Yksilöllinen kehitys(ontogeneesi), ontogeneesin periodisointi.
Kaikilla organismeilla on tietty elinkaari. Seksuaalisesti kehittyvillä organismeilla se alkaa tsygootin ilmestymishetkestä ja päättyy organismin luonnolliseen kuolemaan.
Organismien elinkaaren aikana tapahtuvien prosessien kokonaisuus määritellään yksilön kehitykseksi tai ontogeneesiksi.
Ontogeny sisältää 3 jaksoa:
1 jakso. Preembryonaalinen tai gametogeneesi.
2 jaksoa. Alkionaikainen.
3 jaksoa. Postembryonic.
1. Organismien elinkaaret. Kehitys on toukkaa ja suoraa.
Organismien kehitys on suoraa ja epäsuoraa transformaation kanssa.
epäsuora kehitys tapahtuu toukkavaiheen kautta. Toukassa muodostuu tiettyjä itu- tai väliaikaiselimiä, jotka tarjoavat organismin elintärkeän toiminnan tässä kehitysvaiheessa.
Korkeammilla selkärankaisilla kehitys on suoraa, mutta myös tilapäisiä elimiä muodostuu alkionkehityksen aikana. Nisäkkäillä nämä ovat alkion kalvot (amnion, suonikalvo, allantois, istukka) ja keltuainen pussi.
2. Kehityksen esialkion aika (esi-alkio, progenesis). Gametogeneesin vaiheet. Alkion varhaiseen kehitykseen liittyvät muutokset oogeneesissä (geenin monistuminen, munasolujen segregaatio, ravinteiden kertyminen).
Preembryonaalinen ajanjakso tai gametogeneesi sisältää useita vaiheita: eristäminen, lisääntyminen, kasvu, kypsyminen, muodostuminen (jälkimmäinen vain siittiöissä).
Oogeneesin aikana tapahtuvat tärkeimmät tapahtumat, jotka ovat välttämättömiä tulevan organismin kehitykselle.
1 tapahtuma. Oogeneesin aikana tapahtuu rRNA-geenien monistumista tai rRNA:sta vastuussa olevien geenien kopioiden lukumäärän kasvua. Tämä prosessi tapahtuu meioosin 1 profaasissa. rRNA-geenien kopioita voi olla jopa miljoona.
Sitten nämä kopiot erotetaan kromosomeista, kelluvat vapaasti karyoplasmassa, niiden ympärille muodostuu nukleoleja ja ribosomialayksiköitä syntetisoidaan nukleoleissa, jotka tulevat sytoplasmaan. Siten ribosomien määrä kasvaa jyrkästi etukäteen munassa.
2 tapahtuma. Oogeneesin aikana erityyppisiä mRNA:ta syntetisoidaan meioosin 1 profaasin aikana. Transkriptioprosessit tapahtuvat despiralisoiduissa kromosomien osissa. Meioosin 1 profaasivaiheessa olevia kromosomeja kutsutaan lamppuharjakromosomeiksi.
3 tapahtuma. Ravinteet varastoituvat munaan keltuaisena.
4 tapahtuma. Munalle on ominaista ooplasminen segregaatio, eli aineiden jakautuminen koko munan sytoplasmaan, mikä johtaa sytoplasman kemialliseen heterogeenisyyteen. Uskotaan, että tämä on välttämätöntä solujen varhaiselle erilaistumiselle.
5 tapahtuma. Sukupuolisolut ovat kehon erityisiä soluja, koska niillä on totipotenssi eli yhtäläinen perinnöllisyys. Vain sukusolut, samoin kuin ihmisen blastomeerit, 2 blastomeerin vaiheessa synnyttävät kaikentyyppisiä soluja. Esimerkiksi kokeet blastomeerien erottamisesta, silmukoitumisesta tai sekoittamisesta katkaisuvaiheessa ovat osoittaneet, että lajeissa, joissa on säteittäinen katkaisutyyppi, useiden sukupolvien blastomeerit, jos ne on eristetty ja sijoitettu sopiviin olosuhteisiin, osoittavat totipotenssia, ts. kehittyä täydelliseksi organismiksi. Ihmisalkioiden solujen yhtäläisestä perinnöllisyydestä ja totipotenssista 2–4 blastomeerin vaiheeseen asti puhuvat tapaukset, joissa on syntynyt kaksi, kolme, neljä identtistä kaksoset.
3. Alkion kehitysvaihe, periodisointi.
Ontogeneesin alkion jakso sisältää useita vaiheita:
1 lannoitusvaihe.
Tsygootin 2. vaihe.
3 Murskausvaihe (yksikerroksisen alkion muodostuminen).
4 Gastrulaation vaihe (kaksi- ja kolmikerroksisen alkion muodostuminen).
5 Vaihe histo- ja NOIN rganogeneesi (kudosten ja elinten muodostuminen).
4. Hedelmöityminen ja tsygootin muodostuminen. Nisäkkäiden ja ihmisten hedelmöityksen ominaisuudet.
lannoitusvaihe Tämä on munasolun ja siittiön fuusioprosessi, jonka seurauksena muodostuu diploidinen tsygootti, josta kehittyy diploidi organismi. Tässä prosessissa on 3 vaihetta:
1 vaihe- sukusolujen lähentyminen. Tässä munasolun ja siittiöiden erittämillä aineilla on tärkeä rooli. Niitä kutsutaan gamoneiksi (sukusolujen, gynogamonien ja androgamonien hormoneiksi). Lisäksi on useita epäspesifisiä tekijöitä, jotka lisäävät siittiöiden kohtaamisen ja vuorovaikutuksen todennäköisyyttä munasolun kanssa. Nämä sisältävät
miesten ja naisten hedelmöitysvalmiuden alkamisen koordinointi,
urosten ja naaraiden käyttäytyminen, joka varmistaa parittelun ja siemennyksen,
ylimääräinen siittiöiden tuotanto
iso munasolu,
gamonien läsnäolo, jotka edistävät sukusolujen lähentymistä ja vuorovaikutusta,
paritteluelinten läsnäolo, jotka tarjoavat sisäisen keinosiemennyksen.
Nisäkkäillä hyvin tärkeä sillä on siittiöiden viipymistä naaraan sukupuolielimissä, minkä seurauksena miehen sukusolut saavat hedelmöityskyvyn, ts. kyky akrosomaaliseen reaktioon.
Vaihe 2 - sukusolujen aktivaatio tapahtuu niiden kosketuksen jälkeen. Siittiöiden aktivoitumista kutsutaan akrosomaaliseksi reaktioksi. Munan aktivoituminen on aivokuoren reaktio.
Akrosomaalisen reaktion ydin : Siittiöiden läpäisevyys akrosomissa muuttuu plasmakalvon silta ja entsyymejä, spermatolysiinejä, vapautuvat akrosomista. Nämä entsyymit rentouttavat munasolua ympäröivien follikulaaristen solujen välisiä sidoksia. Siittiöt kulkevat follikulaaristen solujen kerroksen läpi, sitten pellucid-vyöhyke tuhoutuu ja siittiö kulkee tämän alueen läpi.
Kortikaalisen reaktion ydin: Se koostuu monimutkaisista rakenteellisista ja fysikaalis-kemiallisista muutoksista. Koska siittiösolun kalvo läpäisee natriumioneja, jälkimmäiset alkavat päästä munasoluun muuttuen kalvopotentiaali soluja. Sitten sukusolujen kosketuspisteestä etenevän aallon muodossa Ca 2+ -ionien pitoisuus lisääntyy (hyaloplasmassa ne poistuvat varastosta - EPS, retikulum) ja munassa käynnistyvät biokemialliset prosessit, jonka jälkeen aivokuoren rakeet myös liukenevat aaltoon. Samaan aikaan vapautuvat spesifiset entsyymit edistävät keltuaisen kalvon irtoamista; hän kovettaa sitä lannoitustuppi.
Yksi kortikaalisen reaktion merkityksistä on polyspermian ehkäisy, ts. useamman kuin yhden siittiön tunkeutuminen munasoluun. Nisäkkäillä kortikaalinen reaktio ei aiheuta hedelmöityskalvon muodostumista, mutta sen olemus on sama.
Munan aktivoituminen päättyy proteiinisynteesin alkamiseen translaatiotasolla, koska mRNA, tRNA, ribosomit ja energia varastoituvat jopa oogeneesissä.
3 vaihe- sukusolujen fuusio eli syngamia. Tässä tapauksessa siittiöön ja munasoluun muodostuu yhteinen plasmakalvo. Naisen ja miehen pronukleus lähentyvät ja sulautuvat yhteen (synkaryoni) muodostaen yhteisen metafaasilevyn. Tämä on sukusolujen lopullisen fuusion - syngamian - hetki.
Lannoituksen ominaisuudet eri organismilajeissa.
1 esimerkki. Nisäkkäillä ja ihmisillä siittiösolu sitoutuu munasoluun alueella, jossa zona pellucidassa on reseptori. Tämän vuorovaikutuksen jälkeen jäljellä olevat reseptorit estyvät.
2 esimerkki. klo merisiili hedelmöittymisen jälkeen munasolussa muuttuu dramaattisesti sähköinen potentiaali plasmakalvo, ja sitten muodostuu hedelmöityskalvo, joka estää polyspermian.
tsygoottivaihe. Tunkeutumisen jälkeen miehen ydintä kutsutaan urospronukleukseksi. Kromatiini löystyy siinä, DNA:n replikaatio tapahtuu. Naisen ydintä kutsutaan naisen esiytimeksi. Siinä tapahtuu samat tapahtumat. Nisäkkäillä ja ihmisillä ydinfuusiota ei tapahdu, mutta metafaasilevy muodostuu välittömästi.
5. Eläinten ja ihmisten munan keinosiemennys, biologiset ja lääketieteelliset näkökohdat.
Eläinten munien keinohedelmöityksellä on suuri tieteellinen merkitys lääketieteen kannalta, koska sen tutkimuksen aikana kehitetään tapoja ja mekanismeja ihmisten hedelmättömyyden hoitoon.
Keinotekoista siemennystä käytetään useita muotoja sekä miesten että naisten hedelmättömyys, jota on vaikea hoitaa. Esimerkiksi kun miehellä on liian vähän siittiöitä tai he ovat käytännössä liikkumattomia, kun naisen läpinäkyvyys on heikentynyt munanjohtimia tai sisäisissä sukupuolielimissä on muita vaurioita, joihin liittyy kumppanien immunologinen yhteensopimattomuus.
6. Yleiset luonteenpiirteet murskaamalla. Eri eläinlajeille ominaiset murskaustyypit. Blastulan pilkkoutuminen ja muodostuminen istukan nisäkkäillä.
murskausvaihe. Tämä on yksikerroksisen alkion - blastulan - muodostumisvaihe. Blastulan sisällä on onkalo, jota kutsutaan blastocoeliksi.
Murskausominaisuudet:
Solut jakautuvat mitoosilla.
Kunkin jakautumisen aattona tapahtuu DNA:n replikaatio.
Jakautuvat solut eivät kasva.
Murskaustapa riippuu munatyypistä.
Täydellinen tasainen murskaus lansetissa:
Ensimmäinen halkeamisura kulkee pystysuunnassa, muodostuu kaksi blastomeeria. Toinen ura kulkee myös pystysuorassa ja muodostuu neljä blastomeeria. Kolmas uurre kulkee vaakatasossa, muodostuu kahdeksan blastomeeria ja sitten vuorottelevat pysty- ja vaakaurat. 12 syklin jälkeen murskaus muuttuu asynkroniseksi. Tietyssä kehitysvaiheessa alkio on solukimpale tai morula. Sitten solujen väliin ilmestyy rakoja, ja muodostuu ontelo - blastokoel. Lansetissa murskaamisen aikana muodostuu blastula, jota kutsutaan coeloblastulaksi, eli yksikerroksiseksi palloksi.
Täydellinen epätasainen pirstoutuminen sammakkoeläimissä:
Sammakkoeläinsolut ovat kohtalaisen telolesitaaleja. Solun eläinnapalla pirstoutuminen etenee nopeammin kuin vegetatiivisessa napassa. Tämän seurauksena eläinnavan solut ovat pienempiä - mikromeereja. Vegetatiivisessa napassa solut ovat suurempia - makromeerit. Sammakkoeläinten blastulaa kutsutaan amfiblastulaksi. Blastocoel sijaitsee eläinpaalussa.
Murskaamisen ominaisuudet nisäkkäillä ja ihmisillä:
Halkeaminen on täysin epätasaista, asynkronista ensimmäisistä vaiheista lähtien; tietyssä kehitysvaiheessa alkio on morula (solupala). Sitten suuremmat solut erottuvat reuna-alueille muodostaen trofoblastin ja pienemmät solut keskelle muodostaen alkioblastin. Blastulaa kutsutaan blastokystiksi. Blastocoel on hyvin pieni. Trofoblasti edistää alkion siirtymistä kohdun limakalvolle. Tätä prosessia kutsutaan implantaatioksi. Embryoblasti synnyttää itse alkion ja joitain väliaikaisia elimiä.
7. Gastrulaation yleiset ominaisuudet. Sammakkoeläinten ja lintujen gastrulaation ominaisuudet. Gastrulaatio korkeammilla (istukan) nisäkkäillä.
Gastrulaatiovaihe, tai kaksikerroksisen alkion muodostumisvaihe ja sitten kolmikerroksisen alkion muodostumisvaihe. Tässä vaiheessa alkiota kutsutaan gastrulaksi.
Menetelmät kaksikerroksisen alkion muodostamiseksi:
- Intussusseptio (invaginaatio).
– Delaminaatio (kerrostuminen).
– Maahanmuutto (asutus).
- Epiboly (likaantuminen).
Intussusseptio tai invaginaatio. Tämä menetelmä on tyypillinen lansetille. Tietyllä alueella blastulasolut tunkeutuvat blastokoeliin, mikä johtaa kaksikerroksisen alkion muodostumiseen. Solujen ulompaa kerrosta kutsutaan ektodermiksi, sisäkerroksi endodermiksi. Endodermi rajoittaa primaarisen suolen tai gastrocoelin onteloa. Tämän ontelon sisäänkäyntiä kutsutaan ensisijaiseksi suuksi tai blastoporeksi. Blastoporea ympäröivät huulet.
Delaminaatio tai kerrostuminen. Tämä menetelmä on tyypillinen suolistoeläimille, joissa blastula näyttää morulalta ja blastokoelia ei käytännössä ilmene.
maahanmuutto tai asutus. Jotkut blastulasolut viedään blastokoeliin, minkä jälkeen nämä solut jakautuvat intensiivisesti. Tämän seurauksena näiden solujen ansiosta muodostuu endodermia.
Epiboly (likaantuminen). Mikromeerit jaetaan ja kerrostetaan ikään kuin makromeerien päälle. Mikromeerit muodostavat ektodermin, kun taas makromeerit muodostavat endodermin. Puhtaassa muodossaan näitä menetelmiä ei käytännössä löydy, yleensä ne yhdistetään. Sammakkoeläimet yhdistävät invagination ja epibolian. Linnut ja nisäkkäät yhdistävät delaminaatiota ja maahanmuuttoa.
Alkaen lattamatoista, kolmas itukerros, mesodermi, ilmestyy evoluutioon.
Mesodermin muodostusmenetelmät:
Teloblastinen menetelmä on tyypillinen protostomille. Blastoporen huulten alueella erittyy 2 solua, jotka jakautuvat ja muodostavat mesodermin.
Enterocoel-menetelmä on ominaista deuterostomille (chordateille). 2 taskujen muodossa olevaa soluosaa on symmetrisesti erotettu endodermista. Nämä ovat mesodermaalisia taskuja. Mesodermaalisten taskujen solut jakautuvat ja synnyttävät mesodermin. Mesodermi on itukerros.
8. Histo- ja organogeneesin yleiset ominaisuudet (kudosten ja elinten muodostuminen).
Histon ja organogeneesin vaihe(kudosten ja elinten muodostumisvaihe). Se on perinteisesti jaettu kahteen jaksoon.
1 jakso. Koulutusjakso aksiaaliset elimet alkiossa tämä on hermoputken ja jänteen muodostumista. Siksi tätä ajanjaksoa kutsutaan hermojaksoksi, ja tässä vaiheessa olevaa alkiota kutsutaan neurulaksi.
2 jaksoa. Sille on ominaista muiden kudosten ja elinten muodostuminen. Alkion selkäpuolella (dorsaali) sen koko pituudelta erottuu solujen osa ektodermista, mikä synnyttää hermolevyn. Sitten hermolevyn reunat kohoavat, paksuuntuvat ja muodostuu hermoura, joka vajoaa vähitellen ektodermin alle. Sitten hermouran reunat sulkeutuvat muodostaen hermoputken, jonka sisällä on onkalo, onkaloa kutsutaan neurocoeliksi. Selkärankaisilla hermoputken etuosa laajenee ja synnyttää aivot ja loput selkäytimen. Samanaikaisesti hermoputken alle asetetaan notokordi, joka muodostuu endodermista ja viereisestä mesodermista. Aluksi mesodermi on homogeeninen solumassa, mutta kehittyessään sen segmentoituminen tapahtuu. Muodostetaan rakenteita, joita kutsutaan somiiteiksi. Myöhemmin ne synnyttävät tuki- ja liikuntaelimistön.
Itukerrosten johdannaiset:
Ectoderm - hammaskiille, hermosto ja aistielimet, ihon orvaskesi ja sen lisäosat, etu- ja takasuolen epiteeli.
Endodermi - keskisuolen epiteeli, ruuansulatusrauhaset ja hengityselimet.
Mesoderma - tuki- ja liikuntaelimistö, virtsaelimet, verenkierto- ja imukudosjärjestelmä, kaikki sidekudos.
9. Selkärankaisten alkioiden väliaikaisten elinten ominaisuudet. Korkeampien nisäkkäiden väliaikaiset elimet.
Väliaikaiset elimet toimivat alkiossa, ja ne puuttuvat aikuisen tilassa. Näitä ovat keltuainen pussi ja niin sanotut alkiokalvot - amnion, korioni ja allantois.
Keltuainen pussi. Keltuaispussi suorittaa useita tärkeitä toimintoja: ravitsemus, hengitys, erittyminen, hematopoieesi. Mutta koska munassa on vähän keltuaista, sillä ei ole merkittävää roolia alkion ravitsemuksessa.
Amnion. Lapsivesikalvo osallistuu amnionin muodostumiseen, mikä rajoittaa sikiötä kaikilta puolilta nyt ympäröivää lapsivedellä täytettyä onkaloa. Tästä johtuen alkio kehittyy vesiympäristössä, mikä suojaa sitä mekaanisilta traumaattisilta vaikutuksilta ja kiinnittymiseltä kuoriin.
Allantois muodostuu takasuolen uloskasvuna. Allantoiksen päätehtävä on, että se on erityselin. Se kerää hajoamistuotteita, jotka muodostuvat aineenvaihdunnan aikana alkion kehossa.
Embryogeneesi on monimutkainen kokonaisvaltainen prosessi, joka liittyy tiettyihin ilmiöihin ja mekanismeihin. Monia näistä ilmiöistä ei täysin ymmärretä, vaikka joistakin niistä on saatu tiettyjä tietoja.
1. Molekyyligeneettiset muutokset varhaisessa kehityksessä.
2. Solujen lisääntyminen (solun jakautuminen).
3. Solujen erilaistuminen.
4. Muodostuminen tai morfogeneesi.
1. Molekyyligeneettiset muutokset varhaisessa kehityksessä (tsygootin ja pilkkoutumisen aika), sytoplasmisten tekijöiden rooli munassa.
Varhainen kehitys sisältää tsygootti- ja pilkkoutumisvaiheet.
Näitä vaiheita tutkiessaan tutkijat yrittivät vastata kysymyksiin:
Ensinnäkin, kun alkion omat geenit alkavat toimia.
Toiseksi, onko mRNA- ja proteiinimolekyyleissä laadullisia ja kvantitatiivisia eroja eri osat alkio varhaisessa kehitysvaiheessa.
Tsygootissa geeniaktiivisuus on alhainen, koska DNA liittyy vahvasti histoniproteiineihin. Ensimmäiset tsygootissa syntetisoituvat proteiinit ovat äidin alkuperää, koska ribosomit ja mRNA-molekyylit ovat kertyneet munaan etukäteen. On todettu, että nisäkkäissä alkion omat geenit alkavat toimia 2-4 blastomeerin vaiheessa. Sammakkoeläimissä se on blastula-vaiheessa. Ensimmäisenä toimivat proliferaatiosta ja yleisestä aineenvaihdunnasta vastaavat geenit, myöhemmin solujen ja kudosten erilaistumisesta vastaavat geenit. Esimerkiksi kun tuma poistetaan tsygootista, tapahtuu pilkkoutuminen ja alkio saavuttaa kehityksessään lähes blastula-vaiheen, jonka jälkeen jatkokehitys pysähtyy.
On todettu, että mRNA- ja proteiinimolekyyleissä ei ole laadullisia eroja alkion eri osissa kehityksen alkuvaiheessa. On vain määrällisiä eroja.
Tärkeä rooli murskaamisessa on sytoplasman jakautumisella - sytotomia. Sillä on erityinen morfogeneettinen merkitys, koska se määrittää murskaustyypin. Halkeamisuot kulkevat munasolun yksittäisten osien välisiä rajoja pitkin, mikä heijastaa ooplasmisen segregaation ilmiötä. Siksi eri blastomeerien sytoplasman kemiallinen koostumus eroaa.
2. Solujen lisääntyminen, kasvu.
Solujen lisääntyminen tai jakautuminen tapahtuu koko alkion synnyn ajan. Tähän liittyy kudosten ja elinten kasvu. Alkion kasvu kokonaisuudessaan.
3. Erilaistuminen, erilaistumisen molekyyligeneettiset mekanismit.
Solujen erilaistuminen on joukko prosesseja, joiden seurauksena yhteistä alkuperää olevat solut hankkivat pysyviä morfologisia, fysiologisia, biokemiallisia eroja, jotka johtavat solujen erikoistumiseen. Solujen spesifisyyden määräävät niissä syntetisoituvat proteiinit, ja vastaavat geenit ovat vastuussa proteiineista. Siksi voimme päätellä, että jotkut geenit toimivat joissakin soluissa ja toiset toisissa. Tämä on geenien erilaisen aktiivisuuden hypoteesin ydin.
Varhaisessa vaiheessa solujen erilaistuminen liittyy sytoplasmisten aineiden vaikutukseen vastaavien geenien työhön - tämä on geenien työn säätelyn epigeneettinen taso. Munassa tapahtuu munasolujen erotteluilmiö, jonka seurauksena munan sytoplasman eri osat sisältävät erilaisia aineita. Murskaamisen aikana ilmaantuu blastomeerejä, niissä olevien geenien joukko on sama ja sytoplasman koostumus on erilainen. Myöhemmin nämä sytoplasman aineet johtavat ilmeisesti geenien erilaiseen aktiivisuuteen.
Solujen erilaistumisen karakterisoinnissa käytetään kahta käsitettä - määrittely ja kompetenssi.
päättäväisyys tarkoittaa, että solujen erilaistuminen on geneettisesti ennalta määrättyä ja peruuttamatonta.
Erilaistumisprosessissa alkion anlagien solumateriaali muuttuu tietyksi aikuisen organismin elementiksi. Tarkastellaanpa erottelua käyttämällä esimerkkiä mesodermaalisesta somiitista, joka on jaettu dermatomiin, sklerotomiin ja myotomiin. Dermiksen ihosolut, toinen - rustosolut, kolmas - poikkijuovaiset lihaskuidut. Näin ollen yksittäisten alkioanlagien kehityksen lopputulos on ennalta määrätty tai määritetty.
Pätevyys Tämä on solujen kykyä erilaistua eri suuntiin ympäristötekijöiden vaikutuksesta. Esimerkiksi notochord ja viereinen mesodermi vaikuttavat ektodermiin, mikä johtaa hermoputken muodostumiseen ektodermista. Jos tällaista vaikutusta ei ole, ektodermi synnyttää ihon orvaskeden.
4. Morfogeneesi (muotoilu), sen pääprosessit:
Muodostuminen tai morfogeneesi. Morfogeneesi on joukko prosesseja, joiden seurauksena alkio saa ominaisen ulkoisen ja sisäinen rakenne. Morfogeneesi puolestaan liittyy:
a) solujen morfogeneettinen liike
Alkion synnyn aikana yksittäiset solut tai soluryhmät liikkuvat. Solut liikkuvat muiden solujen pinnalla (ameboidiliikkeen mekanismin vuoksi), joissa on erityisiä molekyylejä, jotka osoittavat liikkeen suunnan. Jotkut solutyypit liikkuvat kemiallista pitoisuusgradienttia pitkin (kemotaksis), mutta tämä mekanismi on paljon harvinaisempi.
Solujen migraation rikkominen alkion synnyn aikana johtaa elinten alikehittymiseen tai muutokseen sen normaalissa sijainnissa. Molemmat ovat synnynnäisiä epämuodostumia. Esimerkiksi solujen - neuroblastien - vaeltamisen vastaisesti valkoiseen aineeseen ilmestyy harmaa-aineen saarekkeita, ja samalla solut menettävät kykynsä erilaistua.
Siten solujen migraatio on toisaalta geneettisen hallinnan ja toisaalta ympäröivien solujen ja kudosten vaikutuksen alaisena.
b) alkion induktio
Tämä on yhden kudoksen (induktorin) vaikutus toiseen kudokseen, minkä seurauksena indusoidun kudoksen kehitys muuttuu laadullisesti uudeksi. Ensimmäinen ja merkittävin induktio on notokoordin ja mesodermin vaikutus ektodermiin, mikä johtaa hermoputken muodostumiseen. Ilman hermoputkea koko ektodermi muuttuu orvaskedeksi. Tämä on primaarinen alkion induktio, ensimmäinen vaihe peräkkäisten (toissijaisten, tertiääristen) induktioprosessien ketjussa jatkokehityksessä.
On todettu, että on olemassa "spesifisiä induktoreita", ts. aineet, joilla on indusoiva vaikutus vähäisinä pitoisuuksina ja jotka eroavat toimintansa lopputuloksesta. Siten nisäkkäiden maksan uute indusoi pääasiassa aivorakenteiden kehittymistä ja luuytimen uute - mesodermaalisia.
Alkion alkion kykyä havaita induktioärsyke kutsutaan kompetenssiksi.
c) solujen väliset vuorovaikutukset
Tämä on kosketuksissa tai etäisyydellä olevien solujen tai kerrosten vuorovaikutusta. Vuorovaikutus etänä tapahtuu biologisesti aktiivisten aineiden (BAS) kanssa.
Nämä voivat olla proteiineja, hormoneja jne. Alkion kehityksen alkuvaiheessa nämä ovat äidin hormoneja, koska alkiolla ei ole omia umpirauhasia. Hormonit eivät aiheuta uutta erilaistumista, mutta vahvistavat sitä.
Solujen välisistä vuorovaikutuksista johtuen tapahtuu sellaisia ilmiöitä kuin morfogeneettinen soluliike, alkion induktio ja soluadheesio.
d) tarttuvuus- solujen kyky tarttua yhteen. Kokeessa ektodermin, mesodermin ja endodermin solut erotettiin ja sekoitettiin keskenään. Sitten ne taas kokoontuvat erillisiin ryhmiin, joista jokainen on homogeenisten solujen soluaggregaatti. Muodostuu jälleen kolme alkiokerrosta, jotka sijaitsevat normaalisti suhteessa toisiinsa.
Erityiset proteiinimolekyylit osallistuvat kiinnittymisprosessiin. Niitä kutsutaan soluadheesiomolekyyleiksi (MAC), ja niitä on noin 100 tyyppiä.
Toinen hypoteesi väittää, että samanlaisten solujen väliset kontaktit ovat vahvempia kuin vieraiden solujen välillä.
Tietyn itukerroksen solujen selektiivinen tarttuminen toisiinsa on normaalin kehityksen välttämätön edellytys.
e) solukuolema on välttämätön prosessi, koska yksittäisten rakenteiden (kanavien, kanavien, reikien jne.) muodostuminen edellyttää osan soluista tuhoutumista.
On olemassa kaksi pohjimmiltaan erilaista solukuolemaa: apoptoosi(käännetty kreikaksi "putoaminen pois") ja nekroosi.
Apoptoosi on fysiologinen, geneettisesti ennalta määrätty solukuolema. Yhdessä muiden morfogeneesimekanismien kanssa se edistää tietylle biologiselle lajille ominaisten morfofunktionaalisen organisaationsa ominaisuuksien saavuttamista. Siksi apoptoosi on luonnollinen, evoluutionaalisesti määrätty ja geneettisesti kontrolloitu morfogeneesin mekanismi.
Nekroosi - ei-fysiologinen solukuolema, joka johtuu haitallisten tekijöiden vaikutuksesta (mekaaninen, kemiallinen, fyysinen jne.). Nekroosiin liittyy yleensä tulehdus ja se on patologinen prosessi.
