(meristemaattiset solut), ja niitä löytyy kasvien osista, joissa kasvu tapahtuu.
1. Meristeemien sytologiset piirteet
Erotetut kasvisolut eivät yleensä voi jakaa tai muuttua muun tyyppisiksi soluiksi. Siksi solujen jakautuminen meristeemeissä on välttämätöntä, jotta saadaan aikaan uusia soluja muiden kudosten kasvulle, uusien elinten muodostumiselle ja kasvien kehon rakenteen varmistamiseksi. Meristemaattisten solujen toiminta on samanlainen kuin niiden eläinten kantasoluilla, jotka eivät erota tai erota hiukan toisistaan \u200b\u200bja kykenevät jatkuvaan solujakautumiseen. Meristemaattiset solut ovat pieniä, ja sytoplasma ja ydin täyttävät solun täysin. Vacuulit ovat erittäin pieniä, ja sytoplasma ei sisällä erilaistuneita plastideja (kloroplastit tai kromoplastit), vaikka niitä onkin alkeellisessa muodossa (protoplastides). Meristemaattiset solut on tiiviisti pakattu, eikä melkein ole solunulkoista tilaa. Soluseinä on erittäin ohut primaarinen soluseinä.
Meristeemien fysiologinen aktiivisuus vaatii tasapainon kahden antagonistisen prosessin välillä: uusien kudosten muodostuminen ja tuki meristemaattisten solujen populaation uudistumiselle.
meristeemialtistuksen - muodostamalla kasvakudos, josta kaikki muut kudokset voidaan muodostaa, meristemaattiset solut säilyttävät pitkään kyvyn jakaa uusien erikoistumattomien solujen muodostumisen myötä.
Meristemisolut ovat litistettyjä, pieniä, tiheästi sijoitettuja vierekkäin, ilman interklinejä, ydin sijaitsee solujen keskellä, tyhjiöt ovat pieniä, lukuisia, solumembraani on primaarinen, on vain plastidi-esiasteita.
Rypälemeristeemien sytologiset piirteet ilmaistaan \u200b\u200btyypillisesti apikaalisissa meristeemeissä. Solut ovat isodiametrisiä monikulmioita, eivät erotettuja solujen välisiä tiloja. Soluseinät ovat ohuet, ja niissä on pieni selluloosapitoisuus. Sytoplasma on tiheä, ydin on suuri, sijaitsee keskellä. Sytoplasmassa on suuri ribosomien ja mitokodrioiden kalkisti (proteiineja ja muita aineita syntetisoidaan intensiivisesti). Lukuisat tyhjöt ovat hyvin pieniä.
Sivuttaisten meristeemien solut eivät ole kooltaan ja muodoltaan samanlaiset. Tämä johtuu niistä muodostuvien pysyvien kudosten solujen eroista. Joten esimerkiksi kambiumissa on parenkyymisiä ja prosenkymaalisia soluja. Parenymmaalisista alkukirjaista muodostuu johtavien kompleksien parenkyyma, prosenkymalista - varsinaisesti johtavista elementeistä.
Meristeemit luokitellaan asemansa mukaan tehtaalla:
Ja alkuperä:
- ensisijainen
- toissijainen.
Ensisijaiset meristeemit liittyvät läheisesti apikaaliseen meristemiin ja suoraan niistä, etenkin protodermia, päämeristeemiä ja prokabiumia muodostuu. Protodermi on yksikerroksinen heikosti erilaistuneiden solujen kerros, joka peittää kärjen ulkopuolelta, myöhemmin siitä muodostuu orvaskesi. Ydin muodostetaan päämeristeemistä ja proambiumista muodostetaan keskussylinteri, jonka solut ovat pitkänomaisia \u200b\u200bvarren akselia pitkin.
Toissijaiset meristeemit tunkeutuvat kypsien kasvien elinten erikoistuneisiin soluihin ja varmistavat niiden lateraalisen kasvun (felogeeni, perisykli, kambium). Ensisijaisesta meristeemistä muodostettuja kudoksia kutsutaan primaarisiksi ja sekundaarisiksi sekundaarisiksi. Toissijaisiin meristeemiin tulisi kuulua myös ns. Traumaattiset meristeemit, jotka muodostuvat kasvin rungon vauriopaikoissa ja tarjoavat uudistumisen.
Luettelossa lueteltujen meristeemityyppien joukossa apikaalinen meristeemi ilmenee ensin ontogeneesissä, josta tapahtuu kaikkien muiden kudosten erilaistuminen. Se sijaitsee varren ja juuren yläosissa (APEX) ja niiden sivuhaarat. Kasvi-organismin elintärkeän toiminnan piirre on erityisesti geneettisen aktiivisuuden säilyminen apikaalisissa meristeemeissä koko ontogeneesin ajan, siksi sanotaan, että kasvit kykenevät rajoittamattomaan kasvuun.
Apikaalisen meristemin rakenne (kasvupisteet)
1 - Keskiosa
2 - oheisosa
2. Apikaali meristeemi
4. Kalarienvälinen meristeemi
Intercalar meristem (synonyymi - insert meristem) - solut sijaitsevat erilaistuneiden kudosten alueiden välissä tarjoamalla lisätyn kasvun.
Esimerkiksi kalaarienvälinen meristeemi sijaitsee viljan solmukoissa, petioelien juuressa, heteissä. Nämä ovat jäännösprimaarisia meristeemejä. Ne ovat peräisin uskovista meristeemistä, mutta niiden muuntaminen pysyväksi kudokseksi viivästyy muihin kudokudoksiin verrattuna. Nämä herkät meristeemit ovat erityisen havaittavissa viljoissa. Leivän kunnon tapauksessa ne tarjoavat varren nostamisen, koska solut jakautuvat epätasaisesti olkojen ala- ja yläpuolelta.
5. Haava meristeemi
Muodostuu vaurioittaessaan kudoksia ja elimiä. Vaikuttavia alueita ympäröivät elävät solut muuttuvat erilaistuneiksi ja alkavat jakaa, eli ne muuttuvat toissijaiseksi meristeemiksi. Meristemien haava muodostuu Kalyus - tiheä, vaalean- tai kellertävä kudos, joka koostuu erikokoisista parenkyymisoluista, joita ei ole järjestetty. Kallussoluilla on suuret ytimet ja suhteellisen paksut soluseinät. Kalus voi aiheuttaa minkä tahansa kasvikudoksen tai -elimen. Reunalle on muodostettu tulppa, kallusolut voivat erottua muihin kudoksiin. Kalyusaan voidaan juurtua ylimääräisiä juuria ja silmuja. Kalyus esiintyy, kun kasvit ympätään, se antaa kasvikselle lehden ja kannan; pistokkaiden ytimessä. Kaliusa käytetään myös eristettyjen kudosviljelmien saamiseen.
Koulutuskangas koulutuskankaat
(meristeemit), kasvakudokset, jotka koostuvat soluista, jotka säilyttävät kykynsä jakaa pitkään. Näiden solujen jakautumisen vuoksi kasvit kasvavat koko elämänsä ajan (joissakin puissa se on satoja tai jopa tuhansia vuosia). Meristeemisolujen toinen tärkeä ominaisuus on, että ne synnyttävät erikoistuneita soluja, jotka muodostavat pysyviä kudoksia - yhtenäisiä, emäksisiä, johtavia, mekaanisia, erittyviä. Neljä meristeemityyppiä erotellaan riippuen syntyvän kasvin rungosta. Apikaaliset tai apikaaliset meristeemit aikaansaavat versojen ja juurten kasvun pituuksina. Sivusuuntaiset tai lateraaliset meristeemit aiheuttavat varten ja juurten kasvua paksuusina, ja niitä kutsutaan cambium. Välialueiden väliset, tai kalatalien väliset meristeemit varastoidaan väliaikaisesti varren sisäisissä osissa ja nuorten lehtien emäksissä varmistaen näiden alueiden kasvun, mutta muuttuvat sitten pysyviksi kudoksiksi. Haavoittuneita tai traumaattisia meristemejä syntyy kasvien vauriopaikoissa, joissa ne muodostavat suojaavan kovettuma. Kysymys on kuinka eri kudokset muodostuvat samoista meristemaattisista soluista, ts. miten solujen erilaistuminen tapahtuu, on edelleen ratkaisematta.
.(Lähde: "Biologia. Nykyaikainen kuvitettu tietosanakirja." Toimittanut A. P. Gorkin; M .: Rosman, 2006.)
