Eläimet ovat asettuneet melkein koko maapallolle. Koska liikkuvuus, kyky evoluuttisesti sopeutua kylmempiin olosuhteisiin, koska heillä ei ole suoraa riippuvuutta auringonvalosta, eläimet käyttivät enemmän elinympäristöjä kuin kasveja. On kuitenkin muistettava, että eläimet ovat riippuvaisia \u200b\u200bkasveista, koska kasvit toimivat ravintolähteenä (kasvissyöjille ja saalistajat syövät kasvissyöjiä).
Ymmärrämme tässä eläinten elinympäristöjen yhteydessä eläinten elinympäristö.
Eläinympäristöjä on yhteensä neljä. Näitä ovat 1) maa-ilma, 2) vesi, 3) maaperä ja 4) muut elävät organismit. Maa- ja ilma-elämän ympäristöstä puhutaan joskus maa- ja erikseen ilmasta. Jopa lentävät eläimet istuvat ennemmin tai myöhemmin maassa. Lisäksi liikkuessaan maassa eläin on myös ilmassa. Siksi maa- ja ilmaympäristö yhdistetään yhdeksi maa-ilmaksi.
On eläimiä, jotka elävät kahdessa ympäristössä kerralla. Esimerkiksi, monet sammakkoeläimet (sammakot) elävät sekä vedessä että maassa; monet jyrsijät elävät maaperässä ja maan pinnalla.
Maa-ilman elinympäristö
Ilmassa, useimmat eläinlajit. Maa osoittautui tietyllä tavalla heidän elämänsä mukavimmaksi ympäristöksi. Vaikka evoluutiossa, eläimet (ja kasvit) nousivat veteen ja tulivat myöhemmin pinnalle.
Useimmat madot, hyönteiset, sammakkoeläimet, matelijat, linnut ja nisäkkäät elävät maalla. Monet eläinlajit kykenevät lentämään, joten ne viettävät osan elämästään yksinomaan ilmassa.
Ilma-maa-ympäristön eläimille on yleensä ominaista korkea liikkuvuus, hyvä näkö.
Pohja-ilmaympäristölle on ominaista laaja valikoima elinolosuhteita (trooppiset metsät ja lauhkeat metsät, niityt ja stepit, aavikot, tundra ja paljon muuta). Siksi tämän elinympäristön eläimille on luonteenomaista suuri valikoima, ne voivat erota toisistaan \u200b\u200bsuuresti.
Vesieliöstö
Vesieliöstö eroaa ilmasta tiheydessä. Täällä eläimillä on varaa olla erittäin massiivisia ruumiita (valaita, haita), koska vesi tukee niitä ja helpottaa heidän ruumiinsa. Siirtyminen tiheässä ympäristössä on kuitenkin vaikeampaa, joten vesieläimissä virtaviivainen kehon muoto on useimmiten.
Auringonvalo tunkeutuu tuskin meren syvyyteen, joten syvänmeren eläimissä näköelimet voivat olla huonosti kehittyneitä.
Vesieläimet jaetaan planktoniin, nektoniin ja pohjaeliöihin. plankton passiivisesti ui vesipatsaassa (esimerkiksi yksisoluinen), nekton - nämä ovat aktiivisesti uima-eläimiä (kalat, valaat jne.), pohjaeliöstölle asuu pohjassa (korallit, sienet jne.).
Maaperän elinympäristö
Maaperä elinympäristönä on hyvin tiheää ja puuttuu auringonvalosta. Tässä eläimet eivät tarvitse näköelimiä. Siksi niitä ei ole joko kehittynyt (matoja) tai vähentyneitä (myyrät). Toisaalta maaperällä ei ole niin merkittäviä lämpötilaeroja kuin pinnalla. Monet madot, hyönteisten toukat ja muurahaiset elävät maaperässä. Nisäkkäiden joukossa on myös maaperää: moolit, myyrirotit, urossa olevat eläimet.
VESIYMPÄRISTÖ
Elämän vesiympäristö (hydrosfääri) hallitsee 71% maapallosta. Yli 98% vedestä on keskittynyt meriin ja valtameriin, 1,24% - napa-alueiden jäätä, 0,45% - jokien, järvien, suiden makeaa vettä.
Valtamerissä erotellaan kaksi ympäristöaluetta:
vesipylväs - pelagiset alueja pohja - benthala.
Noin 150 000 eläinlajia elää vesiympäristössä, eli noin 7% niiden kokonaismäärästä ja 10 000 kasvilajea - 8%. Erota seuraava hydrobionttien ekologiset ryhmät.Pelagiaalinen - organismien asuttama nektoni ja planktoni.
Necton (Nectos - kelluva) - tämä on joukko pelagisia aktiivisesti liikkuvia eläimiä, joilla ei ole suoraa yhteyttä pohjaan. Nämä ovat pääasiassa suuria eläimiä, jotka voivat kulkea pitkiä matkoja ja voimakkaita vesivirtauksia. Niille on ominaista virtaviivainen kehon muoto ja hyvin kehittyneet liikuntaelimet (kalat, kalmarit, sorkkaeläimet, valaat) .Makeissa vesissä kalojen lisäksi nektoniin kuuluvat sammakkoeläimet ja aktiivisesti liikkuvat hyönteiset.
Plankton (vaeltava, huiman) - tämä on joukko pelagisia organismeja, joilla ei ole kykyä nopeasti aktiivisiin liikkeisiin. Ne jaetaan kasvien ja eläinplanktonien joukkoon (pienet äyriäiset, alkueläimet - foraminifera, radiolaarit; meduusat, siipiset nilviäiset). Kasviplanktoni - piimat ja vihreät levät.
neuston - joukko organismeja, jotka asuvat veden pintakalvossa ilman rajalla. Nämä ovat havupuiden, ruukkien, simpukoiden, kotilojen ja simpukoiden, piikkinahkaisten ja kalojen toukkia. Toukkavaiheen ohitettuaan ne jättävät pintakerroksen, joka toimi heidän turvapaikkanaan, ja siirtyvät asumaan pohjaan tai pelagisiin lajeihin.
Playston -tämä on joukko organismeja, joiden ruumiin osa on veden pinnan yläpuolella, ja toinen vedessä - duckweed, siphonophores.
Bentos (syvyys) - joukko organismeja, jotka elävät vesistöjen pohjassa. Se on jaettu fytobentosiin ja zoobenthosiin. Kasvibentos - levät - piimat, vihreät, ruskeat, punaiset ja bakteerit; rannikon edustalla, kukkivat kasvit - zoster, rupia. Eläinbentos - foraminifera, sienet, suolisto, madot, nilviäiset, kalat.
Vesieliöiden elämässä veden pystysuunnassa liikkumisella, tiheydellä, lämpötilalla, valolla, suolalla, kaasulla (happi ja hiilidioksidi) ja viini-ionien pitoisuuksilla (pH) on tärkeä merkitys.
Lämpötilatila: Se eroaa vedessä ensinnäkin pienemmästä lämpövirrasta ja toiseksi suuremmasta vakaudesta kuin maassa. Osa veden pinnalle tulevasta lämpöenergiasta heijastuu, osa kuluu haihtumiseen. Veden haihtuminen säiliöiden pinnalta, jolla kuluu noin 2263,8 J / g, estää alempien kerrosten ylikuumenemisen, ja sulamisen lämpöä vapauttavan jään muodostuminen (333,48 J / g) hidastaa niiden jäähtymistä. Lämpötilan muutos virtaavissa vesissä seuraa sen muutoksia ulkoilmassa, jotka eroavat pienemmästä amplitudista.
Järjestelmissä, joissa on kohtalainen leveysaste, järvien ja lampien lämpötila asetetaan tunnetulla fysikaalisella ilmiöllä - veden enimmäistiheys on 4 ° C. Niissä oleva vesi on selvästi jaettu kolmeen kerrokseen:
1. epilimnion- yläkerros, jonka lämpötilassa on voimakkaita kausivaihteluita;
2. metalimnion - siirtymäkerros lämpötilahyppyä, huomattava lämpötilaero havaitaan;
3. alusveden - syvänmeren kerros, joka saavuttaa pohjan, jossa lämpötila muuttuu hieman vuoden aikana.
Kesällä lämpimimmät vesikerrokset sijaitsevat pinnalla ja kylmimmät pohjassa. Tämän tyyppistä kerros kerrokselta tapahtuvaa lämpötilan jakautumista säiliössä kutsutaan suora kerrostuminen. Talvella lämpötilan laskiessa käänteinen kerrostuminen: pintakerroksen lämpötila on lähellä 0 ° C, alhaalla lämpötila on noin 4 ° C, mikä vastaa sen maksimitiheyttä. Lämpötila nousee siten syvyydellä. Tämä ilmiö kutsui lämpötilan dichotomia havaittu useimmissa lauhkean vyöhykkeen järvissä kesällä ja talvella. Lämpötilaerotuksen seurauksena pystysuuntainen kierto on häiriintynyt - tilapäinen pysähtymisjakso alkaa - pysähtyneisyys.
Keväällä pintavesi lämmitetään 4 ° C: seen tiheämmäksi ja vajoaa syvemmälle, ja lämpimämpi vesi nousee paikoilleen syvyydestä. Tällaisen vertikaalisen kiertämisen seurauksena säiliössä tapahtuu homotermiaa, ts. jonkin aikaa koko vesimassan lämpötila on tasaantunut. Lämpötilan noustessa edelleen, ylemmät kerrokset muuttuvat yhä vähemmän tiheiksi eivätkä enää laskeudu - kesästagnaatio. Syksyllä pintakerros jäähtyy ja tiivistyy ja vajoaa syvemmälle, siirtäen lämpimämmän veden pintaan. Tämä tapahtuu ennen syksyn homotermian alkamista. Jäähdytettäessä pintavettä alle 4C: n lämpötilassa ne muuttuvat vähemmän tiheiksi ja jäävät jälleen pintaan. Seurauksena on veden kierto ja talven pysähtyminen.
Vedelle ominainen tiheys (800 kertaa) ilmaa parempi) ja viskositeetti. keskimäärin vesipatsaassa jokaista 10 metriä syvyyttä paine kasvaa 1 atm. Kasveissa nämä piirteet heijastuvat siihen tosiasiaan, että niissä on hyvin vähän tai ei lainkaan mekaanista kudosta, joten niiden varret ovat erittäin elastisia ja taipuisia helposti. Useimmilla vesikasveilla on kelluvuus ja kyky olla suspensiona vesipylväässä, monissa vesieläimissä päällysteet lima, joka vähentää kitkaa liikkumisen aikana, ja vartalo saa virtaviivaisen muodon. Monet asukkaat ovat suhteellisen vakaa ja rajoittuvat tiettyihin syvyyksiin.
Kalvo ja kevyt tila. Tämä vaikuttaa erityisesti kasvien jakautumiseen: mutaisissa säiliöissä ne elävät vain pintakerroksessa. Valojärjestelmä johtuu myös säännöllisestä valon pienenemisestä syvyydessä johtuen siitä, että vesi imee auringonvaloa. Samalla eri aallonpituuksilla olevat säteet absorboituvat eri tavalla: punaisin nopeimmin, kun taas sinivihreä tunkeutuu merkittävään syvyyteen. Väliaineen väri muuttuu, muuttuen vähitellen vihertävästä vihreäksi, siniseksi, siniseksi, sinivioletiksi, korvattu jatkuvalla pimeydellä. Siksi syvyydessä vihreät levät korvataan ruskeilla ja punaisilla, joiden pigmentit on sovitettu sieppaamaan auringonvaloa eri aallonpituuksilla. Myös eläinten väri muuttuu luonnollisesti syvyyden myötä. Kirkkaat ja eriväriset eläimet elävät veden pintakerroksissa, kun taas syvänmeren lajeista puuttuu pigmenttejä. Mannekeissa on eläviä eläimiä, jotka on maalattu väreillä punertavalla sävyllä, joka auttaa heitä piiloutumaan vihollisilta, koska sininen-violetti säde punainen nähdään mustana.
Valon absorptio vedessä on sitä voimakkaampaa, mitä vähemmän sen läpinäkyvyys on. Läpinäkyvyydelle on ominaista äärimmäinen syvyys, jossa erityisesti Secchi-laskettu levy on edelleen näkyvissä (valkoinen levy, jonka halkaisija on 20 cm). Siksi fotosynteesivyöhykkeiden rajat vaihtelevat suuresti eri vesistöissä. Puhtaimmissa vesissä fotosynteesivyöhyke saavuttaa 200 metrin syvyyden.
Veden suolapitoisuus. Vesi on erinomainen liuotin monille mineraaliyhdisteille. Seurauksena on, että tietty kemiallinen koostumus kuuluu luonnonvesimuodostumiin. Tärkeimpiä ovat sulfaatit, karbonaatit, kloridit. Liukenetujen suolojen määrä 1 litrassa vettä makeassa vedessä on enintään 0,5 g, merissä ja valtamereissä - 35 g. Makean veden kasvit ja eläimet elävät hypotonisessa ympäristössä, ts. ympäristö, jossa liuenneiden aineiden pitoisuus on alhaisempi kuin kehon nesteissä ja kudoksissa. Kehon sisällä ja ulkopuolella esiintyvän osmoottisen paine-eron vuoksi vettä tulee jatkuvasti vartaloon, ja makean veden vesieliöt pakotetaan poistamaan sitä intensiivisesti. Tässä suhteessa he ovat korostaneet osmoregulaatioprosesseja. Alkueläimissä tämä saavutetaan eritys tyhjiöiden avulla, monisoluisissa, poistamalla vettä erittymisjärjestelmän läpi. Tyypillisesti meren ja tyypillisesti makean veden lajit eivät siedä merkittäviä muutoksia veden suolapitoisuudessa - stenohaliinijärjestöissä. Euryhallins - makeanveden hauki ahven, lahna, hauki, merestä - monikkoperhe.
KaasutilaVesiympäristön pääkaasut ovat happi ja hiilidioksidi.
happi - Tärkein ympäristötekijä. Se tulee veteen ilmasta ja kasvit erittävät sitä fotosynteesin aikana. Sen pitoisuus vedessä on käänteisesti verrannollinen lämpötilaan - lämpötilan laskiessa hapen liukoisuus veteen (kuten myös muihin kaasuihin) kasvaa. Eläinten ja bakteerien tiheästi asuttamissa kerroksissa hapenpuute voi syntyä lisääntyneen kulutuksen vuoksi. Joten valtamerellä 50–1000 metrin rikkaalle elämänsyvylle on ominaista ilmaston heikko heikkeneminen. Se on 7-10 kertaa pienempi kuin kasviplanktonin asuttamissa pintavesissä. Vesistöjen pohjan lähellä olosuhteet voivat olla lähellä anaerobisia.
Hiilidioksidi - se liukenee veteen noin 35 kertaa paremmin kuin happi ja sen pitoisuus vedessä on 700 kertaa suurempi kuin ilmakehässä. Tarjoaa vesikasvien fotosynteesiä ja osallistuu selkärangattomien eläinten kalkkipitoisten luurankojen muodostumiseen.
Vetyionien pitoisuus (pH)- makean veden uima-altaat, joiden pH \u003d 3,7-4,7, pidetään happamaksi, 6,95-7,3 neutraaliksi ja pH 7,8 alkaliseksi. Makeassa vedessä pH kokee jopa vuorokauden vaihteluita. Merivesi on alkalisempaa ja sen pH muuttuu huomattavasti vähemmän kuin tuoreessa. Syvyyden myötä pH laskee. Vetyionien konsentraatiolla on suuri rooli hydrobionttien jakautumisessa.
Maa-ilman elinympäristö
Pohja-ilma-elämän ympäristö on erityinen, että täällä asuvia organismeja ympäröi kaasumainen ympäristö, jolle on ominaista alhainen kosteus, tiheys ja paine sekä korkea happipitoisuus. Tämän ympäristön eläimet liikkuvat yleensä maaperän (kiinteän substraatin) ympärillä ja kasvit juurtuvat siihen.
Pohja-ilmaympäristön ympäristötekijöillä on useita ominaispiirteitä: suurempi valon voimakkuus verrattuna muihin ympäristöihin, merkittävät lämpötilanvaihtelut, kosteuden muutokset maantieteellisestä sijainnista, vuodenajasta ja vuorokaudenajasta riippuen. Edellä lueteltujen tekijöiden vaikutus liittyy erottamattomasti ilmamassien - tuulen - liikkeeseen.
Evoluutioprosessissa pinta-ilmaympäristön elävissä organismeissa on kehittynyt ominaisia \u200b\u200banatomisia, morfologisia ja fysiologisia mukautuksia.