5. Integrointi kehitykseen, ontogenian eheys. Hormonien rooli kehitysprosessien koordinoinnissa.
Tällä hetkellä tunnetaan useita aineita, jotka saavat solut jakautumaan, esimerkiksi fytohemagglutiniini, jotkut hormonit sekä kudosvaurion aikana vapautuva ainekompleksi. Myös solujen jakautumisen kudosspesifisiä estäjiä on löydetty - keylonit. Niiden toiminta on tukahduttaa tai hidastaa solujen jakautumisnopeutta niitä tuottavissa kudoksissa. Esimerkiksi epidermaaliset kalonit vaikuttavat vain orvasketeen. Koska kalonit ovat kudosspesifisiä, niiltä puuttuu lajispesifisyys. Näin ollen turskan epidermaalinen kalon vaikuttaa myös nisäkkään orvasketeen.
Hormonit ovat tiettyjen solujen tuottamia orgaanisia yhdisteitä, jotka on suunniteltu säätelemään kehon toimintoja, niiden säätelyä ja koordinaatiota.
Hormonien fysiologinen vaikutus on suunnattu:
1) tarjoaa humoraalista, ts. suoritetaan veren kautta, biologisten prosessien säätely;
2) sisäisen ympäristön eheyden ja pysyvyyden ylläpitäminen, kehon solukomponenttien harmoninen vuorovaikutus;
3) kasvu-, kypsymis- ja lisääntymisprosessien säätely. Hormonit säätelevät kaikkien kehon solujen toimintaa. Ne vaikuttavat ajattelun terävyyteen ja fyysiseen liikkuvuuteen, ruumiinrakenteeseen ja kasvuun, määräävät seksuaalisen dimorfismin ja käyttäytymisen merkkien kehittymistä.
6. Perinnöllisyyden ja ympäristön rooli alkion kehityksessä. Kriittiset kehitysjaksot. teratogeeniset tekijät. Anomaliat ja epämuodostumat.
Missä tahansa ontogeneesin vaiheessa organismi on olemassa yhtenäisyydessä ympäristöön. Embryogeneesi ei ole tässä suhteessa poikkeus. Lajin elämään tarvittavien olosuhteiden kirjo voi olla laaja. Minkä tahansa lajin organismeille on kuitenkin olemassa minimi, optimi ja maksimi tarvittavat ehdot kehitystä. Alkion kehitykseen vaikuttavat luonnossa esiintyvien tekijöiden (lämpötila, kosteus, ilmanpaine, säteily, ympäristön kaasukoostumus) vaihtelut.
Joten lämpötilasta riippuen kehitysprosessit hidastuvat tai tehostuvat. Esimerkiksi samasta kytkimestä peräisin olevat sammakonmunat kehittyvät nopeammin korkeammissa lämpötiloissa.
Sukulamadossa kehitys pysähtyy, kun hapen pääsy alkioon loppuu.
Yleissääntö on, että spektrin siniviolettiosasta tulevan valon vaikutuksesta monien eläinlajien alkionkehitys kiihtyy ja punaisesta se hidastuu.
Kohdunsisäisen kehityksen aikana ympäristötekijät ovat erittäin tärkeitä. Jos nämä tekijät johtavat poikkeavuuksien tai kehityshäiriöiden muodostumiseen, niitä kutsutaan teratogeenisiksi. Teratogeeniset tekijät voivat olla fysikaalisia (korkea lämpötila, ionisoiva säteily, röntgen jne.), kemialliset (lääkkeet, raskasmetallien suolat jne.) ja biologiset (virukset, bakteerit). Teratogeeniset tekijät johtavat poikkeavuuksien kehittymiseen tietyillä alkion kehitysjaksoilla, joita kutsutaan kriittisiksi. Nämä sisältävät:
Sukusolujen muodostumisaika (gametogeneesi),
hedelmöitysvaihe,
tsygoottivaihe,
Alkion istutus kohdun seinämään
istukka,
Histogeneesin ja organogeneesin ajanjakso,
Epämuodostumat.
Aplasia - elimen tai sen osan puuttuminen
Hypoplasia - elimen alikehittyminen
Hypotrofia - kehon tai elimen painon lasku
Hypertrofia - elinten massan suhteeton kasvu
Gigantismi - kehon pituuden kasvu
Heterotopia on soluryhmän tai elimen epätyypillinen sijainti kehossa.
Heteroplasia - kudosten erilaistumisen rikkominen
Ahtauma - kanavan tai aukon kapeneminen
Atresia - kanavan tai aukon puuttuminen
Pysyvyys - alkiorakenteiden säilyttäminen
Syystä riippuen synnynnäiset epämuodostumat jaetaan:
perinnöllinen johtuu geenien tai kromosomien muutoksesta vanhempien sukusoluissa, minkä seurauksena tsygootissa on alusta alkaen geeni-, kromosomi- tai genomimutaatio.
eksogeeninen jotka syntyvät teratogeenisten tekijöiden vaikutuksesta: lääkkeet (talidomidi), elintarvikelisäaineet, virukset, teolliset myrkyt jne. Nämä ovat kaikki ympäristötekijöitä, jotka vaikuttavat alkion synnyn aikana häiritsevät kudosten ja elinten kehitystä.
Monitekijäinen vikoja, jotka kehittyvät sekä eksogeenisten että geneettisten tekijöiden vaikutuksesta.
1. Synnytyksen jälkeinen ontogeneesi, sen periodisointi.
Postembryonaalinen kehitys (henkilölle postnataalinen) alkaa syntymähetkestä ja päättyy luonnolliseen kuolemaan tai kuolemaan.
Postembryonaalinen kehitys sisältää useita ajanjaksoja:
1. Lisääntymistä edeltävä (nuori).
2. Lisääntymisikä (kypsyysaika).
3. Lisääntymisen jälkeinen (vanhuus).
2. Lisääntymistä edeltävä aika, sen ominaisuudet. Kehon kasvu tärkeänä ominaisuus ennen lisääntymistä.
Lisääntymistä edeltävä aika alkaa heti syntymän jälkeen. Tällä hetkellä morfogeneesiprosessit päättyvät, ne järjestelmät, jotka eivät toimineet alkiosyntyessä (hengitys, eritys ja monet muut), alkavat toimia.
Lisääntymistä edeltävän ajanjakson tärkeä ominaisuus on organismin kasvu. Tässä tapauksessa koko kehon koko kasvaa, sen pituussuuntaiset mitat kasvavat; kudosten ja elinten koon kasvu.
3. Organismin ja sen yksittäisten osien kasvun luonne.
Organismin kasvun taustalla on kolme pääprosessia:
1. solujen määrän kasvu.
2. solukoon kasvu (hypertrofia).
3. solujen välisen aineen kertyminen.
Kasvuvaihtoehtoja on kaksi: rajoitettu Ja rajoittamaton. Rajoittamaton kasvu jatkuu koko ontogeneesin ajan kuolemaan asti.
Kasvutyyppejä on useita:
Ylimääräinen- kasvu, joka tapahtuu kasvattamalla solujen kokoa.
proliferatiivinen- solujen lisääntymisen kautta tapahtuva kasvu: kertova Ja lisääntyvä.
Kertova kasvulle on ominaista se, että molemmat solut, jotka ovat syntyneet emosolun jakautumisesta, alkavat jälleen jakautua. Multiplikatiivinen kasvu on erittäin tehokasta ja siksi sitä ei esiinny juuri koskaan puhtaassa muodossaan tai se päättyy hyvin nopeasti (esimerkiksi alkiovaiheessa).
lisääntyvä kasvu johtuu siitä, että jokaisen seuraavan jakautumisen jälkeen vain yksi tytärsoluista jakautuu uudelleen, kun taas toinen lopettaa jakautumisen. Tässä tapauksessa solujen lukumäärä kasvaa lineaarisesti. Tällainen kasvu on tyypillistä elimille, joissa tapahtuu solukoostumuksen uusiutumista.
On syytä huomauttaa, että vartalon pitkittäisten mittojen kasvu, joka johtuu pääasiassa pitkien putkiluiden kasvusta, on erityisen tärkeä kasvun karakterisoinnissa. SISÄÄN putkimaiset luut diafyysin ja epifyysin rajalla on eristetty kasvuvyöhyke. Tässä on rustosoluja, joiden jakautumisen aikana luu kasvaa pituudeltaan.
Jokaisen luun lopullinen luutuminen tapahtuu tiettynä aikana. Miehillä kasvu päättyy yleensä 18-20 vuoden iässä, naisilla - 16-18 vuoden iässä. Tällä hetkellä viimeiset kasvuvyöhykkeet katoavat. Silloin luiden kasvu pysähtyy.
On huomautettava, että 30-vuotiaaksi asti ihminen voi kasvaa 3 cm nikamien koon kasvun vuoksi.
Henkilön lineaaristen mittojen kasvua kuvaa S-muotoinen käyrä. Välittömästi syntymän jälkeen kehon kasvu lisääntyy, sitten se vähenee ja kiihtyy jyrkästi 13-14-15-vuotiaana. Tämä on niin sanottu murrosiän kasvupyrähdys (murrosiän aikana). Lisäksi kasvuvauhti hidastuu jonkin verran ja 30-40-45-vuotiaana ihmisen kasvu pysyy vakiona. Tämän järjestelmän mukaan luut, lihakset ja monet sisäelimet(maksa, munuainen, perna).
Ikääntymisen myötä kasvu hidastuu hieman.
Joillakin elimillä on täysin erilainen luonne:
Näitä elimiä ovat aivot ja selkäydin, lymfaattiset elimet, lisääntymiselimet.
Vastasyntyneen aivojen paino on 25% lopullisesta aivojen painosta (aikuisena), 5-vuotiaana - 90%, 10-vuotiaana - 95%.
Kateenkorvan kasvu (kateenkorva) on immuunijärjestelmän keskuselin. Kateenkorvan suhteellinen paino (painoon) saavuttaa maksiminsa 12 vuoden iässä. Absoluuttinen paino saavuttaa maksiminsa 30 vuoden iässä, ja sitten kateenkorvan paino laskee jyrkästi.
4. Geneettinen kasvun hallinta. Hermoston ja endokriinisen järjestelmän rooli kasvuprosessien säätelyssä.
Pituus viittaa geneettisiin ominaisuuksiin, jotka ovat periytyviä, kuten hiusten ja ihon väri, silmien muoto jne. Siksi pitkillä vanhemmilla on yleensä pitkät lapset ja päinvastoin. Kasvu on polygeeninen ominaisuus; useat geenit ovat vastuussa sen ilmenemisestä fenotyypissä. Geenit hallitsevat kasvua asianmukaisten hormonien avulla. Tärkein hormoni on aivolisäkkeen tuottama kasvuhormoni eli somatotropiini.
Somatotropiini stimuloi uusien rustosolujen muodostumista ja osittain niiden luutumista, edistää proteiinisynteesiä solujen rakenteet ah ja uusien kapillaarien muodostuminen. Suuri määrä tätä hormonia tuotetaan yöllä. Lapsen omaa somatotropiinia tuotetaan 3-4-vuotiaana.
Kasvuun vaikuttavat myös kilpirauhashormonit ja sukupuolihormonit.
5. Biologisen ja sosiaalisen vuorovaikutus lapsuuden ja nuoruuden aikana.
Perinnöllisyyden merkitys kasvulle on suuri, mutta tämä ei ole ainoa tekijä. Perinnöllisyyttä tulee pitää likimääräisenä ohjelmana, jonka mukaan henkilön pituus voi olla esimerkiksi välillä 160-180 cm. Se, mikä se todella on, riippuu pitkälti ulkoisista olosuhteista, jotka voivat hidastaa perinnöllistä ohjelmaa tai edistää sen toteuttamista. Ihmisen kasvuun vaikuttavat ympäristöolosuhteet: ravitsemus, fyysinen aktiivisuus, tupakoinnin psykologiset vaikutukset, alkoholi.
Eli väestö kasvaa (tämä on kiihtymistä).
Yksi sen todennäköisistä syistä on elinolojen (ravitsemuksen) paraneminen. On havaittu, että sotien ja luonnonkatastrofien aikana lasten kasvu hidastuu. Kasvuun vaikuttavat hieman ilmasto ja maantieteellinen ympäristö.
6. Perustuslaillisten tyyppien, vartalotyyppien muodostuminen.
Perustuslaillisten ihmistyyppien muodostuminen liittyy ihmisen kasvuun. Tämä tulisi ymmärtää kehon ulkoisten muotojen ominaisuuksiksi, kehon toimintojen ominaisuuksiksi, käyttäytymisen ominaisuuksiksi Tämä henkilö. Kehon rakenteesta riippuen kehon ulkoisista muodoista riippuen erotetaan tietyt fysiikkatyypit. Tällä hetkellä luokituksia on useita. Yksi niistä on M.V. Tšernorutski. Tämän mukaan
Jokainen organismi, riippumatta siitä onko se yksisoluinen vai monisoluinen ja mihin elämän valtakuntaan se kuuluu, kulkee sen läpi yksilöllistä kehitystä, tai ontogeneesi(kreikasta. päälle- olento ja synty- syntymä). Biologian opiskelun ala ontogeneesi, kutsutaan kehitysbiologia.
Ontogeneesi - tämä on organismin elinaika tsygootista (primäärisolusta) kuolemaan.
Monisoluisissa organismeissa ontogeneesi alkaa yleensä tsygootin muodostumisella ja päättyy kuolemaan.
Samaan aikaan keho ei vain kasva, kasvaa kokoaan, vaan myös käy läpi useita erilaisia elämänvaiheita, joista jokaisella on erityinen rakenne, se toimii eri tavalla ja joissakin tapauksissa radikaalisti erilainen elämäntapa. .
Yksisoluisissa organismeissa ontogeneesin alkamisajankohtana pidetään eroamishetkeä emo- tai sisarsolusta. Se jatkuu seuraavaan jakautumiseen tai kuolemaan asti. Samanaikaisesti yksisoluisten organismien ulkoinen ontogeneettisyys ilmenee yleensä vain lievänä solukoon kasvuna, vaikka itse asiassa tämä kätkee täysin erilaisia elämän jaksoja. .
Jokaisella lajilla on oma ontogeneesiohjelmansa. Ja tämä ei ole vain joukko ja sarja kehitysvaiheita, jotka hän käy läpi, vaan myös kunkin niistä kesto. Samaan aikaan jokaisella yksilöllä on yksilöllisiä ontogeneettisiä piirteitä, jotka eivät kuitenkaan ylitä lajia. , ja ne puolestaan noudattavat ontogeneesin lakeja heimo-, perhe-, irrallisuus- ja jopa luokkatasolla .
ontogeny-ohjelma ei ole muuta kuin geeneihin tallennetun perinnöllisen tiedon toteutumista. Siksi ontogenian spesifisyyden yksittäisten yksilöiden tasolla määrittävät yksittäiset geeniyhdistelmät ja lajien, sukujen, perheiden tasolla - erityiset geenit, jotka ovat ominaisia vain kullekin järjestelmälliselle organismiryhmälle.
Mekanismi perinnöllisen tiedon toteuttamiseksi on ennen kaikkea sisällä ero(englannista. erota- vaihtelevat) geenien aktiivisuus. Tämä tarkoittaa, että eri kehitysjaksoissa ja eri kudoksissa monisoluinen organismi geenit ovat aktiivisia, joiden nukleotidisekvensseissä syntetisoidaan tiettyjä mRNA:ita. Tämän seurauksena syntetisoidaan spesifisiä rakenneproteiineja ja entsyymejä, jotka lopulta määräävät solujen toiminnan ja käyttäytymisen ominaisuudet kehon tietyissä kehitysvaiheissa, niiden aineenvaihdunnan ominaisuudet.
Eläimillä ja kukkakasveilla ontogeneettisyys on jaettu kahteen ajanjaksoon. Alkion kehitys, tai embryogeneesi(kreikasta. alkio- sikiö ja synty) kestää tsygootin muodostumisesta syntymään tai munasta poistumiseen, ja sikiön jälkeinen(kreikasta. nopeasti- jälkeen ja alkion) kehitys jatkuu syntymästä tai munasta poistumisesta lähtien ja päättyy organismin kuolemaan.
Alkion synnyn kulku. Alkion synnyn yleinen kaavio sisältää seuraavat vaiheet: materiaalia sivustolta
- tsygootin peräkkäiset jakautumiset, jotka huipentuvat monisoluisen alkion muodostumiseen, joka koostuu sadoista ja jopa tuhansista identtisistä soluista;
- erottelu (englanniksi. erota- erottaa) solut, mikä johtaa kudosten muodostumiseen;
- elinten muniminen ja alkion kasvu.
Aikana ontogeniteetti eliö käy läpi useita vaiheita - tiloja, joissa se eroaa rakenteeltaan, toiminnaltaan ja elämäntavolta. Ontogeneesissä on kaksi vaihetta: embryogeneesi - alkion kehitys ja postembryonaalinen kehitys- organismin elinaika syntymästä (poistuminen munasta) kuolemaan.
Tällä sivulla materiaalia aiheista:
Ontogenian yhteenveto
Yksilöllisen kehityksen ontogeny abstrakti
Ontogeneesin käsite lyhyesti
-
Luku 7. ONTOGENEESIIN PERIODIOINTI
Luku 7. ONTOGENEESIIN PERIODIOINTI
7.1. ontogenian vaiheet, jaksot ja vaiheet
Yksilöllinen kehitys on kokonaisvaltainen jatkuva prosessi, jossa yksittäiset tapahtumat ovat yhteydessä toisiinsa tilassa ja ajassa. Ontogenian periodisaatiosta on olemassa useita muunnelmia, joista jokainen sopii parhaiten tiettyjen tieteellisten tai käytännön ongelmien ratkaisemiseen.
KANSSA yleinen biologinen näkökulmat suuri tapahtuma ontogenia - sukupuolinen lisääntyminen. Jos korreloimme eri ontogeneesin ajanjaksoja yksilön kykyyn suorittaa lisääntymistoiminto, se voidaan jakaa kolmeen ajanjaksoon: lisääntymistä edeltävä, aktiivinen lisääntyminen ja lisääntymisen jälkeinen ajanjakso.
SISÄÄN lisääntymistä edeltävä ajanjakso yksilö on lisääntymiskyvytön. Sen pääsisältö on sukukypsän fenotyypin kehittäminen. Tänä aikana tapahtuvat selkeimmät rakenteelliset ja toiminnalliset muutokset, suurin osa perinnöllisistä tiedoista toteutuu, keho on erittäin herkkä kaikenlaisille vaikutuksille.
Epäaktiivinen lisääntymisaika yksilö suorittaa seksuaalisen lisääntymisen toiminnon, erottuu elinten ja järjestelmien vakaimmasta toiminnasta sekä suhteellisesta vastustuskyvystä vaikutuksille.
Lisääntymisen jälkeinen ajanjakso liittyy kehon ikääntymiseen ja sille on ominaista lisääntymiseen osallistumisen heikkeneminen tai täydellinen lopettaminen. Vähentynyt sopeutumiskyky ja vastustuskyky erilaisille vaikutuksille. Ihmisen ontogeneesin suhteen näille ajanjaksoille on lisäksi ominaista erityiset sosiaaliset tekijät (koulutus, työkyky, luovuus). Jokaisella ajanjaksolla on omat sairauden ominaispiirteensä.
Lisääntymistä edeltävä ajanjakso on jaettu neljään muuhun: alkio, toukka, metamorfoosi ja nuori.
alkio, tai itiö, piste Ontogeneesi alkaa hedelmöityksestä ja jatkuu siihen asti, kunnes alkio vapautuu munakalvoista. Tälle ajanjaksolle on tunnusomaista tsygootin muuttumisprosessien vakavuudella organismiksi, joka pystyy olemaan enemmän tai vähemmän itsenäisesti olemassa. Useimmissa selkärankaisissa se sisältää tsygootin vaiheet (vaiheet), pilkkoutumisen, gastrulaation sekä histo- ja organogeneesin. Sen kesto vaihtelee. Istukan nisäkkäillä se on erityisen lyhentynyt. Yksittäinen munankuori liukenee ennen blastokystan istuttamista kohdun limakalvoon. Tähän mennessä alkiolla on vain aikaa käydä läpi tsygootin ja murskauksen vaiheet. Kaikki muut prosessit etenevät äidin kehon suojeluksessa ja osallistuessa siihen. Näiden piirteiden evolutiivista merkitystä tarkastellaan luvussa 13.2.
Pilkkomista edeltävät gametogeneesi- ja hedelmöitysprosessit, jotka liittyvät suoraan yksilön kehitykseen eivätkä välttämättä edes johda siihen, mutta jotka määrittävät pitkälti alkion jatkokehityksen, jos hedelmöitys tapahtuu. Näitä prosesseja kutsutaan progeneesiksi, joka edeltää varsinaista ontogeneesiä. Sytologisesti gametogeneesi- ja hedelmöitysprosessit edustavat välilinkkiä, joka yhdistää vanhempien ontogeneesin heidän jälkeläistensä ontogeneesiin.
Toukkakausi tyypillisessä muunnelmassa se havaitaan niiden selkärankaisten kehityksessä, joiden alkiot nousevat ulos munakalvoista ja alkavat elää itsenäistä elämäntapaa saavuttamatta organisaation lopullisia (kypsiä) piirteitä. Joten sitä löytyy joistakin alempien selkärankaisten edustajista - nahkiaisista, useimmista luisista kaloista ja sammakkoeläimistä. Toukan tyypillisimpiä piirteitä ovat: sen organisaation alkion luonne, tilapäisten (väliaikaisten) elinten läsnäolo, useiden elinten toiminnan varhainen alkaminen, mikä mahdollistaa itsenäisen olemassaolon. Aktiivisen ruokinnan ansiosta toukka saa mahdollisuuden kehittyä loppuun, ja aktiivisen liikkeen ansiosta sillä on mahdollisuus valita kehitykselle optimaaliset ympäristöolosuhteet ja välttää näin kilpailua omien aikuisten sukulaistensa kanssa. Selkärankaisilla toukkakauden kesto on paljon pidempi kuin alkiojakso.
Metamorfoosi koostuu toukan muuttumisesta nuoreksi. Metamorfoosiprosessissa tapahtuu sellaisia tärkeitä morfogeneettisiä muutoksia kuin osittainen tuhoutuminen, uudelleenjärjestely ja uusien elinten muodostuminen. Muutosaste on mitä suurempi, sitä enemmän
erot toukan ja aikuisen organismin elinympäristön välillä, mitä havainnollistaa hyvin esimerkki hännänttömien sammakkoeläinten kehityksestä (vertaa: nuijapolka ja sammakko).
Nuorten ajanjakso alkaa muodonmuutoksen päättymishetkestä (istukan nisäkkäillä ja ihmisillä - syntymästä) ja päättyy murrosikään ja lisääntymisen alkamiseen. Nuoruusajan piirteet ilmenevät nuoren organismin ravinnon omaperäisyydestä, sen käyttäytymisestä ja riippuvuuden asteesta vanhemmista. Morfologisesta näkökulmasta tälle ajanjaksolle on ominaista voimakas kasvu, lopullisten suhteiden muodostuminen välillä erilaisia osia kehon, luuston, ihon, hampaiden vaihdon, sukurauhasten ja hormonaalisen säätelyn kehittymisen loppuun saattaminen. Nuoruuden kesto selkärankaisilla vaihtelee vähintään 13–18 vuorokaudesta hiirmyyrillä enintään 18–20 vuoteen belugassa, krokotiilissa, albatrossissa ja norsussa. Monilla selkärankaisten edustajilla, erityisesti ihmisillä, murrosiän saavuttaminen ja lisääntymisen alkaminen voivat erota toisistaan merkittävällä aikavälillä.
Sovellus ekologinen ja embryologinen lähestymistapa mahdollistaa ontogenian jakamisen tapahtuviin vaiheisiin ennen syntymää, sen aikana Ja syntymän jälkeen yksilöitä. Itse synnytys, ts. kehittyvän yksilön vapautuminen munankuoresta tai äidin kehosta erilaisia tyyppejä esiintyy eri kypsyysvaiheissa. Samaan aikaan kaikissa lajeissa ennen syntymää elimistö on suojattu munakalvojen tai äidin kehon toimesta, eikä se pysty itse ruokkimaan ja suorittamaan muita tärkeitä toimintoja. Varhaisten, morfofunktionaalisesti epäkypsien vaiheiden suojeleminen varmistaa lajin säilymisen. Syntymän jälkeen yksilö muodostaa yhteydet uuteen ympäristöön, alkaa ruokkia itse, liikkua ja suorittaa kaikkia muita toimintoja.
Yksityiskohtaisempi esitys ihmisen ontogeneesin periodisaatiosta, joka on käytännön arvoa synnytys- ja lastenlääkärin käytännössä katso kohta 7.6.1 ja taulukko. 7.3.
7.2. chordaattien munien morfofysiologiset ja evoluutiopiirteet
7.2.1. MUNIEN ALKUPERÄ
Chordate-munat koostuvat munasolusta, jota joskus kutsutaan myös munaksi, ja munakalvoista. Munasolut muodostuu naisen sukuelimiin
rauhanen - munasarja. He käyvät läpi pitkän kehityspolun, joka alkaa alkiossa ja jatkuu naarasyksilöiden ontogenian aktiivisessa lisääntymisjaksossa.
Ensisijaiset sukupuolisolut alkion alkuvaiheessa eristetään somaattisista soluista. Joten hännänttömissä sammakkoeläimissä gonosyytit eristetään blastula-vaiheessa, linnuissa ne määritetään primaarisen hypoblastin muodostumisen aikana, nisäkkäillä epiblastissa gastrula-vaiheessa, hännän sammakkoeläimissä tämä prosessi tapahtuu myöhemmin, gastrula-vaiheessa. tai jopa neurula (mesodermassa). Sitten nämä solut liikkuvat ja saavuttavat lopulta sukurauhasten alkeet. Nisäkkään gonosyytit löytävät itsensä ensin alkion ulkopuoliselle alueelle keltuaisen pussin suussa ja sitten siirtyvät sukurauhasten anlageen (kuva 7.1). Siirtymämekanismit esitetään kohdassa 6.5.3.
Sukurauhasiin joutuessaan primaariset sukusolut alkavat lisääntyä. Ne jakautuvat mitoosilla ja niitä kutsutaan ovo(oo)gonia. Useimmissa alemmissa selkärankaisissa ovo(oo)gonia säilyttää kykynsä jakautua koko lisääntymisjakson ajan, esimerkiksi kalat vapauttavat tuhansia munia yhdellä kutukerralla, sammakkoeläimet - satoja. Korkeammilla selkärankaisilla samaan aikaan kypsyvien munien määrä on harvoin 15, yleensä niitä on vähemmän, joskus yksi, mikä voi myös selittää ovo(oo)geneesin piirteet.
Naisten ihmisalkioissa ovo(oo)gonia lisääntyy intensiivisimmin kohdunsisäisen 2. ja 5. kuukauden välillä
Riisi. 7.1. Primaaristen itusolujen lokalisointi ihmisen alkiossa 16 somiitin vaiheessa (a) ja niiden siirtyminen sukurauhasten anlageen (b): 1 - etusuola; 2 - takasuoli; 3 - allantois; 4 - primaariset sukusolut; 5 - keltuainen pussi; 6 - sydän; 7 - kehittyvä sukupuolirauhanen (sukupuolielinten tuberkuloosi); 8 - primaarinen munuainen; 9 - kloakki
kehityskausi, jolloin niiden lukumäärä on noin 7 miljoonaa.. 7. kuukauteen mennessä monet ovo(oo)goniat kuolevat ja loput siirtyvät meioosin ensimmäisen jakautumisen profaasiin ja pysähtyvät diakineesin vaiheeseen. Murrosiän alkaessa yksi munasolu ovuloituu joka kuukausi ja saavuttaa meioosin toisen jakautumisen metafaasin. Joidenkin munien kohdalla tämä tapahtuu murrosiän aikana ja toisilla juuri ennen vaihdevuosia. Oosyytti suorittaa meioosin vain, jos hedelmöityminen tapahtuu.
7.2.2. MUNAN SYTOPLASMAN KEMIALLISEN KOOSTUMUKSEN ERITYISUUS JA MERKITYS
Kypsä muna on yleensä suurempi kuin ovo(oo)gonia ja muut somaattiset solut. Munasolun sytoplasmassa ovo(oo)geneesin aikana suuri määrä sen kypsymiseen ja varhaiseen alkiosyntymiseen tarvittavia aineita. Siten kasvukauden aikana hiiren munasolun tilavuus kasvaa noin 40-kertaiseksi ja sammakon munasolun tilavuus yli 400. Aineiden synteesinopeus munasoluissa lisääntyy merkittävästi useiden ominaisuuksien vuoksi. Kypsyvissä munissa tapahtuu yksittäisten geenien (katso osa 4), esimerkiksi rRNA-geenien monistumista (katso kohta 2.4.3.4). sammakkoeläimet (Xenopus laevis) noin 450 rRNA-geeniä löytyy diploidisesta solusta ja yli miljoona ovo(oo)syytistä. Lisäksi näiden solujen genomin transkriptioaktiivisuus lisääntyy. Siten transkriptionopeus kasvavissa hiiren munasoluissa on 10 kertaa suurempi kuin somaattisissa soluissa.
Varastoitujen aineiden toiminnallinen rooli on erilainen.
