Elävässä organismissa on kolme pääasiallista makromolekyyliä: proteiinit ja kahden tyyppiset nukleiinihapot. Niiden ansiosta ylläpidetään elintärkeää toimintaa ja koko organismin asianmukaista toimintaa. Mikä on nukleiinihappo? Mitä ne ovat? Tietoja tästä - tarkemmin artikkelissa.
Yleistä tietoa
Nukleiinihappo on biopolymeeri, orgaaninen yhdiste, jolla on suuri molekyylipaino ja joka muodostuu nukleotiditähteistä. Nukleiinihappojen suorittamisen tärkein tehtävä on sukupolvelta kaiken geneettisen informaation siirto sukupolvesta toiseen. Alla esitetty esitys paljastaa tämän käsitteen yksityiskohtaisemmin.
Tutkimushistoria
Ensimmäinen tutkittu nukleotidi eristettiin sonni lihaksista 1847 ja nimettiin "inosiinihapoksi". Kemiallisen rakenteen tutkimus osoitti, että se on 5'-fosfaatti-ribosidi ja siinä on N-glykosidinen sidos, ja vuonna 1868 löydettiin aine, jota kutsutaan "nukleiiniksi". Sveitsin kemisti Friedrich Miescher avasi sen eräiden biologisten aineiden tutkimuksessa. Tämän aineen koostumus oli fosforia. Yhdisteellä oli happamia ominaisuuksia eikä se hajonnut proteolyyttisten entsyymien vaikutuksen alaisena.  Aine sai kaavan C29H49N9O22P3. Oletus siitä, että nukleiini on osallistunut perinnöllisten tietojen siirtämisprosessiin, esitettiin sen samankaltaisuuden löytämisen seurauksena kemiallinen koostumus kromatiinin kanssa. Tämä elementti on kromosomien pääkomponentti, ja termi "nukleiinihappo" otettiin käyttöön vuonna 1889 Richard Altmann. Hän on kirjoittanut menetelmän näiden aineiden hankkimiseksi ilman proteiinien epäpuhtauksia, ja nukleiinihappojen alkalisen hydrolyysin tutkimuksen aikana Levin ja Jacob paljastivat tämän prosessin tuotteiden tärkeimmät osat. Ne olivat nukleotideja ja nukleosideja. Vuonna 1921 Levin ehdotti, että DNA: lla on tetranukleotidirakenne. Tämä hypoteesi ei kuitenkaan vahvistunut ja osoittautui virheelliseksi.
Aine sai kaavan C29H49N9O22P3. Oletus siitä, että nukleiini on osallistunut perinnöllisten tietojen siirtämisprosessiin, esitettiin sen samankaltaisuuden löytämisen seurauksena kemiallinen koostumus kromatiinin kanssa. Tämä elementti on kromosomien pääkomponentti, ja termi "nukleiinihappo" otettiin käyttöön vuonna 1889 Richard Altmann. Hän on kirjoittanut menetelmän näiden aineiden hankkimiseksi ilman proteiinien epäpuhtauksia, ja nukleiinihappojen alkalisen hydrolyysin tutkimuksen aikana Levin ja Jacob paljastivat tämän prosessin tuotteiden tärkeimmät osat. Ne olivat nukleotideja ja nukleosideja. Vuonna 1921 Levin ehdotti, että DNA: lla on tetranukleotidirakenne. Tämä hypoteesi ei kuitenkaan vahvistunut ja osoittautui virheelliseksi.  Tämän seurauksena syntyi uusi tilaisuus tutkia yhdisteiden rakennetta, ja vuonna 1940 Alexander Todd aloitti yhdessä tieteellisen ryhmänsä kanssa laajamittaisen tutkimuksen kemialliset ominaisuudet, nukleotidien ja nukleosidien rakenne, minkä seurauksena hänelle myönnettiin vuonna 1957 Nobelin palkinto, ja amerikkalainen biokemisti Erwin Chargaff totesi, että nukleiinihapot sisältävät eri tyyppisiä nukleotideja tietyssä kuviossa. Tulevaisuudessa tätä ilmiötä kutsuttiin nimellä ”Chargaff Rule”.
Tämän seurauksena syntyi uusi tilaisuus tutkia yhdisteiden rakennetta, ja vuonna 1940 Alexander Todd aloitti yhdessä tieteellisen ryhmänsä kanssa laajamittaisen tutkimuksen kemialliset ominaisuudet, nukleotidien ja nukleosidien rakenne, minkä seurauksena hänelle myönnettiin vuonna 1957 Nobelin palkinto, ja amerikkalainen biokemisti Erwin Chargaff totesi, että nukleiinihapot sisältävät eri tyyppisiä nukleotideja tietyssä kuviossa. Tulevaisuudessa tätä ilmiötä kutsuttiin nimellä ”Chargaff Rule”.
luokitus
Nukleiinihapot ovat kahdenlaisia: DNA ja RNA. Niiden läsnäolo havaitaan kaikkien elävien organismien soluissa. DNA: ta esiintyy pääasiassa solun ytimessä. RNA on sytoplasmassa. Vuonna 1935 pehmeän DNA: n fragmentoinnin aikana saatiin 4 DNA: ta muodostavaa nukleotidia. Nämä komponentit on esitetty kiteiden tilassa. Vuonna 1953 Watston ja Creek totesivat, että DNA: lla on kaksoiskierre.