Tarkastellaan tärkeimpien ympäristötekijöiden vaikutuksia kasvit ja eläimet ilmassa-maaympäristössä.
Ilmaa. Ilmalle ympäristötekijänä on ominaista vakio koostumus - happea siinä on yleensä noin 21%, hiilidioksidia 0,03%.
Matala ilman tiheysmäärittää sen alhaisen nostovoiman ja lievän tuen. Kaikki ilmaympäristön asukkaat ovat läheisesti yhteydessä maan pintaan, palvelevat heitä kiinnittymisessä ja tukemisessa. Ilman tiheydellä ei ole suurta vastustusta organismeille, kun ne liikkuvat maan pinnalla, mutta vaikeuttaa liikkumista pystysuunnassa. Useimmille organismeille ilmaan jääminen liittyy vain uudelleensijoittamiseen tai saaliin etsimiseen.
Matala ilmahissi määrittää maanpäällisten organismien lopullisen massan ja koon. Suurimmat maan pinnalla elävät eläimet ovat pienempiä kuin vesiympäristön jättiläiset. Suuret nisäkkäät (nykyaikaisen valaan koko ja paino) eivät voineet elää maalla, koska ne murskataan oman painonsa avulla.
Ilman pieni tiheys luo vähän vastustuskykyä liikkumiselle. Monet maa-eläimet ovat käyttäneet evoluution aikana tämän ilmaympäristön ominaisuuden ympäristöhyötyjä saavuttaen lentämiskyvyn. 75% kaikista maaeläimistä kykenee aktiiviseen lentää, lähinnä hyönteisiä ja lintuja, mutta lentolehtisiä löytyy myös nisäkkäistä ja matelijoista.
Ilman liikkuvuuden, ilmakehän alemmissa kerroksissa olevien ilmamassien pystysuorien ja vaakasuorien liikkeiden takia useiden organismien passiivinen lento on mahdollista. Monilla lajeilla on kehittynyt anemochoria - uudelleensijoittaminen ilmavirroilla. Anemochoria on ominaista itiöille, kasvien siemenille ja hedelmille, alkueläinten kystat, pienet hyönteiset, hämähäkit jne. Ilmavirtojen passiivisesti kantamia organismeja kutsutaan kollektiivisesti aeroplanktoniksi analogisesti vesiympäristön planktonisten asukkaiden kanssa.
Vaakasuorien ilmanvaihdosten (tuulet) pääasiallinen ekologinen merkitys on epäsuora sellaisten tärkeiden ympäristötekijöiden kuin lämpötilan ja kosteuden vaikutuksen lisäämiseen ja heikentämiseen maanjäristyksissä. Tuulet lisäävät kosteuden ja lämmön palautumista eläimiin ja kasveihin.
Ilman kaasukoostumus pintakerroksen ilma on melko tasaista (happi - 20,9%, typpi - 78,1%, inertit kaasut - 1%, hiilidioksidi - 0,03 tilavuusprosenttia) johtuen korkeasta diffuusiokyvystään ja jatkuvasta sekoittumisesta konvektiolla ja tuulen virtauksilla. Paikallisista lähteistä ilmakehään saapuvien kaasumaisten, pisaran nestemäisten ja kiinteiden (pöly) hiukkasten epäpuhtauksilla voi kuitenkin olla merkittävä ympäristövaikutus.
Korkea happipitoisuus osaltaan lisäsi aineenvaihduntaa maanpäällisissä organismeissa, ja hapettumisprosessien suuren tehokkuuden perusteella syntyi eläinten homoyotermia. Happi, koska se sisältää jatkuvasti korkean pitoisuuden ilmassa, ei ole rajoittava tekijä maanpäällisessä ympäristössä. Vain paikoin, tietyissä olosuhteissa, väliaikaisia \u200b\u200bpuutteita esiintyy esimerkiksi rappeutuvien kasviromujen rypäleissä, viljavarastoissa, jauhoissa jne.
Edafiset tekijät. Maaperän ja maaston ominaisuudet vaikuttavat myös maaperäisten organismien, pääasiassa kasvien, elinoloihin. Maan pinnan ominaisuuksia, joilla on ympäristövaikutuksia sen asukkaille, yhdistävät nimi edafilaiset ympäristötekijät.
Kasvien juurijärjestelmän luonne riippuu maaperän hydrotermisestä käytöstä, ilmastosta, lisäyksestä, koostumuksesta ja rakenteesta. Esimerkiksi puulajien (koivu, lehtikuusi) juurijärjestelmät ikiroutaa olevilla alueilla sijaitsevat matalassa syvyydessä ja leviävät leveyteen. Siellä, missä ikiroutaa ei ole, näiden kasvien juurijärjestelmät ovat vähemmän levinneitä ja tunkeutuvat syvemmälle. Monissa arojen kasveissa juuret voivat saada vettä suuresta syvyydestä, kun taas samaan aikaan niiden juurien pintajuuret ovat maaperän humushorisontissa, josta kasvit imevät mineraaliravinteita.
Maasto ja maaperän luonne vaikuttavat eläinten liikkumisen erityispiirteisiin. Esimerkiksi avoimissa tiloissa elävät sorkka- ja kavioeläimet, strutot, piiskat tarvitsevat vankkaa maata parantamaan torjuntaa nopean juoksun aikana. Likaisella hiekalla elävissä liskoissa sormet reunustavat sarviasteikkojen reunalla, mikä lisää tuen pintaa. Maan asukkaille, jotka kaivaa reikiä, tiheä maaperä on epäsuotuisa. Maaperän luonne vaikuttaa joissain tapauksissa maaeläinten, jotka kaivaa reikiä, hautaamaan maahan säästääkseen lämmöltä tai petoeläimiltä, \u200b\u200btai munivien munien maaperään, jakautumiseen jne.
Sää- ja ilmasto-ominaisuudet.Sääolosuhteet monimutkaistavat edelleen ilmasto-olosuhteita. Sää on jatkuvasti muuttuva ilmakehän tila maan pinnalla noin 20 km korkeuteen (troposfäärin raja). Säävaihtelu ilmenee ympäristötekijöiden, kuten lämpötilan ja kosteuden, pilvien peittävyyden, sateen, tuulen voimakkuuden ja suunnan, yhdistelmän jatkuvassa vaihtelussa. Säämuutoksille ja niiden säännölliselle vuosisyklin vuorottelulle ovat tyypillisiä epäjaksolliset vaihtelut, mikä monimutkaistaa merkittävästi maaperäisten organismien olemassaoloa. Sää vaikuttaa vesieliöiden elämään paljon vähemmän ja vain pintakerrosten väestöön.
Alueen ilmasto. Pitkäaikaiset sääolosuhteet kuvaavat alueen ilmastoa. Ilmastokonseptiin sisältyy meteorologisten ilmiöiden keskiarvojen lisäksi myös niiden vuotuinen ja päivittäinen kulku, poikkeamat ilmastosta ja niiden esiintymistiheys. Ilmaston määrää alueen maantieteelliset olosuhteet.
Ilmastoalueiden alueellista monimuotoisuutta monimutkaisten tuulien vaikutus, syklonien ja antisyklonien jakautuminen, vuoristoalueiden vaikutus ilmamassajen liikkeeseen, etäisyys merestä ja monet muut paikalliset tekijät.
Suurimmalle osalle maanpäällisiä organismeja, etenkin pieniä, ei ole tärkeätä alueen ilmasto vaan niiden välittömän elinympäristön olosuhteet. Hyvin usein paikalliset ympäristöelementit (reljeefi, kasvillisuus jne.) Muuttavat lämpötilan, kosteuden, valon, ilman liikkumista tietyllä alueella siten, että se eroaa merkittävästi alueen ilmasto-olosuhteista. Tällaisia \u200b\u200bpaikallisia ilmastomuutoksia, jotka taittuvat pintailmakerrokseen, kutsutaan mikroilmastoksi. Kullakin vyöhykkeellä mikroilmasto on hyvin monimuotoista. Mielivaltaisen pienten osien mikroklimatit voidaan erottaa. Esimerkiksi erityinen järjestelmä luodaan kukkakoroloihin, joita siellä asuvat asukkaat käyttävät. Erityinen vakaa mikroilmasto esiintyy uroissa, pesissä, onteloissa, luolissa ja muissa suljetuissa paikoissa.
Sademäärä. Vedenjakelun ja kosteuden varastoinnin lisäksi niillä voi olla toinen ympäristörooli. Siksi rankkasateilla tai rakeilla on joskus mekaanisia vaikutuksia kasveihin tai eläimiin.
Lumipeitteen ekologinen rooli on erityisen monimuotoinen. Päivittäiset lämpötilanvaihtelut läpäisevät lumen paksuuden vain 25 cm asti, syvemmässä lämpötila muuttuu tuskin. -20-30 ° C: n pakkasissa lämpötila on vain vähän alle nollan. Syvä lumipeite suojaa uusiutuvia silmuja, suojaa kasvien vihreitä osia jäätymiseltä; monet lajit menevät lumen alle pudottamatta lehtiä, esimerkiksi karvainen päänahka, lääketieteellinen veronika jne.
Pienet maaeläimet elävät aktiivista elämäntapaa myös talvella, asettaessaan lumen alle ja sen paksuudessa kokonaisia \u200b\u200bgallerioita käytäviä. Joillekin lumisella kasvillisuudella ruokkiville lajeille on ominaista tasainen talvisaapuminen, jota havaittiin esimerkiksi lemmingsissä, metsä- ja keltakarahiirissä, useissa peltohirroissa, vesirottissa ja muissa ritarilintuissa - pähkinänrinta, mustasirra ja ptarmigani -, joka urheili lumessa yön yli.
Suurten eläinten talvinen lumipeite vaikeuttaa ruoan saamista. Monet sorkka- ja kavioeläimet (porot, villisiat, myskin härät) ruokkivat yksinomaan talvella lumen peittämää kasvillisuutta, ja syvä lumipeite ja erityisesti sen pinnalla oleva kova kuori, joka näkyy jäässä, tuomitsee heidät hölynpölyyn. Lumen syvyys saattaa rajoittaa lajien maantieteellistä jakautumista. Esimerkiksi oikeat hirvieläimet eivät tunkeudu pohjoiseen alueille, joilla lumen paksuus talvella on yli 40-50 cm.
Valotila. Maapallon pintaan päästävän säteilyn määrä johtuu maaston maantieteellisestä leveydestä, päivän pituudesta, ilmakehän läpinäkyvyydestä ja auringonvalon esiintymiskulmasta. Eri sääolosuhteissa 42-70% aurinkovakiosta saavuttaa maanpinnan. Valaistus maan pinnalla vaihtelee suuresti. Kaikki riippuu auringon korkeudesta horisontin yläpuolella tai auringonvalon esiintymiskulmasta, päivän pituudesta ja sääolosuhteista sekä ilmakehän läpinäkyvyydestä. Valon voimakkuus vaihtelee myös vuodenajan ja vuorokaudenajan mukaan. Joillakin maapallon alueilla valon laatu ei ole vastaava, esimerkiksi pitkäaaltosäteen (punainen) ja lyhytaalto (sininen ja ultravioletti) säteiden suhde. Lyhytaaltoisten säteiden tiedetään olevan suurempia kuin pitkäaaltosäteiden, ilmakehän absorboimat ja hajottamat.
Maaperä elinympäristönä
Maaperä on löysä ohuen pintakerros, joka on kosketuksissa ilman kanssa. Maaperä ei ole vain kiinteä kappale, kuten suurin osa litosfäärin kivistä, vaan monimutkainen kolmivaiheinen järjestelmä, jossa kiinteitä hiukkasia ympäröi ilma ja vesi. Sitä läpäisevät onkalot, jotka on täytetty kaasujen ja vesiliuoksien seoksella, ja siksi se luo erittäin monipuoliset olosuhteet, jotka ovat suotuisat monien mikro- ja makro-organismien elämälle. Maaperässä lämpötilanvaihtelut tasoittuvat pintailmakerrokseen verrattuna, ja pohjaveden läsnäolo sekä sateiden läpäisy luovat kosteusvarantoja ja muodostavat kosteusjärjestelmän välivaiheen veden ja maaympäristön välillä. Maaperä keskittää orgaanisten ja mineraalisten aineiden varastot, joita kuoleva kasvillisuus ja eläinten ruhot toimittavat. Kaikki tämä määrittelee maaperän suuren kylläisyyden elämästä.
Maaperän olosuhteiden heterogeenisyys on selkein pystysuunnassa. Syvyyden myötä joukko tärkeimpiä maaperän asukkaiden elämään vaikuttavia ympäristötekijöitä muuttuu dramaattisesti. Ensinnäkin tämä liittyy maaperän rakenteeseen. Siinä erotetaan kolme päähorisonttia, jotka eroavat toisistaan \u200b\u200bmorfologisista ja kemiallisista ominaisuuksista: 1) ylempi humuskerääntymishorisontti, johon orgaaniset aineet kerääntyvät ja muuttuvat ja josta osa yhdisteistä pestään pesemällä vettä; 2) pesuhorisontti tai illuviaalinen, jossa ylhäältä pestyt aineet laskeutuvat ja muuttuvat, ja 3) emäkivi tai horisontti, jonka materiaali muuttuu maaperään.
Maaperän hiukkasten välisten onteloiden koko, joka soveltuu eläimille elämiseen, pienenee yleensä nopeasti syvyyden myötä. Esimerkiksi niityn maaperässä onteloiden keskimääräinen halkaisija 0-1 mm: n syvyydessä on 3 mm; 1-2 cm 2 mm ja 2-3 cm syvyydessä - vain 1 mm; syvemmät maahuokoset ovat vielä pienempiä.
Maaperässä on kosteutta erilaisissa olosuhteissa: 1) maa-ainepartikkelien pinta pitää kiinni sitoutuneena (hygroskooppinen ja kalvo); 2) kapillaari vie pieniä huokosia ja voi liikkua niitä pitkin eri suuntiin; 3) painovoima täyttää suurempia tyhjiöitä ja imee hitaasti painovoiman vaikutuksesta; 4) maaperän ilmaan sisältyy höyryä.
Maaperän ilman koostumus on vaihteleva. Syvyyden myötä sen happipitoisuus laskee voimakkaasti ja hiilidioksidipitoisuus kasvaa. Hajoavien orgaanisten aineiden esiintymisen vuoksi maaperässä voi olla korkea pitoisuus myrkyllisiä kaasuja, kuten ammoniakki, rikkivety, metaani jne. Kun maaperää tulvii tai intensiivinen kasvinmädanto pysyi mätää, paikoissa voi esiintyä täysin anaerobisia olosuhteita.
Leikkuulämpötilan vaihtelut vain maaperän pinnalla. Täällä ne voivat olla jopa voimakkaampia kuin ilman pintakerroksessa. Jokaisen senttimetrin syvyydessä päivittäiset ja vuodenaikojen lämpötilan muutokset ovat kuitenkin pienempiä ja niitä ei käytännössä enää näy 1-1,5 m syvyydessä.
Kaikki nämä piirteet johtavat siihen, että huolimatta maaperän ympäristöolosuhteiden suuresta heterogeenisyydestä, se toimii melko vakaana ympäristöna, erityisesti maaperän organismeille. Jyrkkä kosteusgradientti maaprofiilissa antaa maaperän eliöille mahdollisuuden tarjota itselleen sopivat ympäristöolosuhteet pienillä liikkeillä.
Maaperän asukkaat voidaan jakaa suureen ja liikkuvuusasteesta riippuen useisiin ryhmiin:
1. mikrobisto - nämä ovat maaperän mikro-organismeja, jotka muodostavat poikkisuuntaisen ravintoketjun päälinkin, ja ne ovat ikään kuin välipitkä kasvijäämien ja maaperäisten eläinten välillä. Näitä ovat vihreät ja sinilevälevät, bakteerit, sienet ja alkueläimet. Nämä ovat vesieliöitä, ja maaperä heille on mikrovirtausjärjestelmä. Ne elävät maahuokosissa, jotka on täytetty painovoimaisella tai kapillaarisella kosteudella, ja osa elämästä, kuten mikro-organismit, voi olla adsorboituneessa tilassa hiukkasten pinnalla ohuissa kalvokosteuskerroksissa.
2. Mezobiota - tämä on yhdistelmä suhteellisen pieniä, helposti poistettavia maaperästä, liikkuvia eläimiä (maaperäiset nematodit, pienet hyönteisten toukat, punkit jne.). Maaperän mesobiota-edustajien koot ovat kymmenesosista 2–3 mm: iin. Tälle eläinryhmälle maaperä näyttää olevan pieni luolia. Heillä on erityisiä kaivuustyökaluja. Ne indeksoivat maaperän onteloiden seinämiä raajojen tai mutkittelevien matojen avulla. Vesihöyryllä kyllästetty maaperäilma antaa heille hengittää kehon rungon läpi. Eläimet kokevat ajanjaksoja, jolloin maaperä tulvii vedellä, yleensä ilmakuplina. Ilma viipyy vartaloidensa ympärillä, koska useimmissa niistä on hiuksilla ja vaa'oilla varustetun piirustuksen kostumaton vaikutus.