Ensinnäkin nämä ovat replikaatio-, transkriptio- ja translaatioprosesseille välttämättömiä komponentteja, kuten vastaavat entsyymit, ribosomit, mRNA, tRNA ja niiden esiasteet.
Toiseksi, se on joukko erityisiä säätelyaineita, jotka varmistavat kaikkien tallennettujen komponenttien koordinoidun toiminnan. Nämä aineet sisältävät ydinkalvon hajoamistekijän (meioosin ensimmäisen jaon prometafaasi alkaa ydinkalvon tuhoutumisesta); tekijä, joka aiheuttaa kromosomien kondensaation; tekijä, joka muuttaa siittiön ytimen esiytimeksi ja aktivoi DNA-synteesin siinä ennen katkaisua; sytostaattinen tekijä csf, vastuussa meioosin estämisestä metafaasin II vaiheessa (monissa selkärankaisissa tämän lohkon poistaminen tapahtuu vain hedelmöityksen seurauksena); sytoskeletaalinen mikrofilamenttijärjestelmä, joka osallistuu lohkon ylläpitoon
meioosi; sykliset nukleotidit (esim. cAMP), proteiinikinaasit ja muut aineet, joita tarvitaan signalointiin ympäröivistä ovo(oo)syyttisoluista ja solunulkoisesta ympäristöstä; niin kutsutun inositolifosfaattijärjestelmän komponentit, jotka varmistavat munasolun aktivoitumisen hedelmöityksen jälkeen; tekijä, joka on vastuussa sytotomiasta katkaisun aikana. Jotkut niistä, jotka ovat munasolussa jo hedelmöityshetkellä, alkavat toimia vasta gastrulaatiovaiheessa.
Kolmas, se on keltuainen, joka sisältää proteiineja, fosfolipidejä, neutraaleja rasvoja, hiilihydraatteja, mineraalisuoloja. Keltuainen on ravinto- ja energiavarasto, joka on välttämätön alkiokauden varmistamiseksi. Monet maksan tuottamat aineet pääsevät ovo(oo)goniaan kasvunsa aikana munasarjan follikulaaristen solujen kautta. Tämä vaatii naiselta suurta energiankulutusta.
Neljäs, Nämä ovat erityisiä aineita, joita kutsutaan morfogeneettiset tekijät, määrittämällä melko jäykästi niiden solujen (blastomeerien) kohtalon, joihin ne putoavat pilkkomisen aikana. Samanlaisia aineita ei löydy kaikkien eläinten munien sytoplasmasta.
Munasolun sytoplasman kemiallisen koostumuksen kuvattujen ominaisuuksien vuoksi alkio ei joissain tapauksissa käytä murskauksen aikana tsygootin ja blastomeerien ytimien geneettistä tietoa kehitykseen. Erityinen kemiallinen koostumus ja aineiden säännöllinen jakautuminen munasolun sytoplasmaan ovat erittäin tärkeitä alkion alkuvaiheissa. Varastoidut ravintoaineet ja energia-aineet varmistavat alkion kehityksen ilman, että niitä hankitaan ulkopuolelta.
7.2.3. MUNAN KOKO JA NIIDEN ROOLI EVOLUUTIOISSA. MUNALAJIT
Kehitysprosessissa paljastuu säännöllisyys, joka koostuu siitä, että mitä pidempi alkiojakso, sitä enemmän keltuaista tulisi kertyä munaan. Alkion ajanjakson kesto riippuu siitä, missä vaiheessa alkio siirtyy itsenäiseen olemassaoloon ulkoisessa ympäristössä. Jos postembryonaalinen kehitys on suoraa, ts. ilman toukkaa ja metamorfoosia, munassa pitäisi olla enemmän keltuaista. Keltuaisten lukumäärän mukaan jakomunat (taulukko 7.1) jaetaan alecital, oligo-, meso- Ja monisanainen, nuo. jossa on mitätön pieni, pieni, keskikokoinen ja suuri määrä keltuaista (kreikasta. lecithos- keltuainen) (taulukko 7.2).
Taulukko 7.1.Chordata-tyypin systematiikka
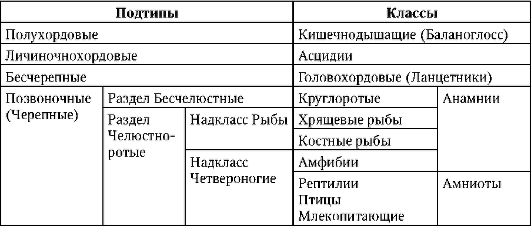
Taulukko 7.2.Oosyyttityypit löytyvät chordaateista

klo lansetti, alempien sointujen edustaja, muna oligolesitaalinen. Useimpien selkärankaisten munat sisältävät huomattavan määrän keltuaista. Alempien selkärankaisten joukossa (Anamnia) suurimmat munat löytyvät hagfishes (luokka Cyclostomes), hait ja kimeerit (luokka Rustokalat) ja eräillä sammakkoeläimillä. Loput sammakkoeläin, samoin kuin sammen munia mesolesitaali, nuo. on keskimääräinen määrä keltuaista. Korkeammissa selkärankaisissa (lapsivesi) kuten matelijat, linnut ja munasolut nisäkkäät, polylesitaalinen, nuo. munassa on paljon keltuaista. Niiden alkion kehitys kestää erityisen kauan.
Tämä kuvio on rikki pussieläimissä ja istukka nisäkkäät, joilla on oligo- Ja alesitaalinen munat, vastaavasti. Pussieläimillä alkio tulee ulos munakalvoista ja kohdusta epätäydellisen organogeneesin kanssa, siirretään pussiin, jossa se jatkaa kehitystä. Istukassa, myös ihmisissä, alkio tulee ulos
munakalvoista jopa aikaisemmin, blastokystivaiheessa, mutta sitten se siirtyy kohdunsisäiseen olemassaoloon, jossa se suorittaa kaikki tärkeimmät kehitysjaksot, jotka valmistelevat sitä syntymään. Vähennä nisäkkään munien keltuaisen määrää voidaan kutsua toissijainen, koska heidän esi-isänsä, jotka hallitsivat maaympäristö, oli, kuten kaikilla amnioteilla, polylesitaaleja munia.
7.2.4. OVA-NAPAUS
Munasolujen polariteetti se on suunniteltu jopa keltuaisen kerääntymisvaiheessa ovo(oo)syytteihin niiden nopean (suuren) kasvun aikana ja kiinnittyy polaaristen (pelkistys)kappaleiden eristämisen aikana. Toisen pelkistyskappaleen eristämisen jälkeen polariteetti muuttuu vakaaksi ja peruuttamattomaksi, mikä on todistettu Guerrierin kokeilla munien sentrifugoinnissa niiden kypsymisen eri vaiheissa. Napaa, jossa pelkistyskappaleet erottuvat, kutsutaan eläin, ja päinvastoin kasvullinen. Munasolun navat eroavat monin tavoin: keskittyminen erilaisia aineita, organellien lukumäärä, solunsisäisten prosessien aktiivisuus ja monet muut. Kyllä, kokeiluja Viime vuosina värähtelevien elektrodien avulla sähkökentät useiden eläinten ja kasvien munasolujen ja munien ympärillä sekä sähkövirtojen virtaaminen niiden sytoplasman läpi. Uskotaan, että tämä johtuu ionikanavien ja oosyyttien vastakkaisissa navoissa olevien pumppujen erilaisista pitoisuuksista. Kynsisammakon munassa ionikanavien pitoisuus on korkeampi eläinnavalla ja pumppujen pitoisuus on suurempi kasvullisessa navassa.
Keltuaisen kerääntyminen munan toimesta- sen polarisaation ensimmäinen ilmentymä. Kun munassa on pieni määrä keltuaista, se jakautuu yleensä tasaisesti sytoplasmaan ja ydin sijaitsee suunnilleen keskellä. Näitä munia kutsutaan isositaalinen(kreikasta. isos- yhtä suuri). Useimmilla selkärankaisilla on paljon keltuaista, ja se on jakautunut epätasaisesti munan sytoplasmaan. Tämä anisolesitaalinen soluja. Suurin osa keltuaisesta kertyy sisään kasvullinen napa. Näitä munia kutsutaan telolesitaalinen(kreikasta. telos- loppu). Päinvastoin eläin keltuaisesta vapaa aktiivinen sytoplasma työnnetään sivuun napaan. Jos keltuainen on edelleen upotettu sytoplasmaan, eikä sitä eristetä siitä erillisenä fraktiona, kuten sammissa ja sammakkoeläimissä, munat ovat ns. kohtalaisen telelesitaalinen. Jos keltuainen on täysin erotettu sytoplasmasta, kuten lapsivedessä, niin tämä jyrkästi telolesitaalinen munat.
Sointumunien koon ja napaisuuden ominaisuudet on ryhmitelty kaavamaisesti taulukkoon. 7.2.
Munan polarisaatioprosesseissa ilmeisesti osallistuu ja aivokuori- Tämä on munasolun sytoplasman pintakerros, joka sijaitsee suoraan plasmakalvon alla. Se sisältää mikrofilamentteja ja kortikaaliset rakeet. Jälkimmäiset sisältävät monia aineita, jotka osallistuvat hedelmöityskalvon muodostukseen ja estävät polyspermiaa. Hedelmöityksen jälkeen havaittu aivokuoren sytoskeleton elementtien kokoaminen ja uudelleenjakautuminen varmistaa sen supistumisen hankkimisen, mikä on välttämätöntä tsygootin pilkkoutumisen jakautumiselle. Useimmissa eläimissä kaksi ensimmäistä halkeamisuraa kulkevat keskenään kohtisuorassa eläin-kasvitason tasoja pitkin (meridiaaneja, jotka yhdistävät eläin- ja kasviperäiset navat).
Yleensä munan eläinnapassa on pääsääntöisesti enemmän vapaata sytoplasmaa, organelleja, varastoitunutta RNA:ta, yleensä ydin sijaitsee myös eläimen napassa tai lähempänä sitä. Vegetatiiviselle napalle on ominaista keltuaisten rakeiden hallitsevuus.
Munan polarisaatioon liittyy ulkonäkö ovo(oo)-plasminen segregaatio munat, ts. munasolun sytoplasman osien sisäisen heterogeenisyyden luominen.
Munan eläin-kasvi-polarisaatiolla on ratkaiseva merkitys kaikille myöhemmille alkiosyntymisprosesseille, koska se määrää alkion tulevan tilaorganisaation. Aikuisilla eläimillä kehon anteroposteriorinen akseli osuu yhteen munan eläin-vegetatiivisen akselin kanssa (esimerkiksi selkärankaisilla) tai on kohtisuorassa siihen nähden (esimerkiksi oligochaeteilla ja joillakin niveljalkaisilla).
7.2.5. MUNANKUORET
Munat peitetään ulkopuolelta yhdellä tai useammalla kalvolla, jotka myöhemmin suorittavat muun muassa kehittyvää alkiota suojaavaa tehtävää.
Erottaa ensisijainen kuori, munasolun itsensä muodostama toissijainen kuori- munasarjojen follikkelisolujen toiminnan tuote ja tertiääriset kuoret, jolla munaa ympäröi sen kulkiessa munanjohtimen läpi.
ensisijainen kuori, joskus kutsutaan keltuainen, löytyy kaikkien eläinten munista. Selkärankaisilla, mukaan lukien nisäkkäät,
Ensisijainen kuori on osa tiheää kuorta ja muodostaa sen sisäosan. Tiheän kuoren ulkoosa tuottaa follikulaarisia soluja toissijainen kuori. Tiheä kuori läpäisee sisäpuolelta munan mikrovillit ja ulkopuolelta follikulaaristen solujen mikrovillit. Heidän optiset ominaisuudet nisäkkäissä sitä kutsutaan kiiltävä kuori(zona pellucida). Joten tämä kuori yhdistää ensisijaisen ja toissijaisen. Päällä zona pellucida on säteilevä kruunu(corona radiata), muodostuu follikulaarisista soluista, jotka kiinnittyvät munasoluun sen ollessa munasarjassa (kuva 7.2).
Zona pellucida on monimutkainen kompleksi solunulkoisia glykoproteiineja, joita nisäkkäissä kutsutaan nimellä ZP. Hiirillä se koostuu kolmesta erilaisesta sulfatoidusta glykoproteiinista - ZP1, ZP2, ZP3. Käyttämällä ZP3 siittiöt sitoutuvat zona pellucidaan, ja yhden siittiön tunkeutumisen jälkeen tämä glykoproteiini modifioituu ja toisen siittiön tunkeutuminen
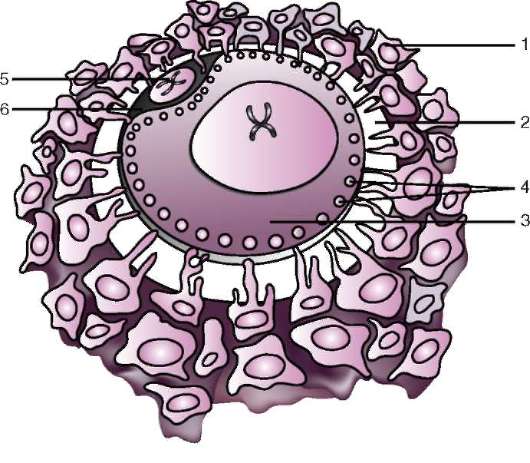
Riisi. 7.2. Nisäkkäiden munasolu (ensimmäisen asteen munasolu): 1 - säteilevä kruunu; 2 - kiiltävä kuori; 3 - sytoplasma; 4 - kortikaaliset rakeet; 5 - naparunko; 6 - keltuainen tila
gih siittiöistä tulee mahdottomaksi. Lisäksi glykoproteiinit ovat lajikohtaisia, mikä estää lajien välisen hedelmöittymisen. Muissa chordaateissa hedelmöityksen lajispesifisyys määräytyy siittiöiden proteiinien vuorovaikutuksesta reseptorien kanssa. keltuainen kalvo. Zona pellucida ei liity suoraan munakalvoon, vaan erottuu perivitelline (periyotelline) tila. Sen jälkeen, kun ensimmäinen siittiö on tunkeutunut munaan, aivokuoren rakeinen ovoperoksidaasi tulee tähän tilaan. Uskotaan, että tämän entsyymin vaikutus muuttuu ZP3 Ja Zp2, mikä johtaa zona pellucidan kovettumiseen. Se säilyy alkion ympärillä koko istutusta edeltävän ajanjakson tai merkittävän osan tästä ajanjaksosta. Kiiltävä kuori estää vierekkäisiä alkioita tarttumasta yhteen ja alkioita tarttumasta munanjohtimen ja kohdun seinämiin. Tiedetään, että murskauksen alkuvaiheessa blastokystiin asti blastomeereillä on korkea tarttuvuus. Jos alkio, josta puuttuu zona pellucida, siirretään munanjohtimeen, blastomeerit tarttuvat väistämättä munanjohtimen seinämään ja alkio kuolee. Lisäksi zona pellucidan ansiosta blastomeerit ovat kompakteja ja järjestyneitä, mikä edistää kontaktien muodostumista ja vuorovaikutusta niiden välillä ja varmistaa alkion normaalin kehityksen tässä vaiheessa. Jos kuori poistetaan, sirpaloituminen jatkuu, mutta blastomeerit asettuvat ketjuun ja niiden tiivistyminen häiriintyy kokonaan tai tulee hyvin myöhään.
Tertiääriset kuoret hyvin kehittyneitä rustokaloissa ja sammakkoeläimissä, mutta ne ovat erityisen monimutkaisia maaselkärankaisilla - matelijoilla, linnuilla ja alemmilla nisäkkäillä. Muodostaminen munanjohtimen rauhasten eritteistä, näillä kuorilla ei ole solurakennetta. Kaikissa selkärankaisissa ne suojaavat alkiota mekaanisilta vaurioilta ja haitallisten bioottisten tekijöiden, kuten bakteeri-, sieni- ja alkueläintekijöiden, vaikutuksilta. Lisäksi maan selkärankaisille ilmaantuu täysin uusia veden ja ravinteiden varastointitoimintoja vastaamaan alkion tarpeita. Matelijoilla kuorikalvo toimii pumppuna, joka ottaa vettä maaperästä ja ilmasta. Linnuilla on vesivarasto proteiinikuori. Veden imeytymistä ja haihtumista säätelevät kuorikalvon huokoset. Kuori sisältää monia mineraalisuoloja, joita tarvitaan alkion luuston kehittymiseen.
7.3. LANNOITUS JA PARTENOGEENESI
Lannoitus- tämä on sukusolujen fuusioprosessi, joka huipentuu niiden geneettisen materiaalin yhdistämiseen. Hedelmöityksen seurauksena syntynyt diploidisolu tsygootti- edustaa uuden organismin kehityksen alkuvaihetta.
Lannoitusprosessi koostuu kolmesta peräkkäisestä vaiheesta:
Sukusolujen etävuorovaikutus ja lähentyminen;
Sukusolujen kontaktivuorovaikutus ja munasolun aktivointi;
Sukusolujen yhdistäminen tai syngamia.
Siittiön lähentäminen munan kanssa ei-spesifisten tekijöiden yhdistelmä, joka lisää heidän tapaamisensa ja vuorovaikutuksensa todennäköisyyttä. Näitä ovat miehen ja naisen hedelmöitysvalmiuden alkamisen koordinointi, urosten ja naarasten käyttäytyminen, joka varmistaa parittelun ja siemennyksen, siittiöiden liiallinen tuotanto, munasolun suuri koko sekä munasolujen ja siittiöiden tuottamat siittiöt. kemialliset aineet, mikä edistää sukusolujen lähentymistä ja vuorovaikutusta. Nämä aineet ns hamons(sukusolujen hormonit) toisaalta aktivoivat siittiöiden liikettä ja toisaalta niiden sitoutumista. Nisäkkään siittiöiden liikkeessä munanjohtimen yläosia pitkin ilmiö reotaxis: niiden kyky liikkua vastaan tulevaa nestevirtausta munanjohtimissa.
Nisäkkäillä siittiöiden pysyminen naisen sukuelinten alueella on erittäin tärkeää, minkä seurauksena siittiöiden kapasitaatio- lannoituskyvyn hankkiminen. Heti sukupuolielimiin pääsyn jälkeen naisen siittiöt eivät pysty tunkeutumaan munasoluihin. Glykoproteiinit ja siemenplasmaproteiinit poistetaan siittiön plasmolemmasta akrosomialueella naisen sukupuolielinten salaisuuden aineiden vaikutuksesta, jotka estävät siittiön plasmolemman reseptorimolekyylien aktiiviset keskukset tunnistaen naisen pinnan. lisääntymissolu. Lisäksi naisen sukupuolielinten albumiinimolekyylit sitoutuvat siittiösolukalvon kolesteroliin. Tämä johtaa siittiön plasmalemman ja sen akrosomaalisen kalvon epävakauteen, mikä helpottaa myöhempää akrosomaalisten entsyymien vapautumista. Havaittiin myös, että kapasitaatioprosessissa siittiöiden pinnan ominaisuudet, kuten sen varaus, muuttuvat. Lisäksi tapahtuu muutos siittiöiden liikkuvuudessa (hyperaktivaatio) ja akrosomaalisten entsyymien aktivoituminen. Ihmisellä kapasitaatio kestää noin 7 tuntia.
Vaiheessa kontaktivuorovaikutusta siittiö tuhoaa munasolun kalvot (nisäkkäillä säteilevän kruunun, läpinäkyvän kalvon) ja munasolun sytoplasmisen kalvon akrosominen reaktio. Kun naarassukusolun kuori joutuu kosketukseen sen aktivoivien aineiden (joista yksi on fertilisiini) vaikutuksesta, kalsiumkationien aktiivinen virtaus siittiön päähän alkaa. Tämän seurauksena tapahtuu siittiöiden solun ja akrosomaalisten kalvojen fokaalinen fuusio ja niiden osittainen tuhoutuminen. Muodostuneiden mikroreikien kautta vapautuu siittiöiden entsyymejä - hyaluronidaasi, penetraasi ja muut, jotka katkaisevat kontaktit säteilevän kruunun solujen välillä sekä niiden ja munasolun välillä. Akrosomaalinen entsyymi akrosiini tuhoaa osan naaraspuolisen sukusolun zona pellucidasta ja siittiöt tunkeutuvat keltuaisen tilaan. Kohdassa, jossa siittiöpää koskettaa munasolun plasmakalvoa, tapahtuu naaras- ja miessukusolujen kalvojen fuusio ja myöhempi tuhoutuminen. Tuloksena olevan sytoplasmisen sillan kautta molempien sukusolujen sytoplasma yhdistyy. Sitten siittiön ydin ja sentrioli siirtyvät munasolun sytoplasmaan, ja siittiökalvo upotetaan munasolun kalvoon. Siittiön häntäosa joko jää ulkopuolelle tai menee myös munasoluun, mutta sitten erottuu ja erottuu ilman, että sillä on merkitystä jatkokehityksessä.
Natriumionit alkavat tunkeutua aktiivisesti munasolun sytoplasmaan siittiösolun kalvon kautta, minkä seurauksena munasolun kalvopotentiaali muuttuu dramaattisesti ja naisen sukusolusta tulee immuuni kosketuksiin muiden siittiöiden kanssa - nopea polyspermian lohko. Natrium-ionien sisäänvirtaus aiheuttaa kalsiumionien vapautumisen solunsisäisistä varastoista ja sen pitoisuuden kasvun munasolun sytoplasmassa, joka leviää aallon muodossa sukusolujen kosketuspisteestä. Tämän jälkeen alkaa kortikaalinen reaktio: aivokuoren rakeiden kalvot sulautuvat munan kalvoon ja niistä vapautuvat proteolyyttiset entsyymit pääsevät keltuaistilaan. Entsyymien vaikutuksesta munankuori paksuuntuu, paksuuntuu, menettää siittiöiden reseptoriproteiineja ja muuttuu lannoituskuori. Lisäksi kortikaalisista rakeista erittynyt glykoproteiini edistää keltuaisen kalvon irtoamista munan plasmolemmasta. Kaiken edellä mainitun seurauksena muiden siittiöiden tunkeutuminen tulee mahdottomaksi - hidas polyspermian esto. Nisäkkäillä
kortikaalinen reaktio ei aiheuta hedelmöityskalvon muodostumista, mutta sen olemus on sama.
Siittiön ja munasolun kosketuksen seurauksena se aktivointi. Se koostuu monimutkaisista rakenteellisista ja fysikaalis-kemiallisista muutoksista. Aktivoinnin alkuvaihe on edellä kuvattu aivokuoren reaktio. Eläimillä, kuten piikkinahkaisilla, luisilla kaloilla ja sammakkoeläimillä, munan sytoplasman muutoksiin liittyy näkyviä morfologisia uudelleenjärjestelyjä. Näitä ilmiöitä kutsutaan kerrostumiseksi tai plasman segregaatio. Sen merkitystä alkion jatkokehityksen kannalta käsitellään jäljempänä. Monissa anuranlajeissa siittiöiden tunkeutuminen munaan johtaa pigmenttirakeiden liikkuminen eläinnapa, ja siittiöiden tunkeutumiskohtaa vasten ilmestyy heikosti värillinen puolikuun muotoinen alue, ns. "harmaa sirppi"(Kuva 7.3).
Munan aktivoituminen päättyy proteiinisynteesin alkamiseen translaatiotasolla, koska mRNA, tRNA, ribosomit ja energia varastoituvat takaisin ovo(oo)geneesiin. Munasolujen aktivaatio voi alkaa ja jatkua loppuun ilman siittiöymaa ja ilman munasolua, mikä on todistettu tsygoottien enukleaatiokokeilla.
Munasolu on tapaamishetkellä siittiöiden kanssa yleensä jossakin meioosin vaiheista tietyn tekijän estämänä. Useimmissa selkärankaisissa tämä lohko suoritetaan
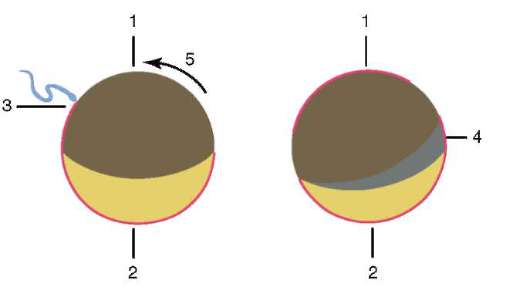
Riisi. 7.3. Sammakkoeläinmunan sytoplasman erottelu siittiöiden tunkeutumisen jälkeen: 1 - eläinnapa; 2 - kasvullinen napa; 3 - siittiön tulopaikka; 4 - harmaa sirppi; 5 - sytoplasman liikesuunta pigmenttirakeiden kanssa
metafaasin II vaiheessa; monissa selkärangattomissa sekä kolmessa nisäkäslajissa (hevoset, koirat ja ketut) esto tapahtuu diakineesivaiheessa (profaasi I). Useimmissa tapauksissa meioottinen tukos poistetaan munasolun aktivoitumisen jälkeen hedelmöittymisen vuoksi. Kun meioosi on päättynyt munassa, siihen tunkeutunut siittiön ydin muuttuu. Se on interfaasin ja sitten profaasiytimen muodossa. Tänä aikana DNA kaksinkertaistuu ja määrä perinnöllinen materiaali V miehen pronukleus tulee p2s, nuo. se sisältää haploidisen joukon toistuvia kromosomeja.
Munasolun ytimestä, joka on suorittanut meioosin, tulee naisen pronukleus, myös hankkimassa p2s. Molemmat proytimet tekevät monimutkaisia liikkeitä, lähestyvät sitten toisiaan ja sulautuvat yhteen. (synkario), muodostaen yhteisen metafaasilevyn. Tämä on itse asiassa sukusolujen lopullisen fuusion hetki - syngamia. Tsygootin ensimmäinen mitoottinen jakautuminen johtaa kahden alkiosolun (blastomeerin) muodostumiseen, joissa on joukko kromosomeja 2p2s kaikissa.
Joissakin tapauksissa kehitys tapahtuu ilman hedelmöitystä - partenogeneesi(kreikasta. parthenoe- Neitsyt). Kun luonnollinen partenogeneesi kehitys etenee munasolun sytoplasman ja esituman perusteella. Munasta muodostuvilla yksilöillä on joko haploidi tai diploidi kromosomisarja, koska yksi kromosomien lukumäärän kaksinkertaistamisen mekanismeista laukeaa. Joissakin tapauksissa naisen sukusolun meioosin aikana kromosomien lukumäärän vähenemisvaihe putoaa ja muna saadaan diploidisella pronukleuksella. Muissa tapauksissa diploidisaatio tapahtuu ensimmäisen katkaisun aikana, jolloin sytotomiaa ei tapahdu.
luonnollinen partenogeneesi, se ei yleensä ole lajin ainoa lisääntymistapa. Se joko vuorottelee normaalin seksuaalisen lisääntymisen kanssa tai esiintyy yksilössä Kisat. Luonnollista partenogeneesiä on havaittu joidenkin äyriäisten ja rotiferien kesäsukupolvilta, mehiläisiltä, ampiaisilta ja useilta perhosilta. Selkärankaisilla partenogeneettistä lisääntymistä on kuvattu kolmessa Armenian kallioliskojen rodussa, jotka koostuvat vain naaraista. 40 % uroksen poissa ollessa munituista kalkkunan munista voi alkaa kehittyä, mutta tämä kehitys harvoin päätyy, useammin se pysähtyy poikkeavuuksien vuoksi. Muiden selkärankaisten lajien luonnollista partenogeneettistä lisääntymistä ei tunneta.
keinotekoinen partenogeneesi näyttää olevan mahdollista kaikissa eläimissä. Partenogeneettisen kehityksen menetelmien kehittäminen on tärkeää
ongelma tieteellisissä ja soveltavissa suhteissa. Suuren panoksen tähän ongelmaan antoivat kotimaiset tutkijat A.A. Tikhomirov, B.L. Astaurov, V.A. Strunnikov. Havaittiin, että siittiön aktivoituminen munasolusta ei ole spesifistä. Monet fysikaaliset ja kemialliset tekijät voivat toimia aktivaattoreina. Silkkiäistoukkien kohdalla osoitettiin, että keinotekoisen partenogeneesin avulla on mahdollista säädellä miesten ja naisten sukupuolen suhdetta väestössä, jolloin saadaan suuri taloudellinen vaikutus.
Luonnollinen partenogeneesi tapahtuu useimmiten epätäydellisen hedelmöityksen yhteydessä, ts. niissä tapauksissa, joissa munasolu aktivoitui, mutta siittiön ydin ei osallistunut hedelmöitykseen. Aktivoiduissa munissa käytetään vain naaraan esitumasta saatua tietoa. Tämän tyyppistä partenogeneesiä kutsutaan gynogeneesi. Keinotekoisella partenogeneesillä on mahdollista poistaa naaraspronukleus, ja sitten kehitys tapahtuu vain miespuolisten proytimien kustannuksella. Tämä an-drogeneesi. Merisiileillä tehdyissä erityisissä kokeissa havaittiin, että jälkeläiset perivät joko vain emon piirteet gynogeneesin aikana tai vain isän piirteet androgeneesin aikana. Tämä osoittaa, että yksilön perinnölliset ominaisuudet määräytyvät pääasiassa ytimestä, ei sytoplasmasta.