Katso mitä "kasvatuskudos" on muissa sanakirjoissa:
KOULUTUSKANKAAT - katso meristemaattiset kudokset ... Kasvitieteellisten termien sanakirja
keskiasteen koulutuskudos - Synonyymit: sekundaariset meristeemit ovat kasvatuskudoksia, jotka syntyvät joko primaarisista meristeemeistä tai pysyvistä kudoksista. Edustettuina kambium-, pellogeeni- ja haavameristeemit ...
peruskoulutuskudos - Synonyymit: ensisijaiset meristeemit, meristeemien kasvatuskudokset, jotka ovat peräisin suoraan alkion meristeemistä; heidän soluillaan on aluksi kyky jakaa. P.: lle. t. sisältää ampun ja juuren apikaaliset meristeemit, Procambius, ... Kasvien anatomia ja morfologia
KASVIKKEET - soluryhmiä tai komplekseja, jotka liittyvät yhteiseen rakenteeseen, alkuperään, toimintaan ja sijaintiin. Tämän mukaisesti erotetaan toisistaan \u200b\u200bopetusaineet, yhdentyvät aineet, perus-, mekaaniset, johtavat ja erittävät aineet. Opetuskankaat tai ... Maatalouden tietosanakirja
kasvakudos - kasvien kudokset, soluryhmät tai -kompleksit, jotka liittyvät yhteiseen rakenteeseen, alkuperään, toimintaan ja sijaintiin. Tämän mukaisesti erottaa koulutus-, kokonaisuus-, perus-, mekaaniset, johtavat ja erittävät T. s. ... Maatalous. Suuri tietosanakirja
Solujärjestelmät, jotka ovat rakenteeltaan, alkuperältään ja toiminnaltaan samanlaisia, eroavat kooltaan, muodoltaan ja sijainniltaan. Kudoksen koostumus sisältää kudosnesteen (täyttää solujen väliset tilat) ja esimerkiksi solujen välissä olevat aineet. kalsiumsuolat ... Biologinen tietosanakirja
Malli apikaalisesta meristeemistä ”tunikatapauksesta”. Meristeemit, koulutuskudokset tai meristemaattiset kudokset (muu kreikkalainen ... Wikipedia
Tunikatapausmalli Meristems, tai kasvatuskudokset, tai meristemaattiset kudokset (kreikalla jaettavat meristot) kasvin kudosten yleisnimi, joka koostuu intensiivisesti jakamisesta ja fysiologisen toiminnan säilyttämisestä ... ... Wikipedia
Tunikatapausmalli Meristems, tai kasvatuskudokset, tai meristemaattiset kudokset (kreikalla jaettavat meristot) kasvin kudosten yleisnimi, joka koostuu intensiivisesti jakamisesta ja fysiologisen toiminnan säilyttämisestä ... ... Wikipedia
1) Kasvatuskudos tai meristemi
Kasvin runko muodostuu toiminnan seurauksena koulutus- kudoksia kutsutaan meristeemit. Meristeemien tärkein ominaisuus on solujen kyky jakaa mitoosilla alkuperäisten solujen läsnäolon takia.
Meristeemien koostumuksessa erota toisistaan: 1) nimikirjaimet, 2) nimikirjaimet. Alkuaineet säilyttävät kyvyn jakaa koko kasvin elinkaaren ajan ja pysyvät aina meristeemien koostumuksessa. Alkuaineiden johdannaiset jaetaan useita kertoja ja muuttuvat sitten pysyviksi kudoksiksi.
Ominaisuudet alkuperäinen: tiheä sytoplasma, suuri ydin, monia mitokondrioita, pieniä tai ei tyhjiöitä, ohut soluseinä.
Alkuperäinen: 1) ensisijainen meristeemialtistuksenjotka ovat peräisin suoraan alkion meristeemistä. Näistä primaariset pysyvät kudokset kehittyvät. 2) toissijainen meristeemialtistuksen, primaarisista tai muista kudoksista. Näistä kehittyy pysyviä sekundaarisia kudoksia.
Neljä meristeemityyppiä erotellaan sijainnista riippuen:
Apikaalinen ( apikaalinen) meristeemit- varren yläosissa ja juurten päissä antaa kasvun pitkäksi.
Sivuttaiset (lateraaliset) meristeemit -aksiaalielinten kehän ympärillä paksuuden kasvu varmistetaan. Ensisijaiset lateraaliset meristeemit ovat Procambius ja pericycle . Toissijaiset lateraaliset meristeemit: cambium- johtuu Procambius - ja korkkikambium (pellogeeni) - muodostettu pysyvien kudosten soluista. Johtavat kudokset muodostuvat procambiumista ja kambiumista ja korvan kambiumista peridermi.
Lisäysmerkinnät (intercalary)ovat versojen ja nuorten lehtien internodeissa. Ne ovat ensisijaisia, koska ne ovat apikaalisten meristeemien jäännöksiä. Lisäysmeristeemeillä ei ole alkukirjaimia kokoonpanossaan, ja ne lopulta muuttuvat pysyviksi kudoksiksi.
Haavan (traumaattiset) meristeemitmuodostuu yleensä kudosten ja elinten vaurioilla. Haava meristeemit muodostuvat kovettuma- erityinen kudos, joka koostuu homogeenisista parenhimaalisista soluista ja peittää vauriokohdan. Mikä tahansa kasvin kudos tai elin voi johtua kalluksesta.
Käytännöllinen käyttö: kukkasia käytetään hajusteissa, lääketieteessä, sisustuksessa.
2) Yksisirkkaisten kasvien mikroskooppinen rakenne:
A) Ensisijainen aivokuori ilmaistuna (lilja, iiris). Näissä tapauksissa varressa voidaan nähdä selkeä erilaistuminen ihoon, primaarikuoreen ja aksiaaliseen sylinteriin monikerroksisella perisykkelillä sklerenymyymirenkaan muodossa. Primaarinen aivokuori koostuu yleensä useista kerroista parenhimaalisia klorofylliä kantavia soluja. Kollenkyema puuttuu yleensä. Maanalaisissa versoissa primaarikuori ja keskisylinteri on rajattu erityisen jyrkästi; ensisijainen aivokuori on paljon leveämpi kuin samojen kasvien maahan varret, endodermissä esiintyy Caspari-renkaita tai jopa kuppimaisia \u200b\u200bsolumembraanien paksunemisia, ja perikkelia edustavat pääasiassa parenkyymisolut.
B) Muilla yksisirkkaisten luokan edustajien palmu-, vilja- ja muissa perheissä ensisijainen aivokuori on niin selvästi ei ilmaistu. Nisäkkeen alla olevassa maissivarressa on joskus klorofyllipitoisia parenhyymia, sklerenymia ympäröi näitä alueita ja vieressä suoraan niiden väliseen epidermiin. Perifeerisissä verisuonen kimppuissa on voimakkaat sklerenymialustat, ja toisinaan orvaskeden lähellä muodostetaan jatkuva sklerenymikerros, johon verisuonen kimpput upotetaan.
3) Korkeampien kasvien yleiset ominaisuudet
Korkeammat kasvit ovat ulkonäöltään, rakenteeltaan ja biologisilta ominaisuuksiltaan hyvin erilaisia. Niihin kuuluvat paitsi kukkivat kasvit, myös kuntosalinpermet, kuten saniaiset, kuorit, kruunut ja sammalit.
Korkeammille kasveille on ominaista joukko merkkejä korkeammasta organisaatiosta. Heidän ruumiinsa on jaettu varroihin ja lehtiin (toisin kuin alemmat). Tässä suhteessa niitä kutsutaan lehtivihaiset kasvit.
Monimutkaisen hajoamisen, elinten (juurin, varren, lehden) muodostuminen johtui kasvien siirtymisestä uuteen maaperäiseen elinympäristöön. Ilmaympäristölle on ominaista paljon korkeampi happipitoisuus kuin veteen. Maaperä - muilla mineraaliravinteiden ja erityisesti veden saannin olosuhteilla. Uudelle olosuhteisiin omaksumiseksi tarvittiin fotosynteettisen laitteen rakenneuudistus. Joten kasvi näytti varsi.
Lisäksi, paremman imeytymisen aikaansaamiseksi, juuset korvattiin juurakoilla. Juuret alkoivat tarjota mineraaliravinteita, ja lehdet - korkeampien kasvien ilmaravinteita.
Elinten normaalin toiminnan kannalta oli välttämätöntä siirtää vesi nopeasti lehtiin ja orgaaniset aineet - lehdestä muihin kasvin osiin. Tämä johti johtavan järjestelmän kehittämiseen erityisten solujen - trakeideiden, verisuonten ja seulaputkien - muodossa. Suojausta varten muodostettiin yhtenäinen kudos, mekaaninen.
Seksuaalinen prosessi on tullut vähemmän riippuvaiseksi vedestä.
Korkeampi: riniofytit, psilofyytit, sammalit, vesimuodot, pipari, saniaiset, kuntosospermit, angiospermit
Kasvien elimet koostuvat kudoksista. Kasvikudos on kokoelma soluja, joilla on samanlainen rakenne, homogeeninen alkuperä ja jotka suorittavat saman toiminnon. Maan korkeampien kasvien kudos saavutti erityisen kehityksen. Kasveissa kasvatuskudokset erotetaan toisistaan: integroituneita, mekaanisia, johtavia, perus- ja erittyviä.