Meso- ja mikrobiotyyppiset eläimet voivat sietää maaperän talvella jäätymistä, mikä on erityisen tärkeää, koska suurin osa heistä ei pääse alas kerroksista, jotka ovat alttiina jäätymislämpötiloille.
3) Makrobiota - Nämä ovat suuria maaperäeläimiä, joiden vartalo on kooltaan 2 - 20 mm (hyönteisten toukat, millipedes, lieroja jne.). Se liikkuu maaperässä, laajentaen luonnollisia kaivoja levittämällä maahiukkasia tai kaivamalla uusia käytäviä. Molemmat liikkumismenetelmät jättävät jäljen eläinten ulkoiseen rakenteeseen. Useimpien tämän ryhmän lajien kaasunvaihto tapahtuu käyttämällä erikoistuneita hengityselimiä, mutta samalla sitä täydennetään kaasunvaihdolla liitoksen kautta.
Karjaavat eläimet voivat poistua kerroksista, jos epäsuotuisat olosuhteet syntyvät. Talven ja kuivuuden aikana ne keskittyvät syvempiin kerroksiin, useimmiten kymmeniä senttimetrejä pinnasta.
4) Megabiota - Nämä ovat suuria kaivauksia, pääasiassa nisäkkäiden keskuudesta. Monet heistä viettävät elämänsä maaperässä (kultahamot Afrikassa, myyrirotit, myyrät Euraasiassa, murupooliset myyrät Australiassa) .Jot tasoittavat maaperään kokonaisia \u200b\u200bkäytävien ja urien järjestelmiä. Sopeutuminen urheiluun maanalaiseen elämäntapaan heijastuu näiden eläinten ulkonäössä ja anatomisissa ominaisuuksissa: Heillä on alikehittyneet silmät, kompakti, turvonnut vartalo, jolla on lyhyt kaula, lyhyt paksu turkki, vahvat kompakti raajat, joilla on vahvat kynnet.
Maaperän pysyvien asukkaiden lisäksi suuret eläimet erottavat usein erillisen ekologisen ryhmän reiän asukkaat (mäyrät, metsänoskat, oravat, jerboat jne.). He syövät pinnalla, mutta kasvattavat, hibernatoivat, lepäävät ja pääsevät vaaroihin maaperässä.
Kävelemällä metsän tai niittyjen läpi, tuskin uskot olevanne ... sisällä maa-air. Mutta juuri tätä tutkijat kutsuvat elävien olentojen taloksi, jonka muodostaa maan ja ilman pinta. Kun ui joessa, järvessä tai merellä, löydät itsesi vesiympäristö - Toinen runsaasti asuttu luonnollinen koti. Ja kun autat aikuisia kaivaa maaperää puutarhassa, näet maaperän jalkojen alla. Asukkaita on myös monia erilaisia. Kyllä, ympärillämme on kolme upeaa taloa - kolme elinympäristöjohon useimpien planeettamme asuttavien organismien kohtalo on erottamattomasti sidoksissa toisiinsa.
Elämässä jokaisessa ympäristössä on omat piirteensä. maa-air tarpeeksi happea, mutta usein ei tarpeeksi kosteutta. Sitä on erityisen vähän stepillä ja aavikoilla. Siksi kuivilla alueilla olevilla kasveilla ja eläimillä on erityisiä laitteita veden uuttamiseen, varastointiin ja taloudelliseen käyttöön. Muista ainakin kaktus, joka tallentaa kosteutta vartaloosi. Ilman ympäristössä tapahtuu merkittäviä lämpötilan muutoksia, etenkin alueilla, joilla on kylmät talvet. Näillä alueilla ympäri vuoden organismien koko elämä muuttuu huomattavasti. Syksyn lehdet, muuttolintujen lento lämpimämpään ilmastossa, turkisten vaihtaminen eläimistä paksummiksi ja lämpimämmiksi - kaikki nämä ovat elävien olentojen mukautumisia luonnon kausivaihteluihin.
Missä tahansa ympäristössä elävien eläinten tärkeä ongelma on liikkuminen. Pohja-ilmaympäristössä voit liikkua maassa ja ilman läpi. Ja eläimet käyttävät sitä. Joidenkin jalat on sovitettu juoksemiseen (strutsi, gepardi, seepra), toisten - hyppäämiseen (kenguru, jerboa). Jokaisesta sadasta tässä ympäristössä elävästä eläinlajista 75 voi lentää. Tämä on suurin osa hyönteisistä, lintuista ja joistakin eläimistä (lepakoista).
vesiympäristö jotain, ja vettä on aina tarpeeksi. Lämpötila muuttuu tässä vähemmän kuin ilman lämpötila. Mutta happi ei usein riitä. Jotkut organismit, kuten taimenkalat, voivat elää vain happea sisältävässä vedessä. Toiset (karppi, ristikarppi, hamppu) kestävät hapen puutetta. Talvella, kun monet lammikot ketjuutetaan jään avulla, kalat voivat kuolla - niiden joukkokuolema tukehtumisesta. Joten happi tunkeutuu veteen, jäässä on reikiä.
Vesiympäristössä on vähemmän valoa kuin ilmassa. Valtamereissä ja merillä alle 200 m: n syvyydessä - hämärän valtakunta ja vielä alhaisemmassa ikuinen pimeys. On selvää, että vesikasveja löytyy vain sieltä, missä on riittävästi valoa. Vain eläimet voivat elää syvemmälle. He syövät "putoamisesta" eri meren asukkaiden kuolleiden jäännösten ylemmistä kerroksista.
Monien vesieläinten näkyvin piirre on uimavälineet. Kaloilla, delfiineillä ja valailla on evät. Puutarhoissa ja hylkeissä on läppä. Majavilla, saukkoilla, vesilinnuilla ja sammakoilla on kalvot sormensa välissä. Uimakuoriaisilla on airomaiset uimajalat.
Maaperän ympäristö - kotona monille bakteereille ja alkueläimille. Tässä ovat sieneli sienet, kasvien juuret. Erilaisia \u200b\u200beläimiä - matoja, hyönteisiä, kaivamiseen sovitettuja eläimiä, esimerkiksi mols - ovat myös asuttaneet maaperä. Maaperän asukkaat löytävät ympäristölle heille välttämättömät olosuhteet - ilma, vesi, mineraalisuolat. Totta, happea on vähemmän ja hiilidioksidia enemmän kuin raikkaassa ilmassa. Ja joskus vettä on liian paljon. Mutta lämpötila on tasaisempi kuin pinnalla. Mutta valo ei tunkeudu syvälle maaperään. Siksi eläimillä, jotka asuvat siinä, on yleensä hyvin pienet silmät tai heillä ei ole täysin näköelimiä. Heidän haju- ja kosketus auttaa.
Pohjailma
Näissä luvuissa erilaisten elinympäristöjen edustajat "tapasivat". Luonnossa he eivät pääse yhteen, koska monet heistä asuvat kaukana toisistaan, eri mantereilla, merillä, makeassa vedessä ...
Mestari lentojen nopeudessa lintujen keskuudessa - nopea. 120 km tunnissa on hänen tavanomainen nopeutensa.
Kolibri-linnut siipivät siipiään jopa 70 kertaa sekunnissa, hyttyset - jopa 600 kertaa sekunnissa.
Eri hyönteisten lentonopeus on seuraava: nauhoituksessa - 2 km tunnissa, kotikärpässä - 7, toukokuoriaiskuoressa - 11, kimalaisessa - 18 ja haarukkaperhosessa - 54 km tunnissa. Joidenkin havaintojen mukaan suuret sudenkorennot kehittävät nopeutta jopa 90 km tunnissa.
Lepakkomme ovat pienikokoisia. Kuumissa maissa heidän sukulaistensa elää siivekäisiä lintuja. Ne ovat 170 cm: n siipien ulottuvilla!
Suuret kengurut tekevät hyppyjä jopa 9 ja joskus jopa 12 metriin. (Mittaa tämä etäisyys luokassa lattialla ja kuvittele kenguruhyppy. Se on vain henkeäsalpaava!)
Gepardi on nopeinjalkainen eläin. Sen nopeus on jopa 110 km tunnissa. Strutsi voi ajaa jopa 70 km / h nopeudella 4-5 metrin askelin.
Vesiympäristö
Kalat ja rapu hengittävät kiduksilla. Nämä ovat erityisiä elimiä, jotka erottavat veteen siihen liuenneen hapen. Sammakko veden alla hengittää ihoa. Mutta vesiympäristön hallitsevat eläimet hengittävät kevyesti nouseen veden pinnalle inspiraatiota varten. Vesivirheet käyttäytyvät samalla tavalla. Vain heillä, kuten muilla hyönteisillä, ei ole keuhkoja, mutta erityisissä hengitysputkissa - henkitorvessa.

Maaperän ympäristö
Molin, zocorin ja myrkän kehon rakenne viittaa siihen, että ne kaikki ovat maaympäristön asukkaita. Moolin etujalat ja zocor ovat tärkeimmät kaivuustyökalut. Ne ovat litteitä, kuten lapioita, erittäin suurilla kynnillä. Ja myyrirotilla on normaalit jalat, se puree maahan voimakkailla etuhampailla (niin että maa ei pääse suuhun, huulet peittävät sen hampaiden takana!). Kaikkien näiden eläinten vartalo on soikea, kompakti. Tällaisella rungolla on kätevää liikkua maanalaisia \u200b\u200bkäytäviä pitkin.

Testaa tietosi
- Lista luontotyypit, jotka tapasit oppitunnissa.
- Mitkä ovat organismien elinolosuhteet ilma-maa-ympäristössä?
- Kuvaile vesiympäristön elinolosuhteita.
- Mitkä ovat maaperän ominaisuudet elinympäristönä?
- Anna esimerkkejä organismien sopeutumiskyvystä elämään eri ympäristöissä.
Think!
- Selitä, mitä kuvassa näkyy. Missä ympäristöissä luulet elävän eläimiä, joiden ruumiinosat on esitetty kuvassa? Voitko nimetä nämä eläimet?
- Miksi merellä elävät vain eläimet?

On maapallon ilman, veden ja maaperän elinympäristöjä. Jokainen organismi on sopeutunut elämään tietyssä ympäristössä.
Yleiset ominaisuudet. Maa-ilmaväliaine kehitettiin evoluution aikana paljon myöhemmin kuin vesiväline. Elämä maalla vaatii sellaisia \u200b\u200bmukautuksia, jotka olivat mahdollisia vain suhteellisen korkealla organisointitasolla sekä kasveille että eläimille. Pohja-ilma-elämän ympäristölle on ominaista, että täällä asuvia organismeja ympäröi ilma ja kaasumainen ympäristö, jolle on ominaista alhainen kosteus, tiheys ja paine, korkea happipitoisuus. Tämän ympäristön eläimet liikkuvat yleensä maaperän (kiinteän substraatin) ympärillä ja kasvit juurtuvat siihen.
Pohja-ilmaympäristön ympäristötekijöillä on useita ominaispiirteitä: suurempi valon voimakkuus verrattuna muihin väliaineisiin, merkittävät lämpötilanvaihtelut, kosteuden muutokset maantieteellisestä sijainnista, vuodenajasta ja vuorokaudenajasta riippuen (taulukko 5.3).
Taulukko 5.3
Ilman ja vesieliöiden elinolot
(mukaan D.F. Mordukhai-Boltovsky, 1974)
|
olosuhteet elinympäristö |
Edellytysten merkitys organismeille |
|
|
ilmaympäristö |
vesiympäristö |
|
|
kosteus |
Erittäin tärkeä (usein vajaa) |
Ei (aina enemmän) |
|
tiheys |
Pieni (pois lukien maaperä) |
Suuri verrattuna sen rooliin ilman asukkaiden kannalta |
|
paine |
Lähes ei |
Suuri (voi saavuttaa 1000 ilmakehän) |
|
lämpötila |
Merkittävä (vaihtelee erittäin suurissa rajoissa (-80 - +100 ° С ja enemmän) |
Pienempi kuin arvo asukkaille (vaihtelee paljon vähemmän, yleensä välillä -2 - + 40 ° C) |
|
happi |
Pieni (enimmäkseen yli) |
Välttämätöntä (usein vajaa) |
|
keskeytetty aineet |
merkityksetön; ei käytetä elintarvikkeissa (pääasiassa mineraaleja) |
Tärkeää (ruoan lähde, erityisesti orgaaninen aine) |
|
Ympäristössä liuenneet aineet |
Jossain määrin (merkityksellinen vain maaperän ratkaisuissa) |
Tärkeää (tietyn määrän välttämätöntä) |
Edellä mainittujen tekijöiden vaikutus liittyy erottamattomasti ilmamassien - tuulen - liikkeeseen. Evoluutioprosessissa pohja-ilmaympäristön elävät organismit kehittivät ominaisia \u200b\u200banatomisia, morfologisia, fysiologisia, käyttäytymiseen liittyviä ja muita mukautumisia. Esimerkiksi on ilmennyt elimiä, jotka tarjoavat ilmakehän hapen suoran assimilaation hengityksen aikana (eläinten keuhkot ja henkitorvi, kasvien vatsat). Luuston muodostumia (eläinten luuranko, kasvien mekaaniset ja tukikudokset), jotka tukevat vartaloa ympäristössä, jonka tiheys on alhainen, on kehitetty voimakkaasti. Mukautukset kehitettiin suojaamaan haitallisilta tekijöiltä, \u200b\u200bkuten elinkaaren jaksolta ja rytmiltä, \u200b\u200brakenteiden monimutkaiselta rakenteelta, lämmön säätelymekanismeilta jne. Maaperään (eläimen raajat, kasvin juuret) muodostui läheinen suhde, eläinten liikkuvuutta kehitettiin etsimään ruokaa ja ilmaan ilmaantuivat virtaukset kasvien siemenet, hedelmät ja siitepöly, lentävät eläimet.
Mieti tärkeimpien ympäristötekijöiden vaikutuksia kasveihin ja eläimiin maan ja ilman elämän ympäristössä.
Matala ilman tiheys määrittelee sen alhaisen nostovoiman ja merkityksettömän kiistanalaisen. Kaikki ilmaympäristön asukkaat ovat läheisesti yhteydessä maan pintaan, palvelevat heitä kiinnittymisessä ja tukemisessa. Ilman tiheydellä ei ole suurta kehon vastuskykyä, kun ne liikkuvat maan pinnalla, mutta vaikeuttaa liikkumista pystysuunnassa. Useimmille organismeille ilmaan jääminen liittyy vain uudelleensijoittamiseen tai saaliin etsimiseen.
Matala ilmahissi määrittää maanpäällisten organismien lopullisen massan ja koon. Suurimmat maan pinnalla olevat eläimet ovat pienempiä kuin vesiympäristön jättiläiset. Suuret nisäkkäät (nykyaikaisen valaan koko ja paino) eivät voineet elää maalla, koska ne murskataan oman painonsa avulla. Jättiläiset mesozoiset lisat johtivat puolivettäiseen elämäntapaan. Toinen esimerkki: Korkeissa, pystyssä olevissa sekvoiakasveissa (Sequoja sempervirens), joiden leveys on 100 metriä, on voimakas tukipuu, kun taas jättiläismäisten ruskelevien Macrocystis-talien thalissa, joka kasvaa jopa 50 metriä, mekaaniset elementit ovat vain erittäin heikosti eristetty talon ytimessä.
Ilman alhainen tiheys luo vähän vastustuskykyä liikkumiselle. Monet maa-eläimet ovat käyttäneet evoluution aikana tämän ilmaympäristön ominaisuuden ympäristöhyötyjä saavuttaen lentämiskyvyn. 75% kaikentyyppisistä maaeläimistä kykenee aktiiviseen lentoon. Nämä ovat pääosin hyönteisiä ja lintuja, mutta löytyy myös nisäkkäitä ja matelijoita. Maalliset eläimet lentävät pääasiassa lihaksen avulla. Jotkut eläimet voivat myös suunnitella ilmavirran vuoksi.