7.4 alkion kehitys
7.4.1. EROTA
7.4.1.1. Murskausvaiheen ydin
Erota- tämä on sarja tsygootin ja tuloksena olevien blastomeerien peräkkäisiä mitoottisia jakautumisia, jotka päättyvät muodostumiseen monisoluinen yksikerroksinen alkio- blastula. Ensimmäinen pilkkoutumisjako alkaa esitumien perinnöllisen materiaalin yhdistymisen ja yhteisen metafaasilevyn muodostumisen jälkeen. Pilkkomisen aikana muodostuneita soluja kutsutaan blastomeerit(kreikasta. räjähtää- verso, alkio). Murskauksen mitoottisten syklien ominaisuus on, että jokaisella jakautumisella solut pienenevät ja pienenevät, kunnes ne saavuttavat somaattisille soluille tavanomaisen ytimen ja sytoplasman tilavuuksien suhteen. Esimerkiksi merisiilissä tämä vaatii 6 jakautumista ja alkio koostuu 64 solusta. Peräkkäisten jakautumisten välillä ei tapahdu solujen kasvua (jakso G1 poissa), mutta DNA on välttämättä syntetisoitu. Kaikki DNA:n esiasteet ja tarpeelliset
Himmennettävät entsyymit kertyvät oogeneesiprosessiin. Tämän seurauksena mitoosisyklit lyhenevät ja jakautumiset seuraavat toisiaan paljon nopeammin kuin tavallisissa somaattisissa soluissa. Useiden jakautumisjaksojen jälkeen muodostuneet blastomeerit rajoittuvat toisiinsa muodostaen soluklusterin ns. morula. Sitten solujen väliin muodostuu ontelo - blastocoel, täynnä nestettä. Solut työnnetään kehälle muodostaen blastulan seinämän - blastodermi. Koska blastomeerikasvua ei esiinny katkaisujakson aikana, alkion kokonaiskoko blastulavaiheessa ei ylitä tsygootin kokoa.
7.4.1.2. Murskauksen morfologia
Yleensä blastomeerit on järjestetty tiukasti toisiinsa ja munan napa-akseliin nähden. Murskausjärjestys tai -menetelmä riippuu keltuaisen määrästä, tiheydestä ja jakautumisesta munassa. Sachs-Hertwigin sääntöjen mukaan soluytimellä on taipumus sijaita sytoplasman keskellä ilman keltuaista, ja solun jakautumiskara - tämän vyöhykkeen suurimman laajuuden suuntaan.
Oligo- ja mesolesitaalimunissa pilkkoutuminen saattaa loppuun, tai holoblastinen. Tämän tyyppistä murskaamista esiintyy lanseteissa, nahkiaisissa, joissakin kaloissa, kaikissa sammakkoeläimissä sekä pussieläimissä ja istukkanisäkkäissä. Täydellisen murskaamisen yhteydessä ensimmäisen jaon taso vastaa kahdenvälisen symmetriatasoa. Toisen jaon taso kulkee kohtisuorassa ensimmäisen jaon tasoon nähden. Kahden ensimmäisen jaon molemmat uurteet ovat meridiaaneja, ts. alkaa eläinnapasta ja leviää kasvulliseen napaan. Munasolu on jaettu neljään suurin piirtein samankokoiseen blastomeeriin. Kolmannen jaon taso kulkee leveyssuunnassa kohtisuorassa kahteen ensimmäiseen nähden. Eläimillä, joilla on isolesitaalisia munia, kaikki kahdeksan muodostunutta blastomeeria ovat suunnilleen yhtä suuret - yhtenäinen murskaus. Mesolecithal munissa se näkyy epäsäännöllisyys murskaamalla. Eläinpaalussa on neljä pienempää blastomeeria - mikrometriä, vegetatiivisella - neljä isompaa - makromeerit. Sitten jako tapahtuu jälleen meridiaanitasoilla ja sitten taas leveysasteilla.
Luisten kalojen, matelijoiden, lintujen ja myös monotreemien nisäkkäiden polylesitaalisissa munasoluissa pilkkoutuminen osittainen, tai meroblastinen, nuo. kattaa vain keltuaisesta vapaan sytoplasman. Se sijaitsee ohuen kiekon muodossa eläinpaalussa, joten tämän tyyppistä murskaamista kutsutaan kiekkomainen.
Murskaustyyppiä karakterisoitaessa otetaan huomioon myös blastomeerien suhteellinen sijainti ja jakautumisnopeus. Jos blastomeerit on järjestetty riveihin päällekkäin säteitä pitkin, murskaus on ns. säteittäinen. Se on tyypillistä chordaateille ja piikkinahkaisille. Luonnossa on muitakin muunnelmia blastomeerien tilajärjestelystä murskaamisen aikana, mikä määrää sen tyypit, kuten esim. kierreäyriäisissä kahdenvälinen sukkulamatossa, anarkistinen medusassa.
Keltuaisen jakautumisen ja eläin- ja kasviperäisten blastomeerien jakautumisen synkronismiasteen välillä havaittiin yhteys. Piikkinahkaisten ja päänahkaisten oligolesitaalisissa munissa pilkkoutuminen on lähes synkronista, esimerkiksi mesolesitaalisissa munissa sammakkoeläimissä synkronia häiriintyy kolmannen jakautumisen jälkeen, koska vegetatiiviset blastomeerit jakautuvat hitaammin suuren keltuaisen määrän vuoksi. Eliöissä, joissa on osittainen pilkkoutuminen (matelijat, linnut), jakautuminen on alusta alkaen asynkronista ja keskeisellä paikalla olevat blastomeerit jakautuvat nopeammin.
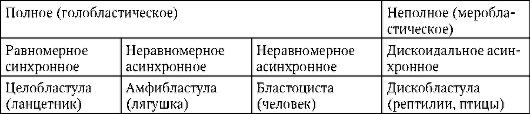
Murskauksen loppuun mennessä, blastula. Blastulan tyyppi riippuu murskaustyypistä ja siten munatyypistä. Jotkin murskaus- ja blastulatyypit on esitetty kuvassa. 7.4 ja kaavio (taulukossa) 7.1. Tarkempi kuvaus pilkkomisesta nisäkkäillä, mukaan lukien ihmiset, katso kohta 7.5.1.
Taulukko 7.3.Murskaustyypit ja chordate blastula -tyypit
7.4.1.3. Molekyyligeneettisten ja biokemiallisten prosessien ominaisuudet murskaamisen aikana
Kuten edellä todettiin, mitoottiset syklit katkaisujakson aikana lyhenevät suuresti, erityisesti aivan alussa. Esimerkiksi merisiilin munien koko fissiosykli kestää 30-40 minuuttia ja kestää n. S-vaiheet vain 15 min.
Gj-, ja monissa eläimissä G 2 - jaksot ovat käytännössä poissa, koska munasolun sytoplasmaan on luotu tarvittava tarjonta kaikista aineista, ja mitä suurempi, sitä suurempi se on. On osoitettu, että tsygootista poistettaessa
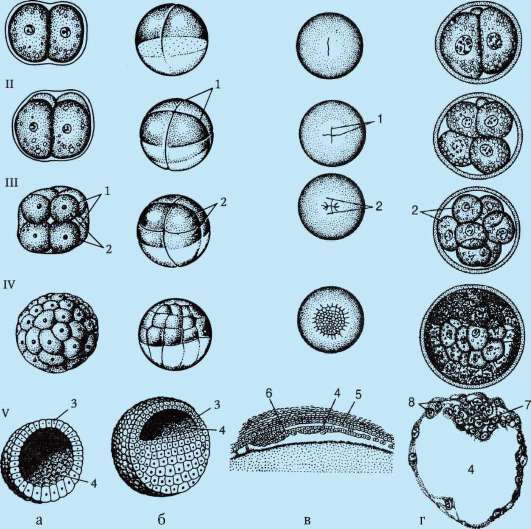
Riisi. 7.4 Halkeaminen chordaateissa erityyppisillä munilla: a - lansetti; b - sammakko; c - lintu; d - nisäkäs: I - 2 blastomeeriä; II - 4 blastomeeriä; III - 8 blastomeeriä; IV - morula; V - blastula; 1 - uurteiden murskaus; 2 - blastomeerit; 3 - blastodermi; 4 - blastocoel; 5 - epiblasti; 6 - hypoblasti; 7 - embryoblasti; 8 - trofoblasti; kuvassa olevien alkioiden koko ei vastaa alkioiden todellista kokoa
ytimen katkeaminen tapahtuu ja alkio saavuttaa kehityksessään lähes blastula-vaiheen. Jatkokehitys pysähtyy.
Ennen jokaista jakautumista tapahtuu DNA:n ja histonien synteesi, kuitenkin kesto S-jakso lyhennetty. DNA:n replikaation aikana tapahtuvan fragmentoinnin aikana replikaatiohaarukan etenemisnopeus on normaali. Samaan aikaan blastomeeri-DNA:ssa on enemmän aloituspisteitä kuin sisällä
somaattisia soluja, joten replikonit ovat lyhyempiä. DNA-synteesi tapahtuu kaikissa replikoneissa samanaikaisesti (synkronisesti). Siksi DNA:n replikaation aika ytimessä osuu yhteen yhden, lisäksi lyhennetyn replikonin kaksinkertaistumisajan kanssa.
Pilkkomisen alussa muun tyyppinen ydinaktiivisuus, kuten transkriptio, puuttuu käytännössä. Erityyppisissä munissa geenin transkriptio ja RNA-synteesi alkavat eri vaiheissa. Tapauksissa, joissa sytoplasmassa on monia erilaisia aineita, kuten esimerkiksi sammakkoeläimissä, transkriptio ei aktivoidu heti. RNA-synteesi niissä alkaa varhaisen blastulan vaiheessa. Päinvastoin, nisäkkäillä RNA-synteesi alkaa jo kahden blastomeerin vaiheessa.
Katkaisujakson aikana muodostuu RNA:ta ja proteiineja, jotka ovat samanlaisia kuin oogeneesin aikana syntetisoituneet. Enimmäkseen histonit, proteiinit solukalvot ja solujen jakautumiseen tarvittavat entsyymit. Näitä proteiineja käytetään välittömästi yhdessä munien sytoplasmaan aiemmin varastoituneiden proteiinien kanssa. Tämän lisäksi katkaisujakson aikana proteiinien synteesi, joka ei ollut aikaisemmin mahdollista, on mahdollista, mistä todistavat tiedot alueellisista eroista RNA:n ja proteiinien synteesissä blastomeerien välillä. Joskus nämä aineet alkavat vaikuttaa myöhemmissä vaiheissa.
Tärkeä rooli murskaamisessa on sytoplasman jakautumisella - sytotomia. Sillä on erityinen morfogeneettinen merkitys, koska se määrittää murskaustyypin. Sytotomiaprosessissa muodostuu ensin supistuva mikrofilamenttien renkaan avulla. Tämän renkaan kokoaminen tapahtuu mitoottisen karan napojen suoran vaikutuksen alaisena. Sytotomian jälkeen oligolesitaalien munien blastomeerit pysyvät kytkettyinä toisiinsa vain ohuilla silloilla. Juuri tähän aikaan ne on helpoin erottaa toisistaan. Tämä johtuu siitä, että sytotomia johtaa solujen välisen kosketusalueen pienenemiseen kalvojen rajallisen pinta-alan vuoksi.
Välittömästi sytotomian jälkeen solupinnan uusien osien synteesi alkaa, kosketusvyöhyke kasvaa ja blastomeerit alkavat koskettaa tiukasti, mikä tarjoaa mahdollisuuden solujen väliseen vuorovaikutukseen. Halkaisuurat kulkevat ovoplasman yksittäisten osien välisiä rajoja pitkin, joten eri blastomeerien sytoplasman kemiallinen koostumus eroaa. Tämä kuvastaa ilmiötä ovoplasminen segregaatio.
Murskausjakson päätulos on yksisoluisen alkion - tsygootin - muuttuminen monisoluinen Ja yksikerroksinen alkio - blastula. Lisäksi murskauksen aikana muodostuu
ing solujen välisiä kontakteja ja niiden pinta-alan kasvu, tsygootin sytoplasman eri osien jakautuminen alkion eri blastomeerien välillä, alkion oman genomin asteittainen aktivointi, somaattisille soluille ominaisen tuman ja sytoplasman suhteen palautuminen. Murskausvaiheessa kaikki alkion solut ovat homogeenisia suhteessa geneettisen laitteen toiminnalliseen tilaan. Ero blastomeerien sytoplasmassa ja solujen väliset vuorovaikutukset määräävät (ennalta määrittävät) solun jatkokehityksen suunnan.
7.4.2. GASTRULATION
7.4.2.1. Gastrulaatiovaiheen olemus
Gastrulaatiovaiheen olemus on, että yksikerroksinen alkio - blastula - muuttuu monikerroksinen- kaksi- tai kolmikerroksinen, nimeltään gastrula(kreikasta. gaster- maha ja deminutiivinen jälkiliite "st").
Alkuperäisissä chordaateissa, esimerkiksi lansetissa, homogeeninen yksikerroksinen blastodermi muuttuu gastrulaation aikana ulommaksi itukerrokseksi - ektoderma- ja sisäinen itukerros - endodermi. Endodermi muodostaa ensisijaisen suolen, jonka sisällä on onkalo - gastrocoel. Gastrocoeliin johtavaa reikää kutsutaan blastopore tai ensisijainen suu. Kaksi alkiokerrosta ovat määrittäviä morfologinen ominaisuus gastrulaatio. Niiden olemassaolo tietyssä kehitysvaiheessa kaikissa monisoluisissa eläimissä, coelenteraateista korkeampiin selkärankaisiin, antaa meille mahdollisuuden pohtia itukerrosten homologiaa ja kaikkien näiden eläinten alkuperän yhtenäisyyttä.
Selkärankaisilla kahden mainitun itukerroksen lisäksi muodostuu kolmas gastrulaation aikana - mesoderma, sijaitsee ekto- ja endodermin välissä. Keskimmäisen itukerroksen kehitys, joka on chordomesoderma,- selkärankaisten gastrulaatiovaiheen evoluutiokomplikaatio, joka liittyy niiden kehityksen kiihtymiseen alkion alkuvaiheessa. Primitiivisemmissä sointuissa, kuten lansetissa, chordomesoderma muodostuu yleensä gastrulaation jälkeisen vaiheen alussa - organogeneesi. Jälkeläisten joidenkin elinten kehitysajan muutos suhteessa muihin verrattuna esi-isiisi ryhmiin on ilmentymä heterokroniaa. Muutokset evoluution aikana tärkeimpien elinten muodostumisen ajoituksessa ovat melko yleisiä.
Gastrulaatioprosessille on tunnusomaista tärkeitä solumuutoksia, kuten ryhmien ja yksittäisten solujen suunnatut liikkeet, solujen selektiivinen lisääntyminen ja lajittelu, sytodifferentioitumisen alkaminen ja induktiovuorovaikutukset. Lueteltuja ontogeneesin solumekanismeja käsitellään yksityiskohtaisesti kohdassa 8.2.
Gastrulaatiomenetelmät ovat erilaisia. Erotetaan neljän tyyppisiä tilallisesti suunnattuja soluliikkeitä, jotka johtavat alkion muuttumiseen yhdestä kerroksesta monikerroksiseksi (kaksi tai kolmikerroksiseksi).
Intussusceptio- yhden blastodermin osan invaginaatio sisäänpäin kokonaisena kerroksena. Lansetissa vegetatiivisen navan solut tunkeutuvat, sammakkoeläimillä intussusseptio esiintyy eläimen ja kasvullisen navan välisellä rajalla harmaan puolikuun alueella. Invaginaatioprosessi on mahdollista vain munissa, joissa on pieni tai keskimääräinen keltuainen.
epibolia- likaantuminen kasvullisen navan pienten nopeasti jakautuvien solujen ja suurempien, jakautumisnopeudessa jäljessä olevien ja vähemmän liikkuvien solujen kanssa. Tämä prosessi ilmaistaan selvästi sammakkoeläimissä.
Delaminaatio- blastodermisolujen kerrostuminen kahdeksi kerrokseksi, jotka ovat päällekkäin. Delaminaatiota voidaan havaita alkioiden discoblastulassa, kun eläimiä, kuten matelijoita, lintuja ja munasoluisia nisäkkäitä, murskataan epätäydellisesti. Delaminaatio ilmenee istukan nisäkkäiden alkioblasteissa, mikä johtaa hypoblastien ja epiblastien muodostumiseen.
Maahanmuutto- ryhmien tai yksittäisten solujen liikkuminen, jotka eivät ole yhdistyneet yhdeksi kerrokseksi. Maahanmuuttoa tapahtuu kaikissa alkioissa, mutta se on tyypillisintä korkeampien selkärankaisten (matelijat, linnut ja nisäkkäät) gastrulaation toiselle vaiheelle.
Jokaisessa erityisessä alkion synnyn tapauksessa yhdistetään yleensä useita gastrulaatiomenetelmiä.
7.4.2.2. Gastrulaation morfologia
Tarkempi gastrulaation tutkimus lansetilla, sammakolla, kanalla ja nisäkkäillä auttaa ymmärtämään paremmin evoluutiosuhteita ja ymmärtämään yksilön kehityksen malleja.
7.4.2.2-a. Lansetin gaetrulaatio
gastrulaatio lansetti esitetty kuvassa. 7.5 Eri markkerit blastula-vaiheessa (kuva 7.5, a) merkitty oletettu(väitetty) alkeet. Nämä ovat solumateriaalista peräisin olevia blastulan alueita
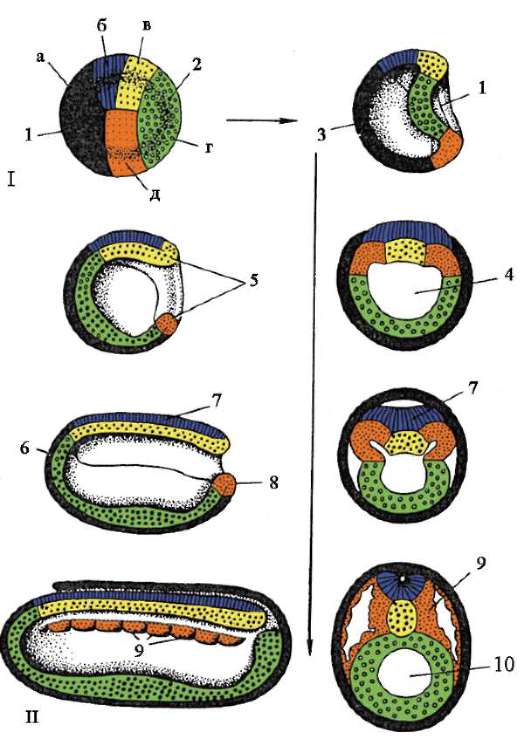
Riisi. 7.5Oletettavat alkeet, gastrulaatio ja hermotus lansetissa. I - oletetut alkeet blastulan (ulkokuva) ja varhaisen gastrulan (leikkausnäkymä) vaiheessa; II - myöhäinen gastrula ja neurulaatio sagittaalisessa (vasemmalla rivillä) ja poikittaisessa (oikea rivi) osassa; III - alkion muovinen malli hermojakson lopussa:
1 - eläintanko; 2 - kasvullinen napa; 3 - blastocoel; 4 - gastro-maali; 5 - blastoporen selkä- ja vatsahuulet; 6 - alkion pää; 7 - medullaarinen levy; 8 - alkion häntäpää; 9 - mesodermin selkäosa; 10 - sekundaarisen suolen ontelo; 11 - segmentoidut somiitit; 12 - mesodermin vatsaosa; a, b, c, d, e - oletettujen ja kehittyvien elinten nimitykset: a - ihon ektoderma; b - hermoputki; in - sointu; d - endodermi, suolen epiteeli; d - mesoderma
joista gastrulaation ja varhaisen organogeneesin (neurulaation) aikana muodostuu yleensä täysin määritellyt itukerrokset ja -elimet (kuva 7.5, b ja c).
Invaginaatio alkaa vegetatiivisesta napasta. Nopeamman jakautumisen ansiosta eläinnavan solut kasvavat ja työntävät vegetatiivisen navan solut blastulaan. Tätä helpottaa sytoplasman tilan muutos soluissa, jotka muodostavat blastoporen huulet ja niiden vieressä. Invaginaatiosta johtuen blastokoel vähenee ja gastrocoel kasvaa. Samanaikaisesti blastokoelin katoamisen kanssa ektodermi ja endodermi joutuvat läheiseen kosketukseen. Lansetti, kuten kaikki muutkin deuterostomes(näihin kuuluvat Echinodermata, Chordata ja eräät muut suhteellisen pienet eläintyypit), blastohuokosalue muuttuu organismin häntäosaksi toisin kuin protostomit, joissa blastopore vastaa pääosaa. Suun aukko deuterostomissa muodostuu alkion päähän vastapäätä blastoporea.
7.4.2.2-6. Gaetrulaatio sammakkoeläimissä
Sammakkoeläinten gastrulaatiolla on paljon yhteistä lanseletin gastrulaation kanssa, mutta koska niiden munien keltuainen on paljon suurempi ja se sijaitsee pääasiassa vegetatiivisessa navassa, amfiblastulan suuret blastomeerit eivät pysty pullistumaan sisäänpäin. Intussusceptio menee vähän toisin. Eläin- ja kasvunapojen välisellä rajalla harmaan sirpin alueella solut venyvät ensin voimakkaasti sisäänpäin ottamalla "pullomaisen" muodon (kuva 7.6, a) ja vetävät sitten pintakerroksen soluja. blastula heidän mukanaan. Näkyviin tulee puolikuun muotoinen ura ja blastohuokosen dorsaalinen (dorsaalinen) huuli (kuva 7.6, b).
Samanaikaisesti pienemmät eläinnavan solut, jotka jakautuvat nopeammin, alkavat liikkua kohti kasvullista napaa. Selkähuulen alueella ne kääntyvät ylös ja tunkeutuvat, ja sirpin muotoista uraa vastakkaisilla sivuilla ja puolella kasvaa suurempia soluja. Sitten prosessi epibolia johtaa blastoporen lateraalisten (lateral) ja ventraalisten (ventraalisten) huulien muodostumiseen. Blastohuoko sulkeutuu renkaaksi, jonka sisällä vegetatiivisen navan suuret valosolut näkyvät jonkin aikaa ns. keltuainen tulppa. Myöhemmin ne upotetaan kokonaan sisäänpäin ja blastohuokos kapenee.
Solujen liikkeitä on tutkittu yksityiskohtaisesti käyttämällä merkintämenetelmää sammakkoeläinten elintärkeillä (vitaali)väreillä
Riisi. 7.6Sammakkoeläinten varhaisen gastrulan blastohuokosen muodostuminen: a - pullon muotoiset solut blastohuokosen selkähuulen alueella; b - yleiskuva falciformisesta urasta ja selkähuulesta; 1 - pullon muotoiset solut; 2 - blastoporen selkähuuli
blastula gastrulaation aikana, on havaittu, että tietyt blastodermin alueet, ns oletettu(alkaen lat. praesumptio- oletus), normaalin kehityksen aikana ne ilmenevät ensin tiettyjen elinten alkuaineissa ja sitten itse elinten koostumuksessa (kuva 7.7). Tiedetään, että hännänttömillä sammakkoeläimillä blastula-vaiheen oletetun notokoordin ja mesodermin materiaali ei ole sen pinnalla, vaan amfiblastulan seinämän sisäkerroksissa, kuitenkin suunnilleen samalla tasolla kuin kuvassa.
Tietty samankaltaisuus sammakkoeläinten ja lansettien gastrulaatioprosessien ja oletettujen elinten alueiden välillä, ts. pääelinten, kuten hermoputken, notochordin, sekundaarisuolen, homologia osoittaa niiden fylogeneettisen suhteen.
7.4.2.2-c. Gastrulaatio linnuilla
Gastrulaatiolla alkioissa, joissa on meroblastinen pilkkoutuminen ja kehitys, on omat ominaisuutensa. klo lintuja se alkaa blastulan halkeamisen ja muodostumisen jälkeen alkion kulkiessa munanjohtimen läpi. Kun muna munitaan, alkio koostuu jo useista kerroksista: yläkerrosta kutsutaan epiblasti, alempi - primaarinen hypoblastooma(katso kuva 7.4, c). Niiden välissä on kapea rako - blastocoel. Sitten muodostettiin sekundaarinen hypoblasti, joiden muodostumistapa ei ole täysin selvä. Siitä on näyttöä ensisijassa
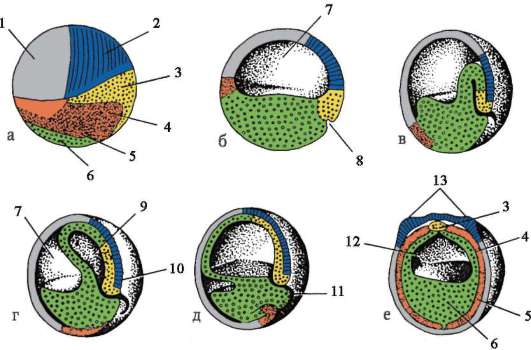
Riisi. 7.7. Kartta alueista, joissa oletetut elinten alkuaineet sammakkoeläinten alkionkehityksen varhaisissa vaiheissa: a - blastula-vaihe (vasen kuva); b-e - peräkkäiset gastrulaation vaiheet (sagittaaliset osat); e - neurulaation alku (poikkileikkaus); 1 - ihon ektoderma; 2 - hermoputki; 3 - sointu; 4 - somiitti mesodermi; 5 - splanknotomien mesodermi; 6 - endodermi; 7 - blastocoel; 8 - puolikuun ura; 9 - gastrocoel; 10 - dorsaalihuulen blastopore; 11 - keltuainen tulppa; 12 - sekundaarisen suolen ontelo; 13 - hermorullat
Lintujen alueella primaariset sukusolut ovat peräisin ja sekundaariset muodostavat alkion ulkopuolisen endodermin. Primaarisen ja sekundaarisen hypoblastin muodostumista pidetään gastrulaatiota edeltävänä ilmiönä.
Gastrulaation päätapahtumat ja kolmen itukerroksen lopullinen muodostuminen alkavat munanpoiston jälkeen inkuboinnin alkaessa. Epiblastin takaosaan on kertynyt soluja, mikä johtuu solujen jakautumisen epätasaisesta nopeudesta ja niiden liikkumisesta epiblastin sivuosista keskustaan, toisiaan kohti. Niin kutsuttu päälinja, joka ulottuu päätä kohti. Keskellä ensisijainen putki muodostuu päävao, ja reunoja pitkin ensisijaiset rullat. Ensisijaisen nauhan päähän ilmestyy paksuuntumaa - Hensen-solmu, ja siinä - ensisijainen kuoppa(Kuva 7.8).
Kun epiblastisolut tulevat primääriuraan, niiden muoto muuttuu. Ne muistuttavat muodoltaan sammakkoeläinten gastrulan "pullon muotoisia" soluja. Nämä solut muuttuvat sitten tähtiksi ja uppoavat epiblastin alle muodostaen mesodermin. Osa siirtyvistä soluista, jotka uppoavat hypoblastiin, synnyttävät myöhemmin ituen endodermin (kuva 7.9). Siten endodermi muodostuu primaarisen ja sekundaarisen hypoblastin pohjalta lisäämällä uuden sukupolven endodermaalisia soluja, jotka siirtyvät blastodermin ylemmistä kerroksista. Useiden sukupolvien endodermaaliset solut osoittavat gastrulaatiojakson pitkittymistä ajassa.
Osa epiblastista Hensenin solmun kautta siirtyvistä soluista muodostaa tulevan notokordin. Samanaikaisesti jänteen venymisen ja venymisen kanssa Hensenin solmu ja ensisijainen juova siirtyvät vähitellen etupäästä kaudaaliseen päähän ja katoavat. Tämä vastaa blastohuokosen kapenemista ja sulkeutumista. Kun ensisijainen juova supistuu, se jättää jälkeensä alkion aksiaalisten elinten muodostuneet osat päässä päästä hännän osiin. Vaikuttaa järkevältä pitää kanan alkion soluliikkeitä homologisina sammakkoeläinten liikkeen kanssa ja ensisijaista juovaa ja Hensenin solmua homologisina näiden eläinten gastrulan blastohuokoselle ja dorsaaliselle (dorsaaliselle) huulelle.