Kasvatuskudokset eli meristeemit palvelevat kasvussa kasvua varten, ne ovat ensisijaista alkuperää ja toissijaista. Peruskoulutuskudos on juurien ja varren kasvun kartioissa. Pysykykäämme ensin juurikartio (Kuva 1, I).
Kuvio 1. Ensisijainen koulutuskudos juurikasvustossa:
Minä - juurikartio: 1 - kasvatuskudoksen solut; 2 - juurikannen solut. II - ytimen ja solun kompleksisen fission peräkkäiset vaiheet (karyokinesis tai mitosis): 1 - välivaihe; 2 - profaasi; 3 - metafaasi; 4-5 - anafaasi; 6-7 - telofaasi; 8 - sytokiineesi.
Juurikärjellä on kartiomainen muoto, peitetty juurikannella, jonka alla on ensisijainen kasvatuskudos. Koulutuskudoksen solut ovat tetraedrisiä tai monipuolisia, niissä on tiheä protoplasma, suuret ytimet ja ohuet selluloosamembraanit. Niiden tyhjiöt eivät ole kehittyneitä tai heikosti kehittyneitä. Näillä soluilla on kyky jakaa energisesti. Solunjakoa edeltää monimutkainen ydinjako, jossa tapahtuu ominaisia \u200b\u200bmuutoksia. Valmiissa, hyvin valmistetuissa ja värjätyissä valmisteissa voi nähdä ytimiä, jotka ovat monimutkaisen fission eri vaiheissa, joita kutsutaan muuten karyokineesiksi (“karyon” on ydin, “kinesis” on liike) (kuva 1, II). Lukuisat havainnot ovat osoittaneet, että ytimessä olevasta kromatiinista syntyy ensin ohuita kromatiiniffilamentteja, jotka lyhenevät, paksenevät ja muuttuvat erillisiksi partikkeleiksi - kromosomeiksi (“kromo” - maali, “monni” - runko), jotka ovat usein kaarevat hevosenkengän muodossa (fissioprofaasi). . Jokainen profaasissa oleva kromosomi jakaa pituussuunnassa kahteen puolikkaaseen, mutta tosiasiassa kromosomit kaksinkertaistuvat (pelkistys). Ydin ja ydinmembraani katoavat vähitellen. Kromosomit sijaitsevat ytimen keskiosassa yhdessä tasossa levyn muodossa. Kunkin kromosomin puolikkaat (“tytärkromosomit”) ovat tiiviisti vierekkäin, ja tässä vaiheessa (metafaasi) juurin poikittaisissa osissa kromosomien lukumäärä voidaan laskea helposti ja niiden muodon huomioon ottaminen. Ytimen napoissa (jopa ennen ydinkuoren katoamista) näkyy kahdessa korkissa ohut akromatiini (värjäämätön) filamenttijärjestelmä. Langat kasvavat ja ovat karan muodossa. Tämän jälkeen kromosomipuolisten (tytärkromosomien) jakautuminen solun kahteen vastakkaiseen napaan (anafaasi) alkaa. Kromosomien erottelu päättyy tytärkromosomeihin, jotka kerääntyvät solun napoihin toisiinsa, muuttuen kahdeksi sotkuksi (teofaasi). Samaan aikaan pektiinin sakeutumisia esiintyy akromatiiniffilamenteissa solun keskiosassa - muodostuu fragmoplasti, ts. Poikittaiskalvon perustila. Lopuksi, ytimiä ympäröi membraani, nukleoleja (yksi tai useampia) muodostuu niihin, ja protoplasma erotetaan poikittaisella kalvolla (sytokiineesi), minkä seurauksena syntyy kaksi uutta solua.
Ytimen ja solun jakamisprosessi tapahtuu useammin yöllä, sen nopeus riippuu lämpötilasta ja kosteudesta. Jakamisprosessi kestää keskimäärin 75-120 minuuttia, mutta joskus se kestää useita tunteja, ja suurin osa ajasta vietetään ensimmäiseen vaiheeseen (profaasi). Solunjako solunjakautumisen aikana tapahtuu protoplasman ja ytimen elintärkeän toiminnan tuloksena. Keskimmäinen levy näkyy aluksi. Se koostuu solujenvälisestä aineesta, joka tarttuu yhteen primaariselluloosakalvoihin, jotka kuuluvat kahteen soluun.
Ensimmäistä kertaa kromosomien hajaantuminen solun napoihin, samoin kuin muut ytimen ja solun hajoamisen yksityiskohdat, määritteli vuonna 1875 Moskovan yliopiston professori I. D. Chistyakov.
Ytimen ja solun kompleksisen fission, jota kutsutaan myös mitoosiksi, ydin on ilmeisesti siinä, että jakamalla kromosomit tasaisesti kahteen puolikkaaseen (tai tarkemmin sanoen kaksinkertaistamalla kromosomit) tytärisolut saavat äidiltä enemmän tai vähemmän yhtä suuret määrät kromatiinia (deoksiribonukleiini) happo - DNA), joka siirtää perinnöllisyyden emosolusta tytärsoluihin. ("Mitos" on säie. On yleisesti hyväksyttyä, että mitoosi \u003d karyokineesi + sytokiineesi). Kromosomien muoto ja lukumäärä ovat erityisiä piirteitä jokaiselle kasvilajeelle (pehmeä vehnä 2n \u003d 42 kromosomia, durumvehnä 2n \u003d 28, ohra 2n \u003d 14 jne.). Joskus kromosomien lukumäärä voi kuitenkin vaihdella, ja yhdessä organismissa voit tarkkailla erilaisia \u200b\u200bkromosomisoluja.
Mitoosin (tai karyokineesin) lisäksi kasveissa on suora ytimenjako (amitoosi) - yksinkertainen ytimen vetäminen ja joskus koko solu kahteen puolikkaaseen.
Mitoosin ohella ytimen jakautumisen väheneminen, meioosi, on yleinen kasveissa, joissa uusien solujen kromosomien lukumäärä puolittuu emosoluun verrattuna. Tämä jakautuminen tapahtuu kasveissa itiöiden muodostumisen aikana, jonka jälkeen uros- ja naissukusolut (sukusolut) syntyvät ja kehittyvät. Viimeksi mainittujen ytimessä on puolet kromosomien lukumäärästä, ja sellaisia \u200b\u200bytimiä kutsutaan haploideiksi toisin kuin vegetatiivisten solujen diploidiset ytimet, joista kasvien vegetatiiviset elimet koostuvat. Kasvien seksuaalinen prosessi on kahden sukusolujen (uros ja naaras) fuusio. Jos kromatiiniainetta ei vähentynyt, ts. Sen massa laski puoleen, niin jokaisessa uudessa seksuaalipolvessa tämä ainemassa ja siten kromosomien lukumäärä kasvaisi väistämättä. Sukusoluissa (sukusoluissa) ytimet ovat haploideja, ja vasta hedelmöitysprosessin jälkeen solun ytimistä tulee taas diploideja.
Tutkija V. I. Belyaev löysi pelkistysjakauman, ja tutkija S. G. Navashin löysi kaksinkertaisen hedelmöityksen kukinnan kasveissa. Hän on koko tutkijakoulun perustaja, joka on mukana tutkimassa kaikkia ydin- ja solujakautumisen yksityiskohtia.
Peruskoulutuskudos synnyttää muita kudoksia, koska sen solut muuttuvat, eriytyvät. Solujen kyky jakaa intensiivisesti kaksisirkkaisten kasvien varreissa ja juurissa säilyy koko elämän ajan meristemaattisissa soluissa, joita kutsutaan kambiumiksi, jolloin varsi ja juuri paksenevat. Keskiasteen koulutuskudoksiin kuuluvat ne, jotka syntyvät muista kudoksista. Esimerkki on falogeeni, joka johtuu usein taustalla olevasta kudoksesta.
Kudosta. KANKAIDEN LUOKITTELU.
Korkeampien kasvien organisointi perustuu solujen erikoistumisen periaatteeseen, joka koostuu siitä, että kehon jokainen solu ei suorita kaikkia luontaisia \u200b\u200btoimintojaan, vaan vain joitain, mutta täydellisemmin ja täydellisemmin.
kankaat- vakaa, säännöllisesti toistuva solukompleksi, alkuperältään, rakenteeltaan samanlainen ja mukautettu suorittamaan yksi tai useampi toiminto.
Kudoksia on erilaisia \u200b\u200bluokituksia, mutta ne ovat kaikki melko mielivaltaisia.
Päätoiminnosta riippuen erotellaan useita kasvien kudosryhmiä.
1. Koulutuskangas,tai meristemi- kyky jakaa ja muodostaa kaikki muut kudokset.
2. Peitekudokset:
ensisijainen;
toissijainen;
Kolmannen asteen.