Alemmassa ilmakehässä olevan ilman liikkuvuuden vuoksi ilmamassien pystysuoraan ja vaakasuoraan liikkumiseen on mahdollista kehittää tiettyjen eliölajien passiivinen lento anemochoria - uudelleensijoittaminen ilmavirtojen avulla. Ilmavirtojen passiivisesti kuljettamia organismeja kutsutaan yhdessä aeroplankton, analogisesti vesiympäristön planktonisten asukkaiden kanssa. Passiiviselle lennolle N.M. Chernova, A.M. Bylova (1988) -organismeilla on erityisiä mukautuksia - pienet vartalokoot, pinta-alan suureneminen johtuen uloskasvuista, vahva hajoaminen, siipien suuri suhteellinen pinta, rainan käyttö jne.
Kasvien anemokorisilla siemenillä ja hedelmillä on myös erittäin pienikokoisia (esimerkiksi tulileväsiemeniä) tai erilaisia \u200b\u200bpterygoideja (Acer pseudoplatanum vaahtera) ja laskuvarjojen muotoisia (Taraxacum officinale voikukka) lisäyksiä
Tuulen pölyttämissä kasveissa on useita laitteita, jotka parantavat siitepölyn aerodynaamisia ominaisuuksia. Niiden kukkapeite on yleensä pienentynyt, ja portaat eivät ole suojattu tuulelta.
Kasvien, eläinten ja mikro-organismien asutuksessa pääroolia ovat vertikaaliset perinteiset ilmavirrat ja heikko tuuli. Myrskillä ja hurrikaaneilla on myös merkittävä ympäristövaikutus maaperäisiin organismeihin. Melko usein voimakkaat tuulet, etenkin samaan suuntaan puhaltavat tuulet, taivuttavat puun oksia, taipuvat kääntöpuoli ja aiheuttavat lipunmuotoisia kruunun muotoja.
Alueilla, joilla voimakkaat tuulet puhaltavat jatkuvasti, pienten lentävien eläinten lajien koostumus on yleensä heikko, koska ne eivät pysty vastustamaan voimakkaita ilmavirtoja. Joten hunaja mehiläinen lentää vain tuulen voimakkuudella jopa 7–8 m / s, ja kirvoja lentävät erittäin heikolla tuulella, joka ei ylitä 2,2 m / s. Näiden paikkojen eläimillä on tiheät peitteet, jotka suojaavat vartaloa jäähdytykseltä ja kosteuden menetykseltä. Merellisillä saarilla, joissa on jatkuvaa voimakasta tuulta, linnuilla ja etenkin hyönteisillä, jotka ovat menettäneet lentämiskyvyn, ei ole siipiä, koska ne, jotka pystyvät lentämään ilmassa, räjäyttävät meren ja kuolevat.
Tuuli aiheuttaa muutoksen kasvien imeytymisnopeudessa ja on erityisen voimakasta kuivassa tuulessa, joka kuivaa ilmaa, mikä voi johtaa kasvien kuolemaan. Vaakasuorien ilmanvaihdosten (tuulet) pääasiallinen ekologinen merkitys on epäsuora ja muodostuu sellaisten tärkeiden ympäristötekijöiden kuin lämpötilan ja kosteuden vaikutuksen vahvistamisesta tai heikentämisestä maanpäällisille organismeille. Tuulet lisäävät kosteuden ja lämmön palautumista eläimiin ja kasveihin.
Tuulen lämpö on siedettävämpää ja vaikeampaa - pakkaset, nopeampi organismien kuivuminen ja jäähtyminen.
Maalaisorganismeja esiintyy suhteellisen alhaisessa paineessa, mikä johtuu ilman alhaisesta tiheydestä. Kaiken kaikkiaan maaperäiset organismit ovat enemmän stenoboituja kuin vesieliöt, koska niiden ympäristössä tavalliset paineenvaihtelut ovat ilmakehän murto-osia, ja esimerkiksi suurelle korkeudelle nouseville lintuille ne eivät ylitä yhtä kolmasosaa normaalista.
Ilman kaasukoostumuskuten aikaisemmin ajateltiin, ilmakehän pintakerroksessa se on melko monotoninen (happi - 20,9%, typpi - 78,1%, m. kaasumaiset kaasut - 1%, hiilidioksidi - 0,03 tilavuusprosenttia) johtuen korkeasta diffundiostaan konvektion ja tuulen virtausten kyky ja jatkuva sekoittuminen. Samanaikaisesti paikallisista lähteistä ilmakehään saapuvien kaasumaisten, pisaranesteiden ja pöly (kiinteiden) hiukkasten epäpuhtauksilla on usein merkittävä ympäristövaikutus.
Happi, koska se sisältää jatkuvasti korkean pitoisuuden ilmassa, ei ole rajoittava tekijä maanpäällisessä ympäristössä. Korkea happipitoisuus osaltaan lisäsi aineenvaihduntaa maanpäällisissä organismeissa, ja hapettumisprosessien suuren tehokkuuden perusteella syntyi eläinten homoyotermia. Vain paikoissa, tietyissä olosuhteissa, syntyy väliaikainen hapenpuute, esimerkiksi rappeutuvissa kasvinjäännöksissä, viljavarastoissa, jauhoissa jne.
Joillakin pinta-ilmankerroksen alueilla hiilidioksidipitoisuus voi vaihdella melko merkittävissä rajoissa. Joten jos suurissa teollisuuskeskuksissa ja kaupungeissa ei ole tuulta, sen pitoisuus voi kasvaa kymmeniä kertoja.
Pintakerrosten hiilihappopitoisuuden päivittäiset muutokset johtuvat kasvien fotosynteesin rytmistä (kuva 5.17).
Kuva 5.17. Päivittäiset muutokset pystysuunnassa
hiilidioksidipitoisuus metsäilmassa (valmistaja W. Larcher, 1978)
Esimerkiksi metsäilman hiilidioksidipitoisuuden vertikaalisen profiilin päivittäisistä muutoksista osoitetaan, että päivällä hiilidioksidi kuluu fotosynteesiin puiden kruunujen tasolla ja tuulen puuttuessa hiilidioksidin huono alue (305 ppm), joka vastaanottaa CO ilmakehästä ja maaperä (maaperän hengitys). Yöllä perustetaan ilman vakaa kerrostuminen lisääntyneellä hiilidioksidipitoisuudella maakerroksessa. Hiilidioksidin kausivaihtelu liittyy muutoksiin elävien organismien, useimpien maaperän mikro-organismien, hengitysnopeudessa.
Korkeissa pitoisuuksissa hiilidioksidi on myrkyllistä, mutta luonteeltaan sellaiset pitoisuudet ovat harvinaisia. Alhainen hiilidioksidipitoisuus estää fotosynteesiä. Kasvihuoneiden ja kasvihuoneiden (suljetussa maassa) fotosynteesin nopeuden lisäämiseksi hiilidioksidipitoisuutta lisätään usein keinotekoisesti.
Suurimmalle osalle maanpäällisestä ympäristöstä ilmatyppi on inertti kaasu, mutta mikro-organismeilla, kuten kyhmybakteerit, atsotobakteerit, klostridiat, on kyky sitoa sitä ja osallistua biologiseen kiertoon.
Tärkein nykyaikainen ilmakehän fysikaalisen ja kemiallisen pilaantumisen lähde on ihmisen toiminta: teollisuus- ja kuljetusyritykset, maaperän eroosio jne. Rikkidioksidi on siis myrkyllistä kasveille pitoisuuksilla, jotka ovat viidenkymmenen tuhannesosan miljoonasta miljoonasosaan ilman tilavuudesta. Jäkälät kuolevat jo rikkidioksidin jäämien perusteella ympäristössä. Siksi erityisen herkkiä kasveja SO 2: lle käytetään usein ilmaisun pitoisuuteen. Savulle herkkiä ovat tavallinen kuusen ja männyn, vaahteran, pärnun, koivun.
Valotila. Maapallon päästävän säteilyn määrä johtuu maaston maantieteellisestä leveydestä, päivän pituudesta, ilmakehän läpinäkyvyydestä ja auringonvalon esiintymiskulmasta. Eri sääolosuhteissa 42–70% aurinkovakiosta saavuttaa maanpinnan. Auringon säteily, joka kulkee ilmakehän läpi, on muuttunut paitsi määrällisesti myös koostumuksessa. Lyhytaaltoinen säteily absorboituu otsoniseulaan ja ilmakehän happea. Infrapunasäteet absorboivat ilmakehään vesihöyry ja hiilidioksidi. Loput suoran tai sironneen säteilyn muodossa saavuttavat maapallon pinnan.
Suoran ja hajanaisen aurinkosäteilyn yhdistelmä on 7 - 7 “kokonaissäteilystä, kun pilvisinä päivinä hajakuormitus on 100%. Korkeilla leveysasteilla hajollinen säteily on vallitsevaa, kun taas tropiikissa - suora. Hajanainen säteily keskipäivällä sisältää kelta-punaista säteilyä jopa 80%, suora - 30–40%. Selkeinä aurinkoisina päivinä maan pinnalle päästävä auringonsäteily koostuu 45% näkyvästä valosta (380–720 nm) ja 45% infrapunasäteilystä. Vain 10% on ultraviolettisäteilyä. Ilmakehän pölyllä on merkittävä vaikutus säteilytilaan. Joissakin kaupungeissa pilaantumisen takia valaistus voi olla 15% tai vähemmän kaupungin ulkopuolelta.
Valaistus maan pinnalla vaihtelee suuresti. Kaikki riippuu auringon korkeudesta horisontin yläpuolella tai auringonvalon esiintymiskulmasta, päivän pituudesta ja sääolosuhteista, ilmakehän läpinäkyvyydestä (kuva 5.18).

Kuva 5.18. Auringonsäteilyn jakauma riippuen
auringonkorkeudet horisontin yläpuolella (A 1 - korkea, A 2 - matala)
Vuodenajasta ja vuorokaudenajasta riippuen myös valon voimakkuus vaihtelee. Joillakin maapallon alueilla valon laatu ei ole sama, esimerkiksi pitkäaaltosäteen (punainen) ja lyhytaalto (sininen ja ultravioletti) säteiden suhde. Lyhytaaltoisten säteiden tiedetään olevan suurempia kuin pitkäaaltosäteiden, ilmakehän absorboimat ja hajottamat. Siksi vuoristoisilla alueilla on aina enemmän lyhytaaltoaallon säteilyä.
Puut, pensaat, kasvikasvit peittävät maaston, luovat erityisen mikroilmaston, heikentäen säteilyä (kuva 5.19).

Kuva 5.19. Säteilyn heikkeneminen:
A - harvinaisessa mäntymetsässä; B - maissikasveissa Tulevasta fotosynteettisesti aktiivisesta säteilystä 6–12% heijastaa (R) istutuksen pinnasta
Siten erilaisissa luontotyypeissä säteilyvoimakkuuden lisäksi ei ole vain spektrikoostumusta, kasvien valaistuksen kestoa, eri intensiteetin valon alueellista ja ajallista jakautumista jne. Vastaavasti organismien mukautukset elämään maaympäristössä yhden tai toisen valomoodin alla vaihtelevat. . Kuten aiemmin totesimme, valossa on kolme pääryhmää kasveja: photophilic (Heliophyte) sävy (sciosyytit) ja varjoa sietävät. Fotofiiliset ja varjoa rakastavat kasvit eroavat toisistaan \u200b\u200bekologisen optimaalisen aseman suhteen.
Fotofiilisissä kasveissa se sijaitsee täyden auringonvalon kentällä. Vahva varjostus vaikuttaa heihin masentavasti. Ne ovat avoimen maan tai hyvin valaistujen arojen ja niittyjen ruohoja (ruohokerroksen ylempi kerros), kallio jäkälät, lehtimetsien varhaiskevään ruohokasvit, eniten viljeltyjä avomaan ja rikkaruohojen kasveja jne. Varjostusta rakastavilla kasveilla on optimi heikossa valaistuksessa, eivätkä ne voi seistä. voimakas valo. Nämä ovat pääasiassa monimutkaisten kasviyhteisöjen alempia varjostettuja kerroksia, joissa varjoaminen on seurausta korkeampien kasvien ja asukkaiden "valloittamasta" valosta. Tähän sisältyy monia sisä- ja kasvihuonekasveja. Suurimmaksi osaksi ne tulevat trooppisten metsien nurmettuneesta peitteestä tai epifyyttisestä kasvistosta.
Ympäristökäyrä suhtautuminen valoon ja varjoa sietävään on hieman epäsymmetrinen, koska ne kasvavat ja kehittyvät paremmin täydessä valossa, mutta sopeutuvat hyvin heikkoon valoon. Tämä on yleinen ja erittäin muovinen kasviryhmä maanpäällisessä ympäristössä.
Pohja-ilmaympäristön kasveissa on kehittynyt mukautuksia valojärjestelmän erilaisiin olosuhteisiin: anatomisiin, morfologisiin, fysiologisiin jne.
Hyvä esimerkki anatomisista ja morfologisista mukautuksista on muutos ulkomuodossa erilaisissa valaistusolosuhteissa, esimerkiksi kasvien lehtiterien epätasainen koko, joka johtuu systemaattisesta sijainnista, mutta jotka elävät erilaisissa valaistusolosuhteissa (niittykello - Campanula patula ja metsänkello - C. trakelium, peltovioletti - Viola) arvensis, kasvaa pelloilla, niittyillä, reunoilla ja metsävioletteilla - V. mirabilis), kuva. 5.20.

Kuva 5.20. Lehden koon jakautuminen olosuhteiden mukaan
elinympäristökasvit: märästä kuivaan ja varjostetusta aurinkoiseen
Huom. Varjostettu alue vastaa luonnossa vallitsevia olosuhteita.
Liiallisissa olosuhteissa ja valon puutteessa lehtiterien sijoittelu kasvien avaruudessa vaihtelee huomattavasti. Helioyyttikasveissa lehdet ovat suuntautuneet vähentämään säteilyn saapumista kaikkein vaarallisimpiin päiväaikoihin. Lehtilavat sijaitsevat pystysuorassa tai suuressa kulmassa vaakatasoon nähden, joten päivän aikana lehdet saavat enimmäkseen liukuvat säteet (kuva 5.21).
Tämä on erityisen voimakasta monissa steppikasveissa. Mukauttaminen niin sanottujen "kompassi" kasvien (luonnonvarainen salaatti - Lactuca serriola jne.) Saadun säteilyn vaimenemiseen on mielenkiintoista. Luonnonvaraisen salaatin lehdet sijaitsevat samassa tasossa, suunta pohjoisesta etelään, ja keskipäivällä säteily lehden pintaan on minimaalinen.
Varjossa sietävissä kasveissa lehdet on järjestetty saamaan maksimaalinen määrä säteilyä.
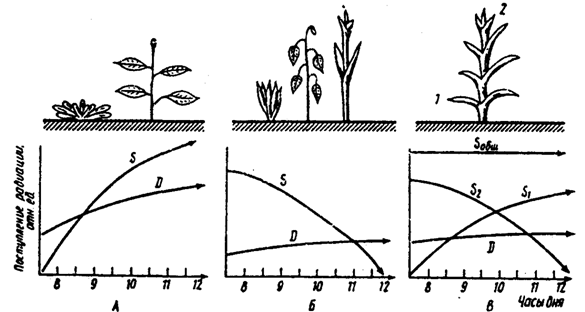
Kuva 5.21. Suoran (S) ja hajaantuneen (D) aurinkosäteilyn saapuminen kasveille, joilla on vaakasuorat (A), pystysuorat (B) ja eri suuntaan suuntautuvat (C) lehdet (I. A. Shulginin mukaan, 1967)
1,2 - lehdet, joilla on erilaiset kallistuskulmat; S 1, S 2 - suoran säteilyn vastaanottaminen heille; S yhteensä - sen kokonaisvirtaus kasviin
Varjostusta kestävät kasvit kykenevät usein suojaamaan liikkeitä: lehtien siipien asennon muutos, kun voimakas valo osuu niihin. Ruohon peittoalueet, joissa happamahapon taitetut lehdet ovat verrattain tarkalleen samoin kuin suurten aurinkosähkön sijainti. Arkin rakenteessa voidaan havaita useita mukautuvia piirteitä aurinkosäteilyn päävastaanottimeksi. Esimerkiksi, monissa heliofyyteissä lehden pinta myötävaikuttaa auringonvalon heijastumiseen (kiiltävä - laakereilla, peitetty vaalealla karvaisella pinnoitteella - kaktus, maitoherkät) tai heikentäen niiden toimintaa (paksu kynsinauha, tiheä karvainen). Lehmän sisäiselle rakenteelle on tunnusomaista palisadekudoksen voimakas kehitys, lukuisten pienten ja kevyiden kloroplastien läsnäolo (kuva 5.22).
Yksi kloroplastien suojareaktioista liialliseen valoon on niiden kyky muuttaa suuntaa ja liikkua solussa, lausutaan kevyissä kasveissa.
