteiinin läsnäololla sisemmässä mitokondriaalisessa membraanissa. Termogeniini sallii aikaisemmin kalvojen väliseen tilaan kuljetettujen protonien käänteisen virtauksen kulkematta ATP-syntetaasijärjestelmän läpi. Siten protonivirran tuottamaa energiaa ei käytetä ATP: n synteesiin, vaan se häviää lämmön muodossa.
KUDEN KYTKENTÄTYYPIT
On kuituisia sidekudoksia ja sidekudoksia, joilla on erityisiä ominaisuuksia. Kuituinen sidekudos (kuitujen lukumäärän ja solunulkoisen matriisin pääaineen mukaan) jaetaan löysäksi ja tiheäksi. Tiheä sidekudos puolestaan \u200b\u200bjaetaan suunnitelluksi ja muotoilemattomaksi, mikä määräytyy kuitujen organisoitumisesta solunulkoiseen matriisiin. Yhdistäviin kudoksiin, joilla on erityisiä ominaisuuksia, ovat alkion (mesenkyymi), rasva ja retikulaariset. Näille kudoksille on ominaista samantyyppisten solujen hallitsevuus.
Kuituinen sidekudos
Löysä sidekudos
Löysä (muotoilematon) sidekudos (kuva. 6-26 ) sijaitsee kaikissa elimissä, muodostaa niiden strooman ja seuraa verisuonia. Tämä kangas sisältää suhteellisen vähän satunnaisesti jakautuneita kollageenia ja elastisia kuituja. Kuitujen välissä on suuri määrä pääainetta, johon on upotettu erilaisia \u200b\u200bsoluja: fibroblastit, muuttuvat ja pysyvät makrofagit, syöttösolut, perisyytit, adiposyytit, plasmasolut, valkosolut. Glikosaminoglykaanien molekyylit, jotka kietoutuvat toisiinsa, muodostavat verkon soluissa ja kanavissa, joiden sisällä on suuri määrä kudosnestettä, johon on liuennut siihen

Kuva 6-26Löysä sidekudos Solujen välinen aine koostuu pääasiassa pääaineesta, jossa on satunnaisesti jakautuneita kuituja, kollageenia ja joustavia. Solun monimuotoisuus on ominaista (fibroblastit, syöttösolut, erilaiset valkosolut, adiposyytit, makrofagit, plasmasolut)
aineita. Valkosolut, makrofagit, masto- ja plasmasolut osallistuvat aktiivisesti suojareaktioihin.
Tiheä sidekudos
Tiheä sidekudos sisältää suuren määrän tiukasti toisistaan \u200b\u200bsijaitsevia kuituja. Perusaineen määrä on suhteellisen pieni.
Paksu muotoilematon sidekudoskoostuu suuresta määrästä tiheästi mutta satunnaisesti järjestettyjä kuituja. Kuitujen välissä on fibroblasteja (fibrosyyttejä), makrofageja, syöttösoluja. Se on ominaista todelliselle iholle, periosteumille.
Tiheä kehystetty sidekudos.Kuidut on järjestetty tiheästi muodostaen yhdensuuntaisia \u200b\u200bkimppuja. Kuitujen välisissä kapeissa tiloissa fibrosyytit rinnastuvat ketjuihin. Tällaisesta kudoksesta muodostuu sidoksia, jänteitä ja kuitumembraaneja.
- joustava joukko(Esim ligamenta flavaja ligamentum nuchae) koostuu elastiinista, joka muodostaa paksuja kuituja. Niiden välissä on ohuita kollageenikuituja ja fibrosyyttejä.