Valintamenetelmät

Kehitetty erilaisia tapoja saada yhdisteitä luonnollisista lähteistä. Näiden tekniikoiden pääolosuhteet ovat nukleiinihappojen ja proteiinien tehokas erottaminen, prosessin aikana saatujen aineiden pienin fragmentoituminen. Nykyään klassista menetelmää käytetään laajalti. Tämän menetelmän ydin on biologisen materiaalin seinien tuhoutuminen ja niiden jatkokäsittely anionisella pesuaineella. Tuloksena on proteiinin sakka ja nukleiinihapot pysyvät liuoksessa. Käytetään toista menetelmää. Tässä tapauksessa nukleiinihapot voidaan sijoittaa geelitilaan käyttämällä etanolia ja suolaliuosta. On oltava varovainen. Erityisesti etanolia tulisi lisätä hyvin huolellisesti suolaliuokseen geelisaostuman saamiseksi. Missä pitoisuudessa nukleiinihappo vapautui, mitä epäpuhtauksia siinä on, se voidaan määrittää spektrofotometrisellä menetelmällä. Nukleiinihapot hajoavat helposti nukleaasilla, joka on erityinen entsyymiluokka. Tällä valinnalla on välttämätöntä, että laboratoriolaitteet hoidetaan pakollisesti estäjillä. Näitä ovat esimerkiksi DEPC-inhibiittori, jota käytetään RNA: n eristämisessä.
Fyysiset ominaisuudet
Nukleiinihappojen liukoisuus veteen on hyvä, ja ne eivät lähes liukene orgaanisiin yhdisteisiin. Lisäksi ne ovat erityisen alttiita lämpötila- ja pH-tasoille. High-nukleiinihappomolekyylit molekyylipainonukleaasi voi hajota mekaanisten voimien vaikutuksesta. Näitä ovat liuoksen sekoittaminen, sekoittaminen.
Nukleiinihapot. Rakenne ja toiminta
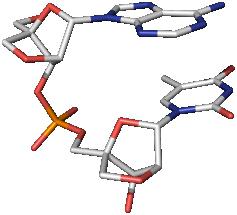
Tarkasteltavien yhdisteiden polymeeri- ja monomeerimuodot löytyvät soluista. Polymeerisia muotoja kutsutaan polynukleotideiksi. Tässä muodossa nukleotidiketjut sitovat fosforihappotähdettä. Kahden tyyppisen heterosyklisen molekyylin, nimeltään riboosi ja deoksoriboosi, pitoisuuden vuoksi hapot ovat vastaavasti ribonukleiini- ja deoksiribonukleiinisia. Niiden avulla tapahtuu perinnöllisten tietojen varastointi, siirto ja toteutus. Nukleiinihappojen monomeerisistä muodoista suosituin adenosiinitrifosfaattihappo. Hän osallistuu solun signaalin siirtoon ja energian varastointiin.
DNA-
Deoksiribonukleiinihappo on makromolekyyli. Sen avulla geneettisen tiedon siirto ja toteutusprosessi. Nämä tiedot ovat välttämättömiä elävän organismin kehittämis- ja toimintaohjelmalle. Eläimissä, kasveissa, sienissä DNA on osa kromosomeja, jotka sijaitsevat solun tumassa, samoin kuin mitokondrioissa ja plastideissa. Bakteereissa ja arkistoissa deoksiribonukleiinihapon molekyyli tarttuu solukalvoon sisältä. Tällaisissa organismeissa on pääasiassa pyöreitä DNA-molekyylejä. Niitä kutsutaan "plasmideiksi". Kemiallisen rakenteen mukaan deoksiribonukleiinihappo on nukleotideista koostuva polymeerimolekyyli. Nämä komponentit puolestaan koostuvat typpipitoisesta emäksestä, sokerista ja fosfaatista. Kahden viimeisen elementin kautta muodostetaan yhteys nukleotidien välille, joka luo ketjuja. Yleensä DNA-makromolekyyli on esitetty kaksiketjuisena heliksinä. 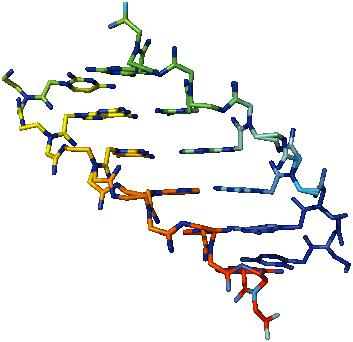
RNA
Ribonukleiinihappo on pitkä ketju, joka koostuu nukleotideista. Ne sisältävät typpipohjaa, riboosisokeria ja fosfaattiryhmää. Geneettinen informaatio koodataan käyttäen nukleotidisekvenssiä. RNA: ta käytetään proteiinisynteesin ohjelmointiin. Ribonukleiinihappo syntyy transkription aikana. Tämä on DNA-templaatin RNA-synteesin prosessi. Se tapahtuu erityisten entsyymien mukana. Niitä kutsutaan RNA-polymeraaseiksi. Tämän jälkeen matriisi ribonukleiinihapot osallistuvat käännösprosessiin. Niin on myös proteiinisynteesin toteuttaminen RNA-matriisilla. Ribosomit osallistuvat aktiivisesti tähän prosessiin. Jäljelle jääneet RNA: t käyvät läpi kemialliset muutokset transkription lopussa. Näiden muutosten tuloksena muodostuu sekundaarisia ja tertiäärisiä ribonukleiinihapporakenteita. Ne toimivat RNA-tyypin mukaan.