Kirkkaassa valossa kloroplastit vievät posturaalisen sijainnin solussa ja niistä tulee "reuna" säteiden suuntaan. Heikossa valossa ne jakautuvat hajanaisesti kennoon tai kertyvät sen alaosaan.
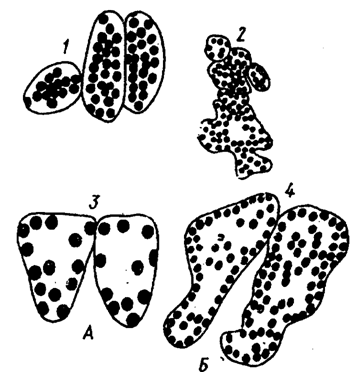
Kuva 5.22. Klooroplastien erilaiset arvot sietokyvyssä
(A) ja valofiiliset (B) kasvit:
1 - marjakuusi; 2 - lehtikuusi; 3 - kavio; 4 - jousipuhdas (kirjoittanut T.K. Goryshina, E.G. Pruzhina, 1978)
Fysiologiset sopeutumiset kasvit ilma-ilma-ympäristön valaistusolosuhteisiin kattavat useita elintärkeitä toimintoja. On todettu, että fotofiilisissä kasveissa kasvuprosessit reagoivat herkemmin valon puutteeseen verrattuna varjoihin. Seurauksena on varren lisääntynyt venytys, joka auttaa kasveja murtautumaan valoon kasviyhteisöjen ylemmissä kerroksissa.
Tärkeimmät fysiologiset mukautukset valoon ovat fotosynteesin kentässä. Yleisessä muodossa fotosynteesin muutos valon voimakkuudesta riippuen ilmaistaan \u200b\u200b”fotosynteesin valokäyrällä”. Seuraavat parametrit ovat ympäristölle tärkeitä (kuva 5.23).
1. Käyrän leikkauspiste ordinaattiakselin kanssa (kuva 5.23, a) vastaa kasvien kokoa ja kaasunvaihdon suuntaa täydellisessä pimeydessä: fotosynteesi puuttuu, hengitys tapahtuu (ei imeytyminen, mutta hiilidioksidipäästö), joten kohta a on abskissa-akselin alapuolella.
2. Valokäyrän ja abskissa-akselin leikkauspiste (kuva 5.23, b) luonnehtii ”kompensointipistettä”, toisin sanoen valon voimakkuutta, jolla fotosynteesi (CO 2: n imeytyminen) tasapainottaa hengitystä (CO 2 -päästö).
3. Fotosynteesin intensiteetti valon lisääntyessä kasvaa vain tiettyyn rajaan, tulevaisuudessa se pysyy vakiona - fotosynteesin valokäyrä saavuttaa "kylläisyyden tasangon".
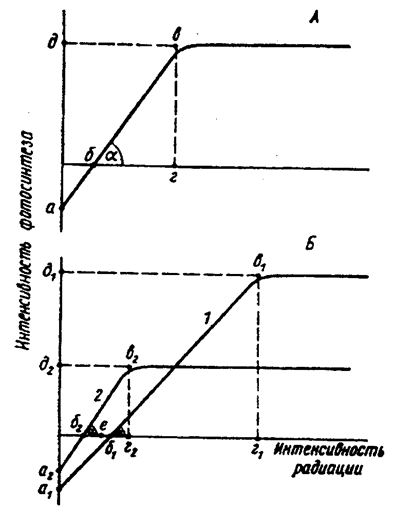
Kuva 5.23. Fotosynteesin vaaleat käyrät:
A on yleinen ääriviiva; B - käyrät fotofiilisille (1) ja varjoa sietäville (2) kasveille
Kuvassa 1 5.23 taivutusalue ilmaistaan \u200b\u200bmielivaltaisesti tasaisella käyrällä, jonka murtuma vastaa pistettä vuonna. Pisteen b projektio abskissa-akselille (piste r) karakterisoi valon “tyydyttynyttä” intensiteettiä, toisin sanoen arvoa, jonka yläpuolella valo ei enää lisää fotosynteesin voimakkuutta. Projektio ordinaattiakselilla (piste e) vastaa tietyn lajin korkeinta fotosynteesinopeutta tietyssä ilmaympäristössä.
4. Valokäyrän tärkeä ominaisuus on kaltevuuskulma (a) abskissaan, mikä heijastaa fotosynteesin lisääntymisastetta säteilyn lisääntyessä (suhteellisen alhaisen valon voimakkuuden alueella).
Kasveissa havaitaan valoreaktion kausittainen dynamiikka. Esimerkiksi karvaisen (Carex pilosa) varhaiskeväällä metsässä hiljattain ilmestyneillä lehdillä on valon kyllästymisen taso 20–25 tuhatta luksia, näissä lajeissa kesällä varjostettaessa fotosynteesin riippuvuus käyristä vastaa ”varjo” -parametreja, t Toisin sanoen lehdet saavat kyvyn käyttää heikkoa valoa tehokkaammin, samat lehdet talvittamisen jälkeen lehdetön kevään metsän katos alla osoittavat taas fotosynteesin ”kevyitä” piirteitä.
Eräs erityinen muoto fysiologisesta sopeutumisesta, jossa on terävää valon puutetta, on kasvin menettämä fotosynteesi, siirtyminen heterotrofiseen ravintoon valmiiden orgaanisten aineiden avulla. Joskus sellaisesta siirtymästä tuli peruuttamaton, koska kasvit menettivät klorofylliä, esimerkiksi varjoisien kuusimetsien orkideat (Goodyera repens, Weottia nidus avis) ja Cyclarum (Monotropa hypopitys). Ne elävät kuolleista orgaanisista aineista puulajeista ja muista kasveista. Tätä ravitsemustapaa kutsutaan saprofyyttiseksi ja kasveja saprofyyttejä.
Suurimmalle osalle maalla eläviä eläimiä, joilla on päivä- ja yöaktiviteetteja, visio on yksi suuntausmenetelmistä, se on tärkeä saaliin etsinnälle. Monilla eläinlajeilla on myös värinäkö. Tässä suhteessa eläimillä, etenkin uhreilla, on mukautuvat piirteet. Näitä ovat suojaava, peittävä ja varoittava värjäys, suojaava samankaltaisuus, matkiminen jne. Korkeampien kasvien kirkkaanväristen kukinten esiintyminen liittyy myös pölyttäjien visuaalisen laitteen erityispiirteisiin ja viime kädessä ympäristön valojärjestelmään.
Vesitila. Kosteuden puute on yksi maanpäällisen ilmaympäristön merkittävimmistä piirteistä. Maaperäisten organismien kehitys tapahtui sopeutumalla kosteuden tuottamiseen ja säilyttämiseen. Maan kosteusmuodot ovat erilaisia \u200b\u200b- ainakin ilman täydellisestä ja jatkuvasta kyllästymisestä vesihöyryllä, jolloin useita tuhansia millimereitä sateita (päiväntasaajan ja monsuunitrooppisen ilmaston alueet) putoavat melkein täydelliseen puutteeseen kuivassa autiomaassa. Siksi trooppisissa autiomaissa keskimääräinen vuotuinen sademäärä on vähemmän kuin 100 mm vuodessa, ja sade ei kuulu joka vuosi.
Vuotuinen sademäärä ei aina anna mahdolliseksi arvioida organismien vesivarantoa, koska sama sademäärä voi olla ominaista autiomaalle (subtroopilla) ja erittäin kostealle (arktiselle alueelle). Suuri rooli on saostumisen ja haihtumisen suhteella (vuotuinen kokonaishaihtuminen vapaalta veden pinnalta), joka vaihtelee myös maapallon eri alueilla. Alueita, joilla tämä arvo ylittää vuotuiset sademäärät, kutsutaan hedelmätön (kuiva, kuiva). Esimerkiksi kasveista puuttuu kosteutta suurimman osan kasvukaudesta. Alueita, joilla kasveilla on kosteutta, kutsutaan kostea, tai märkä. Usein eristetyt siirtymävyöhykkeet - puolikuiva (Semiarid).
Kasvillisuuden riippuvuus vuotuisesta keskimääräisestä sademäärästä ja lämpötilasta on esitetty kuvassa. 5.24.
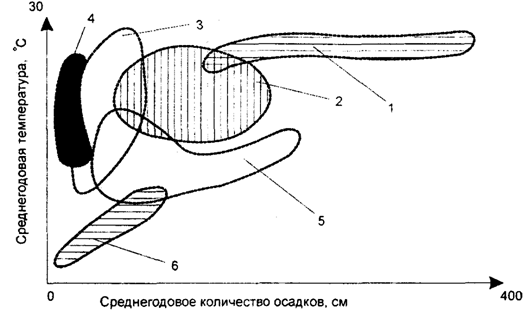
Kuva 5.24. Kasvillisuuden riippuvuus keskimääräisestä vuositasosta
sademäärä ja lämpötila:
1 - trooppinen metsä; 2 - lehtimetsä; 3 - steppi;
4 - autiomaa; 5 - havumetsät; 6 - arktinen ja vuoristo tundra
Maalla olevien organismien vedenjakelu riippuu sademoodista, vesistöjen läsnäolosta, maaperän kosteusvarannoista, pohjaveden läheisyydestä jne. Tämä on myötävaikuttanut monien sopeutumisten kehittämiseen maaperäisten organismien erilaisiin vesihuoltojärjestelmiin.
Kuvassa 1 Kuva 5.25 vasemmalta oikealle osoittaa siirtymisen alemmista vedessä elävistä lehdistä, joissa soluissa ei ole tyhjiöitä, primaarisiin poikilohydrisiin maanpäällisiin leviin, tyhjiöiden muodostumisesta vesivihreissä ja hiilevälehdissä, siirtymä tyhjiöitä sisältävistä talofyytteistä homohydrisiin forageofyyteihin (sammal - hydrofyyttien jakautuminen on edelleen rajoitettu elinympäristöihin, joissa on korkea ilmatila) , kuivissa elinympäristöissä sammalista tulee toinen poikyylihydrinen); saniaisia \u200b\u200bja angiospermejä (mutta ei kuntosolujen keskuudessa) on myös toisia poikilohydrisiä muotoja. Suurin osa lehtikasveista on homohydrisiä johtuen niiden kutikulaarisesta suojasta transpiraatiota ja solujensa voimakkaan imuroinnin vuoksi. On huomattava, että eläinten ja kasvien kserofilismi on ominaista vain pohja-ilmaympäristölle.
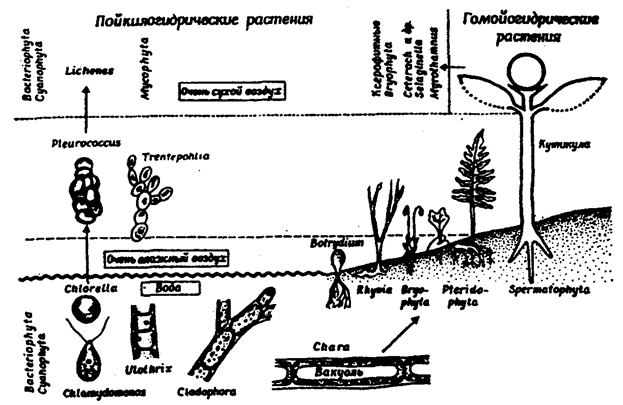
Kuva 5.25. Kasvien vedenvaihdon mukauttaminen maahan
elämäntapa (W. Larhera, 1978)
Sademäärä (sade, rake, lumi) vedenjakelun ja kosteusreservien muodostumisen lisäksi on usein toinen ympäristörooli. Esimerkiksi rankkasateiden aikana maaperällä ei ole aikaa imeä kosteutta, vesi virtaa nopeasti voimakkaisiin puroihin ja puhaltaa usein heikosti juurtuneita kasveja, pieniä eläimiä ja hedelmällistä maakerrosta järviin ja jokiin. Tulva-alueilla sateet voivat aiheuttaa tulvia ja siten vaikuttaa haitallisesti täällä eläviin kasveihin ja eläimiin. Määräajoin tulvissa paikoissa muodostuu ominainen tulva-eläimistö ja kasvisto.
Rakeella on negatiivinen vaikutus kasveihin ja eläimiin. Yksittäisten peltojen kasvustot tuhoutuvat joskus kokonaan tämän luonnonmullistuksen vuoksi.
Lumipeitteen ekologinen rooli on monimuotoinen. Kasveille, joiden uudistumispumput sijaitsevat maaperässä tai lähellä sen pintaa, ja monille pienille eläimille, lumi on lämmöneristävä peite, joka suojaa sitä matalilta talvelämpötiloilta. Jos pakkaset ovat yli -14 ° C, 20 cm lumenkerroksen alla, maaperän lämpötila ei laske alle 0,2 ° C. Syvä lumipeite suojaa kasvien vihreiden osien, kuten Veronica officinaliksen, kavioheinän jne. Jäätymiseltä, jotka menevät lumen alle pudottamatta lehtiä. Pienet maaeläimet elävät aktiivista elämäntapaa talvella, asettamalla lumen alle ja sen paksuudessa lukuisia gallerioita käytäviä. Lumisten talvien väkevöityjen ruokien läsnä ollessa jyrsijät (metsä- ja keltakaareiset hiiret, joukko ääniä, vesirotit jne.) Voivat siellä kasvaa. Hanna, puoliväri ja musta siru piiloutuvat lumen alla voimakkaiden pakkasten aikana.
Suurille eläimille talvinen lumipeite häiritsee usein ruuan hankkimista, etenemistä, kun pinnalle muodostuu jääkuori. Joten hirvi (Alces alces) ylittää vapaasti lumikerroksen, jonka syvyys on jopa 50 cm, mutta sitä ei ole saatavana pienemmille eläimille. Usein lumisten talvien aikana havaitaan mätijen ja villisikojen kuolemaa.
Voimakkaalla lumisateella on negatiivinen vaikutus kasveihin. Lumenmurtajien tai lumipallojen muodossa olevien mekaanisten vaurioiden lisäksi paksu lumikerros voi aiheuttaa kasvien lämpenemisen ja lumen sulamisen aikana, etenkin pitkittyneellä keväällä, kasvien kastelua.
 Kasvit ja eläimet kärsivät alhaisista lämpötiloista voimakkaiden tuulien aikana kevyinä lumisina talvina. Joten vuosina, jolloin lunta on vähän, hiiren kaltaiset jyrsijät, myyrät ja muut pienet eläimet kuolevat. Samaan aikaan leveysasteilla, joilla lumisateet putoavat talvella, kasvit ja eläimet ovat historiallisesti sopeutuneet elämään lumissa tai sen pinnalla, kun he ovat kehittäneet erilaisia \u200b\u200banatomisia, morfologisia, fysiologisia, käyttäytymiseen liittyviä ja muita ominaisuuksia. Esimerkiksi joillakin eläimillä jalkojen tukipinta kasvaa talvella likaamalla ne jäykillä karvilla (kuva 5.26), höyhenillä ja kiimaisilla kilpeillä.
Kasvit ja eläimet kärsivät alhaisista lämpötiloista voimakkaiden tuulien aikana kevyinä lumisina talvina. Joten vuosina, jolloin lunta on vähän, hiiren kaltaiset jyrsijät, myyrät ja muut pienet eläimet kuolevat. Samaan aikaan leveysasteilla, joilla lumisateet putoavat talvella, kasvit ja eläimet ovat historiallisesti sopeutuneet elämään lumissa tai sen pinnalla, kun he ovat kehittäneet erilaisia \u200b\u200banatomisia, morfologisia, fysiologisia, käyttäytymiseen liittyviä ja muita ominaisuuksia. Esimerkiksi joillakin eläimillä jalkojen tukipinta kasvaa talvella likaamalla ne jäykillä karvilla (kuva 5.26), höyhenillä ja kiimaisilla kilpeillä.
Toiset muuttuvat tai joutuvat inaktiiviseen tilaan - nukkumaan, lepotilaan, vaihteluun. Useat eläimet siirtyvät tietyntyyppisiin ruokiin.
 Lumikatteen valkoisuus paljastaa tummat eläimet. Valkoisten ja tundrankuoriaisten, ermiinin (kuva 5.27), valkoisen jänisen, lusikan, arktisen kettu, vuodenajan värinmuutos liittyy epäilemättä taustavärin peittämiseen.
Lumikatteen valkoisuus paljastaa tummat eläimet. Valkoisten ja tundrankuoriaisten, ermiinin (kuva 5.27), valkoisen jänisen, lusikan, arktisen kettu, vuodenajan värinmuutos liittyy epäilemättä taustavärin peittämiseen.