3. Peruskankaat- muodostavat suurimman osan kasvin rungosta. Seuraavat pääkankaat erotetaan toisistaan:
Assimilaatio (klorofylliä sisältävä);
varastojen;
Ilmassa (aerenchyma);
Pohjavesiesiintymiin.
4. Mekaaniset kankaat(tuki, luuranko):
collenchyma;
Sclerenchyma.
5. Johtavat kankaat:
Xylem (puu) - ylävirran kangas;
Phloem (otsa) - kangas, jonka virta on alaspäin.
6. Erityskudos:
ulkoa:
Rauhaskarvat;
Hydatodes - vesistoma;
nectaries;
kotimainen:
Erityisöljyt, eteeriset öljyt, hartsit, tanniinit;
Monisoluiset eritteiden säilytysastiat, maito.
Solujen jakautumiskyvyn perusteella erotetaan kaksi kudostyyppiä: koulutus- jatai meristemija pysyvä- yhtenäinen, erittävä, perus, mekaaninen, johtava.
Kangas nimeltään yksinkertainen,jos kaikki sen solut ovat muodoltaan ja toiminnaltaan samanlaiset (parenkyyma, sklerenchyma, kollenkyema). monimutkainenkudokset koostuvat soluista, joiden muoto, rakenne ja toiminta eroavat toisistaan, mutta jotka liittyvät yhteiseen alkuperään (esimerkiksi ksylem, phloem).
Kudokset on myös luokiteltu niiden alkuperän perusteella (ontogeneettiset). Tämän luokituksen mukaan primaariset ja sekundaariset kudokset erotetaan toisistaan. Ensisijaisesta meristeemistä, joka sijaitsee ampumisen yläosassa ja juuren kärjessä, sekä siemen-itusta, muodostuu primaarivakiotkudos (orvaskesi, kollenkyema, sklerenyema, assimilaatiokudos, epiblema). Pysyvät kudossolut eivät kykene jakautumaan edelleen. Erikoistuneen meristeemin soluista - procambium - ovat muodostuneet ensisijaisesti johtavakudokset (primaarinen ksyleemi, primaarifloemi).
Toissijaisesta meristeemistä - cambium - ovat muodostuneet toissijainenkudokset: sekundaarinen ksyleemi, sekundäärifloemi; alkaen phellogen muodostuu korkki, fellodermi, linssit, joita esiintyy, kun varsi ja juuri paksenevat. Toissijaisia \u200b\u200bkudoksia esiintyy tyypillisesti kuntosoluissa ja kaksisirkkaisissa angiospermissä. Toissijaisten kudosten voimakas kehitys - puu ja hartio - ovat ominaisia \u200b\u200bpuumaisille kasveille.
KOULUTUSKANKAAT
Koulutuskangassolujen jatkuvan mitoottisen jakautumisen vuoksi ne varmistavat kaikkien kasvakudosten muodostumisen, ts. todella muodostavat hänen ruumiinsa. Mikä tahansa solu kehityksessään käy läpi kolme vaihetta: alkion, kasvu ja erilaistumisvaihe (ts. Solun hankkima tietty toiminto). Alkion erottuessa ensisijainen meristeemi pysyy vain tulevan verson kärjessä (kasvikartiossa) ja juuren kärjessä - apikaalinen (apikaalinen) meristeemialtistuksen.Minkä tahansa kasvin alkio koostuu meristeemisoluista.
Meristeemien sytologinen ominaisuus.Tyypilliset merkit korostuvat voimakkaimmin apikaalisissa meristeemeissä. Nämä meristeemit koostuvat isodiametrisistä monikerroksisista soluista, joita ei ole erotettu solujen välisillä tiloilla. Heidän kuoret ovat ohuet, sisältävät vähän selluloosaa ja pystyvät venymään.
Kunkin solun ontelo on täynnä tiheää sytoplasmaa, jolla on suhteellisen suuri ydin, joka on keskeisessä asemassa ja jakaa intensiivisesti mitoosin. Hyaloplasmassa on monia hajanaisesti siroteltuja ribosomeja, proplastideja, mitokondrioita ja dicososomeja. Vakuumia on vähän, ja ne ovat pieniä. Johtavat kudokset muodostetaan meristeemistä, jolla on prosentuaalinen muoto ja suuret tyhjiöt, kuten procambium ja kambium. Procambium -solut ovat poikkileikkaukseltaan monikulmaisia, kambiumsolut ovat suorakulmaisia.
Solut, jotka säilyttävät meristemaattiset ominaisuutensa, jatkavat jakautumistaan \u200b\u200bmuodostaen yhä useampia soluja, joita kutsutaan nimikirjaimet.Jotkut tytärsoluista erilaistuvat ja muuttuvat eri kudosten soluiksi, niitä kutsutaan nimikirjaimet.Alkusolut voivat jakaa määräämättömästi useita kertoja, ja nimikirjaimien johdannaiset jakautuvat yhden tai useamman kerran ja muuttua pysyviksi kudoksiksi.
Alkuperään ensisijainen ja toissijainen meristeemi erotetaan toisistaan.
Ensisijaiset meristeemit
Ensisijaiset meristeemit ovat peräisin suoraan alkion meristeemistä, ja niillä on kyky jakaa. Kasvien sijainnin mukaan ensisijaiset meristeemit voivat olla apikaalisia (apikaalisia), kalarienvälisiä (intercalary) ja lateraalisia (lateraalisia).
Apikaaliset meristeemit- sellaiset meristeemit, jotka sijaitsevat aikuisissa kasveissa varten varsilla ja juurten kärjillä ja varmistavat vartalon kasvun pituudessa. Kasvun kartion varreista erotetaan kaksi meristemaattista kerrosta: tunika, josta muodostuu yhtenäinen kudos ja primaarikuoren kehäosa, ja runko, josta muodostuu primaarikuoren sisäosa ja keskiaksiaalinen sylinteri (kuva 2.3).
Kuva 2.3.Apikaalinen varren meristeemit: ja- pitkittäisleikkaus: 1 - kasvikartio; 2 - lehden ensisija; 3 - aksillaarisen munuaisen tuberkle;
Juuren kärjessä erotetaan kolme kerrosta:
1) dermatogeeni, josta muodostuu primaarinen immuunikudos - rhizoderma;
2) ongelma, josta primaarikuoren kudokset kehittyvät;
3) keuhkoputki, joka muodostaa keskiaksiaalisen sylinterin kudoksen.
alkuperän mukaan ne voivat olla primaarisia ja toissijaisia; aksiaalielinten poikkileikkauksessa ne näyttävät renkailta. Esimerkki ensisijaisesta lateraalisesta meristeemistä on Procambius ja pericycle. alkaen procambiumkambium ja verisuonikuitumaisten kimppujen primaarielementit muodostuvat (primaarifloemi ja primaarinen ksyleemi), kun taas prokambiasolut erilaistuvat suoraan primaarisesti johtavien kudosten soluiksi.
Sivuttaiset meristeemit sijaitsevat yhdensuuntaisesti elimen pinnan kanssa ja aikaansaavat aksiaalisten elinten kasvun paksuudessa.
Lisäysmerkinnät (intercalary) useammin primaarisia ja varastoidaan erillisinä osioina aktiivisen kasvun alueilla kasvin eri osissa (esimerkiksi lehtilehtien kannassa, interoodien pohjalla). Viljassa olevien interoodien juuressa tämän meristeemin aktiivisuus johtaa interoodien pidentymiseen, mikä varmistaa varren kasvun pituudessa.
Toissijaiset meristeemit
Toissijaiseen sisältyy lateraalinen ja haavan meristeemi.
Sivuttaiset (lateraaliset) meristeemitesittelee cambiumja fellogenom.Ne muodostuvat promeriasisista (procambium) tai pysyvistä kudoksista erittämällä. Kambiumsolut jaetaan osioilla, jotka ovat yhdensuuntaiset elimen pinnan kanssa (perikliinisesti). Kambiumin ulospäin sijoittamista soluista kehittyy sekundaarifloemin elementtejä ja soluista, jotka ovat talletettu sisäänpäin, sekundaarinen ksylemi. Kambiumia, jota syntyy pysyvästä kudoksesta erilaistumisen kautta, kutsutaan dobavochnm.Rakenteeltaan ja toiminnaltaan se ei eroa kambiumista, joka syntyi promeriemistä. Fellogeeni muodostuu pysyvistä kudoksista, jotka sijaitsevat subepidermaalisissa kerroksissa (orvaskeden alla). Jakamalla perikliinisesti fellogeeni erottaa putken solut (laskusoma) ulospäin ja phallodermisolujen sisäpuolelta. Siten falogeeni muodostaa sekundaarisen yhtenäisen kudoksen - peridermin. Sivuttaiset meristeemit sijaitsevat yhdensuuntaisesti elimen pinnan kanssa ja aikaansaavat aksiaalisten elinten kasvun paksuudessa.