Sivu 1
Nukleiinihappojen biologinen rooli on se, että DNA tallentaa kehon perinnöllisen informaation deoksiribonukleotidien sekvenssinä, jotka eroavat typpipohjaisissa emäksissä. Kaikkien kehon proteiinien DNA on koodattu DNA: ssa. Jokaisella aminohapolla, joka on osa proteiineja, on oma koodi DNA: ssa, nimittäin kolme spesifistä nukleotidia. RNA-molekyylit siirtävät tietoa DNA: sta solun paikkoihin, joissa tapahtuu proteiinisynteesi.
Nukleiinihappojen biologinen rooli alkoi ilmaantua 40-luvun lopulla ja 50-luvun alussa, kun ensin havaittiin, että yhdestä bakteerilajista otettu ja toiseen lajiin tuodut DNA aiheuttaa jälkeläisiä ensimmäisen lajin piirteillä. Tästä seurasi, että perinnölliset tiedot siirrettiin yhdessä DNA: n kanssa - jollain tavalla koodattu järjestys tietyntyyppisten proteiinimolekyylien rakentamiseksi. Näistä teoksista tuli alkuvaihe molekyyligeeniikan nopealle etenemiselle, mikä toi meidät lähemmäksi proteiinisynteesin prosessia soluissa, solujen lisääntymistä jakamalla ja lopulta toistamalla koko monimutkainen eläin- tai kasviorganisaatio muodossa, joka on ominaista tämän organismin vanhemmille. Yksityiskohtainen keskustelu näistä ongelmista johtaisi meidät pitkälle biokemian alaan, mutta yleensä DNA: n ja RNA: n rooli näyttää seuraavalta. DNA-molekyylit sijaitsevat solunytimissä, ne sisältävät perinnöllistä informaatiota eri nukleotidisekvenssin muodossa. DNA: lla on rooli matriisissa, jolla on painettu RNA-molekyylien kopiot, jotka ovat suoraan mukana proteiinien synteesissä. Siten RNA-molekyylit toimivat lähettiminä DNA: sta solupaikkoihin, joissa proteiinisynteesi suoritetaan suoraan.
Nukleiinihappojen biologinen rooli alkoi kehittyä vuosisadan loppupuolella ja 50-luvun alussa, kun ensin todettiin, että yhdestä bakteerilajista otettu ja toiseen lajiin tuotu DNA aiheuttaa jälkeläisiä, joilla on ensimmäisen lajin ominaisuudet. .
Nukleiinihappojen biologinen rooli alkoi kehittyä vuosisadan loppupuolella ja 50-luvun alussa, kun ensin todettiin, että yhdestä bakteerilajista otettu ja toiseen lajiin tuotu DNA aiheuttaa jälkeläisiä, joilla on ensimmäisen lajin ominaisuudet. . Tästä seurasi, että DNA: n ohella siirrettiin perinnöllisiä tietoja - järjestys tietyntyyppisten proteiinimolekyylien rakentamiseksi.
Mutta ennen kuin ryhdymme tarkastelemaan nukleiinihappojen biologista roolia, on välttämätöntä pysyä niiden koostumuksessa ja rakenteessa. Tämä on erityisen tärkeää, koska nukleiinihappojen biologiset ominaisuudet johtuvat pääasiassa niiden kemiallisesta koostumuksesta ja rakenteesta.
Nukleiinihappojen biologinen rooli alkoi kehittyä tämän vuosisadan 40-luvulla, kun ensin todettiin, että yhdestä bakteerilajista otettu ja toiseen lajiin tuodut DNA pakottaa jälkimmäisen tuottamaan piirteen, jolla on ensimmäisen lajin ominaisuudet. Tästä seurasi, että perinnölliset tiedot siirrettiin yhdessä DNA: n kanssa - jollain tavalla koodattu järjestys tietyntyyppisten proteiinimolekyylien rakentamiseksi.
Nukleiinihappojen biologinen rooli alkoi ilmaantua tämän vuosisadan 40-luvulla, kun ensin todettiin, että yhdestä bakteerilajista otettu ja toiseen lajiin tuodut DNA aiheuttaa jälkimmäisten tuottamisen ensimmäisen lajin ominaisuuksilla. Tästä seurasi, että perinnölliset tiedot siirrettiin yhdessä DNA: n kanssa - jollain tavalla koodattu järjestys tietyntyyppisten proteiinimolekyylien rakentamiseksi.