Saostus, sen välittömän vaikutuksen lisäksi organismeihin, määrää yhden tai toisen ilmankosteuden, jolla, kuten jo todettiin, on tärkeä rooli kasvien ja eläinten elämässä, koska se vaikuttaa niiden veden aineenvaihdunnan voimakkuuteen. Haihtuminen eläimen kehon pinnalta ja imeytyminen kasveihin etenevät intensiivisemmin, sitä vähemmän ilma on kyllästynyt vesihöyryyn.
 Sateen muodossa laskevan tippuvan nestemäisen kosteuden ja ilman höyryisen kosteuden imeytyminen maapallon yläosiin tapahtuu trooppisten metsien epifyyteissä, jotka imevät kosteuden koko lehtien ja ilmajuurten pinnalle. Ilman kosteushöyryt voivat absorboida joidenkin pensaiden ja puiden oksat, esimerkiksi saxaul - Halaxylon persicum, H. aphyllum. Korkeammissa itiöissä ja etenkin alemmissa kasveissa kosteuden imeytyminen maanpäällisistä osista on tavanomainen vedenjakelumenetelmä (sammalit, jäkälät jne.). Koska sammalissa ei ole kosteutta, jäkälät pystyvät selviytymään pitkään tilassa, joka on lähellä ilmakuivia ja joutuneen keskeytettyyn animaatioon. Mutta sateen arvoinen, sillä nämä kasvit imevät nopeasti kosteuden kaikissa maanosissa, muuttuvat pehmeiksi, palauttavat turgorin ja jatkavat fotosynteesin ja kasvun prosesseja.
Sateen muodossa laskevan tippuvan nestemäisen kosteuden ja ilman höyryisen kosteuden imeytyminen maapallon yläosiin tapahtuu trooppisten metsien epifyyteissä, jotka imevät kosteuden koko lehtien ja ilmajuurten pinnalle. Ilman kosteushöyryt voivat absorboida joidenkin pensaiden ja puiden oksat, esimerkiksi saxaul - Halaxylon persicum, H. aphyllum. Korkeammissa itiöissä ja etenkin alemmissa kasveissa kosteuden imeytyminen maanpäällisistä osista on tavanomainen vedenjakelumenetelmä (sammalit, jäkälät jne.). Koska sammalissa ei ole kosteutta, jäkälät pystyvät selviytymään pitkään tilassa, joka on lähellä ilmakuivia ja joutuneen keskeytettyyn animaatioon. Mutta sateen arvoinen, sillä nämä kasvit imevät nopeasti kosteuden kaikissa maanosissa, muuttuvat pehmeiksi, palauttavat turgorin ja jatkavat fotosynteesin ja kasvun prosesseja.
Hyvin kostutettujen maan luontotyyppien kasvien on usein poistettava ylimääräinen kosteus. Tämä tapahtuu pääsääntöisesti, kun maaperä on lämmitetty hyvin ja juuret imevät aktiivisesti vettä, eikä siellä ole verenvuotoa (aamulla tai sumussa, kun kosteus on 100%).
Ylimääräinen kosteus poistetaan guttations - tämä on veden vapautumista erityisten erityskennojen kautta, jotka sijaitsevat lehden reunaa tai kärkeä pitkin (kuva 5.28).

Kuva 5.28. Guttation tyypit eri kasveissa
(mukaan A. M. Grodzinsky, 1965):
1 - viljojen, 2 - metsä mansikoiden, 3 - tulppaanien, 4 - maitoraudan,
5 - Sarmatian Bellevaliassa, 6 - apila
Hygrofyyttien lisäksi myös monet mesofyytit kykenevät suolistumaan. Esimerkiksi Ukrainan steppeissä suolistumista löytyi yli puolella kaikista kasvilajeista. Monet niittyrohut suolistuvat niin paljon, että ne kostuttavat maaperän pintaa. Joten eläimet ja kasvit mukautuvat sateiden vuodenaikojen jakautumiseen, määrään ja luonteeseen. Tämä määrittelee kasvien ja eläinten koostumuksen, tiettyjen vaiheiden virtauksen ajoittumisen niiden kehitysjaksossa.
Kosteuteen vaikuttaa myös vesihöyryn tiivistyminen, jota tapahtuu usein ilman pintakerroksessa lämpötilan muuttuessa. Kastehäviö tapahtuu, kun lämpötila laskee illalla. Kastepisarat ovat usein niin suuria, että se kostuttaa kasveja runsaasti, valuu maaperään, lisää ilmankosteutta ja luo suotuisat olosuhteet eläville organismeille, varsinkin kun muita sateita on vähän. Kasvit edistävät kasteen laskeutumista. Yöllä jäähdyttäen ne kondensoivat vesihöyryä itseensä. Sumu, paksut pilvet ja muut luonnonilmiöt vaikuttavat merkittävästi kosteuteen.
Kun kasvien elinympäristö määritetään vesifaktorilla, käytetään indikaattoreita, jotka heijastavat kosteuden pitoisuutta ja jakautumista paitsi ilmassa, myös maaperässä. Maaperän vesi tai maaperän kosteus, on yksi tärkeimmistä kasvien kosteuden lähteistä. Maaperän vesi on pirstoutuneessa tilassa, sen välissä on erikokoisia ja -muotoisia huokosia, sillä on suuri rajapinta maaperän kanssa, se sisältää useita kationeja ja anioneja. Siksi maaperän kosteus on heterogeeninen fysikaalisten ja kemiallisten ominaisuuksien suhteen. Kaikkia maaperän vettä ei voida käyttää kasveissa. Kasvien fysikaalisen tilan, liikkuvuuden, saatavuuden ja merkityksen mukaan maavesi jaetaan painovoimaiseksi, hygroskooppiseksi ja kapillaariseksi.
Maaperä sisältää myös höyryävää kosteutta, joka vie kaikki vedetön huokoset. Se on melkein aina tyydyttyneitä vesihöyryjä (paitsi aavikon maaperää). Kun lämpötila laskee alle 0 ° C, maaperän kosteus siirtyy jääksi (aluksi vapaa vesi ja myöhemmin jäähdyttäen osa sitoutuneesta vedestä).
Veden kokonaismääräksi, jonka maaperä voi pidättää (se määritetään lisäämällä ylimääräistä vettä ja odottamalla sitten, kunnes se lakkaa putoamasta), kutsutaan kentän kosteuskapasiteetti.
Siksi maaperän veden kokonaismäärä ei voi luonnehtia sitä, missä määrin kasveille tarjotaan kosteutta. Sen määrittämiseksi kokonaismäärästä vettä on tarpeen vähentää kertymiskerroin. Fysikaalisesti saavutettavissa oleva maavesi ei kuitenkaan ole fysiologisesti aina saatavissa kasveille alhaisen maaperän lämpötilan, happea puuttumisen vuoksi maavedessä ja maaperän ilmassa, maaperän happamuuden ja korkean pitoisuuden ollessa mineraalisuoloja liuenneina maaveteen. Juurten veden imeytymisen ja sen lehtien palaamisen välinen epäsuhta johtaa kasvien kuivumiseen. Maanpäällisten osien lisäksi myös kasvien juurten kehitys riippuu fysiologisesti saatavan veden määrästä. Kuivassa maaperässä kasvavissa kasveissa juurijärjestelmä on yleensä haaroittunut, voimakkaampi kuin märissä kasveissa (kuva 5.29).

Kuva 5.29. Talvvehnän juurijärjestelmä
(mukaan V.G. Khrzhanovsky ja muut, 1994):
1 - runsaasti sadetta; 2 - keskimäärin;
3 - pienille
Yksi maaperän kosteuden lähteistä on pohjavesi. Matalalla tasolla kapillaarivesi ei päästä maaperään eikä vaikuta sen vesitilaan. Maaperän kosteutuminen vain ilmakehän sateiden vuoksi aiheuttaa voimakkaita kosteuden vaihteluita, mikä vaikuttaa usein negatiivisesti kasveihin. Liian korkea pohjaveden pitoisuus on haitallista, koska se johtaa maaperän vettymiseen, hapen kulutukseen ja rikastumiseen mineraalisuoloilla. Maaperän jatkuva kosteus sääolosuhteista riippumatta tarjoaa pohjaveden optimaalisen tason.
Lämpötilatila. Pohja-ilmaympäristön erottuva piirre on suuri lämpötilavaihtelu. Suurimmalla osalla maata päivittäiset ja vuotuiset lämpötilan amplitudit ovat kymmeniä asteita. Erityisen merkittävät ilman lämpötilan muutokset autiomaassa ja mantereen ympäröivillä alueilla. Esimerkiksi kausilämpötila-alue Keski-Aasian aavikoissa on 68-77 ° С ja päivittäinen lämpötila on 25-38 ° С. Jakutskin läheisyydessä tammikuun keskilämpötila on -43 ° С, heinäkuun keskilämpötila on + 19 ° С ja vuotuinen vaihteluväli on -64 ... + 35 ° С. Trans-Uralissa vuosilämpötilan vaihtelut ovat teräviä ja yhdistyvät suuresti talven ja kevään kuukausien lämpötilojen vaihteluihin eri vuosina. Kylmin on tammikuussa, keskimääräinen ilman lämpötila on -16 - -19 ° C, joissakin vuosissa se laskee -50 ° C: seen, lämpimin kuukausi on heinäkuu ja lämpötila on 17,2 - 19,5 ° C. Enimmäislämpötilat ovat 38-41 ° C.
Lämpötilan vaihtelut maaperän pinnalla ovat vielä merkittäviä.
Maalaiskasvit vievät vyöhykkeen, joka on maaperän pinnan vieressä, toisin sanoen ”rajapinnassa”, jolla tapahtuvat säteet siirretään väliaineesta toiseen tai toisella tavalla - läpinäkyvästä läpinäkymättömään. Tälle pinnalle luodaan erityinen lämpötila: päivällä - voimakas lämmitys lämmonsäteiden imeytymisen takia, yöllä - voimakas jäähdytys säteilystä. Tästä eteenpäin pintakerroksessa ilmassa on voimakkaimmat vuorokauden lämpötilan vaihtelut, jotka ovat voimakkaimpia paljaalla maaperällä.
Esimerkiksi kasvien elinympäristön lämpötilaluonnehditaan suoraan kasvillisuuspeitteessä olevien lämpötilamittausten perusteella. Ruohoisissa yhteisöissä mittaukset tehdään nurmikasteen sisällä ja pinnalla sekä metsissä, joissa on tietty vertikaalinen lämpötilagradientti, useissa pisteissä eri korkeuksilla.
Maa-organismien ympäristön lämpötilan muutosten vastustuskyky on erilainen ja riippuu tietystä elinympäristöstä, jossa niiden elämä tapahtuu. Joten jauhetut lehtikasvit kasvavat suurimmaksi osaksi laajalla lämpötila-alueella, ts. Ne ovat eurythermisiä. Niiden elinikä aktiivisessa tilassa pidentää yleensä 5 - 55 ° C, kun taas 5 - 40 ° C nämä kasvit ovat tuottavia. Manneralueiden kasvit, joille on ominaista selkeä päivälämpötila, kehittyvät parhaiten, kun yö on 10–15 ° C kylmempi kuin päivä. Tämä pätee useimpiin lauhkean alueen kasveihin - lämpötilaero on 5-10 ° C, ja trooppisiin kasveihin, joiden amplitudi on vielä pienempi - noin 3 ° C (kuva 5.30).
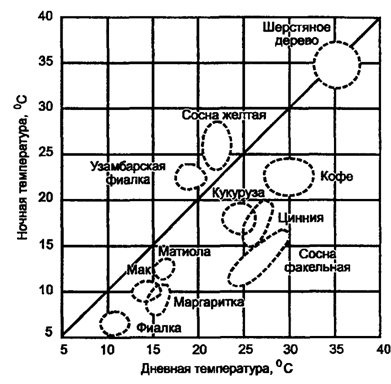
Kuva 5.30. Alueet, joissa optimaaliset lämpötilat kasvuun ja
erilaisten kasvien kehittäminen (Wentin, 1957 mukaan)
Poikilotermisissä organismeissa, joissa lämpötila nousee (T), kehityksen kesto (t) lyhenee yhä nopeammin. Vt: n kehitysnopeus voidaan ilmaista kaavalla Vt = 100 / t.
Tietyn kehitysvaiheen saavuttamiseksi (esimerkiksi hyönteisillä - munasta), ts. nukkuminen, mielikuvitusvaihe, vaatii aina tietyn lämpötilan. Efektiivisen lämpötilan (lämpötilan, joka on kehityksen nollapisteen yläpuolella, ts. T-To) korkeuskehitys kehitysajan perusteella (t) antaa spesifisyyden tälle tyypille lämpövakio kehitys c \u003d t (T-To). Tätä yhtälöä käyttämällä on mahdollista laskea tietyn kehitysvaiheen, esimerkiksi kasvintuholaisen, alkamisen aika, jolloin taistelu on tehokasta.
Poikilotermisinä organismeina olevilla kasveilla ei ole omaa vakaan kehon lämpötilaa. Niiden lämpötila määräytyy lämpötasapainon mukaan, toisin sanoen imeytymisen ja energiansiirron suhteen. Nämä arvot riippuvat sekä ympäristön (säteilyn saapumisen koko, ympäröivän ilman lämpötila ja sen liikkuvuus) että kasvien itsestään (kasvin väri ja muut optiset ominaisuudet, lehtien koko ja sijainti jne.) Monista ominaisuuksista. Ensisijaisella roolilla on verenpoiston jäähdytysvaikutus, joka estää kasvien voimakasta ylikuumenemista kuumissa elinympäristöissä. Edellä esitetyistä syistä kasvien lämpötila poikkeaa yleensä (usein melko huomattavasti) ympäröivän ilman lämpötilasta. Tässä on mahdollista kolme tilannetta: kasvin lämpötila on korkeampi kuin ympäristön lämpötila, sen alapuolella, yhtä suuri tai lähellä sitä. Kasvin lämpötilan yli ilman lämpötilaa esiintyy paitsi lämpimästi myös kylmempissä elinympäristöissä. Tätä helpottavat kasvien tumma väri tai muut optiset ominaisuudet, jotka lisäävät auringon säteilyn imeytymistä, sekä anatomiset ja morfologiset ominaisuudet, jotka osaltaan vähentävät verenkiertoa. Arktisista kasveista voi tulla varsin huomattavia (kuva 5.31).
Toinen esimerkki on kääpiöpaju - Salix arctica Alaskassa, jolla on 2–11 ° C lämpimämpiä päivisin lehtiä ja 1–3 ° C jopa napojen ympäri vuorokauden ympäri.
Varhaisen kevään efemeroidit, ns. Lumikellat, kuumentaen lehtiä, tarjoavat mahdollisuuden melko intensiiviseen fotosynteesiin aurinkoisilla, mutta silti kylmissä kevätpäivissä. Kylmissä elinympäristöissä tai vuodenaikojen lämpötilan vaihteluihin liittyvissä asioissa kasvien lämpötilan nostaminen on ympäristölle erittäin tärkeää, koska fysiologiset prosessit saavat samalla itsenäisyyden tietyissä rajoissa ympäröivästä termisestä taustasta.
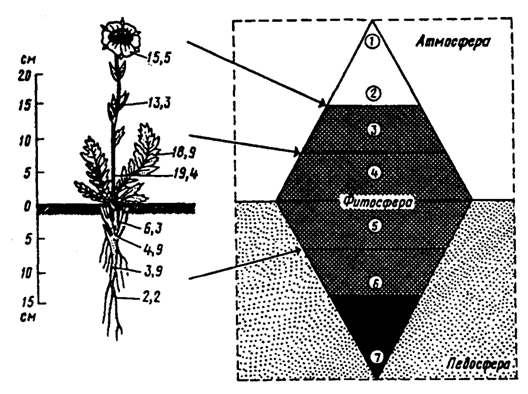
Kuva 5.31. Lämpötilan jakautuminen arktisen tundran (Novosieversia glacialis) ruusukasveessa aurinkoisella kesäkuun aamulla ilman lämpötilassa 11,7 ° C (B. A. Tikhomirovin mukaan, 1963)
Oikealla on biosfäärissä elintärkeiden prosessien voimakkuus: 1 - kylmin ilmakerros; 2 - verson kasvun yläraja; 3, 4, 5 - alue, jolla elintärkeitä prosesseja esiintyy eniten ja orgaaninen aine on kertynyt mahdollisimman paljon; 6 - ikirouta ja juurtumisen alaraja; 7 - alue, jolla on alhaisimmat maaperän lämpötilat
Kasvien lämpötilan laskua suhteessa ympäröivään ilmaan havaitaan useimmiten maapallon voimakkaasti valaistuilla ja lämmitetyillä alueilla (autiomaa, aroa), joilla kasvien lehtipinta on voimakkaasti pienentynyt, ja tehostettu verenkierto auttaa poistamaan ylimääräisen lämmön ja estää ylikuumenemisen. Yleisesti voidaan todeta, että kuumissa elinympäristöissä kasvien ilmaosien lämpötila on matalampi ja kylmissä - korkeampi kuin ilman lämpötila. Kasvin lämpötilan ja ympäristön lämpötilan yhteensopivuus on harvinaisempaa - olosuhteissa, joissa ei ole voimakasta säteilyn virtausta ja voimakasta kulkeutumista, esimerkiksi metsäkatoksen alla olevissa nurmikasveissa ja avoimissa tiloissa pilvisellä säällä tai sateella.