Haava meristeemitmuodostuu kudosten ja elinten vaurioilla. Vaurion ympärillä elävät solut erottuvat, alkavat jakautua ja muuttua siten sekundaariseksi meristeemiksi. Heidän tehtävänään on muodostaa tiheä suojakudos, joka koostuu parenyymisoluista, - känsä.Tämä kudos on vaaleaa tai kellertävää, sen soluissa on suuret ytimet ja melko paksut soluseinät. Kallus esiintyy rokotettaessa, mikä varmistaa lehmän kasvun kannan kanssa ja pistokkaiden juuressa. Siihen voidaan asettaa kliinisiä juuria ja munuaisia, joten sitä käytetään eristettyjen kudosten viljelyyn.
KANSI
Ensisijainen kudos
K ensisijainen kansikudokset kantavat epidermaaliset oikeat, periostikaaliset, takana olevat stomata-solut ja trikomit.
Soluun saapuva pektiini ja selluloosa voivat olla limittyneet muodostumiseen limaja kumit.Ne ovat polymeerihiilihydraatteja, jotka liittyvät sukupetiiniaineisiin, ja ne erottuvat kyvyltään turvota voimakkaasti kosketuksessa veden kanssa. Turvoksissa oleva kumi on tahmeaa ja sitä voidaan vetää lankoihin, kun taas lima on hyvin epäselvää eikä sitä voida vetää lankoihin. Pektiinilima löytyy lilja-, ristiinnauhe-, vaalea-, linden-, vaaleanpunaisten perheiden perheistä, toisin kuin selluloosalima, joka on paljon harvemmin esiintyvä (esimerkiksi orkideamilmassa).
 stomata
ovat erittäin erikoistuneita orvaskeden muodostelmia, jotka koostuvat kahdesta papun muotoisesta sulkemissolusta ja vatsan halkeamasta (eräänlainen solujen välinen tila niiden välillä). Niitä esiintyy pääasiassa lehtiä, mutta myös varsi (kuva 2.6).
stomata
ovat erittäin erikoistuneita orvaskeden muodostelmia, jotka koostuvat kahdesta papun muotoisesta sulkemissolusta ja vatsan halkeamasta (eräänlainen solujen välinen tila niiden välillä). Niitä esiintyy pääasiassa lehtiä, mutta myös varsi (kuva 2.6).
Kuva 2.6.Stomata-rakenne: a, b- timjamilehden kuori (ylhäältä päin ja poikkileikkaus); sisään- kuori viljan varresta (kaktusperhe); 1 - oikeastaan \u200b\u200bepidermaaliset solut; 2 - takana olevat stomata-solut; 3 - taudin halkeama; 4 - ilmaontelo; 5 - klorofyylipitoiset parenhyymisolut; A - kynsinauha; B - kutinakerros - kuori suberiinilla ja vahalla; B - seinämän selluloosakerros; G - ydin, jossa on nukleoli; D - klooriplastit
Takaosan solut paksenevät epätasaisesti: halkeaman (vatsan) suuntaiset seinät paksenevat merkittävästi verrattuna halkeamasta (vatsan) suuntautuneisiin seiniin. Rako voi kasvaa ja supistua säätelemällä verenkiertoa ja kaasunvaihtoa. Raon alla on suuri hengitysontelo (solujen välinen tila), jota ympäröivät lehden mesofyllisolut.
 Jäljellä olevia soluja ympäröivät parotid-solut, jotka muodostavat yhdessä vatsakompleksi(Kuva 2.7). Seuraavat tyypilliset vatsakompleksit erotetaan toisistaan:
Jäljellä olevia soluja ympäröivät parotid-solut, jotka muodostavat yhdessä vatsakompleksi(Kuva 2.7). Seuraavat tyypilliset vatsakompleksit erotetaan toisistaan:
Kuva 2.7.Vatsakalustotyyppien päätyypit: 1 - anomosyyttinen (kaikissa korkeissa kasveissa, paitsi pipar); 2 - diasyytti (saniaisissa ja kukinnan aikana); 3 - paracytic (saniaisissa, kukkaroissa, kukinnassa ja tukahduttavassa); 4 - anisosyytti (vain kukinnan aikana); 5 - tetrasyytti (pääosin yksisirkaisissa); 6 - syklisyyttinen (saniaisissa, kuntosoluissa ja kukinnassa)
1) anomotsitny(virheellinen) - sulkevilla soluilla ei ole äänekäs parostolisia soluja; kaikille korkeille kasveille ominaisia, havupuita lukuun ottamatta;
2) anizotsitny(ei-soluväli) - stomaattien takana olevat solut ympäröivät kolme parotid-solua, joista yksi on paljon suurempi (tai pienempi) kuin muut;
3) paracytic(rinnakkaissolu) - yksi parotidisolu (tai useampi) on samansuuntainen sulkemisen kanssa;
4) diatsitny(ristisolu) - kaksi parotid-solua sijaitsevat kohtisuorassa sulkemiseen nähden;
5) tetratsitny(Kreikan kielestä tetra- neljä) - pääosin yksisirkkaisissa;
 Vatsat sijaitsevat lehden alapuolella, mutta kelluvilla lehdillä varustetuissa vesikasveissa ne ovat vain lehden yläosassa. Lehden epidermaalisten solujen muodon ja stomaattien sijainnin perusteella yksisirkkaiset kasvit kaksisirkkaisista voidaan erottaa (kuva 2.8). Kaksisirkkaisten kasvien varsinaiset epidermaaliset lehtisolut ovat muodoltaan aaltoilevia (kuva 2.9), kun taas yksisirkkaisissa ne ovat pitkänomaisia, rombimuotoisia.
Vatsat sijaitsevat lehden alapuolella, mutta kelluvilla lehdillä varustetuissa vesikasveissa ne ovat vain lehden yläosassa. Lehden epidermaalisten solujen muodon ja stomaattien sijainnin perusteella yksisirkkaiset kasvit kaksisirkkaisista voidaan erottaa (kuva 2.8). Kaksisirkkaisten kasvien varsinaiset epidermaaliset lehtisolut ovat muodoltaan aaltoilevia (kuva 2.9), kun taas yksisirkkaisissa ne ovat pitkänomaisia, rombimuotoisia.
Kuva 2.8.Stomanan sijainti orvaskentässä (näkymä pinnalta): ja- kaksisirkkaiset kasvit:1 - alkukirjain; 2 - vesimeloni; b- yksisirkkaiset kasvit:3 - maissi; 4 - iiris
Stomatatyypit sijainnin suhteen orvaskeden pintaan voidaan jakaa seuraavasti.
1.7.1. Stomata sijaitsee samassa tasossa kuin orvaskesi. Yleisin tyyppi, jota ei yleensä ilmoiteta lääkekasvien materiaalien mikroskopian kuvauksessa, ts. tämä kohta on jätetty pois. Diagnostiset merkit ovat joko ulkonevia tai upotettuja vatsakipuja.
1.7.2. Ulkoneva stomata - hamakuoren yläpuolella sijaitseva stomata. Yleensä, kun mikroskooppista mikroruuvia kierretään (kun linssi lasketaan), tällaiset stomaatit havaitaan ensin, ja sitten ilmaantuvat epidermaaliset solut, minkä vuoksi on lähes mahdotonta kaapata niitä arkin pinnalta valokuvassa ja kuvata niitä kuvassa. Samassa tasossa orvaskeden kanssa tällainen stomata voidaan nähdä poikittaisissa osissa, mutta tätä varten osan on kuljettava vatsan läpi, jota on vaikea saada aikaan, kun ne sijaitsevat harvoin levyllä. Tällaiset oireet ovat tyypillisiä esimerkiksi karhunlehden lehdille.
1.7.3. Upotettu Stomata - orvakehälle upotettu stomata. Kun epidermaaliset solut havaitaan mikroskoopin alla samalla, kun pyöritetään mikroruuvia (kun linssiä lasketaan), epidermaaliset solut havaitaan ensin selvästi, minkä jälkeen on mahdollista nähdä selkeämmin stomaatan muodot. Valokuvia ja piirroksia huumausaineista pinnalta, niitä on myös vaikea näyttää. Löydetty mäyrälehdet, katsella lehdet, eukalyptus lehdet. Joskus syvennykset, joissa stomata sijaitsevat, ovat vuorattuja tai peitettynä karvilla ja niitä kutsutaan vatsakryptat.
1.8. Stomatalisolujen tyypit
Kirjallisuus kuvaa 19 tyyppiä. Valitsimme vain sellaisia, joita käytetään lääkekasvien materiaalien analyysissä **.

Kuva 63.Stomatalisolujen tyypit. A - lento; B - pallomainen; B - korkin muotoinen; D - scaphoid
1.8.1. linssinmuotoisia - 2 identtistä kuun muotoista solua sijaitsevat symmetrisesti. Etutasossa kuoren paksuus on melkein tasainen. Kiinnitysväli (kuva 63, A). Stomatal solutyyppi on ominainen useimmille kasveille.