Muutama vuosikymmen sitten nukleiinihappojen biologinen rooli oli täysin epäselvä, mutta tällä hetkellä niiden ensisijainen merkitys luonnonvaraisissa eläimissä on luotu.
Saksalainen tiedemies Misher löysi noin 100 vuotta sitten vuonna 1869 - 1870 nukleiinihappoja, mutta mielenkiintoisimmat tiedot niiden rakenteesta, ominaisuuksista ja biologisesta roolista saatiin viimeisten 10 - 15 vuoden aikana, kun biokemiassa ja biologiassa ne yleistyivät. soveltaa uusimpia fysikaalisia ja kemiallisia tutkimusmenetelmiä. Nämä tutkimukset ovat paljastaneet nukleiinihappojen tärkeimmän biologisen roolin ja osoittaneet, että näiden happojen biologinen rooli riippuu niiden koostumuksesta ja rakenteesta.
Paljon monimutkaisempi on nukleotidien sekvenssin määrittäminen nukleiinihappojen polymeeriketjussa. Tätä kysymystä, joka on äärimmäisen tärkeä nukleiinihappojen biologisen roolin tutkimukselle, ei ole vielä kehitetty riittävästi. Tämän ongelman ratkaisemiseksi on tarpeen löytää selektiivisiä menetelmiä nukleiinihappojen makromolekyylin jakamiseksi, joka on nyt yksi tämän luokan yhdisteiden kemian tärkeimmistä tehtävistä. Tällä hetkellä nukleotidien sekvenssi määritetään vain yhdelle pienimolekyylipainoiselle ribonukleiinihapolle.
Tietojemme nykyinen tila ei anna meille lupaa lopulta päättää, onko nukleiinihapoilla lajispesifisyys. Samaan aikaan nukleiinihappojen biologisen roolin arviointi riippuu tämän ongelman ratkaisusta.
Nykyään on vaikea nimetä luonnontieteiden aluetta, joka ei olisi kiinnostunut nukleiinihappojen rakenteen ja toimintojen ongelmasta. Huolimatta viime vuosikymmeninä tapahtuneesta valtavasta edistymisestä nukleiinihappojen kemiallisen koostumuksen ja rakenteen tutkimuksessa on vielä ratkaistava monia ongelmia selventääkseen nukleiinihappojen rakenteen ja biologisen roolin välistä suhdetta. Ei ole epäilystäkään siitä, että nukleiinihappojen tutkimisen tällä tiellä on löydöksiä, jotka ovat erittäin tärkeitä biologian, lääketieteen ja koko elintason kannalta. Nukleiinihappojen komplementaarisuusperiaatteen epohakuinen löytäminen mahdollisti tunkeutumisen näiden biopolymeerien hienon rakenteen salaisuuksiin, mutta myös biologisten makromolekyylien synteesin ja lisääntymisen mekanismeihin. Nukleiinihapot suorittavat useita tärkeitä biologisia toimintoja, jotka eivät ole ominaista muille polymeerisille aineille. Erityisesti ne tarjoavat perinnöllisen informaation tallennusta ja siirtoa ja ovat suoraan mukana mekanismeissa tämän informaation toteuttamiseksi ohjelmoimalla kaikkien solun proteiinien synteesi. Nukleiinihappojen rakennekomponentit toimivat myös kofaktorina (koentsyymi A, uridiinidifosfaatti glukoosi jne.), Allosteeriset tehostajat, ovat osa koentsyymejä (nikotiiniamidideniinidinukleotidi, flavinadeniinidinukleotidi jne.), Mikä siten myös suoraan metaboliaan. energian kertymiseen (kertymiseen), siirtoon ja muuntamiseen. Ne ovat toissijaisten lähettäjien (sanansaattajien), syklisten mononukleotidien (cAMP ja cGMP) esiasteita, joilla on tärkeä rooli solunsisäisten signaalien siirrossa.
Viime vuosisadalla havaittiin, että solunytyt sisältävät suurimolekyylisiä aineita, jotka sisältävät typpipitoisia heterosyklisiä emäksiä, hiilihydraatteja ja fosforihappoa. Kaksi vuosikymmentä sitten nukleiinihappojen biologinen rooli oli täysin epäselvä, mutta tällä hetkellä niiden ensisijainen merkitys villieläimissä on luotu.
Nukleiinihapot - deoksiribonukleiini ja ribo nukleiinihapot- kaikkien elävien organismien yleismaailmalliset komponentit, jotka ovat vastuussa geneettisen tiedon tallennuksesta, siirrosta ja lisääntymisestä (toteutumisesta). Kaikilla kahdella tyypillä nukleiinihapotjaettuna molekyylien hiilihydraattikomponentilla: deoksiribroosi deoksiribonukleiinihapoissa (DNA) ja riboosi ribonukleiinihapoissa (RNA). DNA: n biologinen rooli useimmissa organismeissa on geneettisen informaation tallentamisessa ja lisääntymisessä, ja RNA: n tämän informaation toteuttamisessa molekyylien rakenteessa proteiineja synteesin prosessissa.