Yleisesti ottaen maa-organismit ovat vesieliöihin verrattuna enemmän eurooppalaisia.
Ilmaympäristössä elinolot ovat monimutkaisia säämuutokset. Sää on jatkuvasti muuttuva ilmakehän tila maan pinnalla noin 20 km korkeuteen (troposfäärin raja). Säävaihtelu ilmenee ympäristötekijöiden, kuten lämpötilan ja kosteuden, pilvien peittävyyden, sateen, tuulen voimakkuuden ja suunnan, jne. Yhdistelmän jatkuvassa vaihtelussa (kuva 5.32).
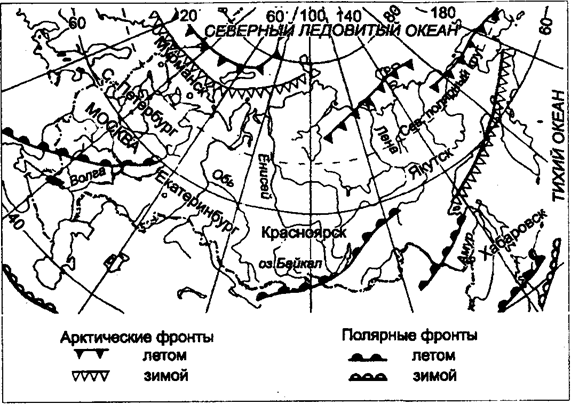
Kuva 5.32. Ilmakehän rintamat Venäjän alueen yli
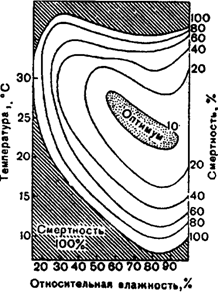
Säämuutoksille ja niiden säännölliselle vuosisyklin vuorottelulle ovat ominaisia \u200b\u200bepäjaksolliset vaihtelut, jotka monimutkaistavat huomattavasti maaperäisten organismien olosuhteita. Kuvassa 1 Kohdassa 5.33 esitetään kuolleisuuden riippuvuus lämpötilasta ja suhteellisesta kosteudesta Carpocapsa pomonella -kukkakoiran esimerkissä.
Kuva 5.33. Carpocapsa-siipikarjan toukkien kuolevuus
pomonella lämpötilasta ja kosteudesta riippuen (R. Dajon mukaan, 1975)
Tästä seuraa, että tasa-arvoisen kuolleisuuden käyrät ovat muodoltaan samankeskisiä ja että optimaalista vyöhykettä rajoittaa suhteellinen kosteus 55 ja 95% ja lämpötila 21 ja 28 ° C.
Kasvien valo, lämpötila ja ilmankosteus eivät yleensä määrittele stomaattien enimmäisarvoa, vaan keskimääräistä avautumisastetta, koska kaikkien niiden avautumiseen johtavien olosuhteiden sattuma on yhtä harvinaista.
Pitkäaikaiset sääolosuhteet ovat ominaisia alueen ilmasto. Ilmasto-käsitteeseen ei kuulu meteorologisten ilmiöiden keskimääräisten arvojen lisäksi myös niiden vuotuiset ja päivittäiset vaihtelut, poikkeamat ilmastosta ja niiden esiintymistiheys. Ilmaston määrää alueen maantieteelliset olosuhteet.
Tärkeimmät ilmastolliset tekijät ovat lämpötila ja kosteus, mitattuina sademäärällä ja ilman kyllästymisellä vesihöyryllä. Joten merestä syrjäisissä maissa tapahtuu asteittainen siirtyminen kosteasta ilmastosta puoliksi kuivasta välivyöhykkeestä satunnaisin tai ajoittaisin kuivina ajanjaksoina kuivalle alueelle, jolle on ominaista pitkäaikainen kuivuus, maaperän ja veden suolapitoisuus (kuva 5.34).
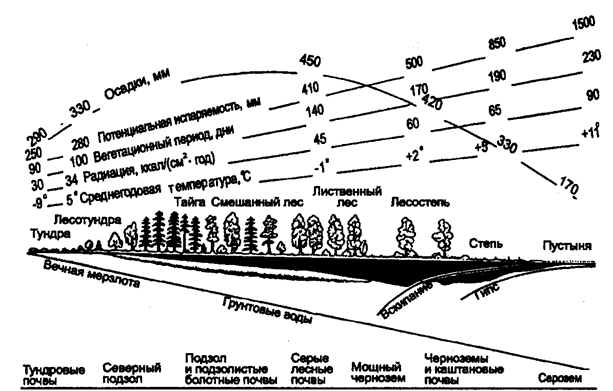
Kuva 5.34. Ilmaston, kasvillisuuden ja maaperän muutosprofiili pitkin Venäjän Euroopan osan päämaisemaa luoteesta kaakkoon Kaspian ala-alueelle (V.N. Sukachevin, 1934 mukaan)
Huomautus: Jos sadekäyrä ylittää nousevan haihtumislinjan, kostean (vasemman) ja kuivan (oikean) ilmaston välinen raja sijaitsee. Humushorisontti on esitetty mustalla, illuiaalinen horisontti on varjostettu
Jokaiselle elinympäristölle on ominaista erityinen ekologinen ilmasto, ts. Pintailmakerroksen ilmasto, tai eKOKLIMAT.
Kasvillisuudella on suuri vaikutus ilmastollisiin tekijöihin. Joten metsäkatoksen alla ilmankosteus on aina korkeampi ja lämpötilanvaihtelut ovat pienempiä kuin lahtia. Näiden paikkojen valotila on myös erilainen. Eri kasviyhdistykset muodostavat oman valon, lämpötilan, kosteuden, ts. Erikoisen phytoclimate.
Tietyn elinympäristön ilmasto-olosuhteiden karakterisoimiseksi ekologinen ilmasto- tai kasvillisuutta koskevat tiedot eivät aina riitä. Ympäristön paikalliset elementit (helpotus, altistuminen, kasvillisuus jne.) Muuttavat hyvin usein valon, lämpötilan, kosteuden, ilman liikkumisen tilaa tietyllä alueella siten, että se voi poiketa merkittävästi alueen ilmasto-olosuhteista. Ilmapintakerroksessa taittuvia paikallisia ilmastomuutoksia kutsutaan mikroilmaston. Esimerkiksi puun kuoren alla elävien hyönteisten toukkien elinolot ovat erilaisia \u200b\u200bkuin metsässä, jossa tämä puu kasvaa. Tavaratilan eteläpuolen lämpötila voi olla 10 - 15 ° C korkeampi kuin sen pohjoispuolen lämpötila. Vakaan mikroilmaston hallitsevat eläinten asuttamat urvet, puiden ontot ja luolat. Ympäristö- ja mikroilmaston välillä ei ole selviä eroja. Ekologisen ilmaston uskotaan olevan suurten alueiden ilmasto ja mikroilmasto on yksittäisten pienten alueiden ilmasto. Mikroilmasto vaikuttaa tietyn alueen, paikan eläviin organismeihin (kuva 5.35).
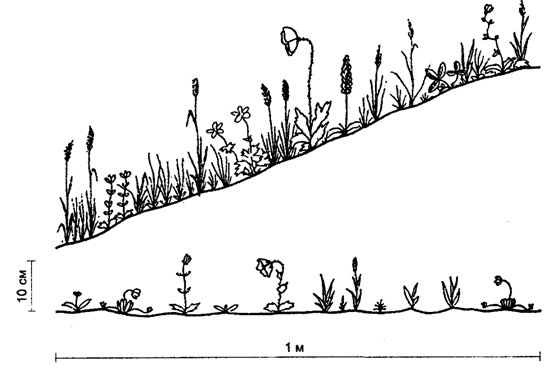
Kuva 5.35. Mikroilmaston vaikutus tundran kasvillisuuteen
(Yu. I. Tšernovin, 1979 mukaan):
yllä - eteläisen valotuksen hyvin lämmitetty kaltevuus;
alapuolella - vuoristoalueen vaakasuora osa (kukkakoostumus on sama molemmissa osissa)
Monien mikroilmastojen esiintyminen yhdessä paikassa varmistaa lajien rinnakkaiselon epätasa-arvoisten ympäristövaatimusten kanssa.
Maantieteellinen ja kaavoitus. Elävien organismien leviäminen maapallolla liittyy läheisesti maantieteellisiin alueisiin ja vyöhykkeisiin. Vyöllä on leveysisku, mikä tietenkin johtuu pääasiassa säteilyrajoista ja ilmakehän luonteen luonteesta. Maapallon pinnalla on 13 maantieteellistä vyöhykettä, jotka sijaitsevat mantereilla ja valtamerellä (kuva 5.36).
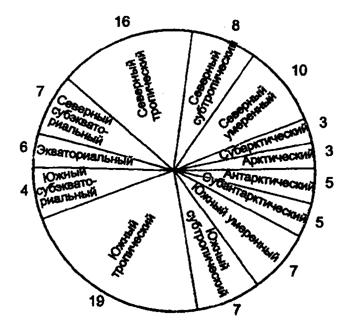
Kuva 5.36. Erilaisten käyttämien maa - alueiden suhde
fyysiset ja maantieteelliset alueet,% (N.F. Reimersin, 1990) mukaan
Se on sellaista kuin arktinen, antarktinen, subarktinen, subantarktinen, pohjoiseen ja etelään kohtalaista, pohjoiseen ja etelään subarktinen,pohjoiseen ja etelään trooppinen, pohjoiseen ja etelään subequatorial ja eQUATORIAL. Vyöt sisältä erittyvät maantieteelliset alueet missä säteilyolosuhteiden ohella otetaan huomioon maan pinnan kostuminen ja tälle vyöhykkeelle ominainen lämmön ja kosteuden suhde. Toisin kuin valtameri, jossa kosteus on täydellistä, mantereilla lämmön ja kosteuden suhteella voi olla merkittäviä eroja. Siksi maantieteelliset alueet ulottuvat mantereille ja valtamereille ja maantieteelliset alueet - vain mantereille. erottaa leveysvyöhyke ja pituuspiiri tai pituusaste luonnolliset alueet. Entinen ulottuu lännestä itään, jälkimmäinen pohjoisesta etelään. Pituussuunnassa leveysvyöhykkeet jaetaan subzone ja leveysasennossa maakunnassa.
Luonnollisen vyöhykkeen tutkimuksen perustaja on V. V. Dokuchaev (1846–1903), joka perusti kaavoituksen yleismaailmalliseksi luonnonlakiin. Kaikki biosfäärin ilmiöt ovat tämän lain alaisia. Kaavoituksen pääasialliset syyt ovat maan muoto ja sen sijainti aurinkoon nähden. Leveysasteen lisäksi lämmön jakautumiseen maan päällä vaikuttavat helpotuksen luonne ja alueen korkeus merenpinnan yläpuolella, maan ja meren suhde, merivirrat jne.
Myöhemmin A. A. Grigoriev ja M. I. Budyko kehittivät säteilypohjat maapallon vyöhykkeen muodostamiseksi. Lämpö- ja kosteussuhteen kvantitatiivisen ominaisuuden määrittämiseksi eri maantieteellisille alueille määritettiin joitain kertoimia. Lämmön ja kosteuden suhde ilmaistaan \u200b\u200bsuhteena pinnan säteilytasapainosta piilevän haihtumislämpötilan ja sademäärän suhteena (kuivuuden säteilyindeksi). Laadittiin laki, jota kutsutaan jaksollisen maantieteellisen alueellisuuden lakiksi (A. A. Grigoriev - M. I. Budyko), jossa todetaan että maantieteellisten alueiden muuttuessa samanlainen maantieteellinen (maisema, luonnollinen) vyöhykkeet ja jotkut niiden yhteisistä ominaisuuksista toistetaan ajoittain.
Jokainen vyöhyke on rajattu tiettyyn indikaattoriarvojen joukkoon: geomorfologisten prosessien erityisluonne, erityinen ilmasto, kasvillisuus, maaperä ja villieläimet. Seuraavat maantieteelliset alueet havaittiin entisen Neuvostoliiton alueella: jää, tundra, metsä-tundra, taiga, sekametsät. Venäjän tavallinen, Kaukoidän monsuuni sekoitettu metsä, metsä-stepit, stepit, puoliväylät, lauhkeat aavikot, subtrooppisen alueen aavikot, Välimeren alue ja kosteat subtroopit.
Yksi tärkeistä olosuhteista organismien vaihtelevuudelle ja niiden alueelliselle jakautumiselle maan päällä on väliaineen kemiallisen koostumuksen vaihtelevuus. Tässä suhteessa A. P. Vinogradovin opetukset aiheesta biogeokemialliset maakunnat, jotka määritetään maaperän kemiallisen koostumuksen alueellisuuden perusteella, samoin kuin biosfäärin ilmasto-, fytogeografinen ja geokemiallinen vyöhyke. Biogeokemialliset maakunnat ovat maapallon pinta-aloja, jotka eroavat kemiallisten yhdisteiden pitoisuuksista (maaperässä, vesissä jne.), Jotka liittyvät tiettyihin paikallisen kasviston ja eläimistön biologisiin reaktioihin.
Maan ympäristössä olevan horisontaalisen vyöhykkeen lisäksi se ilmenee selvästi korkealla tai pystysuora zonation.
Vuoristoisten maiden kasvillisuus on rikkaampaa kuin viereisillä tasangoilla, ja sille on ominaista endeemisten muotojen lisääntynyt leviäminen. Joten, O. E. Agakhanyantsin (1986) tietojen mukaan Kaukasuksen kasvistoon kuuluu 6350 lajia, joista 25% on endeemisiä. Keski-Aasian vuorten kasvistoksi arvioidaan 5500 lajia, joista 25–30% on endeemisiä, kun taas eteläisten autiomaiden viereisillä tasangoilla on 200 kasvilajia.
Vuorikiipeilyssä toistuu sama vyöhykkeiden muutos kuin päiväntasaajasta napoihin. Aavikon juurella sijaitsevat yleensä aavikot, sitten stepit, leveälehtiset metsät, havumetsät, tundra ja lopulta jää. Täydellistä analogiaa ei kuitenkaan vielä ole. Vuorikiipeilyssä ilman lämpötila laskee (ilman lämpötilan keskimääräinen gradientti on 0,6 ° C / 100 m), haihtuminen vähenee, ultravioletti säteily, valaistuksen lisääntyminen jne. Kaikki tämä saa kasvit mukautumaan kuiviin tai märkiin haitoihin. Tyynynmuotoiset elämämuodot, monivuotiset, jotka ovat kehittyneet sopeutumiseen voimakkaaseen ultraviolettisäteeseen ja vähentävät verenkiertoa, hallitsevat kasveissa.
Ylämaan villieläimet ovat myös ominaisia. Alennettu ilmanpaine, merkittävä auringon säteily, terävät päivä- ja yölämpötilojen vaihtelut, ilmankosteuden muutokset korkeuden kanssa myötävaikuttivat vuoristoeläinten organismin erityisen fysiologisen sopeutumisen kehitykseen. Esimerkiksi eläimissä sydämen suhteellinen tilavuus kasvaa, veren hemoglobiinipitoisuus kasvaa, mikä antaa mahdollisuuden imeä enemmän happea ilmasta. Kivinen maaperä vaikeuttaa tai melkein eliminoi eläinten hautaamista. Monet pienet eläimet (pienet jyrsijät, pikot, liskoja jne.) Löytävät turvapaikan kivien rakoihin, luoliin. Lintuista vuoristoalueille on luonteenomaista kalkkunat (Ulars), vuoripihat, haurat ja suuriin lintuihin kuuluvat parrakas, korppikotkat ja kondorit. Suurten nisäkkäiden vuoristossa elävät lampaat, vuohet (mukaan lukien lumi vuohet), säämiskä, jakit jne. Petoeläimiä edustavat lajit, kuten susit, ketut, karhut, ilveset, lumileopardi (lumileopardi) jne.
| edellinen |
Luento 3 Luontotyypit ja niiden ominaisuudet (2 tuntia)
1.Vesiympäristö
2. Maa-ilman elinympäristö
3. Maaperä elinympäristönä
4. Organismi elinympäristönä
Historiallisen kehityksen prosessissa elävät organismit ovat hallinneet neljä elinympäristöä. Ensimmäinen on vesi. Vedessä elämä syntyi ja kehittyi monien miljoonien vuosien aikana. Toinen - maa-ilma - maalla ja ilmakehässä kasveja ja eläimiä syntyi ja sopeutui nopeasti uusiin olosuhteisiin. Maan ylemmän kerroksen - litosfäärin - muuttuessa vähitellen he loivat kolmannen elinympäristön - maaperän, ja heistä itsestään tuli neljäs elinympäristö.