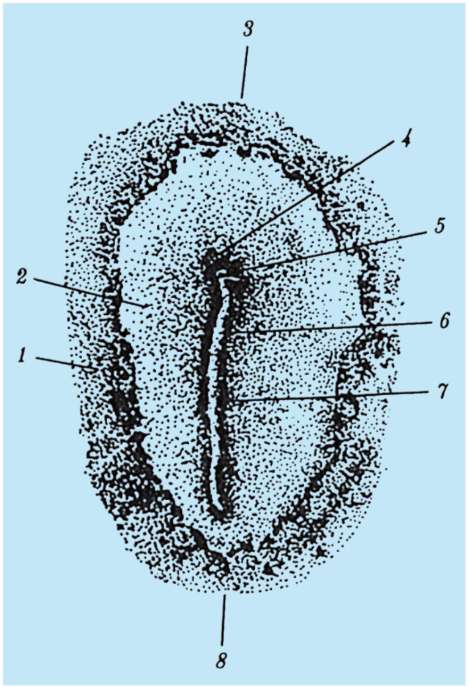
Riisi. 7.8Kanan alkio primaarisen juovan vaiheessa (selkänäkymä): 1 - tumma alue; 2 - itulevyn läpikuultava alue; 3 - pää; 4 - Hensenin solmu; 5 - primaarinen kuoppa; 6 - ensisijainen ura; 7 - ensisijainen rulla; 8 - häntäpää
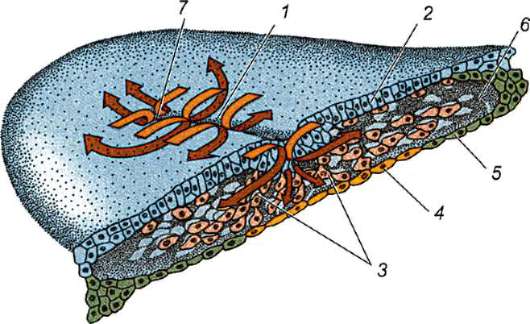
Riisi. 7.9.Kanan alkio ensisijaisen nauhan vaiheessa (poikkileikkaus): 1 - ensisijainen ura; 2 - epiblasti (ektoderma); 3 - vaeltavat solut (mesodermi); 4 - germinaalinen endodermi; 5 - hypoblasti (endodermi); 6 - blastocoel; 7 - Hensenin solmu
7.4.2.2-d. Gastrulaatio nisäkkäillä
On mielenkiintoista huomata, että nisäkäsalkioiden solut (katso kohta 7.5.1) säilyttävät gastrulaatiovaiheessa niille ominaiset liikkeet huolimatta siitä, että näissä eläimissä munassa on vähän keltuaista ja pirstoutuminen on täydellistä. matelijoiden ja lintujen alkiot. Tämä vahvistaa ajatuksen nisäkkäiden alkuperästä esi-isien ryhmästä, jonka munat olivat runsaasti keltuaista.
7.4.2.3. Gastrulaatiovaiheen ominaisuudet
Gastrulaatiolle on ominaista erilaiset soluprosessit. Mitoottinen jatkuu solujen lisääntyminen, lisäksi sillä on erilainen voimakkuus alkion eri osissa. Mitoottisten syklien välivaiheet sisältävät kaikki jaksot (G 1 , S, G 2), siksi gastrulaatiovaiheesta alkaen havaitaan kehittyvän organismin kasvua. Kuitenkin eniten ominaisuus gastrulaatio koostuu solumassojen liikkuminen. Tämä johtaa muutokseen alkion rakenteessa ja sen muuttumiseen blastulasta gastrulaksi. menossa lajittelu soluja kuulumalla eri itukerroksiin, joiden sisällä ne "tunnistavat" toisensa.
Gastrulaatiovaihe alkaa erilaistuminen soluja, mikä tarkoittaa siirtymistä oman genomin biologisen tiedon aktiiviseen käyttöön. Yksi geneettisen aktiivisuuden säätelijöistä on alkiosolujen sytoplasman erilainen kemiallinen koostumus, joka muodostuu ovo(oo)plasmisen segregaation seurauksena, molemmat ovat peräisin munan vegetatiivisesta napasta.
Gastrulaation aikana rooli on erittäin suuri alkion induktio- kehittyvän alkion solukompleksien (osien) väliset vuorovaikutukset. On osoitettu, että primaarisen juovan ilmaantuminen linnuissa johtuu hypoblastin ja epiblastin välisestä induktiivisesta vuorovaikutuksesta. Hypoblastilla on polariteetti. Hypoblastin sijainnin muutos suhteessa epiblastiin aiheuttaa muutoksen primitiivisen juovan orientaatiossa. Kaikki nämä prosessit kuvataan yksityiskohtaisesti luvussa 8.
7.4.3. ELIMIEN JA KUDOSTEN MUODOSTUMINEN 7.4.3.1. Organogeneesin vaiheen olemus
organogeneesi, Yksittäisten elinten muodostuminen muodostavat alkiokauden pääsisällön. Ne jatkuvat toukissa ja päättyvät nuoruuteen. Organogeneesi erottuu monimutkaisimmista ja monipuolisimmista morfogeneettisistä (muotoilu) muunnoksista. Välttämätön edellytys siirtymiselle organogeneesiin on se, että alkio saavuttaa gastrulavaiheen, nimittäin itukerrosten muodostumisen. Ollessaan tietyssä asemassa toisiinsa nähden, itukerrokset muodostavat kosketuksessa ja vuorovaikutuksessa sellaisia suhteita eri soluryhmien välille, jotka stimuloivat niiden kehitystä tiettyyn suuntaan. Tämä ns alkion induktio- tärkein seuraus alkiokerrosten välisestä vuorovaikutuksesta.
Organogeneesin aikana solujen muoto, rakenne ja kemiallinen koostumus muuttuvat, soluryhmiä eristetään, jotka ovat tulevien elinten alkeita. Tietty elinten muoto kehittyy vähitellen, niiden välille muodostuu avaruudellisia ja toiminnallisia yhteyksiä. Morfogeneesiprosesseihin liittyy erilaistuminen
solujen erilaistuminen ja kudosten muodostuminen, kudosten ja rakenteiden erilaistuminen sekä niiden selektiivinen ja epätasainen kasvu. Organogeneesin edellytys solujen lisääntymisen, kulkeutumisen ja lajittelun ohella on niiden valikoiva kuolema (katso luku 8).
7.4.3.2. Neurulaatio
Organogeneesin alkua kutsutaan neurulaatioksi. Neurulaatio kattaa prosessit hermolevyn muodostumisen ensimmäisten merkkien ilmaantumisesta sen sulkeutumiseen hermoputkessa (kuva 7.10). Samaan aikaan, sointu Ja sekundaarinen suolisto, ja jänteen sivuilla makaava mesoderma halkeaa kraniokaudaalisessa suunnassa segmentoiduiksi parirakenteisiksi - somitit(muodostuu aksiaalisten elinten kompleksi - primaarinen organogeneesi).
Selkärankaisten, myös ihmisten, hermostolle on ominaista päärakennesuunnitelman vakaus kaikkialla evoluution historiaa alatyyppi. Hermoputken muodostumisessa kaikilla chordaateilla on paljon yhteistä. Aluksi erikoistumaton dorsaalinen ektoderma, vasteena chordomesoderman induktiovaikutukseen, se muuttuu hermolevy, lähetetty neuroepiteeli sylinterimäisiä soluja.
Hermolevy ei pysy litistyneenä pitkään. Pian sen sivureunat nousevat ja muodostuvat hermopoimut, jotka sijaitsevat matalan pituussuuntaisen molemmin puolin hermoura. Sitten hermopoimujen reunat sulkeutuvat muodostaen suljetun hermoputken, jonka sisällä on kanava - neurocele. Ensinnäkin hermopoimujen sulkeutuminen tapahtuu selkäytimen alun (selkäytimen ja aivojen välisen rajan) tasolla ja leviää sitten aivoihin ja häntään.
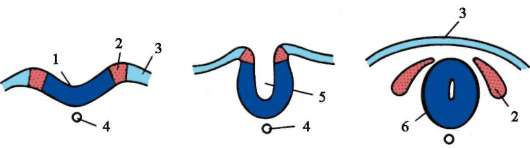
Riisi. 7.10. Peräkkäiset vaiheet hermoputken ja hermoharjan muodostumisessa (alkion poikkileikkaus): 1 - hermolevy; 2 - hermoharja; 3 - ektodermi; 4 - sointu; 5 - hermostunut ura; 6 - neurocoel; 7 - hermolaskokset
ohjeita. On osoitettu, että neuroepiteelisolujen mikrotubuleilla ja mikrofilamenteilla on tärkeä rooli hermoputken morfogeneesissä. Näiden solurakenteiden tuhoaminen kolkisiinilla ja sytokalasiinilla SISÄÄN saa hermolevyn pysymään auki. Hermopoimujen sulkeutumattomuus johtaa hermoputken synnynnäisiin epämuodostumisiin.
Hermopoimujen sulkeutumisen jälkeen muodostuvat solut, jotka alun perin sijaitsivat hermolevyn ja tulevan ihon ektodermin välissä hermoharja. Hermosolut eroavat kyvystään kulkea laajasti, mutta tiukasti säädellysti läpi kehon (katso kohta 8.2.2, kuva 8.9), ja ne muodostavat kaksi päävirtaa. Yhden heistä solut - pinnallinen- sisältyvät ihon orvasketeen tai dermiin, missä ne erilaistuvat pigmenttisoluiksi. Toinen virta kulkee vatsan suuntaan ja muodostaa herkkiä selkäydinhermosolmuja, sympaattisia hermosolmuja, lisämunuaisytimen, parasympaattisia hermosolmuja. Kallon hermonharjan solut synnyttävät sekä hermosoluja että useita muita rakenteita, kuten kidusrustoa, joitain kallon sisäluita.
Endodermi kaikissa alkioissa muodostaa lopulta sekundaarisen suoliston epiteelin ja monet sen johdannaiset. Itse toissijainen suolisto sijaitsee aina jänteen alla.
Siten neurulaation prosessissa kompleksi aksiaaliset elimet: hermoputki, sointu, suolet, jotka ovat kaikkien sointujen kehon organisoinnin tyypillisin piirre. Aksiaalisten elinten sama alkuperä, kehitys ja keskinäinen järjestely paljastaa niiden täydellisen homologian ja evolutionaarisen jatkuvuuden.
Neurulaatioprosessien syvällinen tarkastelu ja vertailu tietyissä sointutyypin edustajissa paljastaa joitakin eroja, jotka liittyvät pääasiassa ominaisuuksiin, jotka riippuvat munien rakenteesta, murskausmenetelmästä ja gastrulaatiosta (kuva 7.11). Huomio kiinnitetään alkioiden erilaiseen muotoon ja aksiaalisten elinten munimisajan siirtymiseen suhteessa toisiinsa, ts. edellä kuvattu heterokronia.
7.4.3.3. mesodermin erilaistuminen
Mesodermi, joka sijaitsee notochordin sivuilla ja ulottuu pidemmälle ihoektodermin ja sekundäärisen suolen endodermin välillä, on jaettu selkä- ja ventraalisiin alueisiin. Selkä
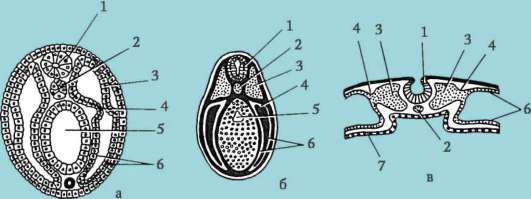
Riisi. 7.11 Erilaisten sointujen hermot: a - lansetti; b - sammakko; c - kana; 1 - hermoputki; 2 - sointu; 3 - somiitti; 4 - somiittijalka; 5 - sekundaarinen suoli; 6 - sivulevy; 7 - endodermi
osa on segmentoitu ja esitetty pareittain somitit. Somiittien muniminen tapahtuu päästä hännänpäähän. Mesoderman ventraalista segmentoimatonta osaa, joka näyttää ohuelta solukerrokselta, kutsutaan ns. sivulautanen. Somiitit on yhdistetty sivulevyyn segmentoidun välimesodermin avulla somiten jalat.
Mesodermin kaikki alueet eroavat vähitellen. Muodostumisen alussa somiiteilla on konfiguraatio, joka on ominaista epiteelille, jonka sisällä on onkalo. Painosta ja hermoputkesta tulevan induktiovaikutuksen alaisena somiittien ventromediaaliset osat - sklerotoomit- muuttuvat sekundaariseksi mesenkyymiksi, häädetään somiitista ja ympäröivät notokordin ja hermoputken vatsaosan. Lopulta niistä muodostuu nikamia, kylkiluita ja lapaluita.
Somiittien dorsolateraalinen osa erottuu kahteen suuntaan. Somiittien alueet, jotka ovat ektodermimuotoa päin dermatomit, jotka synnyttävät ihon sisäkerroksen - dermiksen. Soluja, jotka sijaitsevat somiittien dorsolateraalisen osan sisäpuolella, muodostuvat myotoomit, josta kehittyvät vartalon ja raajojen poikkijuovaiset luurankolihakset. Alkeellisilla somiittien jalkojen alueelta nefrotoomi Ja gonotome muodostuu erityselimiä ja sukupuolirauhasia.
Segmentoimattomat sivulevyt jakautuvat kahteen lehteen rajoittaen toissijaista kehon onteloa - yleisesti. sisälevy, endodermin vieressä olevaa kutsutaan viskeraaliksi. Se ympäröi suolistoa kaikilta puolilta ja muodostaa suoliliepeen, peittää keuhkojen
parenchyma ja sydänlihas. Ulompi lehti sivulevy on ektodermin vieressä ja sitä kutsutaan parietaalinen. Tulevaisuudessa se muodostaa vatsakalvon, keuhkopussin ja sydänpussin uloimmat levyt.
7.4.3.4. Alkiokerrosten johdannaiset
Ektodermi, mesodermi ja endodermi jatkokehityksen aikana, vuorovaikutuksessa keskenään, osallistuvat tiettyjen elinten muodostumiseen (yksityinen tai paikallinen organogeneesi). Elimen alkukennon ilmaantuminen liittyy paikallisiin muutoksiin vastaavan itukerroksen tietyllä alueella. Niin, ektodermista ihon orvaskesi ja sen johdannaiset (höyhenet, hiukset, kynnet, iho ja maitorauhaset), näkö-, kuulo-, haju-, suuontelon epiteeli ja hammaskiille kehittyvät. Tärkeimmät ektodermaaliset johdannaiset ovat hermoputki, hermoharja ja kaikki niistä muodostuneet hermorakenteet.
Endodermijohdannaiset ovat mahalaukun ja suoliston epiteeli, maksasolut, haiman erittävät solut, suolisto- ja maharauhaset. Alkion suolen etuosan endodermi muodostaa keuhkojen ja hengitysteiden epiteelin sekä aivolisäkkeen, kilpirauhasen ja lisäkilpirauhasen etu- ja keskilohkon erittävät solut.
mesoderma, edellä jo kuvattujen luustorakenteiden, luurankolihasten, ihon dermiksen, eritys- ja lisääntymisjärjestelmän elinten lisäksi se muodostaa sydän- ja verisuonijärjestelmän, imukudoksen, keuhkopussin, vatsakalvon ja sydänpussin. From mesenkyymi, kolmen itukerroksen soluista johtuen sekaperäinen, kehittyy kaikentyyppisiä sidekudoksia, sileitä lihaksia, veri ja imusolmuke (katso lisätietoja kappaleesta 8.2.5).
Tietyn elimen alkuaine muodostuu aluksi tietystä itukerroksesta, mutta sitten elin monimutkaistuu ja sen muodostumiseen osallistuu sen seurauksena kaksi tai kolme itukerrosta.
7.4.4. Selkärankaisten alkioiden väliaikaiset elimet
väliaikainen, tai väliaikainen, elimiä muodostuu useiden selkärankaisten edustajien alkiossa varmistamaan alkion elintärkeät toiminnot, kuten hengitys, ravinto, erittyminen, liike ja muut. Kehittyvimmän eläimen alikehittyneet elimet eivät vielä pysty toimimaan tarkoitetulla tavalla, vaikka niiden on pakko
niillä on jokin rooli kehittyvän koko organismin järjestelmässä. Heti kun alkio saavuttaa tarvittavan kypsyysasteen, jolloin useimmat elimet pystyvät suorittamaan elintärkeitä toimintoja, väliaikaiset elimet imeytyvät tai heitetään pois.
Väliaikaisten elinten muodostumisaika riippuu siitä, mitä ravintoainevarastoja munaan on kertynyt ja millaisissa ympäristöolosuhteissa alkio kehittyy. Esimerkiksi hännänttömässä sammakkoeläimissä munan riittävästä keltuaisen määrästä ja vedessä tapahtuvasta kehityksestä johtuen alkio suorittaa kaasunvaihdon ja vapauttaa dissimilaatiotuotteita suoraan munakalvojen läpi ja saavuttaa nuijapäävaiheen. Tässä vaiheessa muodostuu tilapäiset hengityselimet (kidukset), ruoansulatus ja liike, jotka on mukautettu vesielämään. Luetellut toukkaelimet antavat nuijapäisen jatkaa kehitystään. Kun aikuistyypin elinten morfologinen ja toiminnallinen kypsyys on saavutettu, väliaikaiset elimet katoavat muodonmuutosprosessissa.
Matelijoilla ja linnuilla on enemmän keltuaisvarantoja munassa, mutta alkio ei kehity vedessä, vaan maalla. Tässä suhteessa on erittäin varhaisessa vaiheessa varmistettava hengitys ja erittyminen sekä suoja kuivumiselta. Niissä alkaa jo varhaisessa alkionmuodostuksessa, lähes rinnakkain hermoston kanssa, väliaikaisten elinten muodostuminen, kuten esim. amnion, chorion Ja keltuainen pussi. Hieman myöhemmin muodostuu allantois. Istukan nisäkkäillä nämä samat väliaikaiset elimet muodostuvat vielä aikaisemmin, koska munassa on hyvin vähän keltuaista. Tällaisten eläinten kehitys tapahtuu kohdussa, tilapäisten elinten muodostuminen niissä osuu ajallisesti gastrulaation jakson kanssa.
Amnionin ja muiden väliaikaisten elinten läsnäolo tai puuttuminen on taustalla selkärankaisten jakamiselle kahteen ryhmään: Amniota Ja Anamnia(katso taulukko 7.1). Evoluutioteorian mukaan vanhemmat selkärankaiset, jotka kehittyvät yksinomaan vesiympäristössä ja joita edustavat sellaiset luokat, kuten syklostomit, kalat ja sammakkoeläimet, eivät tarvitse lisää vettä ja muita alkion kuoria ja muodostavat anamniumryhmän. Lapsieläinten ryhmään kuuluvat primaariset maaselkärankaiset eli ne, joiden alkion kehitys tapahtuu maanpäällisissä olosuhteissa. Nämä ovat kolme luokkaa: matelijat, linnut ja nisäkkäät. Ne kuuluvat korkeampiin selkärankaisiin, koska niillä on koordinoidut ja erittäin tehokkaat elinjärjestelmät, jotka varmistavat niiden olemassaolon vaikeita olosuhteita mitkä ovat maan olosuhteet. Nämä
luokkiin kuuluu suuri määrä lajeja, jotka ovat siirtyneet toissijaisesti vesiympäristöön. Siten korkeammat selkärankaiset pystyivät hallitsemaan kaikki elinympäristöt. Tällainen täydellisyys olisi mahdotonta, myös ilman sisäistä hedelmöitystä ja erityisten väliaikaisten alkioelinten muodostumista, joita kutsutaan myös nimellä alkion kalvot. Lapsivesikalvot sisältävät amnionin, chorionin (serosa), keltuaisen pussin ja allantois-kalvon.
Eri lapsivesien väliaikaisten elinten rakenteessa ja toiminnoissa on paljon yhteistä. Kuvattaessa yleisimmin korkeampien selkärankaisten alkioiden väliaikaisia elimiä, on huomattava, että ne kaikki kehittyvät jo muodostuneiden itukerrosten solumateriaalista. Istukan nisäkkäiden alkiokalvojen kehityksessä on joitain piirteitä, joita käsitellään jäljempänä.
Amnion on pussi, joka sisältää alkion ja on täytetty lapsivedellä. Lapsikalvon muodostavat alkion ulkopuolinen ektoderma ja somatopleura. Sen ektodermaalinen osa on erikoistunut alkion kylpevän lapsivesien eritykseen ja imeytymiseen. Amnionilla on ensisijainen rooli alkion suojelemisessa kuivumiselta ja mekaanisilta vaurioilta luoden sille suotuisimman ja luonnollisimman vesiympäristön. Amnionin mesodermaalinen osa synnyttää sileitä lihaskuituja. Näiden lihasten supistukset saavat amnionin sykkimään, ja tässä prosessissa alkiolle välitetyt hitaat värähtelevät liikkeet auttavat ilmeisesti varmistamaan, että sen kasvavat osat eivät häiritse toisiaan.
Chorioni (serosa)- uloin itukalvo kuoren (serosa) tai äidin kudosten (chorion) vieressä, joka syntyy, kuten amnion, ektodermista ja somatopleurasta. Tämä kuori palvelee vaihtoa alkion ja ympäristön välillä. Munakaslajeissa seroosin päätehtävä on osallistuminen hengitykseen (kaasunvaihto); nisäkkäillä korioni suorittaa paljon laajempia tehtäviä, osallistuen hengityksen lisäksi ravintoon, erittymiseen, suodatukseen ja aineiden, kuten hormonien, synteesiin.
Keltuainen pussi muodostuu alkion ulkopuolisesta endodermista ja viskeraalisesta mesodermista ja on suoraan yhteydessä alkion suolistoputkeen. Alkioissa, joissa on suuri määrä keltuaista, se osallistuu ravitsemukseen. Esimerkiksi linnuilla keltuaisen pussin splanchnopleuroniin kehittyy verisuonet. Keltuainen ei kulje keltuaiskanavan läpi, joka yhdistää pussin suoleen. Se käännetään ensin kielelle
liukoinen muoto pussin seinämän endodermaalisten solujen tuottamien ruoansulatusentsyymien vaikutuksesta. Sitten se tulee suoniin ja leviää veren kanssa koko alkion kehoon.
Nisäkkäillä ei ole keltuaisvarastoja, ja keltuaispussin säilyminen voi liittyä tärkeisiin toissijaisiin toimintoihin. Keltuaispussin mesodermi tuottaa alkion verisoluja. Lisäksi nisäkkään keltuainen pussi on täytetty nesteellä, jossa on korkea aminohappo- ja glukoosipitoisuus, mikä osoittaa proteiiniaineenvaihdunnan mahdollisuutta keltuaisessa pussissa.
Keltuaispussin kohtalo eri eläimissä on hieman erilainen. Linnuilla inkubaatioajan loppuun mennessä keltuaisen pussin jäännökset ovat jo alkion sisällä, minkä jälkeen se katoaa nopeasti ja häviää kokonaan 6 päivän kuluttua kuoriutumisesta. Nisäkkäillä keltuainen pussi kehittyy eri tavoin. Petoeläimissä se on suhteellisen suuri, ja siinä on pitkälle kehittynyt alusverkosto, kun taas kädellisillä se kutistuu nopeasti ja katoaa jälkiä ennen synnytystä.
Allantois kehittyy jonkin verran myöhemmin kuin muut väliaikaiselimet. Se on pussimainen uloskasvu takasuolen vatsan seinämästä. Siksi sen muodostaa sisäpuolinen endodermi ja ulkopuolella splanchnopleura. Matelijoilla ja linnuilla allantois kasvaa nopeasti korioniin ja suorittaa useita tehtäviä. Ensinnäkin se on urean ja virtsahapon säiliö, jotka ovat typpeä sisältävän aineenvaihdunnan lopputuotteita. eloperäinen aine. Allantoiksella on hyvin kehittynyt verisuoniverkosto, jonka ansiosta se osallistuu suonikalvon kanssa kaasunvaihtoon. Kuoriutuessaan allantoisin ulompi osa heitetään pois, kun taas sisäosa säilyy rakona.
Monilla nisäkkäillä allantois on myös hyvin kehittynyt ja muodostaa yhdessä suonikalvon kanssa korioallantoisen istukan. Termi istukka tarkoittaa itukalvojen läheistä päällekkäisyyttä tai fuusioitumista emoorganismin kudosten kanssa. Kädellisillä ja joillakin muilla nisäkkäillä allantoiksen endodermaalinen osa on alkeellista, ja mesodermaaliset solut muodostavat tiheän johdon, joka ulottuu kloaakin alueelta suonikalvoon. Suonet kasvavat allantois-mesodermia pitkin suonikalvoon, jonka kautta istukka suorittaa eritys-, hengitys- ja ravitsemustoimintoja.
Alkion kalvojen muodostumista ja rakennetta on helpompi tutkia kanan alkion esimerkin avulla. Neurulavaiheessa kolme alkiokerrosta siirtyy suoraan alkiosta ulkopuoliseen
hengitysosa, rajaamatta millään tavalla. Alkion muotoutuessa sen ympärille muodostuu useita poimuja, jotka ikään kuin leikkaavat alkion, erottavat sen keltuaisesta ja muodostavat selkeät rajat alkion ja alkion ulkopuolisten alueiden välille. Niitä kutsutaan rungon taitokset(Kuva 7.12).
Ensin muodostetaan pääpoimu. Hän leikkaa pään osan alhaalta. Tämän taitoksen takapäät kulkevat sivuttaisrungon taitteisiin, jotka rajaavat alkion rungon sivuilta. Häntäpoimu rajaa alkion takapäätä. Vähitellen kapenee jalka yhdistää keskisuolen ja keltuainen pussi, muodostunut
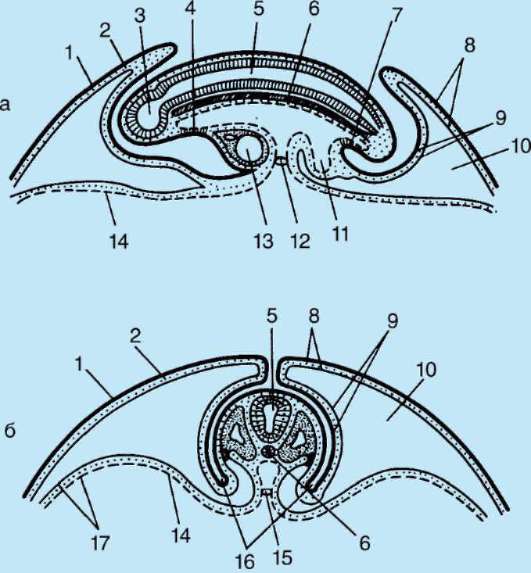
Riisi. 7.12 Runkolaskoksen ja alkion kalvojen muodostuminen kanan alkiossa: a - pituusleikkaus; b - poikkileikkaus; 1 - ektodermi; 2 - mesodermi; 3 - aivojen alkuosa; 4 - nielun kalvo; 5 - hermoputki; 6 - sointu; 7 - kloakaan kalvo; 8 - korioni; 9 - amnion; 10 - eksokoel; 11 - allantois; 12 - navan alue; 13 - sydämen alku; 14 - endodermi; 15 - suolen kirjanmerkki; 16 - vartalon taitokset; 17 - keltuainen pussi
etu- ja takasuolet. Samanaikaisesti ektodermista ja sen viereisestä somatopleurasta muodostuu ensin pääpoimu (kuva 7.13), joka hupun tavoin kasvaa alkion päälle ylhäältä. Päätaitteen päät muodostuvat sivuille lapsivesipoimut. Ne kasvavat alkion päällä toisiaan kohti ja kasvavat yhdessä muodostaen välittömästi alkion viereisen amnionin seinämät ja sen ulkopuolella olevan korionin.
Myöhemmin muodostuu allantois (kuva 7.14). Yleiskuva kanan alkiosta kuudentena inkubaatiopäivänä on esitetty kuvassa. 7.15. Eri nisäkkäillä väliaikaisten elinten muodostumisprosessit ovat enemmän tai vähemmän samanlaisia kuin edellä kuvatut linnuissa. Niiden kehityksen piirteet kädellisissä, myös ihmisissä, esitetään seuraavassa luvussa.
Riisi. 7.13.Kanan alkio noin 40 tunnin inkubaatioon: 1 - amnion pääpoimu; 2 - hermoputki; 3 - somitit
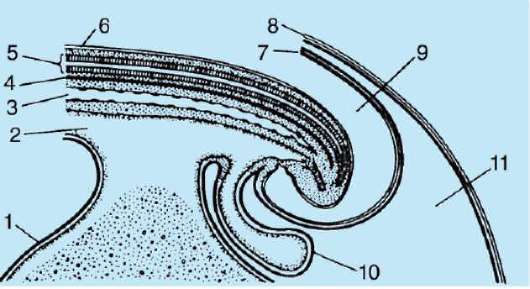
Riisi. 7.14.Allantoiksen muodostuminen kanan alkiossa (hännän pituusleikkaus): 1 - keltuainen pussi; 2 - keskisuoli; 3 - aortta; 4 - sointu; 3 - hermoputki; 6 - ektodermi; 7 - amnion; 8 - korioni; 9 - amnionontelo; 10 - allantois; 11 - eksokoelomi
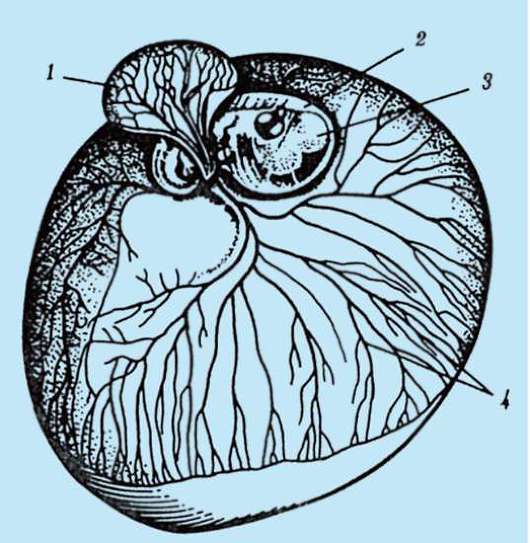
Riisi. 7.15. Kanan alkio kuudentena inkubaatiopäivänä (proteiini ja korioni poistettu, allantois siirretty ylöspäin): 1 - allantois; 2 - amnion; 3 - alkio; 4 - keltuaisen pussin suonet
7.5 nisäkkäiden alkion kehitys
7.5.1. PERIODISTOIMINTA JA ALKIOJEN VARHAINEN KEHITYS
Prenataalisen ja erityisesti alkion (kohdunsisäisen) ihmisen kehityksen tutkiminen on erittäin tärkeää, koska se auttaa ymmärtämään paremmin elinten välistä suhdetta sekä synnynnäisten epämuodostumien mekanismeja. Eri nisäkäslajien alkionkehityksessä on yleiset piirteet, mutta niissä on myös eroja. Esimerkiksi kaikissa istukkaissa varhaisen alkion syntyprosessit eroavat merkittävästi muissa selkärankaisissa aiemmin kuvatuista. Samaan aikaan istukan joukossa on erityispiirteitä.