Nukleiinihapot löydettiin vuonna 1868 g. Sveitsiläinen tiedemies F. Miescher, joka totesi, että nämä aineet ovat paikallisia solujen ytimissä, sisältävät happamia ominaisuuksia ja toisin kuin proteiinit sisältävät fosforia. kemiallisesti nukleiinihapotovat polynukleotidit, ts. monomeerisistä yksiköistä rakennetut biopolymeerit - mononukleotidit tai nukleotidit (niin sanottujen nukleosidien fosforihappoesterit - puriinin ja pyrimidiinipitoisten emästen johdannaiset, D-riboosi tai 2-deoksi-D-riboosi). DNA-molekyyliin sisältyvät puriiniemäkset ovat adeniini (A) ja guaniini (G), pyrimidiini-sytosiini (C) ja tymiini (T). RNA: n nukleosideissa urasiili (U) on läsnä tymiinin sijasta. Polynukleotidiketjussa nukleotidit on liitetty fosfodiesterisidoksen kautta ( riisi. 1 ).
Ensisijainen rakenne nukleiinihapotmääritetään typpipohjaisten vuorottelujen järjestyksessä, ja niiden spatiaalinen konfiguraatio määritetään molekyylin kohtien välisten ei-kovalenttisten vuorovaikutusten avulla: typpipohjaisten vesistöjen väliset sidokset, emäsparien tasojen hydrofobiset vuorovaikutukset, sähköstaattiset vuorovaikutukset osallistumalla negatiivisesti varautuneisiin fosfaattiryhmiin ja -ionioihin.
deoxyribo nukleiinihapoterilaisista organismeista eristetyt eroavat niissä olevien typpipohjaisten emästen suhteen, ts. nukleotidikoostumuksen mukaan, joka kaikelle DNA: lle noudattaa Chargaff-sääntöä: 1) adeniinimolekyylien lukumäärä molekyylissä N on yhtä suuri kuin tymiinimolekyylien lukumäärä, so. A = T; 2) guaniinimolekyylien lukumäärä on yhtä suuri kuin sytosiinimolekyylien lukumäärä, ts. G = C; 3) puriiniemästen molekyylien määrä on yhtä suuri kuin pyrimidiiniemästen molekyylien lukumäärä; 4) 6-aminoryhmien lukumäärä on yhtä suuri kuin 6-ketoryhmien lukumäärä, mikä tarkoittaa, että adeniini + sytosiinin summa on yhtä suuri kuin guaniini + tymiinin summa, ts. A + C = G + T. Chargaffin sääntö on voimassa myös ns. Pieniä typpipohjaisia emäksiä (metyloidut tai muut puriini- ja pyrimidiiniemästen johdannaiset). Siten kunkin DNA: n nukleotidikoostumukselle on tunnusomaista vakioarvo - moolisuhde (spesifisyyskerroin) tai G-C-parien prosenttiosuus, so. ![]() . Viimeisen indikaattorin arvo on lähes sama saman luokan organismeille. Korkeammissa kasveissa ja selkärankaisissa se on 0,55-0,93.
. Viimeisen indikaattorin arvo on lähes sama saman luokan organismeille. Korkeammissa kasveissa ja selkärankaisissa se on 0,55-0,93.
Vuonna 1953 amerikkalainen biokemisti Watson (JD Watson) ja englantilainen biofyysikko Crick (FN Crick), jotka perustuivat Chargaff-sääntöön ja DNA-molekyylien röntgensädeanalyysiin, totesivat, että DNA-molekyylillä on kaksoiskierre, jossa kaksi rinnakkain suunnattua hiilihydraattia - fosfaatiketjut pidetään adeniinin ja tymiinin sekä guaniinin ja sytosiinin välisten vety- sidosten avulla. Yhdessä ketjussa olevien typpipohjaisten emästen sekvenssi määrittää niiden sekvenssin toisessa nukleotidiketjussa komplementaaristen parien А-Т ja Г-sizes koot ovat samat, mikä sallii nukleotidiketjun taittua tavanomaiseksi kaksoiskierrokseksi, joka määrittää DNA: n sekundäärirakenteen. Tällaisen kierteen kierrosta kohti on 10 perusparia.
Geneettistä informaatiota "salataan" DNA-molekyylissä nukleotidien yhdistelmissä, jotka koodaavat tiettyjen aminohappojen sisällyttämistä rakenteessa olevaan polypeptidiketjuun. On todettu, että geneettinen koodi on yleinen kaikille eläville tavaroille; tripletti, ts. kukin aminohappo koodaa nukleotidikolmo (tripletti); ei päällekkäisyyttä, ts. tietty nukleotidi voi päästä vain yhteen niin kutsuttuun kodoniin; yksi aminohappo voidaan koodata useita triplettejä; useimmilla nukleotiditripleteillä on "merkitys", ts. koodaa aminohappoja.