Vesieliöstö - hydrosfääri
Hydrobionttien ekologiset ryhmät. Elämän suurimmalle monimuotoisuudelle on ominaista lämmin meri ja valtameri (40 000 eläinlajia) päiväntasaajalla ja tropiikalla; pohjoiseen ja etelään merien kasvisto ja eläimistö on ehtynyt satoja kertoja. Mitä tulee organismien jakautumiseen suoraan mereen, niiden massa on keskittynyt pintakerroksiin (epipelagial) ja sublittoraaliseen vyöhykkeeseen. Riippuen liikkumistavasta ja pysyvyydestä tietyissä kerroksissa, meren asukkaat jaetaan kolmeen ekologiseen ryhmään: nektoni, planktoni ja pohjaeliö.
Nekton(nektot - uinti) - aktiivisesti liikkuvat suuret eläimet, jotka voivat kulkea pitkiä matkoja ja voimakkaita virtauksia: kalat, kalmarit, sorkkaeläimet, valaat. Makeassa vedessä sammakkoeläimet ja monet hyönteiset kuuluvat nektoniin.
plankton (planktot - vaeltelu, huiman nousu) - joukko kasveja (kasviplanktoni: piimat, vihreät ja sinivihreät (vain makean veden) levät, kasvien siipikarjat, peridinea jne.) ja pieniä eläinorganismeja (eläinplanktoni: pienet äyriäiset, suuret ovat siivekäs nilviäiset, meduusat, ketteroforit, jotkut madot), jotka elävät eri syvyyksissä, mutta eivät kykene aktiivisiin liikkeisiin ja kestämään virtauksia. Planktoniin kuuluu myös eläinten toukkia, jotka muodostavat erityisryhmän - Neustonin. Tämä on passiivisesti kelluva "väliaikainen" kanta ylimmässä vesikerroksessa, jota edustavat erilaiset eläimet (havupuut, sinisorsat ja selkärankaiset, äyriäiset, piikkinahkaiset, moniherokot, kalat, nilviäiset jne.) Toukkavaiheessa. Kasvavat toukat siirtyvät pelageelin alempiin kerroksiin. Pleiston sijaitsee neuston yläpuolella - nämä ovat organismeja, joissa kehon yläosa kasvaa veden yläpuolella ja alaosa vedessä (dubleweed - Lemma, sifonoforit jne.). Planktonilla on tärkeä rooli biosfäärin troofisissa siteissä, koska Se on monien vesieläinten ruokia, mukaan lukien paalen valaiden (Myatcoceti) pääruoka.
pohjaeliöstölle (pohjaeliö - syvyys) - pohjan hydrobiontit. Sitä edustavat pääasiassa kiinnittyneet tai hitaasti liikkuvat eläimet (eläinjalkaat: foramiinifoorut, kalat, sienet, maha-suolikanavat, madot, brachopod-nilviäiset, astsidia jne.), Joita on enemmän matalassa vedessä. Matalassa vedessä bentosiin sisältyy myös kasveja (fytobentos: piimat, vihreät, ruskeat, punalevät, bakteerit). Syvyyksessä, jossa ei ole valoa, fytobentos puuttuu. Lähellä rannikkoa on kukinnan kasvit zoster, rupia. Pohjan rikkaimmat osat ovat rikkaimpia kasvibentosessa.
Järvissä zoobenthos on vähemmän runsasta ja monimuotoista kuin meressä. Sitä muodostavat alkueläimet (silikaatit, daphnia), piilot, nilviäiset, hyönteisten toukat jne. Järvien fytobentos muodostetaan vapaasti kelluvilla piimatomilla, vihreillä ja sinivihreillä. ruskeat ja punalevät puuttuvat.
Järvien rannikkokasvien juurtuminen muodostaa erilliset vyöt, joiden lajien koostumus ja ulkonäkö ovat yhdenmukaisia \u200b\u200bmaa-vesirajavyöhykkeen ympäristöolosuhteiden kanssa. Hydrofyytit kasvavat vedessä lähellä rantaa - vedessä on osittain upotettuja kasveja (nuolenlehti, whitefly, ruoko, piikkisara, siili, trefoil, ruoko). Ne korvataan hydatoofyyteillä - veteen upotetut kasvit, mutta kelluvilla lehdillä (lootus, dubleweed, munakapselit, chilim, takla) ja - edelleen - täysin upotettuina (tuholainen, elodea, hara). Hydatofyyteihin kuuluvat myös pinnalla kelluvat kasvit (duckweed).
Vesiympäristön korkea tiheys määrää elämää tukevien tekijöiden muutosten erityisen koostumuksen ja luonteen. Jotkut niistä ovat samat kuin maalla - lämpö, \u200b\u200bvalo, toiset spesifiset: vedenpaine (syvyyden kasvaessa 1 atm. Jokaista 10 m kohden), happipitoisuus, suolakoostumus, happamuus. Väliaineen suuren tiheyden vuoksi lämmön ja valon arvot korkeusgradientilla muuttuvat paljon nopeammin kuin maalla.
Lämpötila. Vesiympäristölle on ominaista alhaisempi lämmöntuotto, koska merkittävä osa siitä heijastuu, ja yhtä merkittävä osa kulutetaan haihtumiseen. Pinnan lämpötilojen dynamiikan mukaisesti veden lämpötilassa on pienemmät vaihtelut päivä- ja kausilämpötiloissa. Lisäksi vesimuodostumat tasoittavat huomattavasti lämpötilan kulkua rannikkoalueiden ilmakehässä. Jos merestä ei ole jääkuorta kylmällä kaudella, niillä on eristävä vaikutus viereiseen maahan, kesällä ne viilenevät ja kosteuttavat.
Maailman valtameren veden lämpötila-alue on 38 ° (välillä -2 ... + 36 ° C), makean veden lämpötilassa - 26 ° (-0,9 ... + 25 ° C). Syvyyden myötä veden lämpötila laskee voimakkaasti. 50 metriin saakka, päivittäisiä lämpötilanvaihteluita havaitaan, jopa 400 metriin asti - kausiluonteisesti, sitä syvemmäksi muuttuu vakiona, laskevan + 1–3 ° С: seen (arktisella alueella se on lähellä 0 ° С). Koska säiliöiden lämpötila on suhteellisen vakaa, niiden asukkaille on ominaista stenoterminen olosuhde. Pienempiin lämpötilanvaihteluihin toiseen suuntaan liittyy merkittäviä muutoksia vesiekosysteemeissä.
Esimerkkejä: ”biologinen räjähdys” Volgan suistossa Kaspianmeren tason alenemisen vuoksi - lootuspensien (Nelumba kaspium) umpeenkasvu, Primoryen eteläosassa - vanhojen jokien (Komarovka, Ilistaya jne.) Umpeenkasvu, jota pitkin rannat raivattiin ja poltettiin.
Ylä- ja alakerrosten vaihtelevien kuumennusasteiden, vuodenaikojen, virtausten, myrskyjen vuoksi vesikerrokset sekoittuvat jatkuvasti. Veden sekoittamisen merkitys vesieliöille (vesieliöille) on erittäin suuri, koska Samanaikaisesti hapen ja ravinteiden jakautuminen säiliöissä tasaistuu, mikä aikaansaa aineenvaihduntaprosessit organismien ja ympäristön välillä.
|
Laukaisten leveysasteiden seisovissa säiliöissä (järvissä) pystysuuntaista sekoittumista tapahtuu keväällä ja syksyllä, ja näinä vuodenaikoina lämpötila koko säiliössä tasaistuu, ts. on tulossa homothermia. Yläkerrosten lämmityksen tai jäähdytyksen voimakkaan lisääntymisen seurauksena veden sekoittuminen lopettaa kesällä ja talvella. Tätä ilmiötä kutsutaan lämpötilaerotukseksi, ja tilapäisen pysähtymisen aikaa kutsutaan pysähtymiseksi (kesä tai talvi). Kesällä pinnalle jää kevyempiä lämpimiä kerroksia, jotka sijaitsevat raskaiden kylmien yläpuolella (kuva 3). Talvella päinvastoin, lämpimämpi vesi on pohjakerroksessa, koska suoraan jään alapuolella pintavesien lämpötila on alle + 4 ° C ja veden fysikaalis-kemiallisista ominaisuuksista johtuen ne muuttuvat kevyemmiksi kuin vesi, jonka lämpötila on yli + 4 ° C. Pysähtymisjaksojen aikana kolme kerrosta erotetaan selvästi: yläosa (epilimnioni), jonka vesilämpötilassa on voimakkaimpia kausivaihteluita, keskimmäinen (metallimnioni tai termocline), jossa lämpötila nousee voimakkaasti, ja pohja (hypolimnion), jossa lämpötila muuttuu hieman vuoden aikana. Stagnaatiojaksoina happea puuttuu vesipylväässä - kesällä pohjaosassa ja talvella yläosassa, minkä seurauksena kaloja esiintyy usein talvella. |
Valotila.Veden valon voimakkuus heikkenee huomattavasti, koska se heijastuu pinnasta ja absorboi itse vettä. Tämä vaikuttaa suuresti fotosynteettisten kasvien kehitykseen. Mitä alhaisempi veden läpinäkyvyys, sitä enemmän valoa imeytyy. Veden läpinäkyvyyttä rajoittavat mineraalisuspensiot, planktoni. Se vähenee pienten organismien nopean kehityksen myötä kesällä ja maltillisilla ja pohjoisilla leveysasteilla - myös talvella, kun jääpeite on muodostettu ja lumen peittänyt ylhäältä.
Valtamereissä, joissa vesi on hyvin läpinäkyvää, 1% valonsäteilystä tunkeutuu 140 m syvyyteen ja pienissä järvissä 2 m syvyydessä vain kymmenesosa kymmenesosasta. Spektrin eri osien säteitä ei imeydy veteen samalla tavalla, ensin punaiset säteet imeytyvät. Syvyyden myötä se tummenee ja veden väri muuttuu ensin vihreäksi, sitten siniseksi, siniseksi ja lopulta sini-violetiksi, muuttuen täydelliseksi pimeydeksi. Vastaavasti väri ja hydrobiontit muuttuvat, mukautuen paitsi valon koostumukseen myös sen puutteeseen - kromaattiseen sopeutumiseen. Vaaleilla alueilla, matalassa vedessä, vallitsevat vihreät levät (Chlorophyta), joiden klorofylli imee punaiset säteet, syvyydessä ne korvataan ruskealla (Phaephyta) ja sitten punaisella (Rhodophyta). Kasvien pohjaosa puuttuu suuresta syvyydestä.
Kasvit, jotka on mukautettu valon puutteeseen kehittämällä suuria kromatoforeja, jotka tarjoavat alhaisen kompensointipisteen fotosynteesille, samoin kuin lisääntyneiden elinten pinta-alan (lehtien pintaindeksi). Syvänmeren levästä tyypillisesti leikatut lehdet ovat tyypillisiä; lehtien lavat ovat ohuita, läpikuultavia. Heterofylia on ominaista osittain vedenalaisille ja kelluville kasveille - veden yläpuolella olevat lehdet ovat samat kuin maalaiskasvien lehdet, niissä on yksi laatta, vatsakalvo on kehitetty ja lehdet ovat hyvin ohuita vedessä ja koostuvat kapeista filiformisista fraktioista.
Geterofilliya: munakapselit, vesililjat, nuolenpää, chili (vesikastanja).
Eläimet, kuten kasvitkin, muuttavat luonnollisesti väriään syvyydellä. Ylemmissä kerroksissa ne ovat väriltään kirkkain eri väreillä, hämärävyöhykkeellä (meriahven, korallit, äyriäiset) maalattu punaisella sävyllä - se on helpompi piiloutua vihollisilta. Syvänmeren lajeista puuttuu pigmenttejä.
Vesiympäristön, muualla kuin maassa, tunnusomaisia \u200b\u200bominaisuuksia ovat korkea tiheys, liikkuvuus, happamuus, kyky liuottaa kaasuja ja suoloja. Kaikille näille olosuhteille hydrobionteilla on historiallisesti kehitetty sopivia mukautuksia.
2. Maa-ilman elinympäristö
Evolutionin aikana tämä väliaine kehitettiin myöhemmin kuin vesieliö. Sen erikoisuus on siinä, että se on kaasumainen, siksi sille on ominaista alhainen kosteus, tiheys ja paine sekä korkea happipitoisuus. Evoluutioiden aikana elävät organismit ovat kehittäneet tarvittavat anatomiset, morfologiset, fysiologiset, käyttäytymis- ja muut mukautukset.
Pohja-ilmaympäristön eläimet liikkuvat maaperän tai ilman läpi (linnut, hyönteiset) ja kasvit juurtuvat maaperään. Tässä suhteessa keuhkot ja henkitorvi ilmestyivät eläimiin ja vatsakalvo, ts. elimet, joiden avulla planeetan maan asukkaat imevät happea suoraan ilmasta. Luurangan elimet, jotka tarjoavat autonomian liikkumiselle maalla ja tukielinten kanssa kaikilla elimilläan ympäristön alhaisen tiheyden olosuhteissa, tuhat kertaa pienempi kuin vesi, ovat kehittyneet voimakkaasti. Ympäristötekijät pohja-ilmaympäristössä eroavat muista luontotyypeistä korkean valon voimakkuuden, merkittävien lämpötilan ja kosteuden vaihtelun, kaikkien tekijöiden korrelaation suhteen maantieteelliseen sijaintiin, vuodenaikojen ja vuorokaudenajan muutoksen vuoksi. Niiden vaikutukset organismeihin liittyvät erottamattomasti ilman liikkeeseen ja sijaintiin mereihin ja valtameriin nähden, ja ne eroavat suuresti altistumisesta vesiympäristössä (taulukko 1).
Ilman ja vesieliöiden elinolot
(mukaan D.F. Mordukhai-Boltovsky, 1974)
|
ilmaympäristö |
vesiympäristö |
|
kosteus |
Erittäin tärkeä (usein vajaa) |
Ei (aina enemmän) |
|
tiheys |
Pieni (pois lukien maaperä) |
Suuri verrattuna sen rooliin ilman asukkaiden kannalta |
|
paine |
Lähes ei |
Suuri (voi saavuttaa 1000 ilmakehän) |
|
lämpötila |
Merkittävä (vaihtelee erittäin suurissa rajoissa - välillä -80 - + 1OO ° С ja enemmän) |
Pienempi kuin arvo asukkaille (vaihtelee paljon vähemmän, yleensä välillä -2 - + 40 ° C) |
|
happi |
Pieni (enimmäkseen yli) |
Välttämätöntä (usein vajaa) |
|
Suspendoitunut aine |
merkityksetön; ei käytetä elintarvikkeissa (pääasiassa mineraaleja) |
Tärkeää (ruoan lähde, erityisesti orgaaninen aine) |
|
Ympäristössä liuenneet aineet |
Jossain määrin (merkityksellinen vain maaperän ratkaisuissa) |
Tärkeää (tietyn määrän välttämätöntä) |
Maan eläimet ja kasvit kehittivät omat, vähintäänkin alkuperäiset mukautuksensa haitallisiin ympäristötekijöihin: kehon ja sen rakenteiden monimutkainen rakenne, elinkaaren jaksoittaisuus ja rytmi, lämmön säätelymekanismit jne. Kehitettiin eläinten tarkoituksenmukaista liikkuvuutta etsiessään ruokaa, itiöitä, siemeniä ja tuulen kantamia itiöitä, ja kasvien siitepöly, samoin kuin kasvit ja eläimet, joiden elämä on täysin yhteydessä ilmaympäristöön. Muodostui poikkeuksellisen läheinen toiminnallinen, resurssi- ja mekaaninen suhde maaperään.
Edellä olemme pitäneet monia mukautuksista esimerkkeinä abioottisten ympäristötekijöiden karakterisoinnissa. Siksi ei ole järkeä toistaa nyt, koska palaamme heidän luokseen käytännön oppitunneilla