Erota Ihmisen tsygootille on tunnusomaista seuraavat ominaisuudet. Ensimmäisen jaon taso kulkee munan napojen läpi, eli se on muiden selkärankaisten tavoin meridiaani. Tässä tapauksessa yksi tuloksena olevista blastomeereistä on suurempi kuin toinen, mikä osoittaa epäsäännöllisyys murskaamalla. Kaksi ensimmäistä blastomeeria siirtyvät seuraavaan divisioonaan asynkronisesti. Vao kulkee pituuspiiriä pitkin ja on kohtisuorassa ensimmäiseen uraan nähden. Siten on olemassa kolmen blastomeerin vaihe. Pienemmän blastomeerin jakautumisen aikana muodostui pienempien blastomeerien pari käännöksiä 90° siten, että fissiovaon taso on kohtisuorassa kahteen ensimmäiseen nähden.
Riisi. 7.16.Kanin tsygootin katkaisun varhaiset vaiheet: I - ensimmäisen katkaisuvaon taso; IIa - toisen vaon taso, joka murskaa yhden kahdesta ensimmäisestä blastomeerista; IIb - kahdesta ensimmäisestä blastomeeristä toisen toisen katkaisuuran taso
uurteita. Samanlainen blastomeerien järjestely 4-soluvaiheessa on kuvattu hiirillä, kaniineilla, minkeillä ja apinoilla (kuva 7.16). Asynkronisesta pilkkoutumisesta johtuen voi olla vaiheita, joissa on pariton määrä blastomeereja - 3, 5, 7, 9, jotka eroavat koostaan kasvuvaiheen puuttumisen vuoksi. Siis ihmisen murskaamista täydellinen asynkroninen epäyhtenäinen.
Murskaamisen seurauksena muodostuu solujen kertymä - morula. Pinnallisesti sijaitsevat blastomeerit muodostavat solukerroksen, ja morulan sisällä sijaitsevat blastomeerit ryhmittyvät keskeiseen solusolmuun. Pian morulan sisään ilmestyy nestettä, muodostuu ontelo (blastocoel) ja alkio muuttuu blastokysti. Morulan muodostuminen tapahtuu 16 blastomeerin vaiheessa ja kavitaatio (ontelon muodostuminen) alkaen 32 blastomeerin vaiheesta.
SISÄÄN blastokysti erottaa solujen ulomman kerroksen (trofoblasti) ja sisäinen solumassa (ikyhmy tai embryoblasti). Nesteen avulla sisempi solumassa työnnetään sivuun blastokystan yhteen napoihin. Myöhemmin alkaen trofoblasti ulompi itukalvo kehittyy - korioni ja alkaen embryoblasti- itse alkio ja jotkin alkion ulkopuoliset elimet. On osoitettu, että varsinainen alkio kehittyy hyvin pienestä määrästä itukyhmyn soluja.
Murskausvaihe tapahtuu kiiltävän kuoren alla. Kuva 7.17 esittää ihmisalkion alkuvaiheita ja osoittaa, missä alkio sijaitsee äidin kehossa. Ihmisen tsygootin fragmentoituminen ja blastokystan ilmaantuminen on esitetty kaavamaisesti kuvassa. 7.18 ja 7.19.
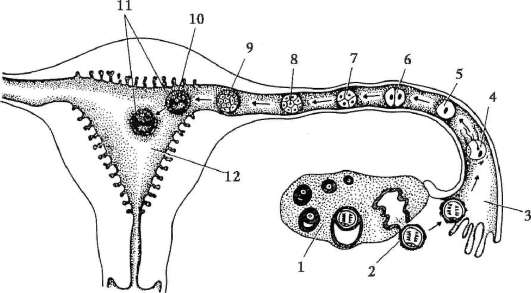
Riisi. 7.17.Ovulaatio, hedelmöitys ja ihmisalkio 1. kehitysviikolla: 1 - munasarja; 2 - munasolujen II kertaluokka (ovulaatio); 3 - munanjohdin; 4 - lannoitus; 5 - tsygootti; 6 - alkio kahden blastomeerin vaiheessa; 7 - alkio neljän blastomeerin vaiheessa; 8 - alkio kahdeksan blastomeerin vaiheessa; 9 - morula; 10, 11 - blastokysti; 12 - kohdun takaseinä
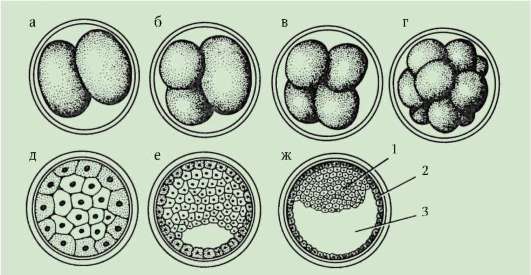
Riisi. 7.18.Ihmisen tsygootin pilkkoutuminen: a - kaksi blastomeeriä; b - kolme blastomeeria; c - neljä blastomeeria; g - morula; e - morulan osa; f, g - varhaisen ja myöhäisen blastokystan osa: 1 - embryoblasti; 2 - trofoblasti; 3 - blastocoel
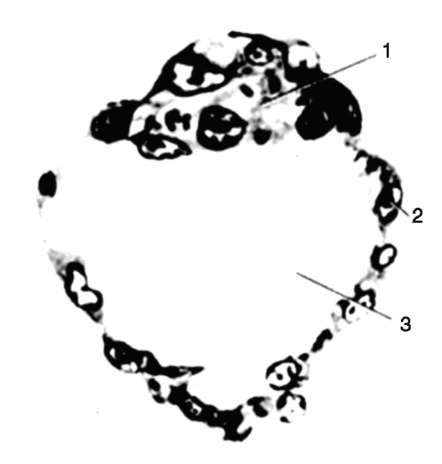
Riisi. 7.19.Ihmisalkion blastokysta (osio): 1 - embryoblasti; 2 - trofoblasti; 3 - blastocoel
Noin 6-7 päivänä hedelmöityksen jälkeen 2-3 vuorokautta kohtuontelossa vapaasti kellunut alkio on valmis istutettavaksi, ts. upottamalla sen limakalvoon. Trofoblastin erittämien entsyymien vaikutuksesta zona pellucida tuhoutuu osittain ja alkio tulee ulos siitä ("kuoriutuu"). Sekä kohdun epiteelin että trofoblastisolujen syntetisoimien integriiniproteiinien osallistuessa blastula kiinnittyy kohdun seinämään alkion napallaan. Kun trofoblastisolut ovat joutuneet kosketuksiin äidin kudosten kanssa, ne lisääntyvät nopeasti ja vapauttavat proteolyyttisiä entsyymejä tuhoavat ne (nämä kudokset). Tämä aiheuttaa kohdun seinämän reaktion, johon liittyy aktiivinen verisuonten muodostuminen istutuskohdassa. Implantaatioon tarvittavien integriinien ja entsyymien synteesin rikkominen tekee tämän prosessin mahdottomaksi, mikä puolestaan johtaa hypoksiaan ja alkion kuolemaan. Implantaation aikana trofoblasti erottuu kahteen kerrokseen: sisäkerrokseen, ns sytotrofoblastooma, koska se säästää solurakenne, ja ulompi nimeltään syncytio-trofoblastooma, koska se on syncytium. Kuva 7.20 esittää ihmisalkion implantaatioprosessissa.
gastrulaatio nisäkkäillä se on läheistä sukua muille alkion transformaatioille. Samalla kun trofoblasti jakautuu kahteen kerrokseen, itukyhmy litistyy ja muuttuu kaksikerroksiseksi sukusolujen suoja. Suojan alin kerros - hypoblasti, tai primaarinen endodermi, useimpien kirjoittajien mukaan, muodostuu sisäisen solumassan delaminaatiosta, suunnilleen samalla tavalla kuin se tapahtuu lintujen alkiolevyssä. Ensisijainen
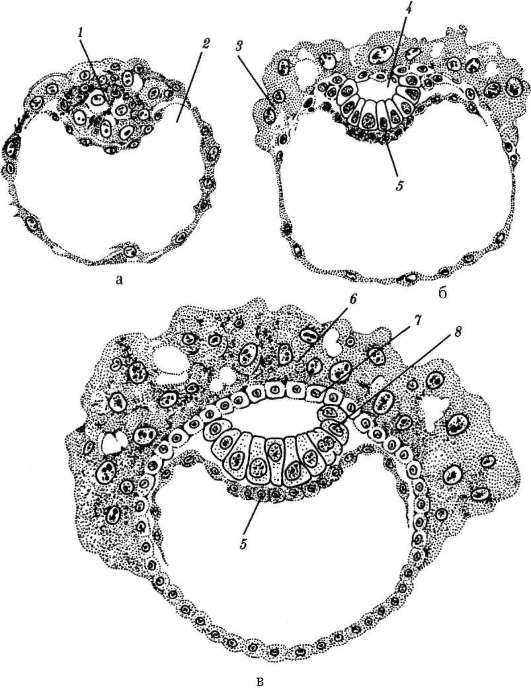
Riisi. 7.20.Ihmisalkion istutuksen ja kehityksen peräkkäiset vaiheet 1. ja 2. viikon lopussa: a - blastokysta; b - blastokysta implantaation alussa (7. kehityspäivä); c - osittain istutettu blastokysta (8. kehityspäivä)
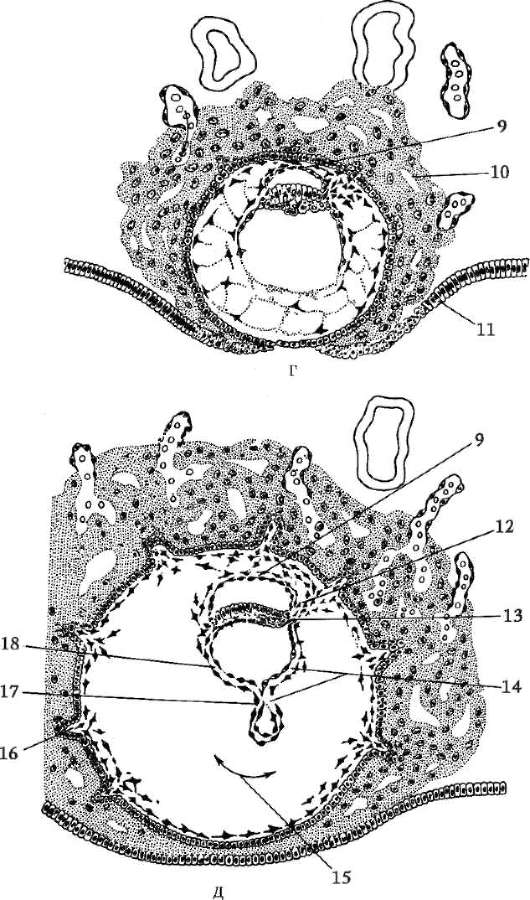
Riisi. 7.20. Loppu: d - alkio 9-10 kehityspäivänä; e - alkio 13. kehityspäivänä: 1 - embryoblasti, 2 - blastocoel, 3 - trofoblasti, 4 - amnionontelo, 5 - hypoblast, 6 - syncytiotrophoblast, 7 - cytotrophoblast, 8 - epiblast, 9 - amnion, 10 - lacuna trofoblasti, 11 - kohdun epiteeli, 12 - vartalon varsi, 13 - allantoismunuainen, 14 - keltuainen, 15 - ylihengittävä coelom, 16 - korionivillus, 17 - primaarinen keltuaispussi, 18 - sekundaarinen keltuaispussi
naya endodermi kuluu kokonaan alkionulkoisen endodermin muodostumiseen. Trofoblastin ontelon vuorauksena se muodostaa nisäkkäiden ensisijaisen keltuaispussin.
Ylempi solukerros epiblasti- tulevan ektodermin, mesodermin ja sekundaarisen endodermin lähde. Kolmannella viikolla epiblasti muodostuu päälinja, jonka kehittymiseen liittyy lähes samat solumassojen liikkeet kuin lintujen primaarijuovan muodostumisessa (kuva 7.21). Ensisijaisen sarjan kärjessä, Hensenin solmu Ja ensisijainen reikä, homologinen muiden selkärankaisten dorsaalisen blastohuulen kanssa. Primaarisen kuopan alueella liikkuvat solut ohjataan epiblastin alle kohti esihordaalilevyä.
esihordaalinen levy sijaitsee alkion päähän ja merkitsee tulevan orofaryngeaalisen kalvon paikkaa. Keskiakselia pitkin liikkuvat solut muodostavat notokordin ja mesoderman alkion ja muodostavat chordomesoderm-prosessi. Hensen-solmu siirtyy vähitellen kohti alkion kaudaalista päätä, ensisijainen juova lyhenee ja notochord rudimentti pitenee. Kordomesodermaalisen prosessin sivuille muodostuu mesodermaalisia levyjä, jotka laajenevat molempiin suuntiin. Kuva 7.22 esittää yleisen kaavion joistakin varhaisen alkion kehityksen prosesseista.
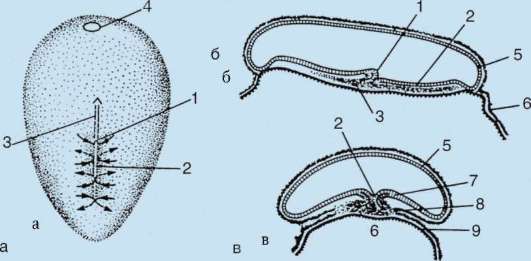
Riisi. 7.21.Ihmisalkion kehitys primaarisessa juovavaiheessa (15-17 päivää): a - näkymä alkiosta ylhäältä (amnion poistetaan); b - pituusleikkaus; c - poikittaisleikkaus ensisijaisen nauhan läpi: 1 - Hensenin solmu; 2 - ensisijainen nauha; 3 - sointu; 4 - esihordaalilevy; 5 - amnion; 6 - keltuainen pussi; 7 - ektodermi; 8 - mesodermi; 9 - endodermi
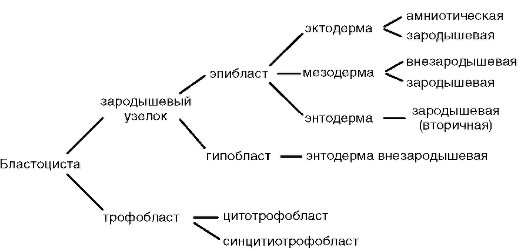
Riisi. 7.22. Nisäkkäiden itukerrosten erilaistuminen
Kolmannen viikon loppuun mennessä alkion ektodermissa notokordin alkupään yläpuolella, hermolevy. Se koostuu korkeista sylinterimäisistä soluista. Hermolevyn keskelle muodostuu muotoon taipuma hermokouru, ja sen sivuilla nousee hermopoimuja. Tämä on hermoston alku. Alkion keskiosassa hermolaskokset sulkeutuvat - a hermostoputki. Sitten sulku leviää pään ja hännän suuntaan. Hermoputki ja viereiset ektodermin osat, joista se myöhemmin kehittyy hermoharja, kokonaan upotettuna ja erotettuna niiden yläpuolella yhdessä kasvavasta ektodermista (ks. kuva 7.10). Hermoputken alla oleva solukaistale muuttuu sointeeksi. Selän mesodermin segmentit - somiitit - näkyvät sikiön keskiosassa olevan notochordin ja hermoputken sivuilla. Neljännen viikon loppuun mennessä ne leviävät pään ja hännän päihin, niiden lukumäärä on noin 40 paria.
Sekundaarisen suolen muodostumisen alku, sydämen vaurio ja keltuaispussin verisuonisto kuuluvat samaan aikaan. Kuvassa 7.23 on esitetty alkion ja alkion ulkopuolisten elinten koon suhde 21. kehityspäivänä. Yksityiskohtaisemmin alkion rungon irtoaminen alkion kalvoista ja elinten asettaminen voidaan nähdä kuvassa. 7.24, joka näyttää paitsi yleiskuvan alkiosta myös leikkaussuunnitelmat. Huomiota kiinnitetään nopeaan (7 päivän kuluessa 4. viikosta) alkion muodostumiseen pitkänomaisen ja kaarevan rungon muodossa, joka on kohotettu ja leikattu pois keltainen pussin runkopoimuista. Takana
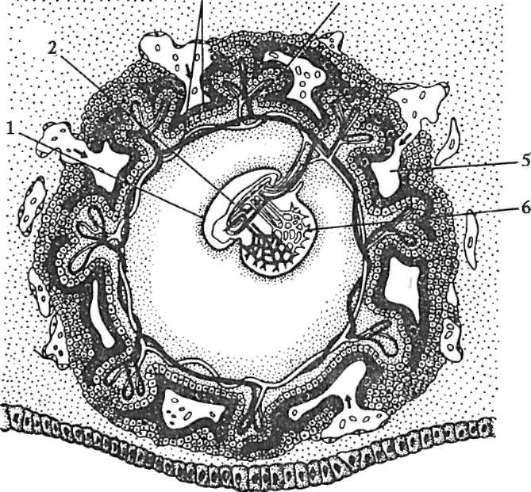
Riisi. 7.23.Ihmisen alkio ja alkion ulkopuoliset elimet 21. kehityspäivänä: 1 - amnion; 2 - alkio; 3 - korioni; 4 - tertiäärinen villus; 5 - äidin veri; 6 - keltuainen pussi
tällä hetkellä asetetaan kaikki somiitit, neljä paria kiduskaavia, sydänputki, raajojen munuaiset, keskisuoli sekä etu- ja takasuolen "taskut".
Seuraavan neljän alkionkehityksen viikon aikana kaikki tärkeimmät elimet asetetaan. Kehitysprosessin rikkominen tänä aikana johtaa vakavimpiin ja moninkertaisimpiin synnynnäisiin epämuodostumisiin.
Kuten edellä todettiin, alkion ulkopuolisten väliaikaisten elinten kehittymisellä nisäkkäillä ja ihmisillä on erityispiirteitä. Nämä elimet muodostuvat hyvin varhain, samanaikaisesti gastrulaation kanssa ja hieman eri tavalla kuin muissa amnioteissa. Korionin ja amnionin kehityksen alku osuu 7-8 päivään, ts. samaan aikaan implantaation alkamisen kanssa. Choorion syntyy trofoblastista, joka on jo jakautunut sytotrofoblasteihin ja synsytiotrofoblasteihin. Jälkimmäinen kasvaa ja tuhoaa sen joutuessaan kosketuksiin kohdun limakalvon kanssa. Toisen viikon loppuun mennessä
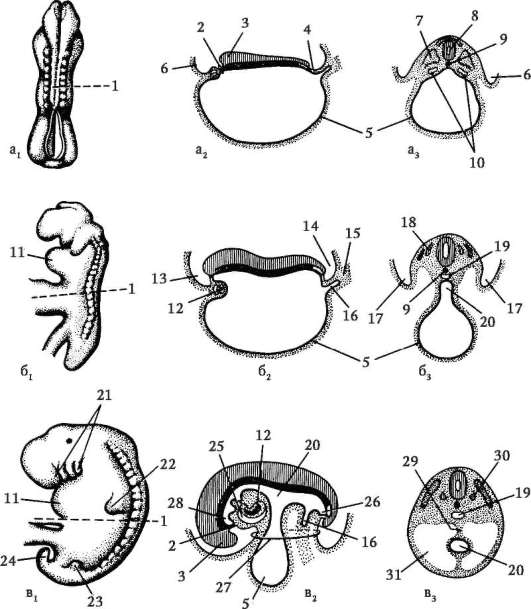
Riisi. 7.24.Ihmisalkion kehitys 4. viikolla: a 1 b 1 in 1 - yleiskuva; a 2 b 2 c 2 - pituusleikkaus; a 3 b 3 c 3 - poikkileikkaus; a 1 a 2 a 3 - 22 päivää; b 1 b 2 b 3 - 24 päivää; in 1 in 2 in 3 - 28 päivää: 1 - poikittaisleikkauksen taso; 2 - orofaryngeaalinen kalvo; 3 - aivot; 4 - kloakaan kalvo; 5 - keltuainen pussi; 6 - amnion; 7 - somitit; 8 - hermoputki; 9 - sointu; 10 - vatsa-aortan parilliset kirjanmerkit; 11 - sydämen reuna; 12 - sydän; 13 - pään rungon taitto; 14 - hännän rungon taitto; 15 - vartalon jalka; 16 - allantois; 17 - sivuttaisrungon taitokset; 18 - hermoharja; 19 - dorsaalinen aortta; 20 - keskisuoli; 21 - kiduskaaret; 22 - eturaajan munuainen; 23 - takaraajan munuainen; 24 - häntä; 25 - sydänpussi; 26 - takasuolen tasku; 27 - napanuora; 28 - etusuolen tasku; 29 - dorsaalinen suoliliepi; 30 - takajuuren hermosolmu; 31 - alkionsisäinen kokonaisuus
korionin primaariset villit muodostuvat sytotrofoblastiepiteelisolujen kerääntymisen muodossa. Kolmannen viikon alussa niihin kasvaa mesodermaalinen mesenkyymi ja ilmaantuu sekundaarisia villit, ja kun verisuonia ilmaantuu sidekudoksen ytimeen 3. viikon lopussa, niitä kutsutaan tertiaariseksi villuksi. Alue, jossa suonikalvon ja kohdun limakalvon kudokset ovat lähellä toisiaan, on ns. istukka.
Ihmisillä, kuten muillakin kädellisillä, istukan emän osan verisuonet menettävät jatkuvuutensa ja suonivilkut todella kylpevät äidin organismin veressä ja imusolmukkeessa. Tätä istukkaa kutsutaan hemochoriaal. Raskauden edetessä villit kasvavat, haarautuvat, mutta sikiön veri alusta loppuun asti eristetään äidin verestä istukkaesteen avulla.
Istukan este koostuu trofoblastista, sidekudoksesta ja sikiön verisuonten endoteelistä. Tämä este läpäisee vettä, elektrolyyttejä, ravinteita ja dissimilaatiotuotteita sekä sikiön punasoluantigeenejä ja äidin vasta-aineita, myrkyllisiä aineita ja hormoneja. Istukan solut tuottavat neljää hormonia, mukaan lukien ihmisen koriongonadotropiinia, jota löytyy raskaana olevan naisen virtsasta 2-3 raskausviikolla.
Amnion tapahtuu sisäisen solumassan epiblastisolujen hajoamisen seurauksena. Ihmisen amnionia kutsutaan skizamnion(katso kuva 7.20) toisin kuin pleuramnion linnut ja jotkut nisäkkäät. Lapsivesionteloa rajoittavat jonkin aikaa epiblastisolut ja osittain trofoblastikohta. Sitten epiblastin sivuseinämät muodostavat ylöspäin suunnattuja taitoksia, jotka myöhemmin sulautuvat yhteen. Ontelo on kokonaan vuorattu epiblastisilla (ektodermaalisilla) soluilla. Ulkopuolelta lapsivesien ektodermia ympäröivät alkion ulkopuoliset mesodermaaliset solut. Lapsivesi sisältää erilaisia aineita, kuten proteiineja, hiilihydraatteja, suoloja, hivenaineita, hormoneja jne. Sen koostumus muuttuu kehityksen aikana, lisäksi sitä päivitetään jatkuvasti. Alkion kehitysprosessissa sillä on tärkeä rooli nenä- ja suuonteloiden, ruoansulatus- ja hengityselinten muodostumisessa.
Keltuainen pussi ilmestyy, kun ohut kerros hypoblastia erottuu sisäisestä solumassasta ja sen alkion ulkopuoliset endodermaaliset solut, liikkuvat, vuoraavat tro-
foblast. Tuloksena oleva primaarinen keltuaispussi romahtaa 12-13 päivänä ja muuttuu toissijaiseksi keltuaispussiksi, joka liittyy alkioon keltuaiskanavan kautta. Endodermaaliset solut peitetään ulkopuolelta alkion ulkopuolisella mesodermilla. Kolmannella kehitysviikolla primaarisia sukusoluja, verisoluja ja verisuonia alkaa havaita keltuaisen pussin seinämässä. 7-8. kehitysviikkoon asti tämä keltuainen pussi on alkion tärkein hematopoieettinen elin.
Allantois syntyy ihmisalkiossa, kuten muissakin lapsisoluissa, taskun muodossa takasuolen vatsan seinämässä, mutta sen endodermaalinen ontelo pysyy alkeellisena rakenteena. Siitä huolimatta sen seiniin kehittyy runsas verisuoniverkosto, joka yhdistyy alkion tärkeimpiin verisuoniin. Allantois-mesodermi yhdistyy korionimesodermiin ja antaa siihen verisuonia, jotka tarjoavat ravintoa kehittyvälle alkiolle noin toiseen alkiokuukauteen asti. Siten tapahtuu korioallantoisen istukan vaskularisaatiota.
Taulukossa on esitetty joitakin ihmisalkioiden elinten kehitysvaiheita ja -ehtoja. 7.4
Taulukko 7.4. Tärkeimmät ajanjaksot ja tapahtumat ihmisen varhaisessa ontogeneesissä
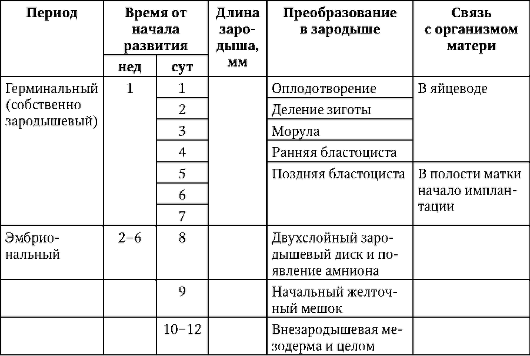
Taulukon jatko. 7.4
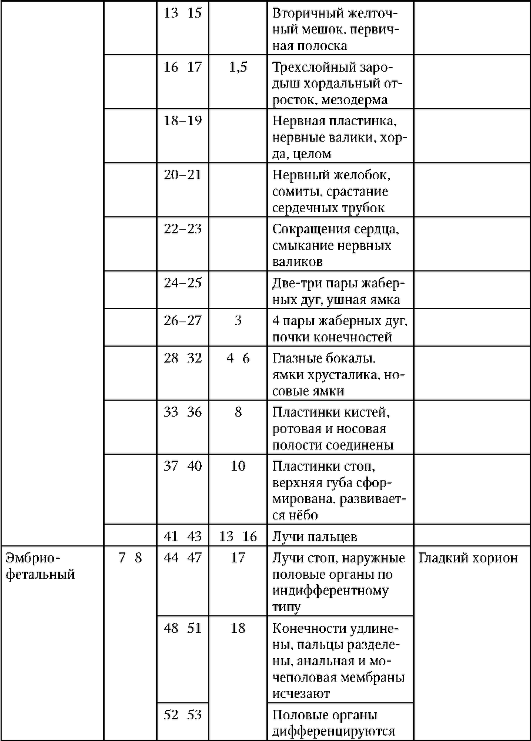
Pöydän loppu. 7.4
7.5.2. ESIMERKKEJÄ LAJIEN EVOLUUTIOTA heijastavasta IHMISELIMISTÄ
Tämän osion tarkoituksena ei ole kuvata kaikkien ihmisen elinten kehitystä. Vain joitain morfogeneettisiä prosesseja tarkastellaan havainnollistaen seuraavia yleisiä biologisia kohtia:
Solujen välisten, kudosten ja elinten välisten vuorovaikutusten arvo morfogeneesissä;
Selkärankaisten organisaation evoluutionaalisesti muinaisempien piirteiden heijastus ihmisen morfogeneesissä;
Organogeneesin piirteet, joiden avulla voidaan arvioida kehityshäiriöiden vaihetta ja mekanismeja tietyissä synnynnäisissä epämuodostumissa.