1.8.2. Sferovidnye - kaksi identtistä, voimakkaasti pyöreästi kaarevaa kennoa sijaitsevat symmetrisesti. Etutasossa kuoren paksuus on melkein tasainen. Rako on pyöreä (kuva 63, B).
1.8.3. Kolpachkovidnye - kahdella identtisellä kuun muotoisella solulla polaariosissa on sakeutumia korkin muodossa. Kiinnitysväli (kuva 63, B). Niitä löytyy digitalisista.
1.8.4. navicular - Stomataalisten solujen sisäseinämät ovat sakeutuneet. Sulakerako (kuva 63, D). Se havaitaan satamissa ruoho, kellon lehdet.
Vatsan mekanismi johtuu solujen osmoottisista ominaisuuksista. Kun arkin pinta valaistaan \u200b\u200bsulkevien solujen kloroplastissa aurinkoa, tapahtuu aktiivinen fotosynteesiprosessi. Solujen kyllästyminen fotosynteesituotteilla, sokereilla merkitsee aktiivista kaliumionien pääsyä soluihin, minkä seurauksena solumehun konsentraatio sulkevissa soluissa kasvaa. Stomataalisten ja jäljessä olevien solujen solupitoisuuden pitoisuuksissa on ero. Solujen osmoottisten ominaisuuksien takia, vedessä vatsasoluista tulee suljin, mikä johtaa jälkimmäisen tilavuuden lisääntymiseen ja turgorin voimakkaaseen nousuun. Takaosan halkeamien suuntaisten takaosan solujen "vatsan" seinämien paksuuntuminen tarjoaa soluseinämän epätasaisen venytyksen; taka-solut saavat selvän papumaisen muodon, ja vatsan aukko avautuu. Kun fotosynteesin intensiteetti vähenee (esimerkiksi illalla), sokerien muodostuminen sulkevissa soluissa vähenee. Kaliumionien virta lakkaa. Solumehun konsentraatio sulkevissa soluissa vähenee verrattuna parotid-soluihin. Vesi osmoosin avulla jättää takasolut, alentaen niiden turgooria, seurauksena suoliston aukko sulkeutuu yöllä.
Orvaskeden solut ovat tiiviisti suljettuja, tämän ansiosta orvaskeni suorittaa useita toimintoja:
Estää taudinaiheuttajien tunkeutumisen kasviin;
Suojaa sisäisiä kudoksia mekaanisilta vaurioilta;
Säätelee kaasunvaihtoa ja verenkiertoa;
Vettä, suoloja vapautuu sen kautta;
Voi toimia imukudoksena;
osallistuu erilaisten aineiden synteesiin, ärsytyksen havaitsemiseen ja lehtien liikkeeseen.
Rihmamaisia - erimuotoisten, rakenteellisten ja toiminnallisten epidermaalisten solujen kasvu: karvat, vaa'at, setit jne. Ne on jaettu piiloon ja rauhasiin. Rauhasrauhaset,toisin kuin peitetyt, heillä on soluja, jotka erittävät erittymiä. Peittävät karvatmuodostaen kasvihuovan, huovan tai muun peitteen, heijastaa osaa auringonsäteistä ja vähentää siten verenkiertoa. Joskus karvat sijaitsevat vain siinä, missä stomata sijaitsee, esimerkiksi jalkalehden alapuolella. Joissakin kasveissa elävät karvat lisäävät haihtumispinta-alaa, mikä auttaa nopeuttamaan verenkiertoa.
Trikomiomit vaihtelevat huomattavasti. Pisimmät trichomit (jopa 5-6 cm) peittävät puuvillan siemenet. Kattavat trikomit ovat yksinkertaisten yksisoluisten tai monisoluisten, haarautuneiden tai tähtikuvioiden muodossa. Peittävät trikomit voivat pysyä hengissä pitkään tai kuolla nopeasti, täyttämällä ilmaa.
Trichomeista, joita esiintyy vain orvaskeden solujen mukana, ne eroavat toisistaan syntymistä, joiden muodostukseen osallistuvat subepidermaalikerrosten syvemmin sijaitsevat kudokset.
Anatomiset ja diagnostiset piirteet, joilla on tärkein merkitys ja suuri variaatio lääkeraaka-aineiden määrityksessä. Hiukset voivat olla yksinkertaisia \u200b\u200bja capitate, mikä puolestaan \u200b\u200bvoi olla yksisoluinen tai monisoluinen. Monisoluiset karvat voivat olla yksirivisiä, kaksirivisiä ja haarautuneita.
1.9.1. Yksinkertaiset karvat.
A. Yksinkertaiset yksisoluiset karvat

Kuva 68.Yksinkertaiset yksisoluiset karvat. A - papillary; B - säikeinen; B - vesikulaarinen; G - piikkimainen; D - koukun muotoinen; E - retortoidi; G - tylsä \u200b\u200blanka aallotettu; Z - akuutti kartiomainen; Ja - suorastaan \u200b\u200bkartiomainen; K - kaksiteräinen; L - kolmiteräinen; M, N, O - monipisteinen; P - mukulainen; P - mailan muotoinen
1. papillaarisen (Kuva 68, A; Kuva 69-74) - epidermisolujen matala, mutta leveä kasvu. Useammin muodostuu terälehdet. Siellä voi olla tylppä ja terävä. Niitä esiintyy timjamilehdillä, mäyräkukkilla, kolmivärivioleilla kukilla, kymmenvuotislehden reunalla, herjakasvien parran ruohossa jne.
2. kartio (Kuva 68, Z, I; kuva 75-79) - karvat, jotka ovat suorat ja kaltevat kulmaan pintaan nähden ja päättyvät tylppään (tylpän kartiomaiseen) tai terävään (terästi kartiomaiseen) päähän. Yleisimmät karvat. Yleensä on suoraa. Pintaan kallistuneita karvoja havaitaan timjamilla. Teräviä kartiomaisia \u200b\u200bkarvoja löytyy sennalehdiltä, \u200b\u200bpuolukan lehtiä, kolmiväriljorten ruohoa (lehtiä), timjamiruohoa jne. Tylsäkoonisia karvoja löytyy aniksen hedelmien orvaskentästä, kolmivärilikarttujen kukien terälehdistä.
3. lankaantuva (Kuva 68, B, G; Kuva 80, 81) - ohuet ja pitkät karvat. Voidaan olla suora ja aallotettu. Suorat filiformiset karvat löytyvät orapihlajan ja vadelman hedelmien orvaskentästä. Aallotettu tylsä \u200b\u200blanka karvoja havaitaan violetti terälehden epidermissä (kuva 82).
4. Retortovidnye (Kuva 68, E; Kuva 83) - karvat, joissa on laajennettu pohja ja kapea kaareva tai suora pää. Niitä löytyy humalan hedelmistä, serratus-ruoosta.
5. vesicular (Kuva 68, B; Kuva 84) - pienten kuplien muodossa olevat karvat. Niitä löytyy esimerkiksi kuolemattomien kukista.
6. koukussa (Kuva 68, D; Kuva 85-87) - kärkeen osoitetut ja koukun muodossa kaarevat hiukset. Nämä karvat ovat karhunlehden juuressa, puolukan, sennan, aniksen hedelmien lehtien pinnalla.
7. fimbriat (Kuva 88, 89) - pitkät orvaskeden solut, jotka sijaitsevat lehden, terälehden tai siipireunan reunaa pitkin. Tällaiset karvat ovat esimerkiksi mäyrä- ja violetti terälehtiä pitkin.
8. spinous (Kuva 68, D; Kuva 90) - karvat ovat muodoltaan lähes pyöristetty terävällä päällä. Nurmikolla havaitaan kolmivärisiä violetteja.
9. Club muotoinen (Kuva 68, P; Kuva 91) - hiukset, joissa on laajennettu päätyosa, joka muistuttaa muskaa. Saatavana timjami terälehdillä, violetti tricolor terälehdet.
10. Kaksi päätä (Kuva 68, K) - hiukset haaroittuneina kahteen päähän. Havaittu paimenen laukun ruohoa.
11. Kolmipisteinen (monipisteinen) (Kuva 68, L-O) - karvat haarautuneet kolmeen (tai useampaan) päähän. Havaittu paimenen laukun ruohoa.
12. mukuloita (Kuva 68, P) - kartiomaiset karvat, joiden pinnalla on ulkonevia osia (tuberkles).
B. Yksinkertaiset monisoluiset yksiriviset karvat
Monisoluisten hiusten ääriviivat voivat olla samat kuin yksisoluisten hiusten, mutta ne sisältävät kaksi tai useampia soluja ja vastaavasti niitä voidaan kutsua samanlaisiksi yksisoluisiksi hiuksiksi, joiden lukumäärä on osoitettu, esimerkiksi 2-soluinen retorttihius, 11-kennoinen säiemainen hiukset, 10-15-soluinen kartionmuotoinen hius.