DNA-molekyylien röntgensädeanalyysi osoitti, että nukleotidin puriini- ja pyrimidiiniemäkset sijaitsevat yhdessä tasossa, joka on lähes kohtisuorassa molekyylin pitkittäisakseliin nähden (ns. DNA-kaksoiskierteen niin sanottu B-muoto, jota sillä on fysiologisissa olosuhteissa), kun taas deoksiriboosijäämät ovat tasossa, joka on lähes kohtisuorassa taso, jossa typpipohjat ovat.
synteesi nukleiinihapotsolussa se suoritetaan matriisimolekyylin kopioinnin periaatteen mukaisesti nukleosiditrifosfaattien polykondensaatioreaktiolla pyrofosfaatin pilkkomisella. Tätä prosessia katalysoi polymeraasientsyymit. Typpikaasujen sekvenssi rakenteessa olevassa molekyylissä määritetään niiden sekvenssillä matriisimolekyylissä. DNA-molekyylien synteesiä kutsutaan replikaatioksi (reduplication), so. replikaattimolekyylien muodostuminen perusmolekyyliin.
Geneettistä informaatiota solusta soluun, sukupolvelta toiselle, välitetään tarkasti DNA-molekyylien replikoinnilla (kaksoiskappaleella), minkä seurauksena yhdestä DNA-molekyylistä muodostuu kaksi sidoksissa olevaa, täysin identtistä äitiä, mikä takaa perinnöllisen informaation täydellisen kompleksin siirtämisen. Reduplikaatioprosessissa nukleotidiparien välillä on rikkoutunut vety- sidoksia, deoksinukleosiditrifosfaatit, jotka sisältävät komplementaarisia typpi-emäksiä, sitoutuvat vapautuneisiin nukleotideihin. Näin ollen kukin toissijainen kaksoiskierre sisältää yhden äidin ja yhden vasta syntetisoidun polynukleotidiketjun. Replikointi on monimutkainen prosessi, johon liittyy monia entsyymejä, proteiini, joka erottaa DNA-säikeet, nukleaasit, ligaasit ja muut proteiinit. Matriisi-DNA: ta tarvitaan sen toteuttamiseen; kaikkien neljän typpipohjan desekä Mg2 + -ionien ( riisi. 2 ).
Uuden ketjun kasvua katalysoi DNA-polymeraasientsyymi. Replikointi alkaa (aloitetaan) tietyissä DNA-molekyylin osissa - replikaattoreissa, joiden ensisijainen rakenne on ominaista korkealle sisällölle höyry AT ja ns. käänteisten toistojen (palindromeja) läsnäolo. Replikoinnin lopettaminen (päättyminen) tapahtuu joko kahden replikoinnin haarukan yhdistämisessä tai ns. Päätepisteissä. Bakteereissa ja eukaryooteissa jokaisen solunjakautumisjakson aikana kaikki DNA: n täytyy yleensä replikoitua ja jopa kerran, siksi on oltava mekanismeja, joilla ohjataan replikaation aloittamista ja mekanismeja, joilla äiti ja tytärmolekyylit eroavat toisistaan. Joskus (normaali ja patologia) voi esiintyä koko genomin useita DNA-replikaatioita (ks. Ks. \\ T geeni) ilman myöhempää solunjakoa, joka johtaa polyploidisten ytimien muodostumiseen tai genomin yksittäisten osien replikoitumiseen ilman koko genomin replikaatiota (ns. ylimääräinen replikaatio). Kuvaillaan tapauksia, joissa osaa DNA: n osaa siitä genomin osasta eukaryoottisoluissa, joissa ei ole geenejä, jotka ovat välttämättömiä solun ylläpitämiseksi. Replikoitumisvaiheita katalysoivien entsyymien samankaltaisuus ja prokaryooteissa ja eukaryooteissa replikoinnin haarassa esiintyvät tärkeimmät prosessit osoittavat, että DNA: n replikaation suuri evoluutiovakavuus ja tiukka geneettinen kontrolli.
DNA: n hajoaminen, kuten RNA, ei ole normaalia elävissä soluissa. Kuolleiden solujen DNA: ta tai soluja, joiden seinämän eheys on rikki, pilkkovat spesifiset nukleaasit (DNaasi I ja DNaasi II), jotka katalysoivat poly- tai oligonukleotidien sisäisten nukleotidisidosten hajoamista ilman epäorgaanisen fosfaatin muodostumista. Nukleaasin vaikutuksen luonteen mukaan fosfodiesteraasi. Nukleaasien merkitys geneettisessä rekombinaatiossa (ks Geenitekniikka), geneettisten vaurioiden korjaus (korjaus), solujen suojaaminen ulkomaalaiselta nukleiinihapoterittäin suuri. Niiden aktiivisuus kudoksissa ja biologisissa nesteissä voi olla diagnostinen testi useille sairauksille. Siten DNA-ase II: n aktiivisuus seerumissa lisääntyy akuutin haimatulehduksen, erityisesti hemorragisen haimatulehduksen myötä, ja RNA-asen aktiivisuus seerumissa lisääntyy uremian myötä. Joidenkin nukleaasien geneettisesti aiheuttama puutos on syy vakavaan perinnöllisiä sairauksia (esimerkiksi pigmentti xeroderma).