Segmentaalinen rakenne selkärangan heijastaa sen alkuperää alkion somiiteista. Lisäksi nikamien muodostuminen liittyy läheisesti notochordiin ja selkäytimeen. Selkärangat muodostuvat sklerotoomeista peräisin olevista mesenkymaalisista soluryhmistä, jotka kulkevat kohti notochordia ja ympäröivät sitä. Kahden vierekkäisen somiittiparin mesenkymaaliset solut ovat mukana nikamakappaleiden muodostumisessa, ja siksi nikamakappaleet eivät sijaitse somiittien tasolla, vaan niiden välissä. Samaan aikaan lihakset kehittyvät somi-
Riisi. 7.25. Selkänikamien selkäkaarien kehitys kanan alkiossa: a - normaali kehitys; b - selkärangan hermosolmujen poistamisen jälkeen: 1 - selkäkaari; 2 - selkärangan runko; 3 - selkärangan solmu; 4 - selkäydin; 5 - sointu; 6 - nikamien segmentoimattomat selkäkaaret
comov, ikään kuin heitettäisiin nikamien välisten nivelten yli ja varmistavat nikamien liikkeen.
Selkäkaaret muodostuvat dorsaaliseen (dorsaaliseen) suuntaan leviävistä mesenkymaalisten sklerotomiasolujen kerääntymistä, kun taas poikittaisprosessit ja kylkiluut muodostuvat sivuttain liikkuvien sklerotomiasolujen kerääntymistä. Selkäydin ja selkäydinhermosolmut osallistuvat kohdunsisäisen kehityksen asianmukaisissa vaiheissa nikamien selkäkaarien morfogeneesiin. Joten jos selkäydinhermosolmukkeet poistetaan, muodostuu selkäkaarien rusto, mutta se näyttää segmentoimattomalta sauvalta (kuva 7.25).
Selkänikamien histologinen rakenne muuttuu vähitellen. Viikon 4-5 aikana muodostuu mesenkymaalisia nikamia, 6. viikolla esiintyy kondrogeneesipesäkkeitä ja sitten 8. viikolla alkaa luutuminen, joka kestää lähes 25 vuotta. Selkärangan sisällä sijaitsevat jänteen osat katoavat vähitellen. Välilevyissä notochord säilyy liman kaltaisena rakenteena, joka tunnetaan ns. nucleus pulposus.
Muodostus keskikorva liittyy ensimmäisen ja toisen ontogeneettisiin transformaatioihin sisäelinten (kidusten) kaaret. Kuva 7.26 esittää kaavamaisesti pään ja kaulan alueita ihmisen kehityksen 4. ja 24. (sikiökehityksen ensimmäisen ja toisen kolmanneksen lopussa) viikoilla. Neljännellä viikolla viskeraaliset (kidus) kaaret asetetaan alkion pään ja kaulan alueelle oikealle ja vasemmalle. Sisäpuolelta, nielun sivulta, niiden väliin muodostuu nielutaskut(Kuva 7.27). Nämä ovat endodermin ulkonemia (tai ulkonemia),
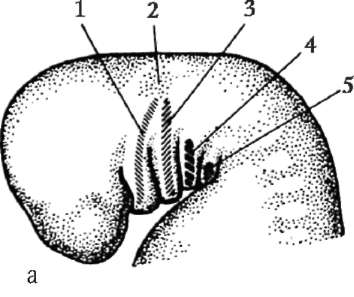
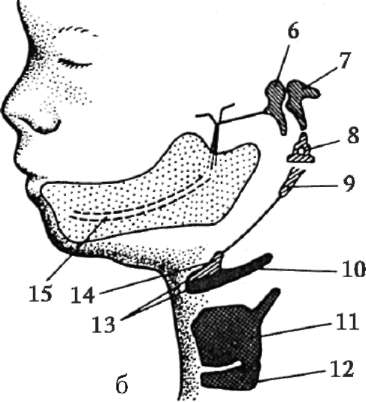
Riisi. 7.26.Ihmisalkion pää ja kaula, sivukuva: a - 4. viikolla; b - viikolla 24: 1 - ensimmäinen kaari; 2 - kehittyvän sisäkorvan sijainti; 3 - toinen kaari; 4 - kolmas kaari; 5 - neljäs niitty; 6 - vasara. 7 - alasin; 8 - jalustin; 9 - styloidiprosessi; 10 - hyoidiluun suuri sarvi; 11 - kilpirauhasen rusto; 12 - cricoid rusto; 13 - hyoidiluun runko; 14 - pieni sarvi; 15 - alaleuka; sisäelinten kaarien (a) ja niistä kehittyvien elinten (b) kalvot on merkitty samalla varjostuksella
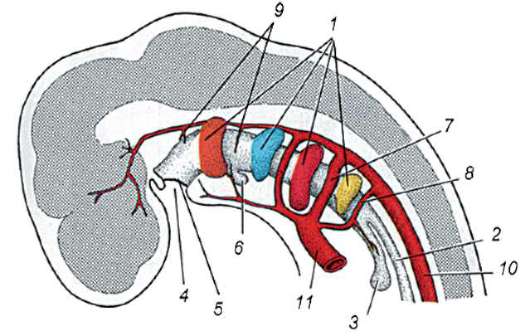
Riisi. 7.27.5 viikon ikäisen ihmisalkion kidusalue (integumentaalinen ektoderma ja mesenkyymi poistettu). 1 - nielun taskut; 2 - ruokatorvi; 3 - henkitorvi ja keuhkomunuaiset; 4 - stomodeum; 5 - stomodaalinen (suullinen) levy; 6 - kilpirauhasen alku; 7 - 4. aortan kaari; 8 - 6. aortan kaari; 9 - pienentämällä 1. ja 2. aortan kaaria; 10 - dorsaalinen laskeva aortta; 11 - vatsan nouseva aortta (valtimopussi)
kasvaa sivuttain. Ulkopuolella nielun kasvun mukaan ektodermaalinen taskut.
Ensimmäisen nielu- ja kidustaskun endodermi ja ektodermi joutuvat kosketuksiin 4. kehitysviikolla (Kuva 7.28, a). Kosketus ei kestä kauan, nielun taskun sokea pää siirtyy pois pinnasta ja sitä ympäröi mesenkyymi (kuva 7.28, b). Nielun taskun distaalisesta osasta alkaa kehittyä keskikorvan ontelo (tympanic cavity) ja kuuloputki (Eustachian) muodostuu proksimaalisesta osasta. Mesenkyymistä erottuvat edelleen ääntä johtavien kuuloluun renkaat (kuva 7.28, c). Malleus ja alasin ovat peräisin ensimmäisen viskeraalikaaren solumateriaalista, jalustin - toisen viskeraalikaaren selkäosan solumateriaalista. Lihakset ja hermot, jotka on liitetty ääntä johtavaan luun laitteistoon välikorvan ossikkelilla, muodostuvat ensimmäisen ja toisen viskeraalisen kaaren alueen solumateriaalista.
Kohdunsisäisen elämän lopussa ja muutaman kuukauden kuluessa syntymästä alkion sidekudos imeytyy täryonteloon. Vasta tämän jälkeen kuuloluut saavat liikkuvuutta ja tärykalvon värähtelyt voivat siirtyä vapaasti sisäkorvan soikean ikkunan kalvoon.
Rakenteiden muodostumisessa kasvojen alue Ja suuontelon ekto- ja mesodermisolut osallistuvat. Suurin osa rakenne
Riisi. 7.28. Välikorvan, etuosan kehitys: a - 4. viikolla; b - 5. viikolla; c - myöhemmät vaiheet; 1 - ensimmäinen kiduslevy; 2 - ensimmäinen tasku; 3 - ensimmäinen viskeraalinen (kiduskaari); 4 - kuulovesikkeli; 5 - ensimmäinen nielun tasku; 6 - ensimmäisen viskeraalisen kaaren johdannaiset; 7 - toisen viskeraalisen kaaren johdannainen; 8 - putken orjaontelo; 9 - ulkoisen kuulokäytävän tulppa. 10 - ensisijainen ulkoinen kuulokanava; 11 - vasara; 12 - alasin; 13 - jalustin; 14 - täryontelo; 15 - kuuloputki (Eustachian).
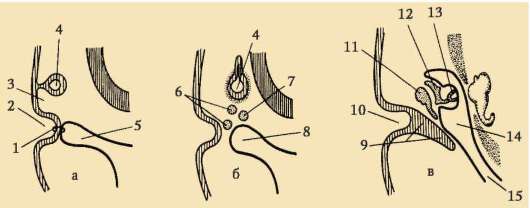
kasvojen alueen kiertue kehittyy ektodermaalista alkuperää olevien mesenkymaalisten solujen massoista, jotka siirtyvät hermonharjasta (4. itukerros). Tästä mesenkyymistä kehittyvät side-, rusto- ja luukudokset sekä hammasmassakudos. Kasvojen ja nielun lihasten poikkijuovaiset lihakset muodostuvat mesodermaalista alkuperää olevasta mesenkyymistä. Tässä on tärkeää ymmärtää, kuinka "toimintojen laajenemis- ja muutosprosessi" tapahtui antropogeneesissä.
Suuontelo muodostuu ektodermaalisesta invaginaatiosta stomo-deum, sokeasti päättyvän etusuolen vieressä. Ne erotetaan ohuista stomodaalinen levy, koostuu ektodermaalisista (stomodeumin puolelta) ja endodermaalisista (suolen puolelta) levyistä (katso kuva 7-26). Tämä levy rikkoutuu neljännen kehitysviikon lopussa ja suuontelo on yhdistetty alun perin sokeasti suljettuun suolistoputkeen.
Korisylkirauhaset kehittyvät stomodeumin ektodermista ja sublingvaaliset ja submandibulaariset sylkirauhaset nielun ventraalisen osan endodermista.
5 viikon ikäisellä sikiöllä suun avaaminen rajoittavat seuraavat rakenteet (kuva 7.29, a): keskiviivalla suuontelon yläpuolella on pyöristetty ulkoneva alue - edestä (etuosa) kieleke; sen sivuilla on hevosenkengän muotoisia kohoumia, jotka ympäröivät hajukuoppia. Näiden korkeuksien keskiprosessit
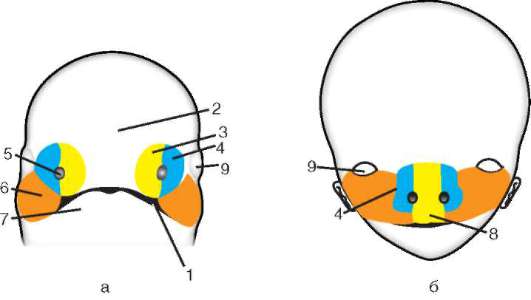
Riisi. 7.29. Kasvojen muodostumisen peräkkäiset vaiheet (näkymä edestä): a - 5. viikolla; b - 10. viikolla. 1 - stomodeum; 2 - etu (etu) reuna; 3 - mediaalinen nenäprosessi; 4 - lateraalinen nenäprosessi; 5 - hajukuoppa; 6 - yläleuan prosessi; 7 - alaleuan kaaren asettaminen; 8 - filtrum; 9 - silmä
nimeltään mediaalinen nenä, ja sivut lateraalinen
nenän prosessit. Suuontelon yläsivukulmista kohti keskiviivaa kasvaa yläleuan prosesseja.
Suunontelon rajoittaminen alhaalta alaleuka muodostettu alaleuan kaaren parilliset kirjanmerkit. Aluksi ne ilmestyvät keskilinjan molemmille puolille, sitten lisääntyvät toisiaan kohti ja kasvavat yhdessä.
Kuudennen viikon aikana kehitys on nopeaa yläleuka. Leuan prosessit kasvavat kohti keskiviivaa. Samaan aikaan nenäprosessit lisääntyvät työntäen niiden välissä sijaitsevan etuosan alaosaa ylös. Erityisen intensiivisesti kasvavat mediaaliset nenän prosessit, jotka siirtyvät kohti keskiviivaa ja sulautuvat toisiinsa ja sulautuvat sitten yläleuan prosesseihin muodostaen näin täydellisen yläleuan kaaren 10. kehitysviikolla. Muodostuu ihon ektodermi yhdistyneiden mediaalisten prosessien päälle filtrum- pituussuuntainen ura ylähuulen yläpuolella nenän alla (kuva 7.29, b).
Toisen kehityskuukauden lopussa, kun yläleuka muodostuu, se alkaa kehittyä kiinteä taivas. Sen muodostumiseen sekä yläleuan kaaren muodostumiseen osallistuvat keskimmäiset nenän ja yläleuan prosessit(Kuva 7.30, a). Yläleuan keskialueelta, joka syntyi keskimmäisten nenän prosessien fuusioitumisen seurauksena, muodostuu pieni kolmion muotoinen osa - ensisijainen kitalaki. Pääosa - toissijainen kitalaki- kehittyy lateraaliset palatiinin kasvut (prosessit), jotka näkyvät yläleuassa (kuva 7.30, b). Aluksi ne on suunnattu alaspäin, ja sitten palatinisten prosessejen reunat nousevat ylös ja kohti keskiviivaa. Ne koskettavat ja sulautuvat muodostaen kitalaen ompeleen. Muodostunut kitalaen pääosa erottaa alkuperäisen stomodaaliontelon ylimmän osan. Toissijainen kitalaki kasvaa yhdessä primaarisen kanssa, ja nenän väliseinä kasvaa niiden kallon pintaan (kuva 7.30, c). Näin ollen samanaikaisesti nenäontelon erottamisen kanssa suuontelosta se jaetaan oikeaan ja vasempaan puolikkaaseen.
Sen muodostumisen alkuvaiheessa (alkion 5. viikolla) Kieli on limakalvon muodostama pussi, joka on täynnä kasvavaa lihasmassaa. Uskotaan, että kielen limakalvo johtuu stomodeum ectoderm, ja lihakset tulevat takaraivo myotomit.
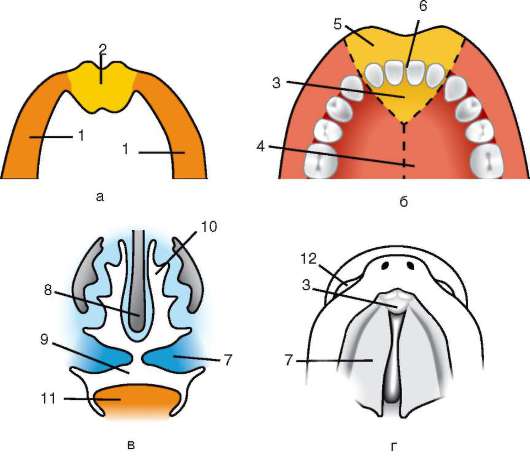
Riisi. 7.30. Kovan kitalaen muodostuminen: a - kaavamainen esitys yläleuasta (näkymä suuontelon sivulta); b - 10. kehitysviikko (näkymä suuontelosta); c - 8. kehitysviikko (etuosa); d - 8. kehitysviikko (näkymä suuontelosta): 1 - yläleuan prosessi; 2 - yhdistetyt mediaaliset nenäprosessit; 3 - ensisijainen taivas; 4 - toissijainen kitalaen (sivusuulake-outgrowths); 5 - huulisuodatin; 6 - yläleuka etuhampailla; 7 - sivusuulakkeen uloskasvu; 8 - nenän väliseinä; 9 - suuontelo; 10 - nenäontelo; 11 - kieli; 12 - silmä
Useat suuontelon pohjan rakenteet osallistuvat kielen limakalvon muodostumiseen. Kielen runko muodostuu parillisten sivuttaispaksuuksien fuusiosta - lateraaliset kielen tuberkuloosit ja niiden välissä sijaitsee pieni keskikorkeus, ns tuberculum impar(Kuva 7.31). Uskotaan, että kielen runko on johdannainen ensimmäisen viskeraalisen kaaren alueen solumateriaalista. Osallistuu kielen juuren muodostukseen corula(Sulkumerkki)- keskikorkeus, sijaitsee takana tuberculum impar ja kytke päälle
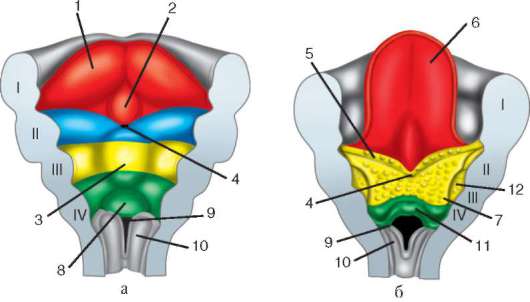
Riisi. 7.31. Kielen kehitys: a - 5 viikon alkio; b - 5 kuukauden ikäinen sikiö; 1 - lateraalinen kielen tuberkuloosi; 2- tuberculum impar 3 - kopula (Sulkumerkki); 4 - foramen coecum; 5 - päätevako; 6 - kielen runko; 7 - kielen juuri; 8 - kurkunruston asettaminen; 9 - äänisana; 10 - arytenoidituberkkeli; 11 - kurkunpää, palatininen risa. I-IV - sisäelinten kaarien lukumäärä
toisen, kolmannen ja neljännen viskeraalikaaren alueiden solumateriaali. Kaikkien kielten fuusion ja kielen lopullisen muodostumisen jälkeen sen runko erotetaan juuresta terminaalivao(sulcus terminalis), jonka keskellä on sokea reikä(foramen coecum)- nielun pohjan invaginaatiojälki, josta kilpirauhanen ilmestyy (katso kuva 7.27). Sitä pidetään kehon ja kielen juuren välisenä rajana.
Myöhemmin kieli erottuu melkein kokonaan suuontelon pohjasta, ja niiden välinen yhteys säilyy limakalvon taitoksen - kielen frenumin - muodossa.
Mielenkiintoinen esimerkki organogeneesistä on muodostuminen hampaat, johon osallistuu integumentaarinen ektoderma - stomodeumin epiteeli (emali) ja hermoharjasta peräisin olevat mesenkymaaliset solut (dentiini, sementti ja pulppu) (kuva 7.32). Kehityksen aikana havaitaan induktiovuorovaikutuksia mesenkymaalisten ja epiteelirakenteiden välillä.
Elämän aikana kehittyy kaksi sukupolvea hampaita. Hampaiden vaihtuminen tapahtuu lapsuudessa, ensimmäisiä hampaita kutsutaan meijeri. Niitä on 20: 10 yläleuassa ja 10 alaleuassa. Maitohampaat puhkeavat täysin noin 2 vuoden iässä. Tämä hammassarja palvelee lasta seuraavat 4 vuotta, jonka jälkeen maitohampaat vähitellen
Riisi. 7.32. Ihmisen pysyvät hampaat: 1 - etuhampaat; 2 - hampaat; 3 - pienet poskihampaat; 4 - suuret poskihampaat
mutta putoaa ja vaihdetaan pysyvät hampaat(toisen sukupolven hampaat), jotka toimivat ihmisessä loppuelämän ajan. Hampaiden vaihtoaika kestää noin 6-12 vuotta. 32 pysyvää hammasta: 16 ylempää ja 16 alempaa (suuret poskihampaat lisätään) (katso kuva 7.32). Muodoltaan ne ovat samanlaisia kuin meijeri, mutta ne ovat kooltaan suurempia.
Kirjanmerkki ja koulutus hammasbakteerit maitohampaat alkavat 6-7 viikon kohdunsisäisen kehityksen jälkeen. Suuontelon epiteeli on upotettu alla olevaan mesenkyymiin tiheän säikeen muodossa, ns. hammaslevy(Kuva 7.33). Hammaslevylle ilmestyy pieniä epiteelin ulkonemia - hammassilmut, josta maitohampaat kehittyvät. Kun hammaslamina kasvaa, jokaisen hampaan alkion koko kasvaa, tunkeutuu syvemmälle mesenkyymiin ja ottaa käänteisen kupin muodon. Tämä rakenne muodostuu emali urut, ja alla oleva mesenkyymi, joka täyttää kulhon ontelon, muodostuu hammaspapilla. Jatkokehityksessä jälkimmäinen muodostaa hampaan massan. Kohdunsisäisen kehityksen kolmannella kuukaudella emalielin kasvaa, muuttaa muotoaan ja irtoaa vähitellen hammaslevystä. Tulevan hampaan muodon määrää mesenkymaalinen komponentti. Yhdistettynä in vitro poskihampaan asettamisen mesenkyymi (poskihammas) etuhampaan asettamisen epiteelikomponentin kanssa, poskihammas kehittyy. Sitä vastoin poskihampaan ektodermin ja etuhampaan mesenkyymin yhdistelmän seurauksena muodostuu etuhammas.
Hammaskudosten histogeneesin aikana, joka alkaa 4. kuukauden lopussa, dentiini, emali ja massa(Kuva 7.33, d). Emali
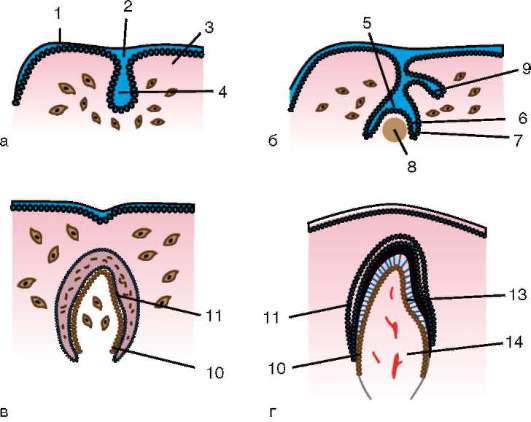
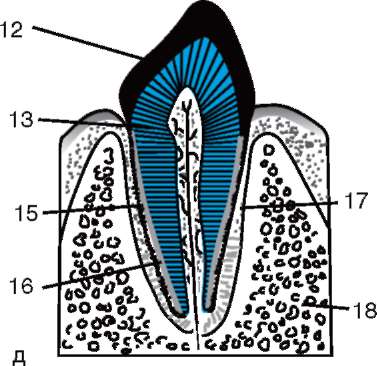
Riisi. 7.33.Hampaiden kehitys: a - 8 viikon alkio; b - 10 viikon ikäinen sikiö; c - 3 kuukauden ikäinen sikiö; g - 6 kuukauden ikäinen sikiö; d - purkauksen jälkeen; 1 - suuontelon epiteeli; 2 - hammaslevy; 3 - mesenkyymi; 4 - maitohampaan alkio; 5 - emali-elin; 6 - emalielimen sisäkerros; 7 - emalielimen ulkokerros; 8 - hammaspapilla; 9 - pysyvän hampaan alkio; 10 - odontoblastit; 11 - ameloblastit; 12 - emali; 13 - dentiini; 14 - hampaan massa; 15 - sementoblastit; 16 - sementti; 17 - parodontaali; 18 - alveolaarinen luu
tuottaa kiilleelimen sisäkerroksen soluja - emali räjäytys. Niiden prosessit erittävät emalin orgaanista perustaa - emaliprismoja, jotka sitten kalkkiutuvat. Kehityksen lähde dentiini- odontoblastit (dentinoblastit), pinnalliset massasolut. Dentinoblastien huipulla on myös prosesseja, jotka erittävät fibrillaarisen rakenteen orgaanisia aineita, ns predentiini ja muodostaa dentiinimatriisin. Viidennen kuukauden lopusta lähtien kalsium- ja fosforisuolat kerrostuvat predentiiniin, ja lopullinen dentiini muodostuu. Dentiinin ja kiilteen muodostuminen eroaa luun kehittymisestä siten, että solut eivät ole tarttuneet sisään solujen välinen aine, mutta siirry pois: ameloblastit - ulospäin, odontoblastit - sisäänpäin. Lisäksi orgaanisten aineiden pitoisuus näissä kudoksissa vaihtelee: luu sisältää noin 45% orgaanista ainetta, dentiini - 28% ja kiille - alle 5%.
Dentiinin ulkopinnalla hammasta ympäröivästä mesenkyymistä (hammaspussi) ilmestyvät vähän ennen hampaiden syntymistä sementoblastit. Nämä solut erittävät kollageenikuituja ja solujen välistä ainetta muodostaen sementti. Se muodostuu vain 4-5 kuukautta syntymän jälkeen hampaiden juurien kehittyessä. Hammaspussin ulkokerroksesta muodostuu hammas. nivelside (parodontium)(Kuva 7.33, e). Siten synnytystä edeltävässä kehityksessä kehittyvät vain maitohampaiden kruunut.
Hampaan puhkeaminen johtuu kolmesta tekijästä. Ensimmäinen ja tärkein on juuren kasvu ja kehitys, mikä johtaa kruunun karkottamiseen limakalvon läpi. Toinen tekijä on massan kasvu, mikä johtaa paineen nousuun hampaan itiön sisällä. Lisäksi hampaiden keuhkorakkuloiden pohjalle muodostuu lisäkertymiä. luukudos kerroksia.
Pysyvien hampaiden teko alkaa hyvin varhain, kohdunsisäisen kehityksen viidennen kuukauden alussa. Pysyvän hampaan itu sijaitsee maitohampaan itun takana, ja jälkimmäisen syntymän jälkeisenä kahden ensimmäisen vuoden aikana kehittyvä juuri vaikuttaa pysyvän hampaan alkion kehittymiseen induktiivisesti. Histologiset prosessit, jotka johtavat maidon ja pysyvien hampaiden muodostumiseen, ovat samat. Kun maitohampaat puhkeavat, pysyvien hampaiden alkuaineisiin muodostuu emali ja dentiini. Jonkin aikaa ne pysyvät piilevässä tilassa ja aktivoituvat vasta, kun leuka saavuttaa riittävän suuren koon. Korvausprosessissa pysyvän hampaan kasvu ja sen kiilteen paine maidon juureen johtavat resorptioon.
pehmeämmän kudoksen osteoklastit (solut, jotka tuhoavat luukudosta) - maitohampaan dentiini. Jälkimmäinen työnnetään ulos ja korvataan pysyvällä.
Osaston kehittäminen ruoansulatus- järjestelmiä on vaikea tarkastella yksityiskohtaisesti, koska suolistoputki on suuresti pitkänomainen, erilaistunut ja kehityksessään yhteydessä moniin muihin järjestelmiin: endokriiniset, hengitys-, eritys-, verenkierto-, hermosto- ja toissijaiset ontelot - coelom. Tarkastellaan vain varhaisimpia ja yleisimpiä suolistoputkessa tapahtuvia prosesseja sekä muutoksia sen yhteydessään keltuaispussiin.
Suolistoputki syntyy keltuaispussin katon endodermista ja viereisestä mesodermin viskeraalisesta levystä, kun alkion runko erotetaan alkion ulkopuolisista osista pään, hännän ja sivuttaisrungon taitteiden avulla (katso kuva 7.24, b 2, b 3). Neljännellä viikolla sitä edustaa suhteellisen yksinkertainen putki, joka koostuu etusuolasta, stomodaalisesta (suun) levystä, joka on sokeasti suljettu edestä, keskisuolesta, joka liittyy keltuaispussiin
keltuaisen varren (kanavan) ja takasuolen läpi, joka on sokeasti suljettu hännän päästä kloaakin (peräaukon) kalvolla
(Kuva 7.34).
Alkion (kohdunsisäisen) kehityksen 4. ja 8. viikon välisenä aikana suolisto pitenee ja alkaa erilaistua. Sen etuosasta muodostuu nielu, ruokatorvi, mahalaukku ja pohjukaissuoli sekä sen johdannaiset (maksa ja haima), ennen kuin yhteinen sappitie virtaa siihen.
Keskisuoli muodostuu U-muotoinen sekalainen silmukka, joka liitetään keltuaispussiin keltuaiskanavan kautta (kuva 7.35, a). Keltuaisen varren ja mahalaukun välinen osa suolesta muuttuu ohutsuoleksi ja osa
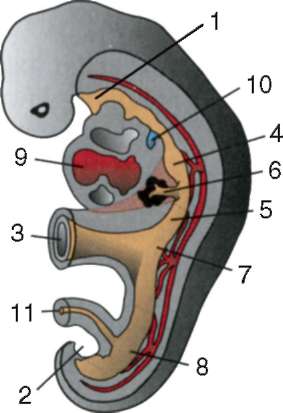
Riisi. 7.34.4 viikon ikäisen alkion ruoansulatusjärjestelmä: 1 - stomo-deal levy; 2 - kloakaan kalvo; 3 - keltuainen kanava; 4 - vatsa; 5 - pohjukaissuoli; 6 - maksan alkuaine; 7 - keskisuoli; 8 - takasuoli; 9 - sydän; 10 - hengityselinten asettaminen; 11 - Urachus
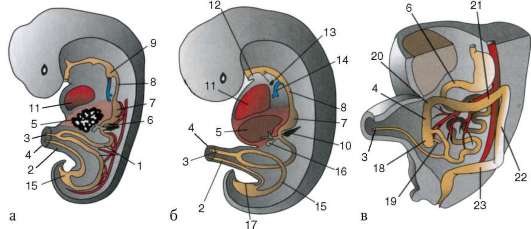
Riisi. 7.35.Ihmisen ruoansulatusjärjestelmän kehitys: a - 32. päivänä; b - 36. päivänä; c - myöhemmät vaiheet; 1 - suolen U-muotoinen silmukka; 2 - urakas; 3 - keltuainen kanava; 4 - vatsan varsi (napanuora); 5 - maksa; 6 - pohjukaissuoli; 7 - vatsa; 8 - ruokatorvi; 9 - kurkunpää; 10 - haima; 11 - sydän; 12 - kieli; 13 - kilpirauhasen kirjanmerkki; 14 - henkitorven ja keuhkojen alku; 15 - takasuoli; 16 - sappirakko; 17 - kloakaan kalvo; 18 - umpisuole; 19 - liite; 20-23 - paksusuolen osat
puristamalla kaudaalista vitelliinitiehyen, - paksusuoleen. Keskisuoli pidentyy, osa siitä työntyy vatsan varren onteloon muodostaen alkion vatsan seinämän herniaalisen ulkoneman.