Kuva 92.Yksinkertaiset monisoluiset karvat. A - kartiomainen; B - vesikulaarinen; Nivel; G - kaksois; D - huokoinen; E - T-muotoinen; F - kaksirivinen; Z - hilseilevä; Ja - palkki; L - yhdensuuntainen; M - haarukka; K - harjasta
1. kartio (Kuva 92, A; 93-96). Yleisimmin löydetyt hiukset. Ne ovat suorat ja kaltevat pintaan, viimeksi mainitut ovat harvinaisia \u200b\u200b(esimerkiksi timjamilla). Kartion muotoiset karvat voivat olla akuutteja ja tylppä-kartiomaisia. Terävät kartiomaiset karvat ovat yleisempää (oreganon yrtissä, mintun, salviaan jne. Lehdessä). Tylkeitä on kalanterikukissa.
2. Retortovidnye - karvat, joissa on laajennettu pohja ja kapea kaareva tai suora pää.
3. vesicular (Kuva 92, B; Kuva 97) - pienten kuplien muodossa olevat karvat. Pinnalla on immortelle-munasarjat.
4. lankaantuva - ohuet ja pitkät monisoluiset karvat.
5. koukussa - kärkeen osoitetut ja koukun muotoiset kaarevat hiukset.
6. Gusenitsevidnye (Kuva 98, 99) - karvat, jotka ovat lähes samanpaksuisia koko pituudeltaan ja koostuvat melkein identtisistä lyhyistä soluista, tylppä pää ja tornia muistuttavat. Nurmikolla havaitaan kolmivärisiä violetteja ruohojonossa.
7. fimbriat (Kuva 100) - pitkät monisoluiset epidermaalisten solujen kasvustot, jotka sijaitsevat lehden, terälehden tai siipin reunaa pitkin. Tapaa ruohoja.
8. Bichevidnye (Kuva 92, D; Kuva 101-104) - hiukset, joissa on monisoluinen emäs, joka koostuu lyhyiden solujen ketjusta ja pitkästä, filiformisesta, siniteetistä terminaalisolusta. Kirjallisuudessa tällaisia \u200b\u200bhiuksia kuvataan piiskamaisiksi, narunkaltaisiksi, langankaltaisiksi huovoiksi. Asteraceae-perheelle ominainen hiustyyppi. Löydetään piikkisirun nurmikosta, kuolemattomista kukista, pohjajalkalehdistä, tansikkokukista jne.
9. jäsentää (Kuva 92, B; Kuva 105, 106) - karvat, joilla on laajentunut solujen perusta niiden nivelkohdassa (muistuttavat niveltä). Samankaltaisia \u200b\u200bkarvoja löytyy emäkukka ruohoista, oregano-terälehdistä, harvoin timjamiruohoista.
10. mukuloita - karvat, joiden pinnalla on ulkonevia osia (tuberkles). Kaksisoluiset mukulakuidut havaitaan esimerkiksi termolehdessä.
B. Yksinkertaiset monisoluiset haarautuneet hiukset
1. yhdensuuntaisia (Kuva 92, L; Kuva 107, 108) - hiukset, jotka koostuvat kahdesta pitkästä suorasta solusta (karvasta), sulatettu pohjaan. Niitä esiintyy lehväkukissa, harvoin ruusun lannoissa ja orapihlajassa.
2. haarukka (Kuva 92, M; Kuva 109, 110) - hiukset, jotka koostuvat kahdesta pitkästä poikkipinnasta (hiuksista), sulatettu pohjaan. Tarkasteltavana lehmäkukissa, hyvin harvoin orapihlajan kukissa ja hedelmissä.
3. stellate (Kuva 111) - karvat, jotka koostuvat kolmesta tai pidemmästä siniaalisestä solusta (karvasta), sulautuneena pohjoihin. Löydetty lehmukukista.
4. skaalattu (Kuva 92, C) - hiukset, jotka koostuvat monisoluisesta levystä (pistorasian muodossa) ja lyhyestä jalasta (jalka voi puuttua). Tällaisia \u200b\u200bkarvoja löytyy tyrnistä. (Tämä karvojen nimi annetaan nykyaikaisen kasvitieteellisen terminologian mukaisesti; muiden lähteiden mukaan näitä hiuksia kutsutaan tähtikuvioiseksi tai corymbose-hilseksi.
5. T- (Kuva 92, E) - kaksiteräinen hiusraja, jolla on yksi monisoluinen jalka. Kirjallisuus kuvaa myös, kuinka torvi-, rokkarimainen.
6. cirrus (Kuva 92, D) - monisoluiset karvat, jotka muistuttavat haarapuuta. Löydetty esimerkiksi mulleinistä.
D. Yksinkertaiset monisoluiset moniriviset karvat
1. Kaksirivinen (Kuva 92, G; Kuva 112, 113) - karvat, joiden solut on järjestetty kahteen riviin. Niitä voidaan havaita kalanterin kukissa, yrttimeen ruohoa.
2. palkki (Kuva 92, I) - hiukset, jotka koostuvat kahdesta tai useammasta solusta, tiiviisti sulautettuina toisiinsa muodostaen kimpun.
3. kankea (Kuva 92 K, kuva 114) - monisoluiset hiukset, jotka koostuvat eripituisista hiuksista, jotka on sulautettu toisiinsa ja joilla on vapaat terävät päät. Tällaiset hiukset ovat yleensä haihtuvia hiuksia (perhe Asteraceae).Kirjallisuudessa niitä kutsutaan myös rosoisiksi monimutkaisiksi, huonomaisiksi.
1.9.2. Vangitse karvat. Capitate hiuksia kutsutaan myös rauhasiksi. Ne voivat olla yksisoluisia ja monisoluisia.

Kuva 115.Vangitse karvat. A - yksisoluinen jalka ja yksisoluinen pää; B - kaksirivinen jalka ja yksisoluinen pää; B - kaksirivinen jalka ja kaksirivinen pää; G - yksisoluinen jalka ja monisoluinen pää; D - monisoluisella jalkalla ja yksisoluisella päällä; W - monisoluinen jalka ja monisoluinen pää; Z - yksisoluinen (palava); Ja - corymbose
A. Yksisoluiset capitate hiukset
Tämän tyyppisten hiusten edustajat voivat olla kartiomaisia \u200b\u200btai jälkimaisia, mutta niiden päässä on aina pää. Tällaisia \u200b\u200bkarvoja on esimerkiksi nokkosenlehdillä. B. Monisoluiset capitate (rauhaset) hiukset
1. Hiukset, joissa on monisoluinen pää ja yksisoluinen jalka (Kuva 115, D; Kuva 116-120). Löydetty emäkukka ruoho, karhunlehdet.
2. Hiuksilla on yksisoluinen pää ja yksisoluinen jalka (Kuva 115, A; Kuva 121-123). Löydetty emäkukka ruoho, salvia lehdet, tomaatti kukat, minttu lehdet.
3. Hiuksilla, joissa on yksisoluinen pää ja monisoluinen jalka (Kuva 115, D; kuva 124). Löydetty emäkukka ruohoa, salvia lehtiä, kalenterikukat ja kamomilla pedicels.
4. Hiukset, joissa on monisoluinen pää ja monisoluinen jalkavarsi (Kuva 115, G; kuva 125). Löydetty emäkukka ruoho, karhunlehden lehdet, linden kukat, kalankurikukka, violetti ruoho.
5. Hiukset, joissa yksisoluinen pää ja kaksirivinen jalka (Kuva 115, B; kuva 126). Löytyi kalanterin ja kuolemattoman kukista.
6. Hiuksissa on kaksirivinen pää ja kaksirivinen jalka (Kuva 115, B; Kuva 127-130). Niitä löytyy kalanterin ja kuolemattoman kukista, marsin kanelin ruohosta.
7. Club muotoinen monisoluiset rauhasmaiset hiukset (kuva 131-133; katso kuva 155, D) - hiukset, joiden muodossa on laajennettu päätyosa, joka muistuttaa muskaa. Niitä voidaan havaita esimerkiksi puolukkalehdissä, violetissa ruohossa.
8. corymbose monisoluiset rauhaskarvat (kuva 115, I; kuva 134) ovat monikulmaisista ohutseinäisistä soluista valmistettu kilpi, joka istuu yhdellä, kaksisoluisella lyhyellä varrella. Sen alla vapautuva eteerinen öljy siirtää kiemuran kyynärpäästä. Niitä löytyy hap-hedelmistä.
1.10. Soluseinien ja karvojen peittävän kynsinauhan paksuuntumisen luonne
A. Paksuuntuneiden soluseinien luonne
1.10.1. ohutseinäinen (Kuva 135-137). Useimmat karvat ovat ohutseinäisiä. Monisoluisten pitkien ohutseinäisten karvojen soluseinät romahtavat toisinaan, häiritseen hiuksen tasaista muotoa. Tällaisia \u200b\u200bkarvoja esiintyy esimerkiksi cacalya-lehden alakerrossa.