Substraatin hyökkäysmenetelmän mukaan nukleaasi on jaettu ekso- ja endonukleaaseihin. Eksonukleaasit katalysoivat mono- tai oligonukleotidien peräkkäistä katkaisua yhdestä polynukleotidiketjun päästä. Jotkut eksonukleaasit katalysoivat sekä DNA: n että RNA: n pilkkoutumista.
Endonukleaasit katalysoivat aukon polynukleotidiketjun sisäisten linkkien välillä ja niillä on paljon suurempi substraattispesifisyys kuin eksonukleaaseilla.
Heikkenee nukleiinihapotnukleosidaasit, jotka katalysoivat fosmononesterisidosten katkeamista mononukleotideissa nukleosidien ja epäorgaanisen fosfaatin muodostamiseksi, osallistuvat myös niiden luonteen mukaan fosfo- monesteraseihin ja hydrolyyttisiin ja fosforolyyttisiin nukleosidaaseihin (nukleosidihydrolaaseihin ja nukleosidifosforylaaseihin).
Ribot nukleiinihapot useimmat organismit tuottavat geneettistä informaatiota, mutta RNA: ta sisältäville viruksille ne voivat olla myös perinnöllisten tietojen, kuten DNA: n, kantajia.
RNA-molekyyli on lineaarinen polymeeri, jonka monomeeriset yksiköt ovat ribonukleotideja (niiden hiilihydraattikomponentti on pentoosi-D-1-riboosi). Joidenkin RNA: iden tyypillisiä rakenteellisia elementtejä edustavat pienet emäkset (vastaavat nukleotidit ovat osa joukkoa RNA: ta hyvin pieninä määrinä).
RNA: n ensisijainen rakenne on tiukasti spesifinen ja ainutlaatuinen jokaiselle luonnolliselle RNA: lle. Se toimii muodossa biologisen informaation tallentamiseen, joka toistetaan ja toistetaan toistuvasti RNA-biosynteesin prosessissa. DNA-molekyyliin matriisina rakennetun syntetisoidun RNA: n rakenne (transkription prosessi) määritetään tällä DNA-molekyylillä, joka on sen polynukleotidiketjuun koodatun geneettisen informaation toteutuksen alkuvaihe. RNA-transkription synteesiä katalysoivat RNA-polymeraasit, jotka lukevat vain yhden, niin sanotun DNA-templaatin kaksoiskierroksen niin sanotun merkittävän juosteen. Transkriptioprosessi tuottaa vastaavan DNA: n tai geenin RNA-kopion RNA-kopion.
RNA-molekyylin sekundääriset ja tertiääriset rakenteet (sen spatiaalinen konfiguraatio) sekä DNA-molekyylit muodostuvat pääasiassa vetysidosten ja typpipohjaisten hydrofobisten vuorovaikutusten vuoksi. Kuitenkin, jos stabiili kaksisäikeinen helix on ominaista DNA-molekyylille, niin RNA-molekyylien sekundääriset ja tertiääriset rakenteet ovat paljon labiilempia ja muuttuvia. RNA-polynukleotidiketjut ovat hyvin joustavia. RNA-molekyylit käyttäytyvät tyypillisten polyelektrolyyttien tavoin liuoksissa, joilla on alhainen ionivahvuus; kun liuoksen ionivahvuus kasvaa, RNA: n turvonnut ketjut puristetaan joissakin joustavan RNA-ketjun osissa, jotka kiertyvät, kiertyvät itsensä ympärille, muodostuu kaksisäikeisistä rakenteista niin sanotun komplementaarisen parin tuloksena, kuten DNA-molekyyleissä. Sellaiset RNA-molekyylien rakenteet stabiloidaan vety- sidoksilla vastakkaisten typpipohjaisten emästen välillä antiparalleeliketju- paikoissa. RNA-molekyylin kaksisäikeisissä osissa olevat typpipohjaiset spesifiset parit ovat, kuten DNA-molekyylissä, A-U, G-C ja G-U (urasiili tymiinin sijasta). Joissakin viruksissa esiintyy RNA-molekyylejä, jotka koostuvat kahdesta komplementaarisesta polynukleotidiketjusta; Lisäksi ne muodostetaan välimuotoina useiden virus-RNA: iden biosynteesin aikana (niin kutsutut RNA: n replikatiiviset muodot).
Jotkut kaksisäikeiset RNA: t, kuten DNA, voivat esiintyä rengasmolekyyleinä, ja jos molemmat polynukleotidiketjut ovat suljettuina kovalenttisesti, ne kykenevät muodostamaan supercoiled-renkaita. RNA voi muodostaa kaksijuosteisia komplekseja, joissa yksi juoste on esitetty polyribonukleotidilla ja toinen. Sellaiset DNA-RNA-hybridikompleksit muodostetaan DNA-replikaation aikana niin kutsutun siemen-RNA-fragmenttien, samoin kuin RNA: n transkription DNA-templaatissa. DNA-RNA-hybridikompleksit syntyvät myös solujen infektoinnin jälkeen tietyillä RNA: ta sisältävillä viruksilla sen DNA: n viraalisen RNA: n synteesin seurauksena, joka on komplementaarinen sen kanssa entsyymin käänteistranskriptaasin (revertase) avulla.