Samaan aikaan jotkin suolen osat, pyörivät, muuttavat sijaintiaan (kuva 7.35, b). Viikolle 10 mennessä suolen silmukan ulkoneva osa vedetään takaisin navan reunan läpi ja ottaa lopullisen asemansa vatsaontelossa. Hieman aikaisemmin ohutsuolen siirtymäkohdassa paksusuoleen kehittyy umpisuole. Aluksi sen koko kasvaa, mutta kolmantena kuukautena sen distaalinen osa alkaa jäädä muiden osien kasvun jälkeen (epätasainen kasvu), minkä seurauksena tämän osan halkaisija on paljon pienempi ja umpilisäke on muodostettu. Takasuolesta muodostuu pernan kulman alapuolelle paksusuoli, mukaan lukien peräsuole (kuva 7.35, c). Takasuolen kloakaan pään kehitys liittyy läheisesti urogenitaalisen aukon kehittymiseen (kuva 7.36). Virtsaperäsuolen väliseinä, joka erottaa kloakan urogenitaaliseen poskionteloon ja peräsuoleen, muodostuu viikolla 5-6. Peräaukon kalvon läpimurto tapahtuu 8. viikolla.
Kohdunsisäisen kehityksen toisena kuukautena ruokatorven ja pohjukaissuolen epiteelin nopea proliferaatio alkaa, mikä
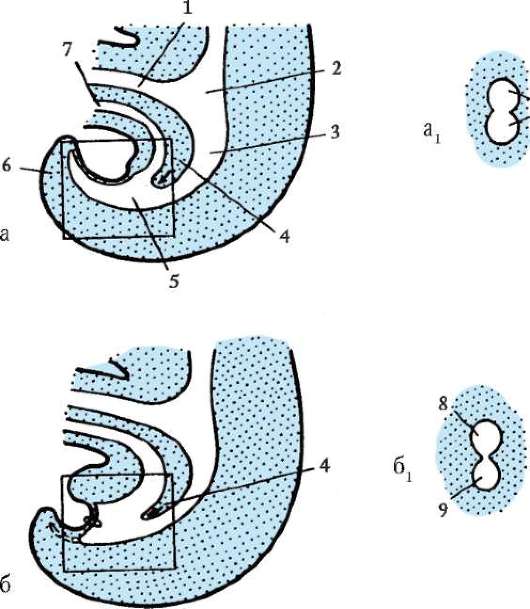
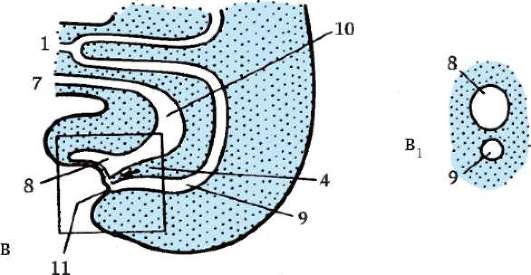
Riisi. 7.36. Ihmisalkion takasuolen kloakaan alueen kehitys: a - 4. viikolla; b - 6. viikolla; c - 7. viikolla; a 1 b 1 ja c 1 - poikittaisleikkaukset kloaakin läpi (leikkaustasot on merkitty neliöillä kuvioissa a; b; c). 1 - keltuaisen varsi; 2 - keskisuoli; 3 - takasuoli; 4 - urogenitaalinen väliseinä; 5 - kloaka; 6 - häntä; 7 - allantois; 8 - urogenitaalinen sinus; 9 - peräsuolen; 10 - virtsarakon; 11 - peräaukon kalvo; nuolet osoittavat kasvun suunnan
johtaa niiden luumenin tilapäiseen sulkeutumiseen. Toisen kuukauden loppuun mennessä tapahtuu näiden elinten rekanalisaatio, ts. läpäisevyyden palauttaminen.
Sydän henkilö alkaa kehittyä 3. viikon lopusta - 4. viikon alusta splanchnomesodermista nielun alla sijaitsevien parillisten alkuruuhojen muodossa (kuva 7.37). Koska itse alkion vartaloa rajaavat runkolaskokset ja etusuola on suljettu vatsan puolelta, parilliset endokardiaaliset putket sulkeutuvat yhdeksi, makaamalla keskiviivalla (ks. kuva 7.37). Sammakkoeläimillä on todistettu endodermin induktiivisen vaikutuksen olemassaolo sydänalassa olevaan mesodermiin. Tästä on osoituksena myös kanan alkiolla tehty koe, kun etusuolen pohjan poistamisen jälkeen saatiin alkio, jossa oli kaksoissydän. Celoomisten onteloiden parilliset anlages yhdistyvät vatsan puolella muodostaen sydänpussin ontelon.
Sydämen alueellinen erilaistuminen alkaa primaarisen sydänputken nopealla venymisellä, mikä johtaa sen taipumiseen ja 5-muotoisen muodon saamiseen (kuva 7.38). On huomattava, että mutkan muodostuminen, jossa solujen muoto muuttuu paikallisesti sydänputkessa, stimuloidaan itse sydänputkesta. Eksplantoituminen (tilanne "kehon ulkopuolella" tai ex vivo), sydänputki säilyttää kyvyn muodostaa 5-muotoisen rakenteen.
Riisi. 7.37. Ihmisen sydämen kehitys 21.-22. päivänä: a - alkion yleiskuva; b - alkion pääkartion pituusleikkaus; c - alkion poikkileikkaus: 1 - leikkaustaso; 2 - hermostunut rulla; 3 - somitit; 4 - ensisijainen nauha; 5 - aivot; 6 - etusuola; 7 - sydänputki; 8 - sydänpussi; 9 - amnion; 10 - orofaryngeaalinen kalvo; 11 - hermoura; 12 - sointu; 13 - selkä-aortan parilliset kirjanmerkit; 14 - sydänputkien yhdistäminen; 15 - keltuainen pussi
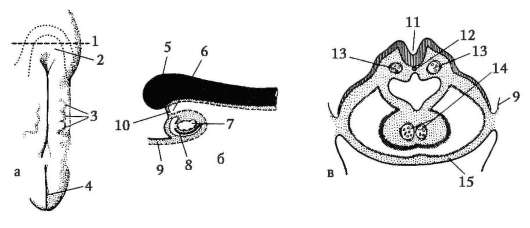
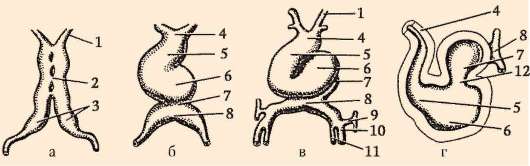
Riisi. 7.38. Kehittyvä ihmisen sydän: a - 21.-22. päivänä; b - 23. päivänä; c - 24. päivänä (näkymä vatsan puolelta); d - 28. päivänä (pitkittäisleikkaus): 1 - ensimmäinen pari aortan kaaria; 2 - sydänputkien yhdistäminen; 3 - ei-sulautuvat sydänputket; 4 - valtimorunko; 5 - sydämen polttimo; 6 - kammio; 7 - atrium; 8 - laskimoontelo; 9 - Cuvier-kanava; 10 - napalaskimo; 11 - keltuainen; 12 - eteiskammiokanava
Alunperin häntäpäässä on laskimoontelo, johon virtaavat suuret suonet: Cuvier-kanavat, napa- ja vitelline. Laskimoontelosta tulee myöhemmin osa oikeaa eteistä. Kalloonteloon nähden eteis muodostuu sydänputken laajennetusta osasta ja kammio kaarevasta keskiosasta. Siirtymäaluetta, jossa kammio kapenee, kutsutaan kartioksi tai sipuliksi. Se liitetään myöhemmin oikean kammion seinämään. Kartio siirtyy valtimorunkoon, josta vatsa-aortan juuret lähtevät.
Atriumin ja kammion muodostumisen jälkeen ilmenee ulkoisia merkkejä sydämen tulevasta jakautumisesta oikeaan ja vasempaan puoliskoon, ilmestyy mediaanisulcus. Tällä hetkellä kehittyvän sydämen kudoksissa havaitaan fokaalinen solukuolema ja tekijöiden solunulkoisten ja solujen elementtien vuorovaikutus. Jonkin ajan kuluttua kammion sisäpinnalle, joka vastaa ulomman uran tasoa, muodostuu väliseinä sydämen kärjestä eteiseen päin kasvavista lihasköydistä. Kapenevan atrioventrikulaarisen kanavan selkä- ja vatsa-seinämille endokardiaaliset tyynyt muodostuvat löysästä mesenkyymistä. Myöhemmin ne muuttuvat tiheiksi sidekudos, kasvavat yhdessä ja jakavat kanavan oikeaan ja vasempaan kanavaan. Samaan aikaan ilmestyy primaarinen mediaani väliseinä, joka myöhemmin korvataan toissijaisella. Siinä on reikä, jota kutsutaan soikeaksi, jonka läpi
veri oikeasta eteisestä virtaa vasempaan. Se on välttämätön sikiön verenkierrolle.
Samanaikaisesti sydämen pääosassa itsenäisenä elimenä tapahtuvien muutosten kanssa valtimorunko jakautuu kahteen kanavaan. Tämä prosessi alkaa vatsa-aortan juuresta neljännen ja kuudennen viskeraalisen kaaren välissä. Erottaminen tapahtuu pitkittäisten taitteiden muodostumisen vuoksi. Poimut ovat järjestetty spiraaliksi, kasvavat valtimorungon sisällä ja jakavat sen kohdatessaan aortaan ja keuhkorunkoon. Lisäksi taitokset leviävät kohti kartiota, jossa aortan ja keuhkon rungon puolikuuläppäimet muodostuvat erikoisalueilta, ja edelleen kammioihin, joissa ne kohtaavat eteiskammiokanavan liikkuvan endokardiaalikudoksen ja eteisväliseinän. Tämä johtaa kammioiden välisen aukon täydelliseen liikakasvuun.
Kohdunsisäisen elämän loppuun asti vain eteisväliseinän foramen ovale ja valtimotiehy, joka yhdistää keuhkon rungon aortaan, pysyvät auki. Siitä keskustellaan tarkemmin alla. Valtimotiehyen tehtävänä on ohjata osa verestä oikeasta kammiosta aortaan ohittaen keuhkot, kunnes ne ovat saavuttaneet tarvittavan kehityksen eivätkä ole saaneet toiminnallista ärsykettä hengitystoiminnan alkaessa (kuva 7.39).
Suuret valtimot kehittyä yhdessä sydämen kanssa 4. viikosta alkaen. Alkion primaariverenkierto on toiminut 5. viikon lopusta lähtien, 8. viikon loppuun mennessä rakenteen perussuunnitelma toteutuu valtimojärjestelmä. Neljännellä viikolla muodostuneet parilliset sisäelimet saavat asianmukaisen valtimovirran aorttakaarien muodossa. Nämä aorttakaaret nousevat valtimon rungosta tai pikemminkin sen laajentuneesta osasta - valtimopussista (kutsutaan myös vatsa- tai nousevaksi aortaksi) - ja päättyvät dorsaalisen eli laskevan aortan juuriin (katso kuva 7.27).
Huolimatta siitä, että asetetaan kuusi paria aortan (viskeraalisia) kaaria, ne eivät ole olemassa samanaikaisesti. Sinä aikana, jolloin kuudes kaaripari muodostuu, kaksi ensimmäistä paria ovat jo rappeutumassa. Viides kaari näkyy vain päällä lyhyt aika alkeellisen aluksen muodossa.
Pääsuonten muodostumisessa vatsa- ja selkäaortan juurilla sekä kolmannella, neljännellä ja kuudennella aorttakaaren parilla on olennainen rooli (kuva 7.40).
Kolmannen parin proksimaaliset osat muodostavat yhteiset kaulavaltimot (kuva 7.41). Vasen neljäs kaari on osa aortan kaaria.
Riisi. 7.39. Sikiön verenkiertokaavio: 1 - soikea ikkuna; 2 - interatrial väliseinä; 3 - keuhkolaskimo; 4 - yläonttolaskimo; 5 - valtimon (botall) kanava; 6 - keuhkovaltimo; 7 - laskeva aortta; 8 - navan valtimot; 9 - napalaskimo; 10 - alempi onttolaskimo; yksitoista - ductus venosus; 12 - maksan porttilaskimo. I - maksa; II - alempi onttolaskimo; III - oikea eteinen; IV - vasen eteinen; V - laskeva aortta
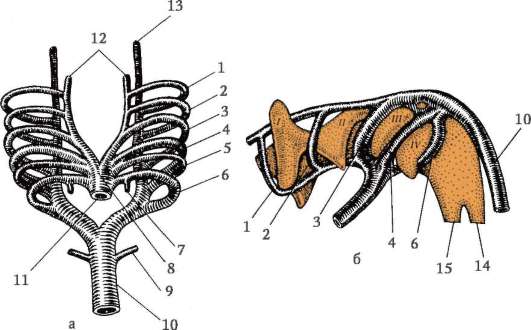
Riisi. 7.40. Aorttakaarien kehitys: a - näkymä vatsan puolelta: 1-6 - aorttakaaret; 7 - keuhkovaltimo; 8 - valtimopussi, vatsa-aortta; 9 - seitsemäs intersegmentaalinen valtimo, subclavian; 10 - dorsaalinen laskeva aortta; 11 - valtimorunko; 12 - ulkoinen kaulavaltimo, vatsan nousevan aortan juuri; 13 - sisäinen kaulavaltimo, dorsaalinen aortan juuri
Oikeasta neljännestä kaaresta tulee oikean subclavian valtimon proksimaalinen osa. Tämän valtimon distaalinen osa syntyy oikeasta dorsaalisesta aortasta. Vasemman kuudennen kaaren proksimaalinen osa muuttuu vasemman keuhkovaltimon proksimaaliseksi osaksi ja distaalinen osa valtimon (botall) kanavaksi. Oikean kuudennen kaaren proksimaalinen osa muodostaa oikean keuhkovaltimon proksimaalisen osan ja distaalinen osa pienenee. Selkä-aortan juuret sulautuvat hyvin varhain parittamattomaan selkä-aorttaan, mutta 7. kehitysviikolla oikean dorsaalisen aorttajuuren distaalinen osa liittyy yleensä oikeaan subclavian valtimoon, kuten edellä mainittiin. Tämä tulee mahdolliseksi, koska sivuston käänteinen kehitys yhdistää oikean juuren vasemmalle. Sydämen ja suurten valtimoiden kehitykseen liittyvän aineiston esittäminen antaa pohjaa vertailulle
Riisi. 7.41. Suurimpien valtimoiden muodostuminen, valtimorungon, aorttapussin, aorttakaarien ja selkä-aortan johdannaiset: a - 6 viikon sikiö; b - 7 viikon ikäinen sikiö; c - 6 kuukauden ikäinen sikiö: 1-6 - aorttakaaret; 7 - valtimorunko; 8 - keuhkovaltimo; 9 - subclavian valtimo; 10 - dorsaalinen laskeva aortta; 11 - aorttapussi; 12 - sisäinen kaulavaltimo; 13 - valtimon (botall) kanava; 14 - nouseva aortta; 15 - keuhkojen runko; 1b - ulkoinen kaulavaltimo; 17 - yhteinen kaulavaltimo
ihmisen ontogeneesin selkärankaisten fylogeneettisen kehityksen kanssa (katso luku 14), ja sen avulla voimme myös päästä lähemmäksi sydämen ja verisuonten synnynnäisten epämuodostumien esiintymisen mekanismeja.
Kysymyksiä itsehillintää varten
1. Millaisia ontogeneesityyppejä on olemassa? Listaa ja kuvaile ontogenian päävaiheet.
2. Mitkä ovat sukusolujen rakenteen ja toiminnan ominaisuudet?
3. Miten lannoitus suoritetaan ja mitä vaiheita tämä prosessi sisältää?
4. Kuvaile eri luokkien edustajien alkionkehityksen vaiheiden olemusta ja morfologisia muotoja
sondit (murskaus, gastrulaatio, organogeneesi ja ennaltaehkäisevien elinten muodostuminen).
5. Mitkä ovat nisäkkäiden alkionkehityksen piirteet?
6. Anna esimerkkejä ihmiselinten alkionkehityksestä, jotka kuvastavat sointujen esi-isien ryhmien kehitystä.
Ontogeneesi on organismin yksilöllinen kehitys, sarja peräkkäisiä morfologisia, fysiologisia ja biokemiallisia muutoksia, jotka organismi käy läpi hedelmöityksestä (sukupuolisen lisääntymisen aikana) tai siitä hetkestä, kun se eroaa äidin yksilöstä (aikana). suvuton lisääntyminen) elämän loppuun asti.
Jakakaa alkiota edeltävät, alkion jälkeiset ja jälkeiset ajanjaksot. Istukan eläimillä ja ihmisillä erotetaan synnytystä edeltävä (antenataalinen) ja synnytyksen jälkeinen (postnataalinen) ajanjakso.
Kaikki tapahtumat, jotka tapahtuvat ovo(oo)geneesissä, mutta uuden organismin yksilöllisen kehitysprosessin nimissä, muodostaa syntyvaiheen sisällön - esisikiö yksilöllisen kehityksen aika. Se alkaa ennen hedelmöittymishetkeä ja tsygootin muodostumista ja liittyy pääasiassa naisen sukusoluun. Jälkimmäinen saa gametogeneesin (ovogeneesin) aikana joitain ominaisuuksia, joita ei käytä itse, vaan yksilökehityksen aloittaneen uuden sukupolven yksilö. Yksi näistä ominaisuuksista, ehkä tunnetuin, on se, että munan sytoplasmaan muodostuu eläintyypistä riippuen suurempi tai pienempi määrä keltuaista, jota käytetään ravintoaineena jälkeläisten kehityksessä. . Useiden geenien toiminnallinen geneettinen aktiivisuus, joka ilmenee niiden transkriptiossa ja transkription jälkeisissä muutoksissa primaarisessa ja (m)RNA-transkriptissa, liittyy myös ajallisesti hedelmöittymistä edeltävään ajanjaksoon. Tämän toiminnan tuloksena muodostuva i(m)RNA järjestää alkion alkuvaiheille tärkeiden proteiinien synteesiä.
Munatyypit.
Ne luokitellaan keltuaisen määrän ja jakautumisen mukaan. Voi olla:
1. Alesitaaliset munat - ei keltuaista tai vain vähän
Esimerkki: nisäkkäillä
2. Isolesitaali - keltuaista on vähän, se jakautuu tasaisesti sytoplasmaan
Esimerkki: lansetti
3. Polylesitaali: a) telolesitaali - keltuainen siirtyy kohti vegetatiivista napaa
b) sentrolesitaali - keltuainen keskellä.
Lannoitus
Siittiöiden fuusio munan kanssa. Virstanpylväät hedelmöitysprosessi: 1) siittiöiden tunkeutuminen munasoluun; 2) erilaisten synteettisten prosessien aktivointi munassa; 3) munasolun ja siittiöiden ytimien fuusio palauttamalla diploidinen kromosomisarja.
Elinikä ja kyky hedelmöittää munasoluja ja siittiöitä ovat rajalliset. Useimmilla nisäkkäillä munasolu säilyttää hedelmöityskyvyn 24 tuntia ja ihmisillä 12–24 tuntia ovulaation jälkeen. Siittiöt säilyttävät hedelmöityskykynsä naisen sukupuolielimissä myös useita tunteja.
Siittiön ja munasolun suora kosketus aiheuttaa molemmissa soluissa säännöllisiä muutoksia, jotka aiheuttavat urospuolisen sukusolun ytimen ja sentriolin tunkeutumisen munasoluun ja sen aineenvaihdunnan aktivoitumisen. Näitä muutoksia siittiön puolelta kutsutaan akrosomaaliseksi reaktioksi ja munasolun puolelta kortikaalireaktioksi. Akrosomaalisen reaktion olemus on seuraava. Siittiöpään yläosassa olevan munan kanssa kosketuksissa plasmakalvo ja sen vieressä oleva akrosomaalisen rakkulan kalvon osa liukenevat. Entsyymien vapautumisen vuoksi akrosomaalisesta rakeesta viereinen munakalvon osa liukenee. Akrosomaalinen kalvo työntyy ulospäin ja muodostaa uloskasvun onton putken muodossa. Jälkimmäinen pitenee, kulkee munakalvojen läpi ja joutuu kosketuksiin munan plasmakalvon kanssa. Tällaisen kosketuksen alueella tapahtuu hedelmöittymisen ulkonema tai tuberkuloosi, jonka jälkeen molempien sukusolujen plasmakalvot sulautuvat ja niiden sisällön yhdistyminen alkaa. Tästä hetkestä lähtien siittiö ja muna ovat yksi solu - tsygootti. Akrosomaalinen reaktio on erittäin nopea.
Munasolujen aktivaatio tai aivokuoren reaktio, joka kehittyy joutuessaan kosketuksiin siittiöiden kanssa. Muutokset munasolun pinnallisessa kortikaalikerroksessa ja hedelmöityskalvon muodostuminen. Tämä kalvo, jota kutsutaan myös keltuaiskalvoksi, syntyy kuoriutumalla pois munan pinnasta. Sen alle muodostuu keltuainen tila, johon kaadetaan munan sytoplasman kortikaalikerroksen rakeiden sisältö. Hedelmöityskalvo suojaa munasolua ylimääräisten siittiöiden, ts. tarjoaa "polyspermian lohkon". Keltuaisen perimätilaan kerääntyvä neste toimii erityisenä ympäristönä, jossa alkion kehitys etenee siihen hetkeen asti, kun se poistuu munakalvoista.
SISÄÄN alkio ajanjaksolla on kolme päävaihetta: murskaus, gastrulaatio ja primaarinen organogeneesi. Alkion eli itämisvaiheen ontogeneesi alkaa hedelmöittymishetkestä ja jatkuu, kunnes alkio tulee ulos munakalvoista. Useimmissa selkärankaisissa se sisältää tsygootin, pilkkoutumisen, gastrulaation, histo- ja organogeneesin vaiheet.
Tsygootti on yksisoluinen alkio. Tsygootissa tapahtuu sytoplasman erilaistumisprosessi. Ooplasman liikettä havaitaan. Siihen ilmestyy alueita, joista myöhemmin muodostuu oletetut (oletetut) alkeet tulevista elimistä.
Pilkkominen - sarja hedelmöittyneen tai aloitetun munasolun peräkkäisiä mitoottisia jakautumisia kehitystä varten. Katkaisu on ensimmäinen alkion kehityksen jakso, joka on läsnä kaikkien monisoluisten eläinten ontogeniassa ja johtaa alkion muodostumiseen nimeltä blastula (yksikerroksinen alkio). Samanaikaisesti alkion massa ja tilavuus eivät muutu, eli ne pysyvät samoina kuin tsygootin, ja muna jaetaan yhä pienempiin soluihin - blastomeereihin. Jokaisen pilkkoutumisjakauman jälkeen alkion solut pienenevät ja pienentyvät, eli ydin-plasmasuhteet muuttuvat: ydin pysyy samana ja sytoplasman tilavuus pienenee. Prosessi jatkuu, kunnes nämä indikaattorit saavuttavat somaattisille soluille tyypilliset arvot. Murskaustapa riippuu keltuaisen määrästä ja sen sijainnista munassa. Jos keltuaista on vähän ja se jakautuu tasaisesti sytoplasmaan (isolesitaaliset munat: piikkinahkaiset, litamadot, nisäkkäät), murskaus etenee täydellisen univormutyypin mukaan: blastomeerit ovat samankokoisia, koko muna murskataan. Jos keltuainen jakautuu epätasaisesti (telolesitaaliset munat: sammakkoeläimet), murskaus etenee täydellisen epätasaisuuden tyypin mukaan: blastomeerit ovat erikokoisia, keltuaisen sisältävät ovat suurempia, muna murskataan kokonaisena. Epätäydellisen murskaamisen yhteydessä munissa on niin paljon keltuaista, että murskausvaot eivät voi erottaa sitä kokonaan. Munan pilkkoutumista, jossa vain eläinnapaan keskittynyt sytoplasman "korkki", jossa tsygootin ydin sijaitsee, murskataan, kutsutaan epätäydellisiksi kiekkomaiseksi (telolesitaaliset munat: matelijat, linnut). Epätäydellisen pinnan murskaantuessa keltuaisen syvyyksissä tapahtuu ensimmäiset synkroniset ydinjakaumat, joihin ei liity solujen välisten rajojen muodostumista. Tumat, joita ympäröi pieni määrä sytoplasmaa, ovat jakautuneet tasaisesti keltuaisessa. Kun niitä on tarpeeksi, ne siirtyvät sytoplasmaan, jossa solujen välisten rajojen muodostumisen jälkeen ilmaantuu blastodermi (keskelitiaaliset munat: hyönteiset).
· Gastrulaatio - solujen liikkumisprosessi, johon liittyy kasvua, lisääntymistä, erilaistumista. Tässä vaiheessa alkiota kutsutaan gastrulaksi.
Aluksi muodostetaan ulompi (ektodermi) ja sisempi (endodermi) levyt. Myöhemmin ilmestyy kolmas, keskimmäinen itukerros (mesodermi), joka sijaitsee alkion rungossa ektodermin ja endodermin välissä.
Ulko- ja sisälevyjen muodostamiseen on 4 päätapaa. Usein gastrulaatiossa on kuitenkin useiden eri tapojen yhdistelmä.
1 tapa invaginaatio, joka piilee siinä, että tietty osa blastodermista, samalla kun se säilyttää kerroksen rakenteen, ruuvataan blastocoeliin. Sitten blastokoel katoaa ja gastrocoel ilmestyy. Aukkoa, jonka kautta tämä onkalo kommunikoi ulkoisen ympäristön kanssa, kutsutaan ensisijaiseksi suuksi tai blastoporeksi.
2-suuntainen epibolia– makromeerien likaantuminen nopeasti jakautuvilla eläinnapaan mikromeereillä. Tällaisten eläinten alkioissa ei aluksi muodostu blastohuokosta eikä gastrocoelia ole.
3-suuntainen maahanmuutto– blastulan seinämän solujen osan laskeminen blastokoelin sisällä.
4-suuntainen delaminointi - ulkopuolella olevat solut muuttuvat kerrokseksi
a-ektoderma; b-endodermi; v-blastocoel.
Kolmannen itukerroksen asettaminen tapahtuu kahdella tavalla: teloblastinen ja enterokoelous.
Teloblastinen - menetelmä mesodermien muodostamiseksi protostomissa erottamalla kaksi mesodermaalista kaistaletta kahdesta primäärisestä alkiosolusta (teloblastista).
Enterocoelous - koostuu siitä, että ulkonemat muodostuvat primaarisen suolen endodermista molemmille puolille - taskut (coelomic pussit), jotka myöhemmin nauhoittuvat ja kasvavat ekto- ja endodermin väliin, muodostaen mesodermin; niiden ontelot sulautuvat toisiinsa ja syntyy toissijainen ruumiinontelo eli coelom.
Histo- ja organogeneesi
Kaksi vaihetta voidaan erottaa.
1 neurulaatio, koostuu aksiaalisten elinten kompleksin muodostumisesta - hermoputki, sointu. Neurulaation vaiheessa olevaa alkiota kutsutaan neurulaksi. Ensin tapahtuu solukerroksen litistyminen, mikä johtaa hermolevyn muodostumiseen, jonka reunat nousevat muodostavat hermolaskokset. Solujen liikkumisen vuoksi hermolevyn keskiviivaa pitkin tapahtuu painauma. Tässä vaiheessa neurulaation, kirjanmerkki hermosto kutsutaan hermouraksi. Samanaikaisesti hermolevy näyttää taittuvan keskiviivaa pitkin ja jonkin verran myöhemmin sen reunat sulkeutuvat. Näiden prosessien seurauksena syntyy hermoputki, jossa on ontelo - neurocoel. harjanteiden sulkeutuminen tapahtuu ensin hermouran keskellä ja sitten takaosassa. Tämä tapahtuu viimeisenä pääosassa, joka on muita alueita leveämpi. Etuosan laajennettu osa muodostaa jatkokehityksessä aivot, loput hermoputkesta - selkäytimestä.
2 histo- ja organogeneesi.
Ektodermista muodostuu ihon epidermis ja sen johdannaiset, rauhaset, suuontelon epiteeli, emättimen epiteeli, hammaskiille ja reseptorihermosolut.
Endodermista - muodostuu hengitysteiden ja keuhkojen epiteeli, osa haiman soluista, erittävät solut, mahalaukun epiteeli, suoli.
Mesodermista - rusto ja luuranko, lihakset, munuaiset, verisuonet.