- jänne(Fig. 6-27 ) koostuu kollageenikuiduista, jotka muodostavat jänteen niput I, II ja III luokkaan. Ensimmäisen asteen kimppujen välissä on rivi fibrosyyttejä (jännesolut), joissa on lamelliprosessit. Ensimmäisen kertaluokan nippujen ryhmät, joita ympäröi löysä sidekudos suonien ja hermojen kanssa, muodostavat toisen luokan niput. Löysä sidekudos yhdistää useita toisen kertaluokan kimppuja kolmannen luokan kimppuiksi. Kun jänne vaurioituu, aktivoidut fibrosyytit ja fibroblastit syntetisoivat kollageenia uusille kuiduille.
- Kuitukalvot- Eräänlainen tiheä, muodostunut sidekudos. Niiden välissä olevat kollageenikuitukimput ja fibrosyytit on järjestetty kerroksiin. Kussakin kerroksessa kollageenikuitujen aaltomaiset kimput kulkevat samansuuntaisesti samaan suuntaan, eri tavalla kuin vierekkäisten kerrosten suunnat. Erilliset kuitukimput kulkevat kerroksesta toiseen yhdistäen ne toisiinsa. Kuitumembraaneihin kuuluvat fascia, aponeurosis, kalvon jännekeskus, sisäelinten kapselit, dura mater, sklera.
Erityinen sidekudos
Yhdistäviin kudoksiin, joilla on erityisiä ominaisuuksia, ovat mesenkyymi, retikulaarinen ja rasvakudos.
mesenkyymasta
Sikiön sidekudos - mesenkyymi (kuva. 6-28 ) - kaikkien sidekudosten solujen lähde. Mesenkymaalisolut ovat tähti- tai karanmuotoisia, ja herkät haarautumisprosessit muodostavat verkon. Soikeassa ytimessä - dispergoitunut

Kuva 6-27jänne - poikkileikkaus B- pitkittäisleikkaus ja D- kaavio jänteen rakenteesta poikittais- ja pitkittäisleikkauksissa
kromatiini ja selvästi näkyvät nukleolit. Geelimäinen solunulkoinen materiaali koostuu melkein yksinomaan pääaineesta ja hajotettujen kuitujen (lähinnä retikuliinin) vähimmäismäärästä.
Verisuonikudos
Retikulaarinen kudos (kuva. 6-29 ) on retikulaarinen rakenne ja koostuu retikuliinikuiduista ja retikulaarisoluista, joilla on pitkät prosessit. Retikulaariset solut yhdessä retikuliinikuitujen kanssa muodostavat löysän verkon. Retikulaarinen kudos luo spesifisen mikroympäristön verisolujen kehittämiselle, lisäksi retikulaariset solut syntetisoivat erilaisia \u200b\u200bhematopoieettisia kasvutekijöitä (ILS, IL7, GM-CSF, G-CSF, M-CSF).
Rasvakudos
Rasvakudosta löytyy monista elimistä. Erota valkoinen ja ruskea rasvakudos.
Valkoinen rasvamuodostaa melkein koko kehon rasvakudoksen. Osallistuu veren imeytymiseen, neutraalien lipidien (triglyseridien) synteesiin, varastointiin ja mobilisointiin. Rasvakudoksen jakautumiseen kehossa vaikuttavat sukupuolihormonit ja lisämunuaisen kuoren hormonit. Rasvasolut (rasvasolut) muodostavat klustereita (lobules), erotettuna irtonaisen sidekudoksen väliseinillä. Jälkimmäisessä verisuonet ja hermot siirtyvät rasvakudokseen. Yksittäisiä rasvasoluja ympäröi retikuliini- ja kollageenikuituverkko. Sidekudosseptumissa esiintyy fibroblasteja ja syöttösoluja.
Ruskea rasvavastasyntyneessä se osallistuu termoregulaatioon. Aikuisella ruskeaa rasvaa löytyy pieninä määrinä välikarsinassa, aortan varrella ja ihon alla lapaluiden välissä. Ruskeaa rasvakudosta varustetaan runsaasti verikapillaareilla, jotka muodostavat verkon kunkin rasvasolun ympärille, ja sillä on selvä sympaattinen hengitys.