RNA: n pitoisuus elävissä soluissa (siittiöitä lukuun ottamatta) on merkittävästi suurempi kuin DNA: n pitoisuus. Suurin osa RNA: sta sijaitsee solun sytoplasmassa: todellisten sytoplasmisten ribosomien koostumuksessa (ks. solu) ja mitokondriot, ja se on läsnä myös ei-ribosomaalisten kompleksien muodossa proteiinien kanssa. Kromatiini tulee RNA-ytimeen (osa ydin-RNA: sta on nykyisen geenin transkription tuote).
RNA: n toiminnot solussa ovat monimutkaisia ja monipuolisia. RNA: n kolme päätyyppiä: ribosomaalinen RNA (rRNA), kuljetus RNA (tRNA) ja messenger-RNA (mRNA tai mRNA). Soluissa on myös joukko ns. Pieniä RNA: ita, joiden toimintoja ei ole vielä selvitetty.
Ribosomaaliset RNA: t muodostavat noin 80% kaikista solun RNA: sta, 50-65% RNA: n kokonais ribosomimateriaalista. Kussakin ribosomi-alayksikössä (suuri ja pieni) RNA toimii luurankona, johon on koottu ribosomaalisia proteiineja. Muodostunut ribonukleoproteiinikompleksi (ns. Ribonukleoproteiini tyas-RNP-tyazh) on järjestetty monimutkaiseksi kompaktiksi hiukkaseksi - todelliseksi ribosomaaliseksi alayksiköksi. RRNA: n rooli solun proteiinisynteesijärjestelmässä ei ole tyhjentynyt sen rakenteellisilla toiminnoilla. Uskotaan, että jotkut rRNA: n osat vaikuttavat ribosomin peptidyylitransferaasikeskuksen muodostumiseen, joka vastaa peptidisidosten muodostumisesta aminohappotähteiden välillä proteiinisynteesin aikana.
Kuljetus RNA: t muodostavat noin 15% solujen RNA: iden kokonaismäärästä. TRNA: n nukleotidiketju sisältää vain 75-90 nukleotidia, suurin osa tRNA: sta muodostuu ketjussa olevien nukleotidien täydellinen sekvenssi. TRNA: n erityispiirre on suhteellisen korkea nukleotidien pitoisuus, mukaan lukien pienet typpipohjaiset emäkset. nämä nukleiinihapoterittäin spesifisten entsyymien, aminoasyyli-tRNA-synteesien avulla kiinnitä yksi tai toinen aminohappo itselleen ja siirrä se ribosomiin. Samaa aminohappoa varten on useita tRNA: ita, joita kutsutaan iso-reseptoriksi. Proteiinin polypeptidiketjun synteesin aikana kuljetus-RNA "tunnistaa" spesifisen aminoasyyli-tRNA-syntetaasin, ottaa siitä aktivoidun aminohapon, kiinnittyy mRNA: han ribosomilla ja takaa siten tiukan spesifisyyden aminohappojen valinnan ja integroinnin kasvavaan proteiinimolekyyliin; koulutuksen jälkeen peptidisidos toimitetun aminohapon ja tRNA: n jo rakennetun polypeptidiketjun välillä tämä ketju pysyy ribosomilla.
Informatiivinen tai messenger, RNA, muodostaa vain 5% solun RNA: n kokonaismäärästä. Niiden biologinen rooli on ohjelmoida kaikkien soluproteiinien synteesi. Jos rRNA ja tRNA kuuluvat solun proteiinisynteesijärjestelmän "palvelevaan" laitteeseen, mRNA on suora välittäjä DNA: n ja proteiinien välillä ja toimii templaattina proteiinien synteesille; MRNA: n muodossa käännetään suurin osa DNA-tiedot.
Polynukleotidiketjun kemiallisen rakenteen mukaisesti kvantitatiivista määritystä varten on kolme ryhmää menetelmiä nukleiinihapot: typpipitoisuuksien (spektrofotometrinen määritys), hiilihydraattikomponentin (spesifisten värireaktioiden) mukaan fosforin määrän mukaan. Toisen ryhmän menetelmät ovat spesifisiä tyypille N. k ja sallia erottaa DNA RNA: sta.
Yksittäisten nukleotidisekvenssien määrittäminen nukleiinihapoton suuri käytännön etu. Tämä on erityisesti tapa tunnistaa mutanttigeenit ja selittää epänormaalien proteiinien synteesin taustalla olevat molekyylimekanismit perinnöllisen patologian eri muodoissa (ks. Perinnölliset sairaudet).
viitteet.: Gaytskhski V.S. Eläinten solujen informatiivinen RNA. M., 1980; Rolan ZH.K., Selosh A. ja Selosh D. Atlas solubiologiasta, trans. ranskaksi, s. 28, M., 1978; Spirin A.S. ja Gavrilova L.P. Ribosome, M., 1968; Watson, J.D., geenin molekyylibiologia, trans. Englanti, M., 1978; Shabarova Z.A. ja Bogdanov L.A. Nukleiinihappojen ja niiden komponenttien kemia.