1.10.2. paksuseinäinen (Kuva 138, 139). Niitä löytyy ruusunmarjoista, peräkkäin olevista ruohoista, Daursky-vaahan ruohoista jne.
1.10.3. Epätasaisesti paksuuntunut. Nurmikolla on mahdollista tarkkailla äiti-virnaa (Kuva 140). Rosehip-karvat, joissa on vino huokoset, kuuluvat samaan luokkaan (kuvat 141, 142).
B. Hiusneulan peittävyys
1.10.4. Sileä pinta (Kuva 143). Tällaisella pinnalla olevia hiuksia havaitaan esimerkiksi suvun, merimäkin ruohon jne. Lehdissä.
1.10.5. Sotainen pinta (katso kuva 1, G; kuva 144, 145) - orvaskesi muodostaa ulkonemat tuberkuloiden (syylät) muodossa. Karvoja on esimerkiksi violetissa ruohoissa, timjamiruohoissa, äitiherkän ruohoissa, mintunlehdissä, sennalehdissä jne. Lisäksi hiusten pinta voi olla hiukan syyläinen, kun muodostuu kynsinauhan pieniä ulkonemia, ja karkeat warteet, kun ulkonemat muodostavat merkittäviä. Ensimmäisessä tapauksessa violetin ruohon karvat voidaan mainita esimerkkinä, toisessa sennalehtien ja timjamiruohojen karvat.
1.10.6. Karkea pinta (katso kuva 68, P) - kynsinauha muodostaa erittäin suuret ulkonemat, esimerkiksi lansetoituneen termopsis-ruohon karvat.
10.7. Ryppyinen pinta - hiusneulan taivut tai aallot pohjasta kärkeen (kuva 146). Sitä havaitaan useammin papillaeissa, esimerkiksi kentaururuohoissa, piikkikukkakukissa. Rypistynyt pinta löytyy kuitenkin tavallisista karvoista, esimerkiksi ruohojonosta. Pohjassa minttulehtikarvoilla on ryppyinen pinta (kuva 147).
1.10.8. Katkoviiva pinta - orvaskentä muodostaa lyhyitä ulokkeita aivohalvauksen muodossa (tuberkles, syylät). Se on harvinainen ja on välivaihtoehto syylän ja ryppyisen pinnan välillä. Se havaitaan esimerkiksi pihlajan kukkien karvoissa (kuva 148).
1.11. Karvojen liittymispaikkojen ominaisuudet
1.11.1. Yleiset kiinnityskohdat (Kuva 149, 150) - karvat kiinnittyvät soluun tai orvaskeden solujen väliin. Yleisintä tyyppiä esiintyy esimerkiksi pohjajalkalehdissä, mintunlehdissä, salvialehdissä, äiti-ruoho-, violetti-ruohoissa jne.
1.11.2. Epidermaalisten solujen kasvu (katso kuva 68, A; 69-74). Niitä havaitaan piikkikukkakukissa, centaury-ruohoissa, äiti-terälehdessä, hermenkaarisien ruohoissa, aniksen hedelmien orvaskedessä jne.
1.11.3. Hiusten juuressa muodostuu epidermaalisten solujen ruusuke (Kuvio 151, 152). Sitä löytyy sennalehdistä, plantainlehdistä, oregano-ruoosta jne.
1.11.4. Laajennettu hiuspohja (Kuva 153). Sitä löytyy suvurakennuksen lehdistä.
1.11.5. Laajennettu osa hiuspohjasta on upotettu subepidermaalisten kudosten - emergenes - koteloon (katso kuva 115, 3). Se voidaan havaita esimerkiksi nokkosenlehdissä.
1.11.6. Monisoluinen hiuspohja (Kuva 154). Voit tarkkailla esimerkiksi ruohojonossa.
Usein hiukset hajoavat ja jättävät kiinnittymispaikkansa orvaskedelle, mikä on myös huomattava lääkekasvien materiaalin anatomisena ja diagnosoivana merkkinä.
TOINEN KANSI
Toissijaista integraalista kudosta kutsutaan peridermiä.Tämä on monivuotinen monivuotinen kudos monivuotisten varsien, juurten ja juurten kanssa. Se korvaa aksiaalisten elinten orvaskeden, joka vähitellen kuolee ja desquamates. Peridermi muodostetaan fallogeenistä (sekundaarinen meristeemi). Pellogeeni on upotettu orvaskesiin, subepidermaaliseen kerrokseen ja jopa aksiaalielinten syviin kerroksiin. Phalogeenin solut jaetaan seuraavasti: korkkisolut makaavat ulospäin ja phallodermin elävät parenkyymisolut ulospäin. Klooroplastit sisältyvät fellodermivarren soluihin.
 Korkki koostuu kuolleista soluista, joissa soluseinä on kyllästetty rasvamaisella aineella suberiinilla. Solut on järjestetty tasaisiin riveihin, ne ovat suorakaiteen muotoisia (poikkileikkaukseltaan), sopivat tiukasti toisiinsa muodostaen monikerroksisen kotelon. Korkki suojaa sisäisiä eläviä kudoksia kosteuden menettämiseltä, suurilta lämpötilanvaihteluilta ja mikro-organismien tunkeutumiselta. Korkin alla elävät kudokset tarvitsevat kaasunvaihtoa ja ylimääräisen kosteuden poistamista. Siksi stomaatan alla, koska subepidermaaliset kerrokset jakautuvat (jopa ennen peridermin ilmestymistä), ja myöhemmin fallogeenissä, elävät, löysästi sijaitsevat, monien solujen välisten tilojen ollessa parenkyymisoluja, joita kutsutaan suorittaa kutoauusi, joka hajottaa orvaskeden ja luo mahdollisuuden kaasunvaihtoon ja verenkiertoon ympäristön kanssa. Tätä rakenteellista muodostumista kutsutaan halkeamat(Kuva 2.12).
Korkki koostuu kuolleista soluista, joissa soluseinä on kyllästetty rasvamaisella aineella suberiinilla. Solut on järjestetty tasaisiin riveihin, ne ovat suorakaiteen muotoisia (poikkileikkaukseltaan), sopivat tiukasti toisiinsa muodostaen monikerroksisen kotelon. Korkki suojaa sisäisiä eläviä kudoksia kosteuden menettämiseltä, suurilta lämpötilanvaihteluilta ja mikro-organismien tunkeutumiselta. Korkin alla elävät kudokset tarvitsevat kaasunvaihtoa ja ylimääräisen kosteuden poistamista. Siksi stomaatan alla, koska subepidermaaliset kerrokset jakautuvat (jopa ennen peridermin ilmestymistä), ja myöhemmin fallogeenissä, elävät, löysästi sijaitsevat, monien solujen välisten tilojen ollessa parenkyymisoluja, joita kutsutaan suorittaa kutoauusi, joka hajottaa orvaskeden ja luo mahdollisuuden kaasunvaihtoon ja verenkiertoon ympäristön kanssa. Tätä rakenteellista muodostumista kutsutaan halkeamat(Kuva 2.12).
Kuva 2.12.Peridermin rakenne linsseillä: 1 - linssikudoksen suorittaminen; 2 - orvaskeden jäänteet; 3 - korkki (putosi); 4 - pellogeeni; 5 - felloderma
Linssit, joilla on pieniä tuberkuloita, erottuvat selvästi puiden ja pensaiden versojen pinnalta (kuva 2.13).
Koivun kannoilla niiden jäännökset havaitaan tyypillisissä poikittaisissa mustissa raidoissa, haavassa ne ovat rommien muodossa.
Tertiäärinen integraatti
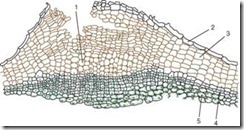
 Kuori (ritidi)edustaa tertiääristä yhtenäistä kudosta, joka muodostuu monivuotisissa kasveissa juuressa, varressa ja juurakossa. Joka vuosi syvempiin kerroksiin laitetaan uusi faaligeenikerros ja muodostetaan peridermi. Haavan ulompi kerros - korkki - eristää kaikki päällä olevat kudokset, minkä seurauksena ne kuolevat. Täten joukko lukuisia peridermia, joiden välissä on kuolleita kudoksia, on kuori (kuva 2.14).
Kuori (ritidi)edustaa tertiääristä yhtenäistä kudosta, joka muodostuu monivuotisissa kasveissa juuressa, varressa ja juurakossa. Joka vuosi syvempiin kerroksiin laitetaan uusi faaligeenikerros ja muodostetaan peridermi. Haavan ulompi kerros - korkki - eristää kaikki päällä olevat kudokset, minkä seurauksena ne kuolevat. Täten joukko lukuisia peridermia, joiden välissä on kuolleita kudoksia, on kuori (kuva 2.14).
Kuva 2.14.Tammenkuori: 1 - korkkikerrokset; 2 - kuitu; 3 - primaarikuoren jäänteet; 4 - kalsiumoksalaatin drusen